

Introduction
Postpartum depression (PPD) is the most common psychiatric disorder following childbirth, with a global prevalence rate of approximately 17.22% (Z. Wang et al., Reference Wang, Liu, Shuai, Cai, Fu, Liu and Yang2021). Women with PPD typically experience insomnia, anxiety, irritability, fatigue, and persistent low mood, causing significant distress to the mothers and profoundly impacting entire families (Stewart & Vigod, Reference Stewart and Vigod2019). Some women with PPD may even develop suicidal and infanticidal ideation (Wisner et al., Reference Wisner, Sit, McShea, Rizzo, Zoretich, Hughes and Hanusa2013). Concurrently, PPD can indirectly affect infants’ emotional and intellectual development, potentially increasing their risk of depression during adolescence (Aoyagi & Tsuchiya, Reference Aoyagi and Tsuchiya2019). Although several functional neuroimaging studies have revealed differences in specific brain regions and connectivity patterns among PPD patients, the neurobiological underpinnings of PPD remain incompletely understood through neuroimaging research.
Over the past decade, blood oxygen level-dependent (BOLD) functional magnetic resonance imaging (fMRI) has developed rapidly, emerging as a promising tool for studying different brain activity regions in patients with postpartum depression (PPD), significantly advancing our understanding of the neurobiological mechanisms underlying this disorder. A task-based fMRI study of mothers’ neural response to their own infant’s emotion faces reported that patients with postpartum depression exhibit blunted dorsal anterior cingulate cortex responses to distress faces of their own infants (Laurent & Ablow, Reference Laurent and Ablow2013). Another task-based fMRI study revealed that patients with postpartum depression exhibited enhanced hippocampal activation in response to negative emotional music stimuli, which was negatively correlated with their subjective ratings of pleasure (X. Wang, Wang, & Liu, Reference Wang, Wang and Liu2014). However, due to variations in experimental designs and task paradigms, the findings from task-based fMRI studies can be highly heterogeneous (Bennett & Miller, Reference Bennett and Miller2010). Resting-state fMRI (rs-fMRI) provides a noninvasive and task-free approach, mitigating some confounding factors associated with variations in task performance, while offering a reliable method for measuring baseline brain activity (Biswal, Yetkin, Haughton, & Hyde, Reference Biswal, Yetkin, Haughton and Hyde1995). Resting-state functional imaging methods encompass various imaging modalities, such as amplitude of low-frequency fluctuation (ALFF)/fractional ALFF (fALFF), positron emission tomography (PET), single-photon emission computed tomography (SPECT), voxel-mirrored homotopic connectivity (VMHC), degree centrality (DC), arterial spin labeling (ASL), and regional homogeneity (ReHo). In SPECT and PET, radioactive tracers are utilized to measure cerebral metabolism. Neuronal activity cannot be directly represented by the BOLD signal. In fact, BOLD contrast relies on changes in cerebral blood flow (CBF), cerebral oxygen metabolism rate, and cerebral blood volume. The integrated combination of these coupled changes constitutes the hemodynamic activation response (Buxton, Uludağ, Dubowitz, & Liu, Reference Buxton, Uludağ, Dubowitz and Liu2004; Liu, Zhuo, & Yu, Reference Liu, Zhuo and Yu2016). Arterial spin labeling (ASL) employs magnetically labeled arterial blood water as an endogenous tracer to quantify CBF. ASL can measure both the BOLD signal and CBF concurrently (Buxton et al., Reference Buxton, Uludağ, Dubowitz and Liu2004). ALFF/fALFF quantified the strength of low-frequency fluctuations in spontaneous neural activity. VMHC assesses the synchronization of spontaneous neural activity between two symmetrical regions across cerebral hemispheres. DC reveals the importance of brain regions within the functional brain network connectivity. ReHo reflects the homogeneity of spontaneous neural activity among adjacent brain regions. Previous resting-state functional imaging studies have identified differences in spontaneous brain activity among patients with PPD. However, these results are inconsistent. On one hand, different studies have reported different brain regions, such as different functional activities in the precentral gyrus (Che et al., Reference Che, Mao, Li, Liu, Ma, Bai and Xie2020), precuneus (Che et al., Reference Che, Mao, Li, Liu, Ma, Bai and Xie2020), anterior cingulate cortex (Xu et al., Reference Xu, Yu, Lv, Zhou, Huang, Xu and Zhao2023), and temporal gyrus (Xiao-Juan et al., Reference Xiao-Juan, Jian, Zhi-Hong, Yan and Shi-Wei2011). On the other hand, the same brain region shows different patterns of activity across different studies. For example, Li et al. (Reference Li, Zhang, Li, Liu and Hou2022) reported decreased activities in the dorsolateral prefrontal cortex in PPD, while Che et al. (Reference Che, Mao, Li, Liu, Ma, Bai and Xie2020) reported hypermetabolism in the dorsolateral prefrontal cortex in PPD. Che et al. (Reference Che, Mao, Li, Liu, Ma, Bai and Xie2020) found lower activities in the inferior occipital gyrus in PPD. Xu et al. (Reference Xu, Yu, Lv, Zhou, Huang, Xu and Zhao2023) reported increased activities in the inferior occipital gyrus in PPD. These discrepancies may be closely related to patients’ onset time, depression severity, and analytical methods. A meta-analysis integrating multimodal functional neuroimaging techniques to investigate spontaneous neural activity in PPD during resting-state remains unreported.
