


Management Implications
Effective management of noxious weeds is a primary challenge in Canadian Aspen Parkland restoration. This study demonstrated that aminopyralid + metsulfuron-methyl treatments using aminopyralid and metsulfuron provided reduced cover of noxious weeds while promoting establishment of seeded native grasses. However, managers should be aware that aminopyralid + metsulfuron-methyl applications also reduced native shrub and forb cover, potentially lowering structural diversity and limiting habitat for pollinators and wildlife. Selective herbicide application combined with protective measures for shrubs (e.g., shielding desirable woody plants or timing applications to avoid sensitive growth stages) may help reduce non-target impacts. Importantly, mowing provided little additional benefit when combined with aminopyralid + metsulfuron-methyl. Aminopyralid + metsulfuron-methyl–alone treatments effectively controlled invasive plants and supported native species recovery while avoiding additional costs and soil disturbance associated with mowing. This finding suggests that, under similar conditions, mowing may not be a necessary component of integrated management programs, thus representing an avoidable expense. While aminopyralid + metsulfuron-methyl application improved conditions for seeded native grasses, the overall seeded species establishment remained limited. Including species adapted to early establishment or disturbance-tolerant conditions may increase establishment reliability under competitive or variable site conditions.
Introduction
Managing invasive plants in disturbed ecosystems remains a major challenge, particularly during vegetation restoration, when seeded native species may have to establish in competition with aggressive invasive plants (Daehler Reference Daehler2003; D’Antonio and Chambers Reference D’Antonio and Chambers2006). Many invasive plants rapidly colonize post-disturbance environments, outcompeting native vegetation and altering soil and resource availability in ways that reinforce their dominance (DiTomaso Reference DiTomaso2000; Pauchard and Shea Reference Pauchard and Shea2006). While seeding native plant species is a widely used strategy to suppress invasives and restore plant communities (Endress et al. Reference Endress, Parks, Naylor, Radosevich and Porter2012; Schuster et al. Reference Schuster, Wragg and Reich2018; Scotton and Andreatta Reference Scotton and Andreatta2021; Torok et al. Reference Torok, Deak, Vida, Valko, Lengyel and Tothmeresz2010; Walsh and Rose Reference Walsh and Rose2022), success remains highly variable, largely depending on post-seeding management. Without targeted strategies to mitigate competition and enhance native seedling establishment, invasive plants often persist, limiting long-term vegetation restoration (Orrock et al. Reference Orrock, Witter and Reichman2008). Therefore, identifying effective post-seeding interventions is essential to improving native plant reestablishment and ensuring sustained restoration success.
In Aspen Parkland ecosystems of the Northern Great Plains of North America, two particularly problematic plant species are Canada thistle [Cirsium arvense (L.) Scop.] and smooth brome (Bromus inermis Leyss.). Both species exhibit aggressive growth habits that allow them to outcompete native plants for light, nutrients, and space (Bossard et al. Reference Bossard, Randall and Hoshovsky2000; Grace et al. Reference Grace, Smith, Grace, Collins, Stohlgren, Moore and Wallace2000; Rana and Rana Reference Rana and Rana2016). This can lead to significant losses in biodiversity and economic value (Otfinowski et al. Reference Otfinowski, Kenkel and Catling2007; Patterson Reference Patterson1995; Tiley Reference Tiley2010). Suppressing these invasive species is a priority for land managers seeking to restore grassland ecosystems. Although herbicide application, mowing, grazing, prescribed burning, and biological control have demonstrated varying degrees of effectiveness in suppressing these species (Bahm et al. Reference Bahm, Barnes and Jensen2011; Cole et al. Reference Cole, King, Oyarzun, Dietzler and McClay2007; Grekul and Bork Reference Grekul and Bork2007), their feasibility differs among species and management contexts.
For C. arvense, repeated mowing can reduce root carbohydrate reserves and create wounds that facilitate biological control pathogen entry (Kluth et al. Reference Kluth, Kruess and Tscharntke2003). Despite these physiological vulnerabilities, biological control of C. arvense has produced limited and inconsistent results, and no effective biological control agents exist for B. inermis (Jacobs et al. Reference Jacobs, Sciegienka and Menalled2006; Muller and Nentwig Reference Muller and Nentwig2011). In industrial, post-disturbance, or construction-associated landscapes, safety, access, and regulatory constraints may restrict the use of prescribed burning or grazing. These limitations may help explain why, across many restoration settings, land managers continue to rely primarily on chemical and mechanical tools as the foundation of invasive plant control. Mowing can complement herbicide treatments by increasing herbicide uptake and improving overall control efficacy when the two methods are integrated (Miller Reference Miller2016). Herbicide application, mowing, and their combined use represent practical and effective strategies for suppressing invasive species while minimizing additional disturbance to establishing native vegetation. Together, these broader management realities provide a clear rationale for emphasizing chemical and mechanical treatments in restoration-focused vegetation management.
