


Introduction
Seed germination is a critical stage in the plant life cycle, directly influencing population demographics, species persistence and vegetation dynamics (Fenner and Thompson, Reference Fenner and Thompson2005). This early life-history stage of plants is strongly influenced by environmental factors that act as cues to initiate or delay the process (Fenner and Thompson, Reference Fenner and Thompson2005). Seeds often do not germinate immediately after dispersal due to dormancy, an adaptive mechanism that delays germination until conditions are expected to be more favourable for seedling survival (Baskin and Baskin, Reference Baskin and Baskin2004, Reference Baskin and Baskin2014).
Seed dormancy can be classified according to the mechanisms involved (classification based on Baskin and Baskin, Reference Baskin and Baskin2004, Reference Baskin and Baskin2014). Physiological dormancy involves internal hormonal regulation and often requires specific environmental cues to be alleviated (e.g., cold stratification under snow for seeds produced in alpine environments). Physical dormancy, on the other hand, is the result of water-impermeable seed coats that require scarification (e.g. through fire or mechanical abrasion). Morphological dormancy occurs when seeds are dispersed with underdeveloped embryos that must grow and mature before they can germinate. In some cases, seeds exhibit complex dormancy related to multiple factors when two or more mechanisms are combined. The type of dormancy a species exhibits, or the lack of it, is mainly thought to be driven by climate conditions under which the species have evolved (Rosbakh et al., Reference Rosbakh, Carta, Fernández-Pascual, Phartyal, Dayrell, Mattana, Saatkamp, Vandelook, Baskin and Baskin2023).
Once dormancy has been broken, temperature and soil moisture become the primary drivers of germination. The extent to which temperature influences the speed and proportion of seed germinating in different species (i.e. their thermal germination niche) is often determined by the environmental conditions in which the species has evolved. High-elevation species tend to germinate within narrower thermal ranges and have higher optimal temperatures than lowland species, which generally exhibit broader thermal niches (Walder and Erschbamer, Reference Walder and Erschbamer2015). These differences in thermal germination responses are ecologically significant, affecting the timing of emergence, seedling survival and, ultimately, population dynamics (Fenner and Thompson, Reference Fenner and Thompson2005; Baskin and Baskin, Reference Baskin and Baskin2014).
In this context, climate change is expected to disrupt environmental requirements for dormancy and germination of many species, particularly in highly seasonal environments such as those at high elevations (Vázquez-Ramírez and Venn, Reference Vázquez-Ramírez and Venn2021; Mondoni et al., Reference Mondoni, Jiménez-Alfaro, Cavieres, Baskin and Baskin2022). For example, the projected reduction in snow cover depth and duration in alpine environments (Hock et al., Reference Hock, Rasul, Adler, Cáceres, Gruber, Hirabayashi, Jackson, Kääb, Kang, Kutuzov, Milner, Molau, Morin, Orlove, Steltzer, Kaser and Mukherji2019) could prevent the natural cold stratification that many alpine species require to break dormancy. As seed dormancy and germination timing have evolved in synchrony with prevailing climatic conditions, rapid changes to environmental conditions could result in phenological mismatches and reduced recruitment success (Walck et al., Reference Walck, Hidayati, Dixon, Thompson and Poschlod2011).
Despite their ecological significance, the dormancy and germination traits of high-elevation species remain poorly understood in many taxonomic groups and mountainous regions of the world, including the Australian Alps (Vázquez-Ramírez and Venn, Reference Vázquez-Ramírez and Venn2021; Mondoni et al., Reference Mondoni, Jiménez-Alfaro, Cavieres, Baskin and Baskin2022). In particular, few studies have directly contrasted these traits among closely related species across elevational gradients (e.g. Walder and Erschbamer, Reference Walder and Erschbamer2015). Understanding how dormancy depth and thermal germination niches vary among congeneric species distributed in contrasting environments may help predict species resilience to climate change.
Here, we investigated the presence of physiological dormancy and the thermal germination niche of different congeneric taxa from contrasting elevations. We considered gibberellic acid (GA3), a plant hormone known to stimulate germination (Kildisheva et al., Reference Kildisheva, Dixon, Silveira, Chapman, Sacco, Mondoni, Turner and Cross2020), as an experimental proxy to detect physiological dormancy. We then subjected both treated and untreated seeds to a temperature gradient. This approach allowed us to address the following research questions: (1) Do high-elevation species exhibit deeper physiological dormancy than those from lower elevations (i.e. a greater response to GA3)? and (2) Do high-elevation species germinate over a narrower and warmer range of temperatures than lowland seeds? We also examined the distribution of seed size within and between taxa to account for any potential effect of this trait on seed dormancy depth and thermal responses.
Material and methods
Study species
We selected 21 species/subspecies that represent four major groups representative and endemic to the Australian flora: Eucalyptus (Myrtaceae; five subspecies of Eucalyptus pauciflora subsp. pauciflora, subsp. acerina, subsp. hedraia, subsp. niphophila and subsp. parvifructa); Callistemon (Myrtaceae; five species; C. kenmorrisonii, C. forresterae, C. nyallingensis, C. pityoides and C. brachyandrus); Brachyscome (Asteraceae; nine species; B. barkerae, B. decipiens, B. dentata, B. foliosa, B. melanocarpa, B. muelleroides, B. nivalis, B. spathulata and B. tadgellii); Deyeuxia (Poaceae; two species; D. affinis and D. talariata) – each representing a particular life form – trees, shrubs, forbs and grasses respectively (Fig. 1). Of these species/subspecies, 14 are characteristic of high-elevation communities (i.e. alpine and subalpine environments; >1,500 m a.s.l.), while the remainder are found at lower elevations (<1,100 m a.s.l.). Of the study taxa, 14 are listed as threatened under the Victorian state-level Flora and Fauna Guarantee Act 1988 and 3 are listed as vulnerable under the Australian national-level Environment Protection and Biodiversity Conservation Act 1999 (see Table S1). Within the genus Brachyscome, seven species are represented by more than one population (see Table S1) to test for variation in germination responses at the population level and contrast this with the species level.