The brain function differences in depression could be driven by serotonergic, dopaminergic, and other neurotransmitter signaling, and (or) are caused by genetic factors (Paul et al., Reference Paul, Östman, Heilig, Mayberg and Hamilton2023). Whole-brain gene expression data, such as the integration of the Allen Human Brain Atlas (AHBA) (Hawrylycz et al., Reference Hawrylycz, Lein, Guillozet-Bongaarts, Shen, Ng, Miller and Jones2012) with brain imaging data, link microscopic molecular pathways to macroscopic brain organization, providing a viable pathway for identifying the genetic mechanisms of various neuroimaging phenotypes (Fornito, Arnatkevičiūtė, & Fulcher, Reference Fornito, Arnatkevičiūtė and Fulcher2019). The integration of AHBA with brain imaging data has given rise to the emerging field of neuroimaging transcriptomics, which refers to the analysis of spatial changes in gene expression associated with spatial changes in a given neuroimaging phenotype (J. Chen et al., Reference Chen, Zhang, Wang, Jiang, Cai, Zhao and Yu2022). A recent study using transcription–neuroimaging association analysis found a correlation between brain dysfunction and specific genes in psychiatric disorders (G.-J. Ji et al., Reference Ji, Sun, Hua, Zhang, Zhang, Bai and Wang2023). However, the neurotransmitters and gene expression profiles associated with brain disruptions in PPD have not been reported.
Therefore, we aim to conduct a whole-brain voxel-wise meta-analysis to identify the most robust findings from published resting-state functional neuroimaging studies. Furthermore, we investigated the spatial correlation between the brain alterations identified in the meta-analysis and the neurotransmitter systems/gene expression. We hypothesize that increased and/or decreased spontaneous functional activity in PPD predominantly localizes to the occipital lobe, precuneus, and frontal lobe, which would spatially correlated with specific neurotransmitters and transcriptional profiles. The findings of this meta-analysis will facilitate further original research and contribute to a deeper understanding of PPD’s pathophysiological mechanisms.
Methods
Data sources, study selection, and quality assessment
We adhere to the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) international guidelines (Vrabel, Reference Vrabel2009) (registration number: CRD420251083433). A comprehensive search of studies published was conducted between January 1990 and August 2025 in the Web of Science, PubMed, WanFang, and Chinese National Knowledge Infrastructure databases using the following keywords: (‘Postpartum Depression’ OR ‘Post Natal Depression’ OR ‘Post-Natal Depression’ OR ‘Post Partum Depression’ OR ‘Postnatal Depression’ OR ‘Post-Partum Depression’) AND (‘neuroimag*’ OR ‘functional magnetic resonance imaging’ OR ‘fMRI’ OR ‘cerebral blood flow’ OR ‘CBF’ OR ‘positron emission tomography’ OR ‘PET’ OR ‘single photon emission computed tomography’ OR ‘SPECT’ OR ‘arterial spin labeling’ OR ‘ASL’ OR ‘amplitude of low frequency fluctuation’ OR ‘ALFF’ OR ‘LFF’ OR ‘LFO’ OR ‘fractional ALFF’ OR ‘fALFF’ OR ‘regional homogeneity’ OR ‘ReHo’ OR ‘reho’ OR ‘voxel-mirrored homotopic connectivity’ OR ‘VMHC’ OR ‘degree centrality’ OR ‘DC’ OR ‘resting-state’ OR ‘rest’ OR ‘resting’). Additionally, we checked the reference lists of included studies and relevant review articles to identify other relevant studies.
Studies that met the following criteria were included in the meta-analysis. (1) Patients diagnosed with PPD according to the Diagnostic and Statistical Manual of Mental Disorders, Fourth/Fifth Edition (DSM-IV/V) criteria, who had not received antidepressants or other psychotropic medications postpartum, (2) between-group comparisons of resting-state functional imaging profiles were performed, (3) spatial normalization utilized three-dimensional coordinates (either Talairach or Montreal Neurological Institute [MNI] standards) for whole-brain resting-state functional imaging analyses, (4) significant results were reported using thresholds either corrected for multiple comparisons or uncorrected thresholds with cluster extent thresholds, and (5) the investigation was published as an original research article (categorized as neither abstract nor letter) in a peer-reviewed journal accepting submissions in either English or Chinese.