Integrated vegetation management, combining herbicide application, mowing, and native species seeding, is commonly used to control invasive plants in North American grasslands. Herbicides such as glyphosate, applied alone or with imazapyr and sulfosulfuron, effectively reduce B. inermis (Bahm et al. Reference Bahm, Barnes and Jensen2011; Brown et al. Reference Brown, Dhar and Naeth2024). Similarly, C. arvense is managed with a variety of herbicides, including amitrole, dicamba, clopyralid, glyphosate, MCPB/MCPA, dichlobenil, 2,4-D, and metsulfuron, which are often applied in combination for greater efficacy. However, broad-spectrum herbicides such as glyphosate can lead to broader declines in non-target plant diversity and shoot growth of native species such as green needle grass [Nassella viridula (Trin.) Barkworth] (Olszyk et al. Reference Olszyk, Pfleeger, Nash and Plocher2024; Power et al. Reference Power, Kelly and Stout2013). Therefore, selective herbicides are preferred during restoration where native grasses are being established.
Some selective herbicides promote native grass establishment; for example, high concentrations of atrazine promote little bluestem [Schizachyrium scoparium (Michx.) Nash] establishment, while 2,4-D enhances blue grama [Bouteloua gracilis (Willd. ex Kunth) Lag. ex Griffiths] in warm-season grass reseeding (Bragg and Sutherland Reference Bragg and Sutherland1989). More recently, aminopyralid has emerged as a preferred alternative due to its lower application rates and reduced site restrictions (Enloe et al. Reference Enloe, Lym, Wilson, Westra, Nissen, Beck and Halstvedt2007). Aminopyralid effectively suppresses C. arvense while also promoting increases in native grass species, both in already-established stands and in pre-seeding restoration contexts (Almquist and Lym Reference Almquist and Lym2010; Lym et al. Reference Lym, Becker, Moechnig, Halstvedt and Peterson2017). Metsulfuron-methyl (Escort® XP, Environmental Science U.S., LLC, Cary, NC, USA) effectively controls C. arvense and is registered for selective weed management in native and forage grass systems, with the label indicating tolerance by a wide range of cool-season perennial grasses, including B. inermis, Festuca spp., N. viridula, and multiple wheatgrass species, which can be safely reseeded within 1 to 4 mo after application, depending on the recommended herbicide rate (14 to 69.2 g ha−1 or higher for certain species; Environmental Science US LLC 2022). Turfgrass studies show that perennial warm-season grasses such as buffalograss [Bouteloua dactyloides (Nutt.) J.T.] tolerate label-rate applications of metsulfuron, with no reduction in establishment or sustained injury relative to untreated controls (McCarty and Colvin Reference McCarty and Colvin1992). However, spring applications of metsulfuron alone and in combination with aminopyralid or other herbicides in tall fescue [Schedonorus arundinaceus (Schreb.) Dumort., nom. cons.] hayfields have been shown to successfully reduce tall fescue height, seedhead production, and forage yield by up to 45%, 88%, and 63%, respectively (Sather et al. Reference Sather, Roberts and Bradley2013). Additionally, metsulfuron can control bahiagrass [Paspalum notatum Flueggé], but tolerance varies among cultivars and across application rates (Bunnell et al. Reference Bunnell, Baker, McCarty, Hall and Colvin2003). The combination of aminopyralid and metsulfuron-methyl broadens control among invasive forbs and certain problematic non-native species while maintaining safety for many native perennial grasses, making it a suitable option in restoration contexts where preserving seeded grass establishment is important. However, the activity of this combination is largely limited to broadleaf species and therefore provides no control of perennial grasses such as B. inermis. Mowing has been shown to reduce vegetative reproduction of B. inermis by limiting rhizome and bud production (Xu et al. Reference Xu, Young, Boe, Hendrickson and Troelstrup2016), and studies indicate that frequent and severe defoliation can further reduce stand vigor (Brown et al. Reference Brown, Dhar and Naeth2024; Palit and DeKeyser Reference Palit and DeKeyser2022). However, lower-intensity mowing is generally more effective when combined with other management treatments (Willson and Stubbendieck Reference Willson1996). Together, the aminopyralid + metsulfuron-methyl combination and mowing can provide complementary control of both invasive forbs and undesirable perennial grasses, offering a more integrated and effective approach for restoration vegetation management.