Collections of seed from sampled study species from different elevations. Coordinates and additional site information for each study species can be found in Table S1. Seed photographs were captured using an Olympus Stereo SZ16 microscope with a fixed scale for each genus.

Figure 1 Long description
The image displays seeds from four plant groups: Eucalyptus, Callistemon, Brachyscome and Deyeuxia. Each group is shown with seeds from different species or subspecies, along with their respective elevations. Eucalyptus pauciflora includes subspecies pauciflora at 47 meters, parvifructa at 1,178 meters, hedraia at 1,612 meters, acerina at 1,687 meters and niphophila at 1,786 meters. Callistemon species include kenmorrisonii at 150 meters, brachyandrus at 150 meters, forresterae at 150 meters, nyallingensis at 320 meters and pityoides at 1,100 meters. Brachyscome species include melanocarpa at 31 meters, dentata at 34 to 116 meters, muelleroides at 113 meters, barkerae at 1,600 meters, foliosa at 1,609 meters, tadgellii at 1,642 to 1,811 meters, decipiens at 1,665 to 1,774 meters, nivalis at 1,600 meters and spathulata at 1,800 meters. Deyeuxia species include affinis at 1,686 meters and talariata at 1,416 meters. The seeds are shown with a scale indicating size, either 1 millimeter or 500 micrometers, depending on the group.
Seed collection and trait measurements
Seeds were collected from wild populations across an elevational gradient ranging from 31 to 1,822 m a.s.l. in Victoria, Australia (Fig. 1). In addition, we included the progeny of one population (B. tadgellii) grown at a lower elevation to explore potential dormancy loss (see Table S1 for collections metadata). Following collection, seeds were processed according to standard seed banking protocols (Martyn Yenson et al., Reference Martyn Yenson, Sommerville, Guja, Merritt, Dalziell, Auld, Broadhurst, Coates, Commander, Crawford, Emery, Funnekotter, Knapp, Makinson, Monks, Wrigley and Offord2024) and stored at the Royal Botanic Gardens Victoria Conservation Seed Bank, where they remained until the start of the experiment (see Table S1). Seed storage conditions followed standards set out by the Millennium Seedbank Partnership and involved dehumidification of seeds at 15% relative humidity and 18℃ post collection and long-term storage at −20℃ (Sutcliffe and Adams, Reference Sutcliffe and Adams2014). Seed age post-collection ranged from 1 to 16 years (see Table S1). Seed storage duration can affect germination capacity, particularly among alpine plants whose seeds are often considered short-lived (e.g. Satyanti et al., Reference Satyanti, Nicotra, Merkling and Guja2018), and this is acknowledged as a limitation (see Section 4.4). However, accessions were stored under controlled conditions prior to use in this experiment, and the viability of ungerminated seeds was assessed at its conclusion, which together limit the influence of storage-related effects on our results (see Section 2.3).
In 2020, prior to the start of the experiment, seeds were retrieved from long-term storage and weighed using an analytical digital balance. Seed length and width were measured from digital images captured on an Olympus Stereo SZ16 microscope and quantified using Olympus cellSens imaging software (Olympus, cellSens 1.17 Build 21199).
Germination experiments
The trials were designed to assess physiological dormancy and the effect of temperature on germination. To test for seed dormancy, two treatments were applied: seeds were either soaked in deionized water for 8 hours (control) or in a GA3 solution (250,000 ppm) for 8 hours. All seeds were then sown onto sterile 90-mm Petri dishes containing a 1% agar medium. Each dish contained 15–100 seeds per species per replicate as seed availability differed across the study species (see Table S2). Each Petri dish was sealed individually with Parafilm to prevent moisture loss. There were four replicates for each treatment and temperature combination.
To test the effect of temperature on germination, both control and GA3-treated seeds were incubated at each of the following six constant temperatures: 5°C, 10°C, 15°C, 20°C, 25°C or 30°C, using Thermoline incubators which were set to a 12-hour light/dark photoperiod. Within each incubator, Petri dishes were randomly positioned on shelves and remained in place throughout the experiment. Constant temperatures were chosen instead of diurnal cycles to avoid confounding species-specific responses, as previous studies have demonstrated that taxonomic groups may respond differently to fluctuating temperatures (Liu et al., Reference Liu, Baskin, Baskin, Bu, Du and Ma2013). Germination was scored every 3–4 days based on radicle emergence (>1 mm). The experiment ran for 56 days across all taxa and incubator settings. At the conclusion of the experiment, all remaining ungerminated seeds were cut-tested to assess viability. An indicator of a viable seed was a firm, white endosperm and a fresh, intact and non-coloured embryo (Commander et al., Reference Commander, Quarmby, Cochrane, Ruiz-Talonia, Craigie, Crawford and Commander2021). Seeds deemed non-viable at the conclusion of the experiment were not considered in the analysis.
Data analysis
Due to almost no germination at our 5°C treatment (which is reflective of cold stratification conditions), we removed this treatment class from our statistical analysis; however, we included these data in plots.