Studies were excluded if (1) past and present medical history of neurological or psychiatric disorders in patients and first-degree relatives, (2) using a region-of-interest-based approach, (3) these data overlap with data from another included publication, (4) data were unavailable even after contacting the authors via phone or email (e.g., missing neuroanatomical coordinates).
The 10-point checklist involved in the previous neuroimaging meta-analysis was used to assess the quality of each study selected in this meta-analysis (Supplementary Table S1) (Z. Q. Chen et al., Reference Chen, Du, Zhao, Huang, Li, Lui and Gong2015; Shepherd et al., Reference Shepherd, Matheson, Laurens, Carr and Green2012). The literature search, study evaluation, and selection were independently completed by two researchers (C.G.M. and T.X.Y.). Any discrepancies would be resolved by the third investigator (H.X.X), who would make the final decision. This study was conducted following the Meta-analysis Of Observational Studies in Epidemiology (MOOSE) guidelines (Stroup et al., Reference Stroup, Berlin, Morton, Olkin, Williamson, Rennie and Thacker2000).
The sample sizes of the PPD group and healthy controls (HCs) group from all studies finally included in the analysis were pooled into two separate groups, respectively, for an independent samples t test. Comparisons of age, years of education, and scores on the Edinburgh Postnatal Depression Scale (EPDS) between the two groups were performed using the same approach.
Data analysis
Analysis of function
Meta-analyses of functional differences (specifically ReHo, ALFF, fALFF, VMHC, and DC) between PPD and HCs groups were performed using the Seed-based d Mapping with Permutation of Subject Images (SDM-PSI software version 6.21) following its standard protocol (www.sdmproject.com). SDM-PSI is a voxel-wise meta-analysis tool that utilizes peak coordinates and their corresponding t values reported in primary studies to compute multiple effect size maps (Hedges’ g) for group comparisons (hyperactivation and hypoactivation) within each study. Subsequently, these maps were pooled into a conventional random-effects model, weighted by sample size, within-study variability, and between-study heterogeneity (Radua et al., Reference Radua, Mataix-Cols, Phillips, El-Hage, Kronhaus, Cardoner and Surguladze2012). SDM-PSI estimates the maximum possible effect size within the lower and upper bounds of possible effect sizes for each study separately using MetaNSUE, and then adds realistic noise (Albajes-Eizagirre, Solanes, & Radua, Reference Albajes-Eizagirre, Solanes and Radua2019). We conducted the analyses as described in the SDM tutorials and related publications, visualizing the SDM maps using MRIcron software (www.mricro.com/mricron/). In line with the recommendations of the SDM software developers, we reported an uncorrected p value threshold of <0.005, a minimum cluster extent of >10 voxels, and a mean SDM-Z >1 to reduce false positives (Radua et al., Reference Radua, Mataix-Cols, Phillips, El-Hage, Kronhaus, Cardoner and Surguladze2012). This parameter provided an optimal balance between false positives and negatives and yielded results comparable to those obtained with corrected thresholds (Müller et al., Reference Müller, Cieslik, Laird, Fox, Radua, Mataix-Cols and Eickhoff2018; Radua et al., Reference Radua, Mataix-Cols, Phillips, El-Hage, Kronhaus, Cardoner and Surguladze2012).
Jackknife sensitivity analysis
To assess the reliability and consistency of the key findings, we conducted a jackknife sensitivity analysis. One study is removed at a time, followed by the repetition of the analyses; subsequently, that study is reinstated, another study is removed, and the analytical process is repeated again, with this cycle continuing sequentially for all studies (Radua & Mataix-Cols, Reference Radua and Mataix-Cols2009). Brain regions that maintained statistical significance throughout most or all of these modified analyses were deemed to show stable and reproducible effects.
Heterogeneity and publication bias analysis
The peak coordinate values were extracted to obtain relevant data for evaluating heterogeneity across studies and potential publication bias. The I 2 statistic was applied to examine interstudy heterogeneity, thereby verifying the robustness of the study findings (I 2 > 50% commonly indicates serious heterogeneity) (Egger, Davey Smith, Schneider, & Minder, Reference Egger, Davey Smith, Schneider and Minder1997). To determine potential publication bias, funnel plots were created to visually examine, and Egger’s tests were conducted (Sterne et al., Reference Sterne, Sutton, Ioannidis, Terrin, Jones, Lau and Higgins2011), whether findings were driven by a small subset of studies or those with limited sample sizes. An asymmetric funnel plot and a p < 0.05 indicated the presence of significant publication bias.
Regression analysis
Additionally, for the thresholded meta-analysis results, we conducted metaregression analyses to assess the influence of age, sex, education years, and EPDS during data preprocessing – factors that may potentially confound the findings. At this stage, we continued using an uncorrected threshold of p < 0.005 with a cluster extent of 10 voxels.