Studies testing invasive plant suppression in the Aspen Parkland, in the Northern Great Plains are limited. The Aspen Parkland consists of a mosaic of woody species and grasslands, is dominated by C3 grasses, and typically has fertile soils and a cold climate (Kupsch et al Reference Kupsch, France, Looen, Burkinshaw, Willoughby and MacNeil2013). Furthermore, the region’s history of agricultural land use has contributed to the persistence and spread of invasive species that complicate restoration efforts. Grasslands within the Aspen Parkland are considered a threatened ecosystem, making their restoration critical for maintaining biodiversity and ecosystem services (Locklear Reference Locklear2017). Restoration is often costly, making it essential to identify successful and cost-effective methods to ensure the long-term success of these efforts. Identifying vegetation management options that enhance seeded species establishment, and existing native species, while suppressing weedy species is important for the restoration of endangered ecosystems, such as grassland areas within the Aspen Parkland ecoregion. This study evaluated the effects of mowing, aminopyralid + metsulfuron-methyl application, and a combined treatment on native and weedy plant biomass, species richness, diversity, cover, and community composition over a 2-yr management period (2023 to 2024). The treatments were applied after the native grass mix had already undergone one full year of establishment following seeding in 2021, allowing us to assess vegetation responses to consecutive years of post-establishment management on a disturbed site in the Aspen Parkland.
Materials and Methods
Study Site
The study was conducted in the river valley floodplain area adjacent to EPCOR’s Kīsikāw Pīsim (“daylight sun” in Cree) solar farm located in Edmonton, AB, Canada (53.46765°N, 113.60856°W). The site lies at an elevation of approximately 630 m, receives an annual precipitation of 476 mm, and average monthly temperatures range from −12 C in January to 15 C in July (Statistics Canada Reference Statistics2024). The study area received an average annual precipitation of approximately 327 mm in 2021, 356 mm in 2022, and 410 mm in 2023, based on data from the Edmonton Blatchford weather station that is 13.2 km from the study site (Environment and Climate Change Canada 2024). Direct precipitation data were not available for the solar farm itself, and some daily records were missing at the Blatchford station; therefore, annual totals represent the sum of available daily observations for each year and may slightly underestimate true precipitation. The site is in the Aspen Parkland ecoregion, historically characterized by a mosaic of forest and grassland ecosystems with high biodiversity and Chernozemic soils (Bird Reference Bird1930). Before restoration, the area consisted of abandoned farmland dominated by weedy species, including C. arvense, field sowthistle (Sonchus arvensis L.), yellow toadflax (Linaria vulgaris Mill.), and B. inermis.
Experimental Design
In 2021 early summer, the study site was drill-seeded with a seed mix comprising seven native prairie grass species at a rate of 30 kg ha−1 (Table 1). Before seeding, all dead vegetation and debris were removed, and the upper 5 cm of topsoil was lightly tilled to improve seed-to-soil contact. Seeding was completed using a Brillion drill seeder (Brillion Farm Equipment, Brillion, WI, USA) at a rate of 30 kg ha−1, with seed placed at a depth of approximately 6 to 12 mm. Seeding included supplemental watering, which was applied to support seed establishment. All seeds were verified as weed-free by accredited seed testing laboratories. “Weed-free” refers to the absence of listed noxious/prohibited weeds per lab certification; a subsample indicated a small quantity of non-target native seed, slender wheatgrass [Elymus trachycaulus (Link) Gould ex Shinners] (∼33 seeds per 25 g); no other contaminants were detected. In July 2022, twenty-four 2 by 4 m plots were established, with a 2-m buffer between plots. Plots were randomly assigned to one of four treatments: (1) aminopyralid + metsulfuron-methyl application only (Aminopyralid + metsulfuron-methyl), (2) mowing application only (Mow), (3) a combination of aminopyralid + metsulfuron-methyl application and mowing (Combination), and (4) no treatment (Control). ClearView® herbicide (Corteva Agriscience Canada, Calgary, AB, Canada), is a selective, postemergence herbicide containing 52.5% aminopyralid (present as the potassium salt) and 9.45% metsulfuron-methyl as active ingredients. It is registered for use in rangeland, pastures, rights-of-way, and other non-crop areas across Canada for control of annual and perennial broadleaf weeds, invasive plants, and shrubs (Corteva Agriscience Reference Agriscience2025). A professional applicator applied ClearView® herbicide (Corteva Agriscience Canada) at 230 g product ha−1, equivalent to 120.8 g aminopyralid ai ha−1 and 21.7 g metsulfuron-methyl ai ha−1 in late July of 2022 and again in late July of 2023 on the Aminopyralid + metsulfuron-methyl and Combination plots, using a backpack sprayer. Mowing treatments were applied to the Mow and Combination plots in late August 2022 and again in late August 2023 using an EGO POWER+ 56 V Powerload String Grass Trimmer with Line IQ (EGO Power+, Anderson, SC, USA) to achieve a vegetation height of 5 to 10 cm.