We ran four Generalized Linear Mixed Models (GLMMs) using the ‘glmmTMB’ package in R (Brooks et al., Reference Brooks, Kristensen, van Benthem, Magnusson, Berg, Nielsen, Skaug, Maechler and Bolker2017; R Core Team, 2022) separated by life-form; trees, shrubs, forbs and grasses. Whether a seed germinated was used as a binary response variable with (GA3 application [binary]) seed treatment (control or GA3), species (or subspecies), temperature (five-level categorical variable: 10°C, 15°C, 20°C, 25°C and 30°C), the interaction between GA3 and species, and the interaction between treatment and temperature as explanatory variables. Seed plate was included as a random effect to account for seeds placed on the same plate potentially experiencing different microclimatic conditions during the experiment. Seed storage time could not be included in the analysis as it was confounded with species (i.e. each species has a unique storage time). Models were validated by comparing Pearson residuals in relation to fitted values and residuals in relation to each variable in the model. Heterogeneity patterns were observed across different treatment levels and we therefore included Treatment in ‘dispformula’ to account for this heteroscedasticity. We plotted proportions of seed germinated to aid with interpretation.
We assessed differences in seed size between populations as we acknowledge this can influence germination; however, we do not have replication of the same species across elevational gradients. We therefore include these results to aid interpretability of our findings, acknowledging the confounding nature of seed size and species. To assess how seed size varied between subspecies, species and populations, we ran a permutational multivariate analysis of variances (PERMANOVAs) of Euclidean distance resemblance matrices of length and width measurements of seeds. Separate models were run for each of the four groups. Significant terms in PERMANOVAs can reflect differences between assemblages, differences in group dispersions or a combination of both. Hence, where significant PERMANOVAs were found, we also ran a permutational test for differences in group dispersions to assist with interpretations. We subsequently ran canonical analyses of principle coordinates to assist in visualizing the effects. These analyses were run in Primer v7 software.
Results
Overall, GA3 increased germination success across most taxa, but high-elevation species exhibited significantly stronger responses, particularly among Eucalyptus and Deyeuxia taxa. In contrast, Callistemon species showed no response to GA3. Thermal responses also varied with seed origin, with alpine species and subspecies generally exhibiting higher germination at warmer temperatures (20–25 °C).
Seed size
Seed size differed between species of Eucalyptus (Pseudo-F (4, 495) = 61.873, p = 0.0001), Callistemon (Pseudo-F (4, 494) = 88.906, p = 0.0001), Brachyscome (Pseudo-F (13, 1486) = 482.2, p = 0.0001) and Deyeuxis (Pseudo-F (1,148) = 394.36, p = 0.0001). E. pauciflora pauciflora and E. p. hedraia had both longer and wider seeds when compared to E. p. niphophila, E. p. acerina and E. p. parvifructa (Figure S1). C. forresterae and C. kenmorrisonii had much longer seeds when compared to C. brachyandrus, C. nyallingensis and C. pityoides (Figure S2). C. pityoides also tended to have wider seeds (Figure S2). B. barkerae had the longest seeds, followed by B. foliosa when compared to all other species, with B. melanocarpa and B. spathulata having the shortest seeds (Figure S3). There was strong clustering of B. nivalis, B. tadgellii and B. dentata. B. decipiens also clustered together, although with slightly longer and wider seeds found in the Pretty Valley wet area compared to the Pretty Valley rocky area (Figure S3). D. talariata had longer and wider seeds when compared to D. affinis (Figure S4).
Trees – Eucalyptus species
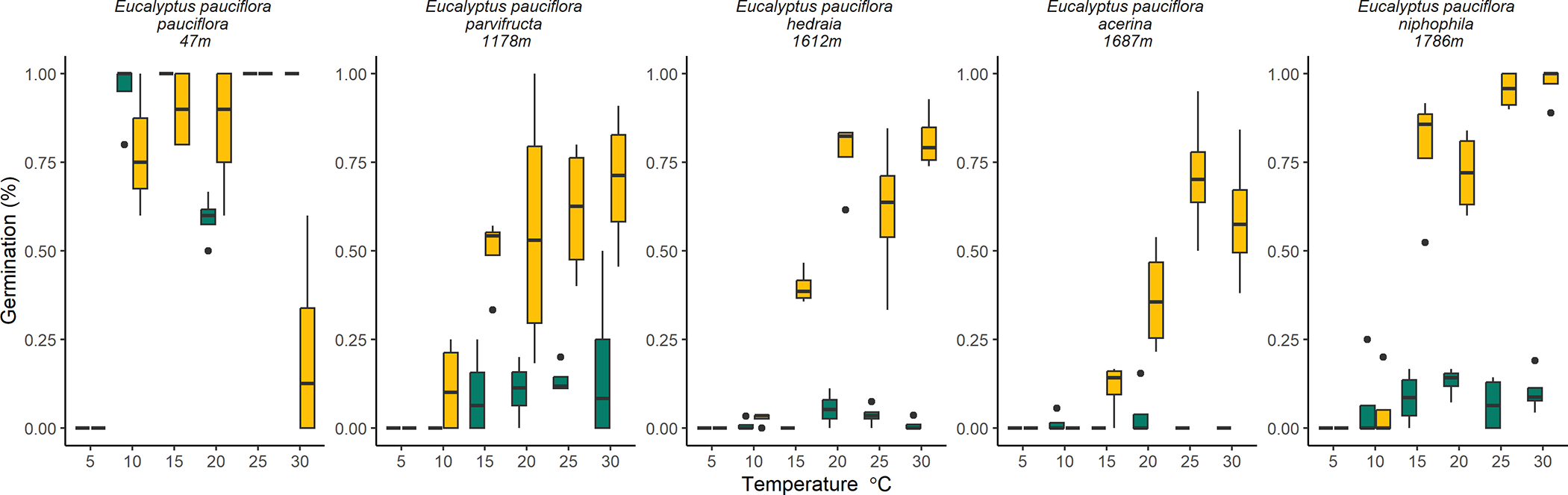
We found an interaction between seed treatment and subspecies (χ2 = 61.178, df = 4, p < 0.001) and between treatment and temperature (χ2 = 23.939, df = 4, p < 0.001). GA3 increased the germination percentage across all E. pauciflora subspecies (E. p. acerina: control = 0.6%, GA3 = 25%; E. p. hedraia: 1.5%, 47.6%; E. p. niphophila: 6.5%, 78.7%; E. p. parvifructa: 6.5%, 45.8%) except for the lowest elevation subspecies E. p. pauciflora where it decreased (control = 91.2%, GA3 = 78%; Fig. 2).