Neurotransmitter analysis
By using JuSpace v1.5, we assessed spatial correlations between different brain functional patterns and distribution patterns of various receptor/transporter systems (https://github.com/juryxy/JuSpace) (computing option 3) (Dukart et al., Reference Dukart, Holiga, Rullmann, Lanzenberger, Hawkins, Mehta and Eickhoff2021). In the study, serotoninergic neurotransmitter targets – the 5-hydroxytryptamine receptor subtype 1a (5-HT1a) receptor, 5-HT1b receptor, 5-HT2a receptor, 5-HT4 receptor, and serotonin transporter (SERT) (Fiore et al., Reference Fiore, Preziosa, Tedone, Margoni, Vizzino, Mistri and Filippi2023); dopaminergic neurotransmitter targets – the D1 receptor, D2 receptor, dopamine transporter (DAT), and FluoroDOPA (FDOPA) (Premi et al., Reference Premi, Pengo, Mattioli, Cantoni, Dukart, Gasparotti and Borroni2023); as well as the vesicular acetylcholine transporter (VAChT) (Giboureau et al., Reference Giboureau, Som, Boucher-Arnold, Guilloteau and Kassiou2010) were both selected for spatial correlation. Exact permutation based p values (10,000 permutations; adjusted for spatial autocorrelation) were calculated for all analyses to test whether the observed mean correlation coefficient across subjects differed significantly from the null distribution (Dukart et al., Reference Dukart, Holiga, Rullmann, Lanzenberger, Hawkins, Mehta and Eickhoff2021). False discovery rate (FDR) correction was applied to all analyses across group comparisons (Dukart et al., Reference Dukart, Holiga, Rullmann, Lanzenberger, Hawkins, Mehta and Eickhoff2021).
Brain gene expression processing
The available microarray-based gene expression data of six postmortem brains with 3702 distinct samples were downloaded from the AHBA dataset (mean age: 42.5 years, 1 female) (Hawrylycz et al., Reference Hawrylycz, Lein, Guillozet-Bongaarts, Shen, Ng, Miller and Jones2012). Because only two of the six brains in the AHBA database had right hemispheres, only the left hemisphere was considered in this study. The abagen toolbox (v.0.1.3; https://github.com/rmarkello/abagen) was utilized to process and map the transcriptomic data onto the functional maps, including updating probe-to-gene annotations and selection, and normalization across tissue samples and donors (details were shown in Supplementary Materials). We performed the functional meta-analysis within the gray matter mask provided by SDM-PSI. Consequently, we restricted the functional analysis to samples within this mask, resulting in a final sample × gene matrix of 1714 × 10,028 (Fang et al., Reference Fang, Cai, Jiang, Zhao, Song, Zhao and Zhu2023). We then defined a 3-mm radius sphere centered on the MNI coordinates of that sample and extracted the mean z value of the voxels within the sphere from the PPD meta-analysis z map.
Spatial associations between transcriptome and functional maps
Partial least squares (PLS) regression analysis was employed to examine the association between gene transcriptomic patterns and the differences observed between groups in the functional maps (Xue et al., Reference Xue, Guo, Zhu, Liang, Xu, Ma and Liu2023). Each PLS component is obtained through a weighted linear combination of the gene expression values, ordered according to the amount of covariance explained between the predictor and response variable sets. The first PLS component (PLS-1) provides the optimal low-dimensional summary of the covariance structure within the high-dimensional data matrix (Abdi & Williams, Reference Abdi and Williams2013). Subsequently, genes reordered according to their adjusted regression coefficients represent their respective contributions to the PLS regression components. We utilized the BrainSMASH toolbox (Burt et al., Reference Burt, Helmer, Shinn, Anticevic and Murray2020) to generate 5000 surrogate maps preserving the spatial autocorrelation of brain functional variation. Finally, we computed the null distribution of gene expression correlations with these surrogate brain maps and compared it against our true observed result, thereby validating the association between the brain and transcriptomic maps.
Gene enrichment analysis
We employed the GOrilla tool (http://cbl-gorilla.cs.technion.ac.il/) to perform gene enrichment analysis for both upregulated and downregulated genes, identifying enriched Gene Ontology (GO) terms (Eden et al., Reference Eden, Navon, Steinfeld, Lipson and Yakhini2009). The evaluation encompassed the GO classifications of biological processes (BPs), molecular functions (MFs), and cellular components (CCs). Enrichment significance was determined using a FDR corrected threshold of q < 0.05.