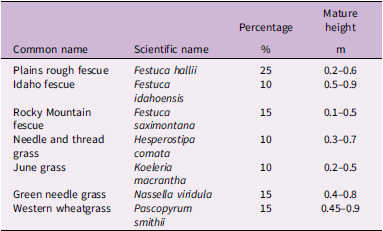
Seeded species, percent of seeds in the mix, and height at maturity used in this study in Edmonton, Alberta, Canada.

Data Collection and Analysis
Vegetation cover surveys and biomass sampling were conducted using 50 by 50 cm and 25 by 50 cm quadrats respectively, with two cover and two biomass quadrats placed in each plot. Baseline vegetation of 1-yr establishment cover was collected in late July 2022, before the application of any treatments. In central Alberta, the growing season typically runs from late May to early September, with both cool-season grasses and forbs reaching peak biomass in July. For this reason, mid-July is the standard monitoring window used by provincial agencies and is appropriate for assessing treatment effects in this region. In 2023 and 2024, vegetation cover surveys and biomass collections were conducted in July, also before treatment application those years. Biomass was clipped at ground level, sorted into four groups (forbs, grasses, B. inermis, and litter), oven-dried at 50 C for 48 h, and weighed. For analysis, B. inermis was combined with grasses to represent total grass productivity, a decision made for practical and analytical reasons: mixed swards can introduce field identification error during harvest. Accordingly, “grass biomass” includes B. inermis, while species-specific responses of B. inermis are assessed from the cover dataset. All clipped biomass was fully removed from the plots following the mowing treatment. For cover estimates, plants were identified to species and then classified into five groups based on their origin and seeding status: seeded (seven native species that were intentionally seeded), noxious (invasive species identified as problematic based on the Weed Control Act (Province of Alberta 2016), native (non-seeded species native to the Aspen Parkland), introduced (species not native to the Aspen Parkland that are not designated as noxious), and native shrubs (non-seeded native woody species). This status-based cover summary aligns with restoration and weed-control objectives and complements the functional grouping used for biomass by capturing species-specific responses and community composition.
To assess the effects of the treatments (Control, Aminopyralid + metsulfuron-methyl, Mow, and Combination) on biomass, plant community composition, species diversity, richness, and species cover, a series of statistical analyses were conducted in R Studio (v. 2024.12.0+467) with the threshold for significance set at α = 0.05. Biomass data, including grasses, forbs, litter, and total biomass (sum of all three), were analyzed using Kruskal–Wallis rank-sum tests or Welch’s ANOVA, depending on whether assumptions of normality and homogeneity of variances were met. Pairwise comparisons were performed using Nemenyi’s test following Kruskal–Wallis tests. For Welch’s ANOVA, a Tukey-type test for unequal variances was applied using the tukeySEM() function from the semTools package, which accounts for heteroscedasticity.
For each functional group (seeded species, noxious weeds, native species, introduced species, and native shrubs), species richness, and Hill’s number for Simpson diversity (q = 2) were calculated at the plot level based on species-level percent cover data (Chao et al. Reference Chao, Gotelli, Hsieh, Sander, Ma, Colwell and Ellison2014). Cover, richness, and diversity metrics were analyzed using the same statistical approach as for biomass: Kruskal–Wallis tests or Welch’s ANOVA, with subsequent Nemenyi or Tukey post hoc tests for unequal variance, respectively, depending on whether test assumptions were met. For examination of seeded species in greater detail, species-level responses were also analyzed using the same approach, to assess whether individual species differed in their sensitivity to treatments or exhibited distinct patterns of establishment.