Percentage germination of seeds across five subspecies of Eucalyptus pauciflora tested at six temperatures. Note: Data for seeds treated with GA3 (yellow) and untreated controls (green) are presented.

Figure 2 Long description
Five box plots display the germination percentage of Eucalyptus subspecies seeds at different temperatures. Each plot represents a subspecies: Eucalyptus pauciflora at 470 meters, Eucalyptus pauciflora parvifructa at 1070 meters, Eucalyptus pauciflora hedraia at 1057 meters, Eucalyptus pauciflora acerina at 1657 meters and Eucalyptus pauciflora niphophila at 1750 meters. The x-axis is labeled 'Temperature' in degrees Celsius and the y-axis is labeled 'Germination' in percentage. Each plot compares germination rates of seeds treated with GA (yellow) and untreated controls (green) across temperatures ranging from 0 to 30 degrees Celsius. The plots show variations in germination rates, with treated seeds generally showing higher germination percentages compared to untreated seeds, except for Eucalyptus pauciflora at 470 meters where untreated seeds have higher germination rates.
Temperature had a quadratic effect on seed germination when seeds were treated with GA3 (ꞵ = −5.817, SE = 1.078, p < 0.001) compared to control seeds, which showed no clear differences (p > 0.05; Table S2). Germination under GA3 increased from 6% at 10°C to 53% at 15°C and peaked at 84% at 25°C before decreasing to 79% at 30°C (Table S3). Differences existed between all temperatures except between 15°Cand 20°C and >20°C (p > 0.05; Fig. 2).
Shrubs – Callistemon species
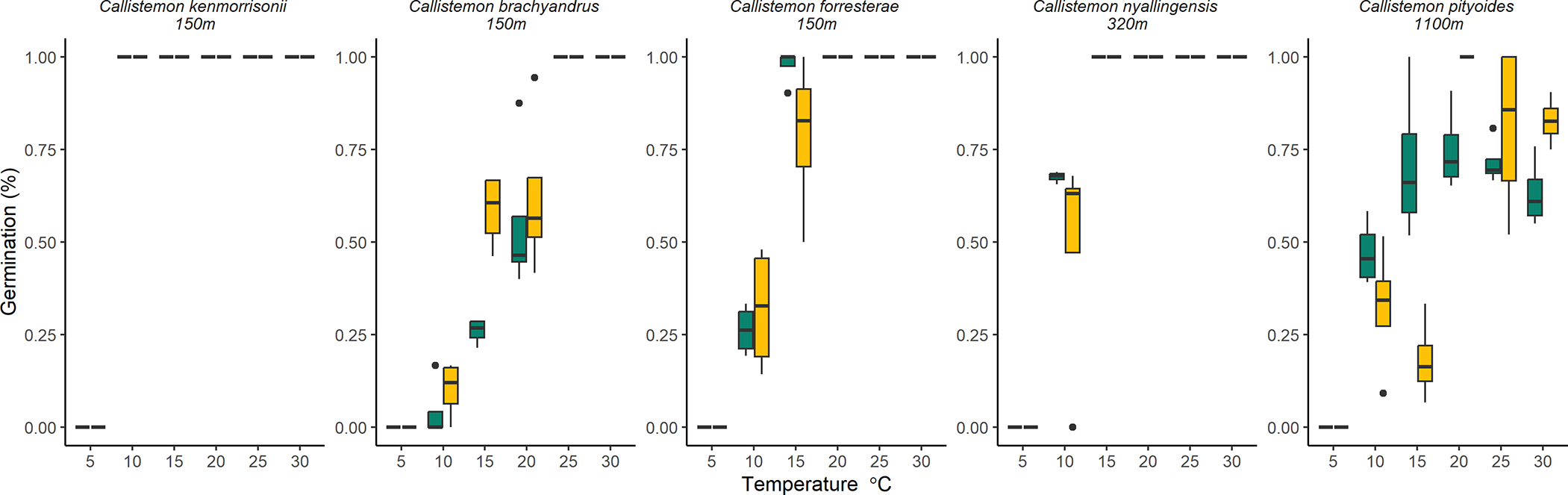
Germination rates differed between Callistemon species (χ2 = 86.374, df = 4, p < 0.001) and across different temperatures (χ2 = 125.354, df = 4, p < 0.001). There was no interaction between species and treatment (χ2 = 9.910, df = 4, p = 0.752) or between treatment and temperature (χ2 = 5.387, df = 4, p = 0.250). Seed treatment had no influence on germination rate (χ2 = 0.123, df = 1, p = 0.726). Germination rate for C. kenmorrisonii, C. nyallingensis and C. forresterae reached near 100% but was lower for C. pityoides and C. brachyandrus (75% and 76%, respectively; p < 0.05; Fig. 3; Table S4). A quadratic relationship existed between germination rate and temperature, germination rates were lowest at 10℃ and increased to a peak at 25℃ before reducing at 30℃ with clear differences between temperatures <25℃ and >25℃ in all comparisons (Fig. 3; Table S2).
Percentage germination of seeds from five Callistemon species tested at six temperatures. Note: Data for seeds treated with GA3 (yellow) and untreated controls (green) are presented.