Results
Included studies and sample characteristics
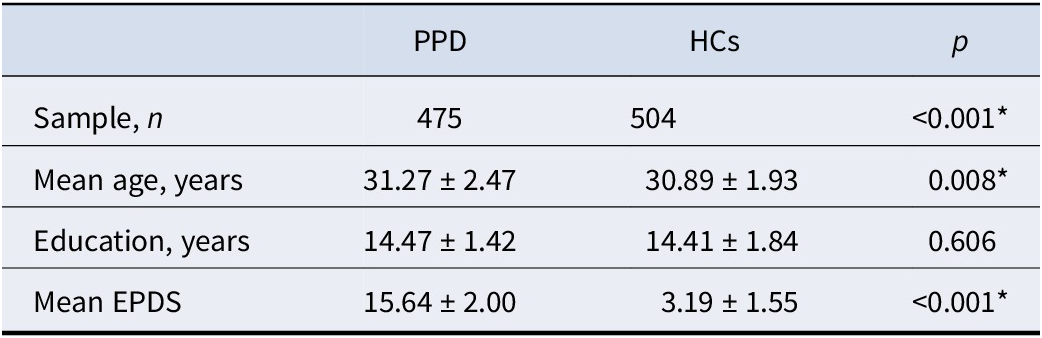
The flowchart illustrating study identification and exclusion is presented in Figure 1. Seventeen experiments from 12 studies comprising 475 patients with PPD and 504 HCs were selected (Table 1). The demographic, clinical, neuroimaging characteristics and quality assessment scores of studies included in this meta-analysis are detailed in Supplementary Table S2.
Flow diagram for the identification and exclusion of resting-state functional imaging studies of patients with PPD. Abbreviations: PPD, postpartum depression; ALFF, amplitude of low-frequency fluctuation; fALFF, fractional amplitude of low-frequency fluctuation; ReHo, regional homogeneity; VMHC, voxel-mirrored homotopic connectivity; DC, degree centrality; CBF, cerebral blood flow; ROI, region of interest.

Demographic information of meta-analysis samples

Note: Means ± standard deviations are reported. All p values obtained using independent sample t test. * indicates statistical significance. PPD, postpartum depression; HCs, healthy controls; EPDS, Edinburgh Postnatal Depression Scale.
PPD meta-analysis
Patients with PPD displayed increased resting-state functional activity in the left inferior occipital gyrus and left precuneus as well as decreased resting-state functional activity in the right amygdala and left precentral gyrus (p < 0.005, uncorrected) (Figure 2 and Table 2). No significant results survived when a more conservative statistical threshold (p < 0.05, familywise error [FWE]-corrected) was used.
Meta-analyses results regarding resting-state functional differences between PPD and HCs, uncorrected, p < 0.005. Areas with increased value are displayed in red, and areas with decreased value are displayed in blue. The color bar indicates the maximum and minimum SDM-Z values. PPD, postpartum depression; HCs, healthy controls; SDM, Seed-based d Mapping.

Meta-analyses results regarding resting-state functional brain activity difference between PPD and HCs

Note: PPD, postpartum depression; HCs, healthy controls; BA, Brodmann area; MNI, Montreal Neurological; CI, confidence interval.
Jackknife sensitivity analysis
In the analysis of PPD, jackknife sensitivity analysis revealed that these results were robust and replicable in most datasets (p < 0.005, uncorrected). Details were shown in Table 2. No statistically significant results were observed in the sensitivity analysis upon using a more rigorous cluster-extent threshold (p < 0.05, FWE-corrected).
Heterogeneity and publication bias analysis
In the PPD meta-analysis, none of the brain regions displaying functional alterations across the included studies showed significant heterogeneity (I 2 < 50%) or publication bias (p > 0.05) (Supplementary Figure S1 and Table 2).
Metaregression analysis
The metaregression analysis revealed no statistically significant results.
Spatial associations between neurotransmitter and brain changes
The patterns of resting-state functional activity differences in patients with PPD associated with 5HT1a (rho = −0.3265, p = 0.013), D1 (rho = −0.4444, p < 0.001), DAT (rho = −0.4857, p < 0.001), FDOPA (rho = −0.4242, p < 0.001), SERT (rho = −0.4769, p < 0.001), and VAChT (rho = −0.6001, p < 0.001) (Figure 3).
(a) Spatial correlations between brain alterations and neurotransmitter distribution maps in PPD compared to HCs. The radar map showing the absolute values of correlation coefficients (vertical axis). (b) The first partial least squares component (PLS-1) identified a profile of genes that were positively correlated with functional difference in PPD. (c) Enrichment analysis related to PPD functional changes (color coded by p value for significant enrichment). PPD, postpartum depression; *p < 0.05, **p < 0.01, ***p < 0.001.

Transcriptome profiles and gene enrichment
PLS-1 accounted for 21.37% of the variance in PPD functional divergence (p = 0.011, corrected for spatial autocorrelation with 5000 permutation tests), and demonstrated a statistically significant association with functional divergence in PPD (r = 0.33, p < 0.001) (Figure 3). The gene enrichment analysis revealed that the genes of PLS1 were enriched in BPs related to inorganic ion transmembrane transport. The enriched genes in MFs were related with ion-gated channel activity, substrate-specific channel activity, cation channel activity, and passive transmembrane transporter activity. The enriched genes in CCs were associated with ion channel complex, transmembrane transporter complex, and cation channel complex (Figure 3 and Supplementary Table S3).