Species evenness was assessed using the
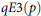 $qE3\left( p \right)$
index, originally proposed by Jost (Reference Jost2010) and generalized by Chao and Ricotta (Reference Chao and Ricotta2019), which is a Hill number–based evenness measure that expresses the normalized slope of the Hill diversity profile between diversity orders q = 0 and q > 0. At q = 2, the index aligns with formulations by Kvaalseth (Reference Kvaalseth1991), and reflects sensitivity to dominant species. When q = 1, it reduces to Heip’s (Reference Heip1974) evenness. This makes
$qE3\left( p \right)$
index, originally proposed by Jost (Reference Jost2010) and generalized by Chao and Ricotta (Reference Chao and Ricotta2019), which is a Hill number–based evenness measure that expresses the normalized slope of the Hill diversity profile between diversity orders q = 0 and q > 0. At q = 2, the index aligns with formulations by Kvaalseth (Reference Kvaalseth1991), and reflects sensitivity to dominant species. When q = 1, it reduces to Heip’s (Reference Heip1974) evenness. This makes
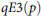 $\;qE3\left( p \right)$
both mathematically interpretable and comparable across communities with differing richness. Higher values of qE3(p) denote higher evenness.
$\;qE3\left( p \right)$
both mathematically interpretable and comparable across communities with differing richness. Higher values of qE3(p) denote higher evenness.
 $qE3\left( p \right) = \;{{{{\left( {\sum\nolimits_{i = 1}^s {p_i^q} } \right)}^{1/{{\left( {1 - q} \right)}}}- 1}} \over {S - 1}}\; = \;{{_\;^qD - 1} \over {S - 1}}$
$qE3\left( p \right) = \;{{{{\left( {\sum\nolimits_{i = 1}^s {p_i^q} } \right)}^{1/{{\left( {1 - q} \right)}}}- 1}} \over {S - 1}}\; = \;{{_\;^qD - 1} \over {S - 1}}$
In this equation, S is the number of species, p is the relative abundance of focal species i. The summation in the numerator of the first term calculates the Hill number, D for order q, where q = 1 to 2.
Plant species composition was analyzed using permutational multivariate analysis of variance (PERMANOVA) with Bray-Curtis dissimilarities to test the effect of treatment, and pairwise PERMANOVA comparisons were conducted to explore specific differences between treatment groups. We specifically analyzed mean percent cover of B. inermis and C. arvense in 0.25-m2 quadrats from 24 plots per treatment: Control, Mow, Aminopyralid + metsulfuron-methyl, and Combination (mowing + aminopyralid + metsulfuron-methyl), and they were resampled at consistent phenology each July in 2022 to 2024. Because zeros were common, we assessed distributional fit (quantile deviation, dispersion, outliers) with DHARMa (Hartig Reference Hartig2016). Time, treatment, and their interaction effects were modeled with generalized linear mixed models (GLMMs) in glmmTMB, using a negative binomial distribution (family = nbinom2, log link) and a random intercept for plot to account for repeated measures. For B. inermis, we fit a model with a constant zero-inflation term (∼1); for C. arvense, we used a zero-inflated negative binomial with the zero-inflation component. Model-estimated marginal means were obtained with emmeans on the response scale (back-transformed means ± 95% CI), with pairwise contrasts among treatments within years and among years within treatments to quantify mean differences. Predicted means were visualized in ggplot2 as treatment-specific trend lines.