Figure 3 Long description
Five box plots display the germination percentage of different Callistemon species at various temperatures. Each plot represents a species: Callistemon kenmorrisonii, Callistemon brachyandrus, Callistemon forresterae, Callistemon nyallingensis and Callistemon pityoides. The x-axis is labeled 'Temperature degrees Celsius' and the y-axis is labeled 'Germination percent'. Each plot compares treated seeds (yellow) and untreated seeds (green) across temperatures ranging from 10 to 30 degrees Celsius. The germination rates vary, with some species showing higher germination at specific temperatures. The plots include data points, whiskers and outliers, illustrating the distribution and variability of germination rates for each species under different conditions.
Grasses – Deyeuxia species
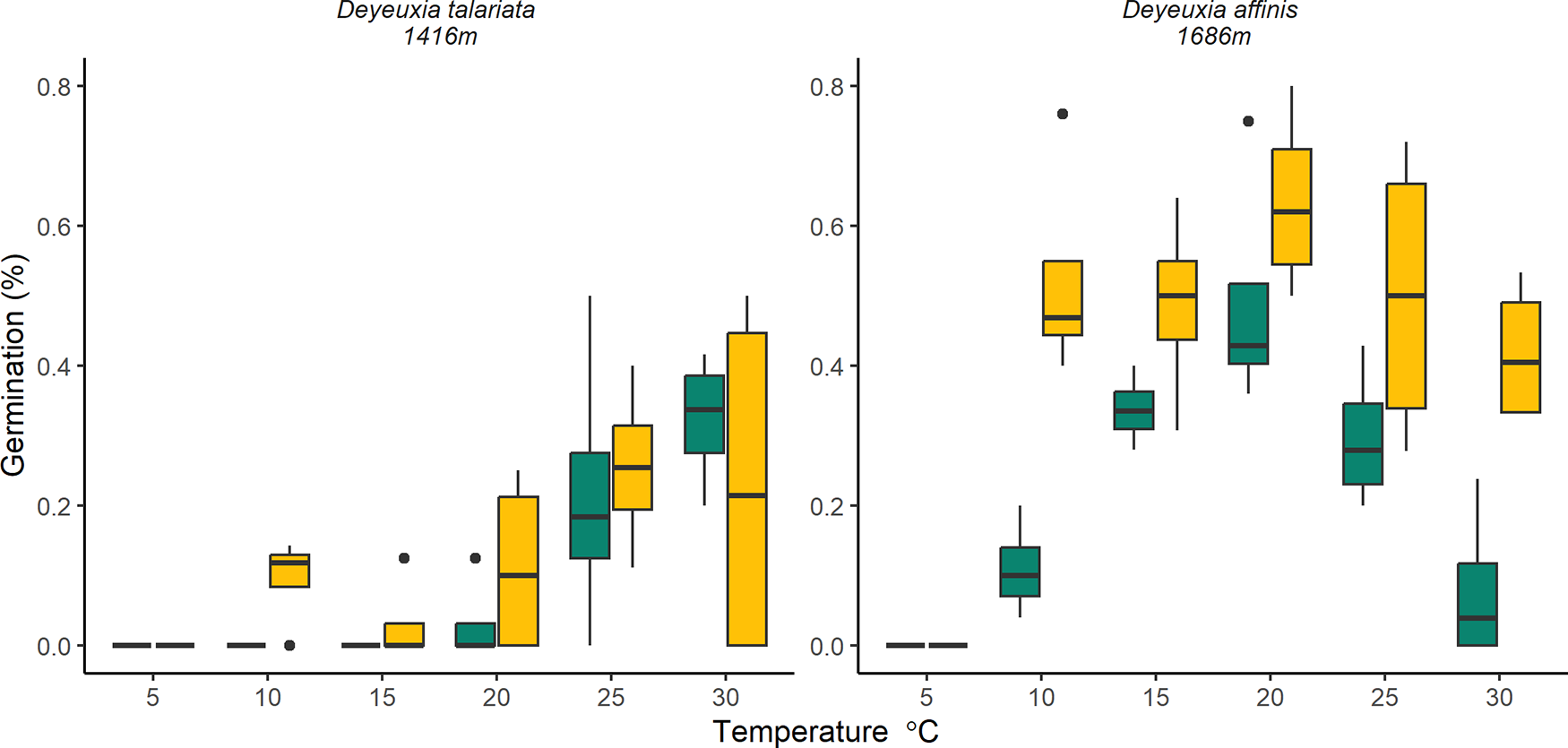
Germination rates were influenced by the interaction between seed treatment and species (χ2 = 5.221, df = 1, p = 0.022) and by temperature (χ2 = 9.681, df = 4, p = 0.046). There was no support for an interaction between treatment and temperature (χ2 = 7.544, df = 4, p = 0.110). D. affinis seeds treated with GA3 had higher germination rates when compared to the control (52% and 24%, respectively); however, this was not the case for D. talariata (GA3 = 14%, control = 11%; Table S3). Germination rates peaked at moderate temperatures with highest germination at 20°C and 25°C (31% and 27%, respectively) compared to 10°C, 15°C and 30°C (13%, 21% and 22%, respectively; Fig. 4; Table S2).
Germination percentage of seeds two Deyeuxia sp. Across six temperatures and seeds treated with GA3 (yellow) or untreated controls (green).

Figure 4 Long description
The image contains two box plots. The left plot represents the germination percentage of Deyeuxia talariata at 1416 meters across temperatures ranging from 5 to 30 degrees Celsius. The x-axis is labeled 'Temperature degrees Celsius' and the y-axis is labeled 'Germination percentage'. The right plot shows the germination percentage of Deyeuxia affinis at 1698 meters over the same temperature range. Both plots include data for seeds treated with GA (yellow) and untreated controls (green). The germination percentages vary with temperature, with visible differences between treated and untreated seeds.