Discussion
This is the first whole-brain meta-analysis of resting-state functional activity in patients with PPD. The main findings were as follows: (i) Patients with PPD showed decreased spontaneous functional activity in the right amygdala and left precentral gyrus, and increased functional activity in the left inferior occipital gyrus and left precuneus. (ii) The functional differences in PPD were spatially associated with serotonergic, dopaminergic, and VAChT transmitter system. (iii) The functional differences in PPD were associated with the genes involved in inorganic ion transmembrane transport, ion-gated channel activity, substrate-specific channel activity, cation channel activity, passive transmembrane transporter activity, ion channel complex, transmembrane transporter complex, and cation channel complex. These findings may provide novel insights into the pathophysiology of PPD.
We found decreased resting-state functional activity in the right amygdala in patients with PPD, indicating impaired intrinsic functional activities in the amygdala in patients with PPD. The amygdala, as a key component of the limbic system, is responsible for emotion recognition, memory integration, and the regulation of stress responses. Several resting-state fMRI studies reported reduced connectivity of amygdala (Chase et al., Reference Chase, Moses-Kolko, Zevallos, Wisner and Phillips2014; Deligiannidis et al., Reference Deligiannidis, Sikoglu, Shaffer, Frederick, Svenson, Kopoyan and Moore2013; Mao et al., Reference Mao, Che, Xie, Li, Wang, Liu and Zhuo2020). These research findings on the weakened functional connectivity of the amygdala raise a possibility that PPD may involve external preventive dysfunction in self-related thinking and psychotheoretical/empathetic processes. A resting-state fMRI study of repetitive transcranial magnetic stimulation showed that the intrinsic functional connectivity between the bilateral amygdala increased in PPD patients following treatment, and this increase was closely correlated with improvements on the postpartum depression scale (Zhang et al., Reference Zhang, Mu, Li, Sun, Ma, Li and Qi2022). PPD is a major depressive disorder (MDD) with postpartum onset and exhibits distinct patterns of neural activation compared to major depressive disorder. For example, previous research (O’ Brien, et al., Reference O’ Brien, Sethi, Gudbrandsen, Lennuyeux-Comnene, Murphy and Craig2021) has indicated that the postpartum period is accompanied by drastic fluctuations in sex hormone levels. The reduction in amygdala functional activity may reflect a failure of functional adaptation or abnormal inhibition under the drastically changing neuroendocrine environment. This also serves as one of the unique neuropathological mechanisms distinguishing postpartum depression from nonperipartum major depressive disorder. In addition, in response to noninfant emotional stimuli, PPD exhibits reduced amygdala activation, whereas MDD shows increased amygdala activation (Barrett et al., Reference Barrett, Wonch, Gonzalez, Ali, Steiner, Hall and Fleming2012; Moses-Kolko et al., Reference Moses-Kolko, Horner, Phillips, Hipwell and Swain2014; Wonch et al., Reference Wonch, de Medeiros, Barrett, Dudin, Cunningham, Hall and Fleming2016). Therefore, based on neuroimaging studies comparing MDD to PPD, we infer that the functional roles of these brain regions may be particularly relevant to maternity and might be specifically influenced by maternal emotions.
The precentral gyrus is the core region of the primary motor cortex, responsible for the planning and execution of movement. It also exhibits functional connectivity with the brain’s advanced networks, indirectly modulating emotion and cognition (Xu et al., Reference Xu, Wei, Bai, Wang, Li, He and Wang2020). In the study, we found decreased resting-state functional activity in the left precentral gyrus in patients with PPD. Unlike MDD outside the perinatal period, PPD patients may undergo unique remodeling within the sensorimotor network (including the precentral gyrus), driven by postpartum hormonal fluctuations (such as sharp declines in estrogen and oxytocin), and the stress of adapting to the maternal role (Tang et al., Reference Tang, Tang, Zhou, Luo, Wen, Yang and Luo2025). For example, studies (Cheng et al., Reference Cheng, Guo, Chen, Lv, Liao, Qu and Wang2022; Dong et al., Reference Dong, Zhang, Chu, Che, Li, Gai and Xie2022) have found that PPD patients exhibit reduced ALFF or functional connectivity strength in the precentral gyrus during the resting state, while MDD patients may show increased activity or no significant changes in this region. The precentral gyrus, through its connections with the prefrontal–limbic system (e.g., anterior cingulate cortex, insula), mediates motor responses within mother–infant interactions (such as coordinated movements like holding an infant). Reduced function in this region may contribute to deficits in maternal behavior (Tang et al., Reference Tang, Tang, Zhou, Luo, Wen, Yang and Luo2025). These findings imply that the precentral gyrus might be a key region in patients with PPD, which could result in clinical symptoms of PPD.