Results and Discussion
Treatment Effects on Plant Biomass
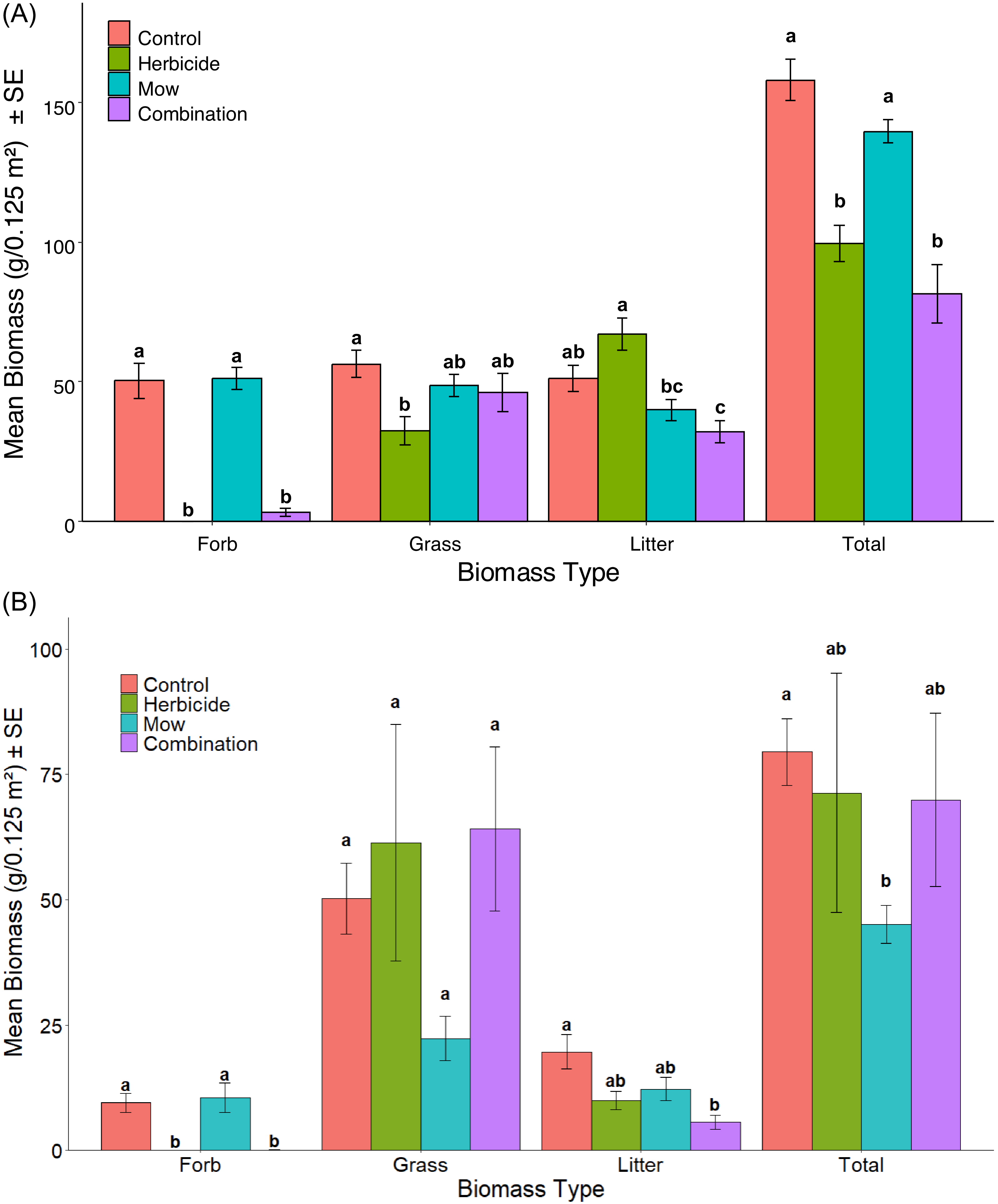
Vegetation biomass was affected by treatments with some variation from year to year among the different plant functional groups (Figure 1A and 1B). The pattern of forb biomass response to treatments in 2023 and 2024 was identical. In both years, forb biomass was significantly lower in the Aminopyralid + metsulfuron-methyl and Combination treatments compared with the Control and Mow treatments, while no significant differences were observed between Control and Mow or between the two herbicide-based treatments. This was supported by Kruskal–Wallis tests (2023: χ2 = 38.56, df = 3, P < 0.001, Figure 1A; 2024: χ2 = 35.92, df = 3, P < 0.001, Figure 1B) and post hoc Nemenyi comparisons, which confirmed significant reductions in forb biomass in both herbicide-based treatments relative to Control and Mow (all P < 0.001, except Combination vs. Mow in 2024 with P < 0.05). In both years, forb biomass in the Aminopyralid + metsulfuron-methyl and Combination treatments was nearly eliminated, whereas forbs remained present in the Control and Mow treatments (Figures 1 and 2). This outcome aligns with aminopyralid’s activity on many broadleaf weeds, while signaling that control is dependent on species and growth stage (Gramig and Ganguli Reference Gramig and Ganguli2015). ClearView® was applied at 230 g product ha−1 in both 2022 and 2023, a within-label yet relatively high application rate; such a repeated application of which should contribute to reductions of targeted forb species.