Forbs – Brachyscome species
Brachyscome species showed different germination rates (χ2 = 967.612, df = 15, p < 0.001) that varied across temperatures (χ2 = 140.579, df = 4, p < 0.001) with a marginally non-significant difference between treatments (χ2 = 3.658, df = 1, p = 0.056). There was no interaction between species and seed treatment (χ2 = 14.704, df = 15, p = 0.473), or between seed treatment and temperature (χ2 = 2.907, df = 4, p = 0.574). Overall germination rates were highest for B. dentata and B. tadgellii (94%, 91%, 97%, 99%, 93%, respectively) compared to B. decipiens and B. spathulata. B. melanocarpa had the lowest germination (0.4%, 2%, 0.5% and 8%, respectively; Fig. 5; Table S4). Germination was highest at 10℃, 15℃ and 20℃ (59%, 65% and 59%, respectively) decreasing at 25℃ (39%) and further at 30℃ (14%). GA3 treatment tended to produce higher germination rates (49%) when compared to the control (42%; Table S2).
Germination percentage of seeds in 13 populations across eight species of Brachyscome across six temperatures with seeds treated with GA3 (yellow) or untreated controls (green).

Figure 5 Long description
Six box plots display germination rates of Brachyscome species across various temperatures. Each plot represents a different species: Brachyscome dentata, Brachyscome tadgellii, Brachyscome decipiens, Brachyscome melanocarpa, Brachyscome radicata and Brachyscome spathulata. The x-axis is labeled 'Temperature' in degrees Celsius, ranging from 0 to 30. The y-axis is labeled 'Germination' as a proportion. Each plot compares germination rates of seeds treated with GA and untreated controls, indicated by different colors. The plots show variations in germination rates across temperatures for each species.
Forbs – Brachyscome populations
We observed differences between populations of the same species. B. nivalis (Bogong High Plains Rocky ) had higher germination rates (76%) than conspecifics B. nivalis (Ruined Castle Rocky) (36%; χ2 = 53.461, df = 1, p < 0.001). Temperature had a parabolic effect (χ2 = 137.081, df = 4, p < 0.001) with higher germination at 15℃ and 20℃ when compared to 25℃ and 30℃ (p < 0.001). There was no effect of GA3 on germination (χ2 = 1.164, df = 1, p = 0.281) nor interactions between populations and treatment (χ2 = 3.891, df = 4, p = 0.421), or species and treatment (χ2 = 0.055, df = 1, p = 0.815; Fig. 6).
Germination percentage of seeds of Brachyscome nivalis, B. decipiens and B. tadgellii from different collections, across six temperatures and seeds treated with GA3 (yellow) and a control (green).

Figure 6 Long description
The image contains six box plots. The first row shows two plots: the left plot displays germination percentage against temperature in degrees Celsius for Brachyscome nivalis (BHR) and Brachyscome decipiens (PVR), with treatments indicated by color. The right plot shows germination percentage for Brachyscome decipiens (PVR) and Brachyscome nivalis (PVR) across temperatures. The second row contains two plots: the left plot illustrates the coefficient of velocity for Brachyscome decipiens (PVR) and Brachyscome nivalis (PVR) against temperature, while the right plot shows the same for Brachyscome decipiens (PVR) and Brachyscome nivalis (PVR). The third row includes two plots: the left plot presents the coefficient of velocity for Brachyscome nivalis (PVR) and Brachyscome nivalis (BHR) across temperatures and the right plot shows the coefficient of velocity for Brachyscome nivalis (PVR) and Brachyscome nivalis (BHR) against temperature. Each plot uses box plots to represent data distribution, with treatments differentiated by color.
B. decipiens (Pretty Valley Wet) had higher germination rates (3.3%) than B. decipiens (Pretty Valley Rocky) (0.9%; χ2 = 26.202, df = 1, p < 0.001; Fig. 6). Temperature also influenced germination rates (χ2 = 18.678, df = 4, p < 0.001) with germination higher at 10°C compared to 15°C and 30°C (p < 0.05); however, no other pairwise comparisons existed. There was no support for an effect of treatment (χ2 = 0.264, df = 1, p = 0.607) or the interactions with species (χ2 = 0.421, df = 1, p = 0.516) or temperature (χ2 = 0.943, df = 4, p = 943).
We found a difference between two collections (A, B) of B. tadgellii from different parts of the same area [Pretty Valley Wet A] and [Pretty Valley Wet B], which showed contrasting germination (A: 96%, B: 39%; χ2 = 86.320, df = 2, p < 0.001; Fig. 5c), while a cultivated low elevation population (progeny from collection A) B. tadgellii [Cranbourne Seed Orchard] had complete germination. Temperature showed higher germination rates at 15℃ and 20℃ (χ2 = 30.841, df = 4, p < 0.001) with lower germination at 10℃ (p < 0.002) and lower at 25℃ and 30℃ (p < 0.001; Fig. 6). No effects were observed with treatment (χ2 = 0.047, df = 1, p = 0.828) or its interactions with species (χ2 = 0.408, df = 2, p = 0.815) or temperature (χ2 = 3.273, df = 4, p = 0.513).
Discussion
We found the thermal responses and dormancy-breaking treatment effect varied among the studied taxonomic groups (trees, shrubs, forbs and grasses), although a common pattern can be drawn. Generally, dormancy was associated with elevation; species from high elevation exhibited a stronger response to GA3 than their lowland counterparts, irrespective of taxonomic group. The response to temperature also differed among species originating from different elevations: the optimum temperature for germination was higher for species from higher elevations, whereas lowland species exhibited favourable germination at a broader range of temperatures. Our findings suggest that dormancy and germination are influenced by environmental factors and evolutionary history, which has implications for the ability of these species to adapt to ongoing climate change.