The inferior occipital gyrus is a key component of the visual cortex, responsible for primary visual information processing and visual processing with emotional relevance (Shang et al., Reference Shang, Zhu, Wu, Xu, Chen, Dou and Ye2023). The precuneus serves as a central hub of the Default Mode Network (DMN), integrating self-referential processing, episodic memory retrieval, and social cognition (Dadario & Sughrue, Reference Dadario and Sughrue2023). In our study, increased resting-state functional activity in the left inferior occipital gyrus and left precuneus were found in patients with PPD. During the postpartum period, mothers require heightened attention to their infant’s visual cues (such as facial expressions and movements). Enhanced activity in the inferior occipital gyrus may represent an adaptive response to the demands of mother–infant interaction (C. Chen et al., Reference Chen, Li, Zhang, Liu, Wang, Xu and Liu2023). Increased activity in the precuneus is associated with excessive rumination (e.g., thoughts like ‘I’m an incompetent mother’), exacerbating negative self-evaluation. The precuneus modulates the processing of infant emotional cues through its connections with the limbic system (Gholampour, Riem, & van den Heuvel, Reference Gholampour, Riem and van den Heuvel2020). Hyperactivity in this region may impair attention to positive infant signals while heightening sensitivity to negative cues, such as infant crying.
Furthermore, the functional differences in PPD were spatially associated with serotonergic, dopaminergic, and VAChT transmitter system. What is more, in terms of biological process, the gene were enriched for inorganic ion transmembrane transport; in terms of molecular functions, the gene were enriched for ion-gated channel activity, substrate-specific channel activity, cation channel activity, and passive transmembrane transporter activity; in terms of cellular components, the gene were enriched ion channel complex, transmembrane transporter complex, and cation channel complex. A previous animal study reported that the sharp postpartum decline in estrogen suppresses tryptophan hydroxylase activity, reducing central serotonin synthesis (Zhao et al., Reference Zhao, Zhou, Shi, Ye, Lyu, Wen and Xu2022). This serotonin deficiency results in weakened functional activity in amygdala-related brain regions, impairing the ability to suppress negative emotions and thereby contributing to the onset of postpartum depression (Haim et al., Reference Haim, Albin-Brooks, Sherer, Mills and Leuner2016; Zhao et al., Reference Zhao, Zhou, Shi, Ye, Lyu, Wen and Xu2022). At the same time, the postpartum decline in oxytocin secretion reduces the activity of dopaminergic neurons, leading to diminished function in the brain’s reward pathway. This ultimately results in psychomotor retardation and anhedonia (Medina, De Guzman, & Workman, Reference Medina, De Guzman and Workman2024). The VAChT is the key protein responsible for the storage and release of ACh within synaptic vesicles. It is primarily expressed in the presynaptic terminals of cholinergic neurons. VAChT regulates cognitive, attentional, and emotional networks by transporting cytosolic ACh into synaptic vesicles, thereby maintaining quantal release of the neurotransmitter (Kanel et al., Reference Kanel, Müller, van der Zee, Sanchez-Catasus, Koeppe, Frey and Bohnen2020; Kipp et al., Reference Kipp, Nunes, Galaj, Hitchcock, Nasra, Poynor and Savage2021). In PPD, dysfunction of the VAChT can disrupt the default mode-limbic system circuitry, exacerbating emotional dysregulation (Pádua-Reis et al., Reference Pádua-Reis, Aquino, Oliveira, Szawka, Prado, Prado and Pereira2017). Previous studies demonstrated that selective serotonin reuptake inhibitors (SSRIs) are commonly used to treat PPD. They likely mediate their antidepressant effects, at least in part, by modulating the activity of voltage-gated cation channels (Sohn et al., Reference Sohn, Brown, Sharma, Ziemann and McGirr2024; Tabka et al., Reference Tabka, Cheikh, Maatoug, Ayeb, Bendahhou and Benkhalifa2020). Furthermore, estrogen withdrawal affects the expression of serotonergic and dopaminergic neurotransmitter transmembrane transporters (Cliburn et al., Reference Cliburn, Dunn, Stout, Hoffman, Lohr, Bernstein and Miller2017). This leads to reduced vesicular neurotransmitter loading and insufficient presynaptic reserves (Cliburn et al., Reference Cliburn, Dunn, Stout, Hoffman, Lohr, Bernstein and Miller2017). In summary, physiological dysfunction in PPD exhibits spatial correlations with serotonergic, dopaminergic, and VAChT neurotransmitter systems, while related genes are enriched in biological processes such as transmembrane ion transport and channel activity. Postpartum hormonal fluctuations contribute to dysfunction in associated brain regions and disruption of emotional networks by affecting the synthesis, activity, and transport of these neurotransmitter systems (Ji et al., Reference Ji, Liao, Chen, Zhang and Wang2017, Reference Ji, Ren, Li, Sun, Liu, Gao and Wang2019, Reference Ji, Sun, Hua, Zhang, Zhang, Bai and Wang2023). Existing therapeutic approaches may partially mediate their antidepressant effects through the modulation of cation channel activity.