Dormancy (responses to GA3)
Across the studied taxa, GA3 treatment promoted the germination of high-elevation species. This pattern was particularly evident in the Eucalyptus and Deyeuxia taxa (trees and grasses), where high-elevation species responded strongly to GA₃ treatment. In contrast, lower elevation species of these groups (E. pauciflora subsp. pauciflora and D. talariata) exhibited similar or higher germination rates without GA3 treatment (i.e. lack of physiological dormancy). These findings are consistent with global evidence that macroclimate shapes dormancy patterns: physiological dormancy (here broken with GA3) is more common in highly seasonal environments such as alpine regions, whereas non-dormancy is more common in stable, mild environments, such as those where the lowland species of our study were collected (Rosbakh et al., Reference Rosbakh, Carta, Fernández-Pascual, Phartyal, Dayrell, Mattana, Saatkamp, Vandelook, Baskin and Baskin2023).
The population of Eucalyptus pauciflora subsp. pauciflora examined in this study is situated at the lowest elevation within the subspecies’ known range. Notably, this subspecies exhibits the broadest elevational distribution among snow gums, extending from near sea level to the upper limits of the montane tree line. The absence of dormancy observed in this low-elevation population may reflect an elevation-dependent physiological response. To elucidate the role of elevation in dormancy expression, future research should investigate populations spanning the full elevational gradient of the subspecies.
In contrast to Eucalyptus and Deyeuxia, no Callistemon species responded to GA₃ regardless of seed origin (high or low elevation), suggesting an absence of physiological dormancy. This aligns with previous studies on Callistemon (Kim et al., Reference Kim, Walck, Hidayati, Merritt and Dixon2009; Kheloufi et al., Reference Kheloufi, Mansouri, Telali and Slimani2020) and supports the view that, although macroclimate influences dormancy, germination traits are also shaped by evolutionary history and tend to be conserved within closely related taxa (Gioria et al., Reference Gioria, Pyšek, Baskin and Carta2020; Carta et al., Reference Carta, Fernández-Pascual, Gioria, Müller, Rivière, Rosbakh, Saatkamp, Vandelook and Mattana2022). In other words, phylogenetically related species often share similar germination strategies (e.g. Hoyle et al., Reference Hoyle, Steadman, Good, McIntosh, Galea and Nicotra2015), which may explain the uniform lack of response to GA₃ observed in this genus. Furthermore, the consistently high germination across all tested temperatures may therefore reflect a ‘no dormancy’ strategy, whereby seeds germinate rapidly following dispersal when conditions are favourable. However, it remains unclear whether this pattern reflects a phylogenetic constraint or population-level variation (Seglias et al., Reference Seglias, Williams, Bilge and Kramer2018), and further work is needed to disentangle local adaptation from species-level effects.
Brachyscome species (forbs) exhibited mixed dormancy patterns; dormancy was present in some high elevation species but absent in others. This is consistent with the findings of Hirst et al. (Reference Hirst, Griffin, Sexton and Hoffmann2017), who observed variable germination responses among seven alpine Brachyscome species in a reciprocal germination experiment. Broad-niche species exhibited stronger germination responses, while narrow-range species displayed longer dormancy periods. Together, these results suggest an overall elevation-dormancy trend, which is heavily influenced by species-specific characteristics and local environmental conditions. Such variation is to be expected, given that alpine habitats are highly heterogeneous even at comparable elevations, due to differences in topography, aspect and soil characteristics (Körner and Hiltbrunner, Reference Körner and Hiltbrunner2021).
Effects of temperature on germination
The thermal germination responses of high-elevation and lowland species differed. High-elevation species generally germinated more successfully at warmer temperatures (20–30°C) and exhibited reduced germination at cooler temperatures (10–15°C). Overall, the requirement for high temperatures for germination is consistent with the findings for other Australian (Hirst et al., Reference Hirst, Vázquez-Ramírez, Rendall and Venn2025) and alpine species globally (Fernández‐Pascual et al., Reference Fernández‐Pascual, Carta, Mondoni, Cavieres, Rosbakh, Venn, Satyanti, Guja, Briceño, Vandelook, Mattana, Saatkamp, Bu, Sommerville, Poschlod, Liu, Nicotra and Jiménez-Alfaro2021) and is likely an adaptive strategy to delay seedling emergence until late spring when the risk of frost following snowmelt is reduced. Conversely, lowland species exhibited broader thermal tolerances, with germination observed at the majority of the tested temperatures, including the cooler ones (10–15°C), suggesting that in these environments other environmental factors, such as water availability (Moeslund et al., Reference Moeslund, Arge, Bøcher, Dalgaard, Odgaard, Nygaard and Svenning2013; Carta et al., Reference Carta, Fernández-Pascual, Gioria, Müller, Rivière, Rosbakh, Saatkamp, Vandelook and Mattana2022) may be more important drivers for germination and seedling establishment than temperature.
The above trend was less consistent among the Brachyscome species (forbs), which exhibited greater variability between different taxa. The differences observed between genera suggest that germination thermal responses are also shaped by evolutionary history, with implications for species responses to climate change. We used constant temperature settings in our methods to avoid confounding species-specific responses, as previous studies have demonstrated that taxonomic groups may respond differently to fluctuating temperatures (Liu et al., Reference Liu, Baskin, Baskin, Bu, Du and Ma2013). Germination can be affected by diurnal temperature changes; however, we did not test how temperature fluctuations affect seed germination in this study as minor fluctuations can significantly impact germination success (Hsu et al., Reference Hsu, Stuke, Bakker and Kim2024).