This study had several limitations. First, the analysis depended on peak coordinates and effect sizes reported in the original studies, not the original statistical maps. This approach results in the loss of the complete spatial continuity contained within those maps, thereby compromising the accuracy of the analytical results (Radua et al., Reference Radua, Mataix-Cols, Phillips, El-Hage, Kronhaus, Cardoner and Surguladze2012). Second, after rigorous screening against the inclusion criteria and meeting the minimum criteria recommended for SDM software analysis, a limited number of studies were ultimately included. The statistical power was insufficient to perform formal subgroup analyses, including imaging metric (e.g., ALFF/fALFF vs. ReHo vs. DC), medication status (medicated vs. unmedicated), postpartum onset timing (early vs. late), and number of postpartum depressive episodes (first vs. recurrent). Future studies with larger sample sizes are warranted to examine their effects on the findings. Third, the scope of this meta-analysis was narrowed to English and Chinese publications due to language proficiency and resource constraints, a limitation that may introduce selection bias. Future work should aim to incorporate studies in additional languages to mitigate this issue and enhance the review’s comprehensiveness and impartiality. In addition, using the JuSpace toolbox, spatial correlations were assessed between the provided neuroimaging maps and atlas-derived neurotransmitter system maps. The correlations observed likely represent overarching neuroanatomical patterns rather than specific neurochemical signatures. Hence, these results should be construed with caution. Moreover, this research did not investigate prognostic outcomes or utilize MR imaging features to study illness trajectories in individuals with a genetic predisposition to psychosis. These aspects represent valuable directions for future inquiry. Finally, for our findings, an uncorrected threshold of p < 0.005 was adopted. While previous studies have suggested that this threshold is recommended when the cluster extent is 10 voxels and the SDM-Z value is greater than 1, as it can effectively control the false positive rate (Radua et al., Reference Radua, Mataix-Cols, Phillips, El-Hage, Kronhaus, Cardoner and Surguladze2012), it is still an approximation of corrected results. Unfortunately, when we further applied a more conservative statistical threshold (p < 0.05, FWE-corrected), no significant results were retained in the meta-analysis. This could be due to the following reasons: (a) the limited number of cases in the included case–control studies; (b) heterogeneity not captured by our meta-analysis, such as the time of postpartum onset. Owing to the limited number of studies, we did not conduct subgroup analyses based on different postpartum onset times. These results should be interpreted with greater caution and require validation in independent datasets.
Conclusion
This study identified that spontaneous functional alterations in the precuneus, amygdala, and primary sensorimotor cortex in PPD are implicated in core clinical features. Specifically, precuneus hyperactivity within the DMN may underlie excessive rumination and negative self-referential thinking, while amygdala hypoactivity could contribute to emotional blunting and impaired mother–infant bonding. These findings suggest a dysregulated interplay between the default mode and salience networks in PPD. Furthermore, the spatial correspondence of these functional changes with serotonergic and dopaminergic systems provides neurobiological correlates for existing pharmacotherapies and highlights potential targets for future interventions, such as neuromodulation of the precuneus. Collectively, these insights advance the neurobiological understanding of PPD and inform the development of more objective assessment tools and targeted treatment strategies.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0033291726103651.
Data availability statement
The data will be made available upon reasonable request.
Acknowledgments
The study was supported by grants from the Guangdong Basic and Applied Basic Research Foundation, China (2023A1515110582); Special Funds for Social Public Welfare and Basic Research in Zhongshan City (Grant No. 2021B1026); National Natural Science Foundation of China (82472057, 81971597, and 82502506); Science and Technology Projects in Guangzhou (2024B03J1299). The funding organizations play no further role in study design, data collection, analysis and interpretation and paper writing.
Author contribution
Guanmao Chen, Xinyue Tang, and Pan Chen: conception and design, analysis and interpretation of data, drafted the article. Ruoyi Chen, Chao Chen, Yongxin Zhang, Zhilong Wang: revised the manuscript critically for important intellectual content. Fangyun Li, Jianfeng Fei, Danyu Lu, and Xiuyu Wang: acquisition of data. Xiaoxing Huang, Ying Wang, and Xuehong Xiao: conception and design, acquisition of data, analysis and interpretation of data, revised the manuscript critically for important intellectual content. All authors have carefully reviewed and revised the manuscript and approved the final version for submission.
Competing interests
The authors declare none.







 Open access
Open access