Population responses in Brachyscome species
When comparing within species, we observed population/collection differences. Beginning with the lowland species, we observed differences in the two B. dentata collections with the most recently wild collected (1 year old, from Amos Lane, northwest Victoria) showing the lowest germination rates (19%) compared to the other collection (10 year old, Truganina, Victoria; Table S1). B.tadgellii populations at one site (Pretty Valley wet area) but from different sites within it (Table S1) showed contrasting germination (A: 93%, B: 39%), while the progeny of Site A grown at a low elevation – B. tadgellii (Cranbourne) – showed contrasting responses to temperature with complete germination >15℃. In B. tadgellii, differences in dormancy and germination between the two collections are likely driven by microclimatic variation: collection A originated from a wetter site at the centre of an extensive seasonal depression, whereas collection B came from a comparatively drier site at the edge of the depression, near the species’ boundary. Similar microclimatic effects on dormancy have been reported previously (Terry et al., Reference Terry, Hardegree, Madsen, Roundy and St. Clair2022; Espinosa Del Alba et al., Reference Espinosa Del Alba, Fernández-Pascual and Jiménez-Alfaro2025). This finding is relevant given that the microclimate of collection B may represent future conditions for this habitat, as these alpine wetlands are expected to become progressively drier under climate change projections (Department of the Environment, 2015). The additional B. tadgellii population (and progeny of site A) was collected from a lowland population (Cranbourne, Victoria). There are substantial differences in the dormancy and germination response of all three, including significant differences in seed size, with potential cross-generation effects dictating thermal responses for the effect seen on the lowland Cranbourne progeny, which was absent of any dormancy mechanism. This potential transgenerational plasticity may benefit this species, indicating an adaptive mechanism to cope with environmental stresses; however, successive generations of B. tadgellii have not been tested at the lowland site to explore if this effect persists.
The high elevation populations of B. nivalis which both naturally occur on rocky outcrops [Basalt Hill] had higher germination rates (76%) than at another site [Ruined Castle] (36%). Microsite differences may also contribute to the contrasting germination responses observed between collections. Although both populations occur on exposed alpine rocky outcrops, they differ in local topography and substrate, which may influence soil moisture availability and the thermal conditions experienced during seed development and after dispersal. The two collections were obtained 1 year apart; however, seed viability remained high following storage, suggesting that differences in germination behaviour are unlikely to be driven by storage age alone. Instead, variation may reflect differences in maternal environmental conditions during seed maturation and fine-scale spatial heterogeneity. Such small-scale spatial and temporal variation has been shown to influence dormancy expression and germination timing in alpine species, even among populations occurring at similar elevations.
Study caveats and next steps
Our study shows that elevation is a good predictor of seed dormancy and thermal germination responses across four Australian plant groups. However, it is important to consider limitations, and we suggest the following areas for further research.
Firstly, the seeds that we used had been stored under seed bank conditions for extended periods (1–16 years), which may have affected their germination capacity and dormancy status. Although our inference is based on within-species comparisons between GA₃-treated and control seeds, the influence of storage effects cannot be entirely ruled out. Future studies comparing fresh and long-stored seeds will help to clarify the role of ex situ conservation in maintaining germination traits and explain seed behaviour under such storage conditions. Secondly, we subjected the seeds to constant temperature regimes to enable standardized comparisons between species. However, it is known that alternating temperatures (i.e. day/night) can influence germination responses, including thermal niche breadth. Incorporating fluctuating temperature regimes in future studies would make them more ecologically relevant. Thirdly, for species with very low germination (e.g. B. melanocarpa), it is difficult to fully distinguish between deep dormancy and reduced seed viability, even when seeds appear filled and viable. Interpretations for these species should therefore be made with caution. Finally, our study is based on single populations per species. Future work should include multiple populations at similar elevations but across different environmental gradients (e.g. soil moisture or snow cover) to better capture variation in dormancy and germination responses. Approaches such as reciprocal-elevation germination experiments would also help disentangle genetic adaptation from phenotypic plasticity, improving ecological inference in a climate change context.
Conclusion
This study demonstrates elevation is a strong predictor of seed dormancy and thermal germination responses across diverse Australian plant taxa. High-elevation species showed deeper physiological dormancy and narrower, warmer germination niches, while lowland species germinated across broader temperature ranges with minimal dormancy. These traits suggest that alpine species are particularly vulnerable to climate change. Their reliance on specific temperature cues and dormancy-breaking conditions – such as cold stratification – makes them sensitive to warming trends, reduced snow cover and shifting seasonality. In contrast, lowland species may be more resilient due to broader thermal tolerances and reduced dormancy constraints. However, population-level variation – especially in B. tadgellii – highlights potential for local adaptation and transgenerational plasticity, offering insights into species resilience. Successful germination of seeds stored for over a decade underscores the value of seed banking for long-term conservation. Together, these findings provide critical insights into how seed traits mediate species’ responses to environmental change and inform strategies for restoration, seed sourcing and biodiversity conservation in a warming world.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S096025852610018X.
Acknowledgements
We acknowledge the traditional owners of the lands on which this research took place. We thank Adam Cross for his insightful comments in early conversations on seed dormancy status and Bob Hare, a Victorian Conservation Seedbank volunteer for assisting with the laboratory set-up.
Author contributions
Megan J. Hirst: Conceptualization, Methodology, Writing – original draft, Writing – review & editing; Daniel J. White: Writing – review & editing; Anthony Rendall: Formal analysis, Writing – review & editing, Visualisation; Ary A. Hoffmann: Methodology, Writing – review & editing; Jerónimo Vázquez-Ramírez: Writing – original draft, Writing – review & editing.
Funding statement
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Competing interests
The authors declare no conflicts of interest.
Data accessibility statement
All the data and R code used in this study are available upon request.






 Open access
Open access