Mean biomass (g 0.125 m− 2) ± SE of forbs, grasses, litter, and total vegetation in July 2023 (A) and 2024 (B) under four vegetation-management treatments in Edmonton, Alberta, Canada: Control (no treatment), Herbicide (aminopyralid + metsulfuron-methyl), Mow, and Combination (mow + herbicide). Different letters above bars indicate significant differences (P < 0.05) among treatments within each biomass type; treatments that share a letter are not significantly different. Comparisons are made within biomass categories (Forb, Grass, Litter, and Total), not across categories. The 2023 biomass data (A) were collected in July 2023, approximately 1 yr after the initial herbicide application before mowing (July 2022) and following one mowing event implemented in 2022. The 2024 biomass data (B) were collected in July 2024, approximately 2 yr after the initial (July 2022) and 1 yr after second (July 2023) herbicide and mowing application.

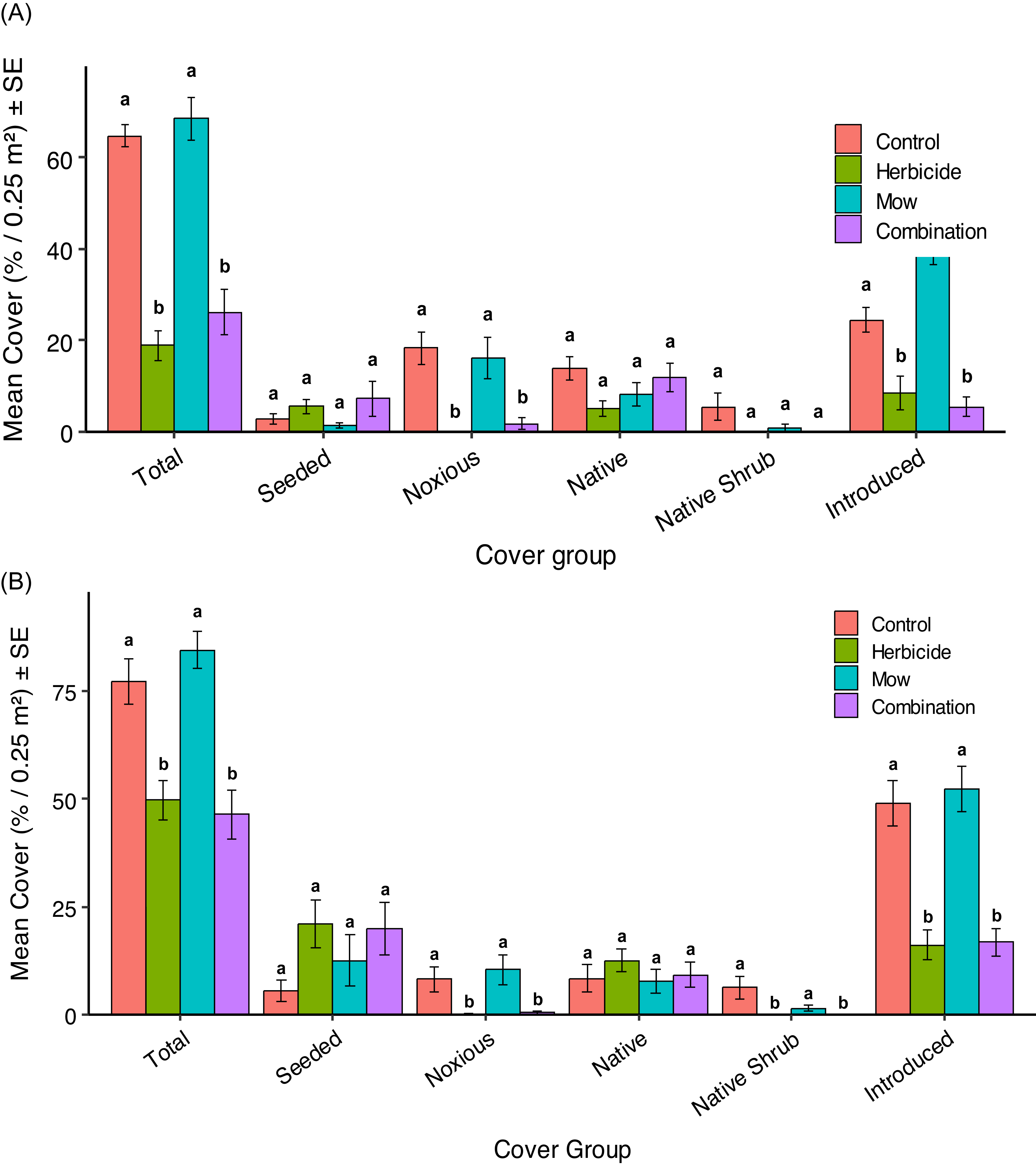
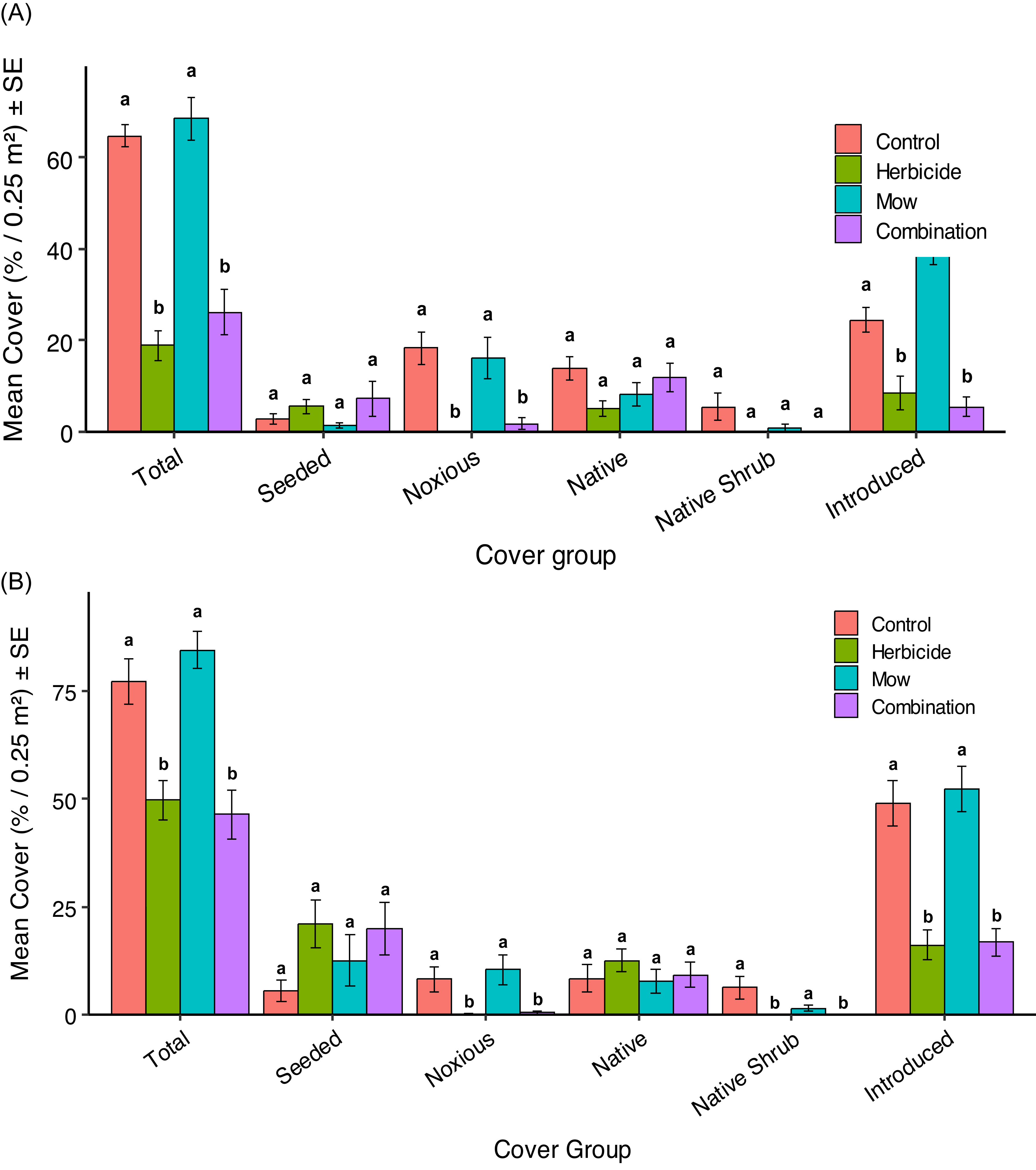
Mean vegetation cover (%0.25 m− 2) ± SE for six plant groups: Total, Seeded, Noxious, Native, Native Shrub, and Introduced in 2023 (A) and 2024 (B) under four vegetation-management treatments in Edmonton, Alberta, Canada: Control (no treatment), Herbicide (aminopyralid + metsulfuron-methyl), Mow, and Combination (mow + herbicide). Different letters above bars indicate significant differences (P < 0.05) among treatments within each cover group. Treatments sharing a letter are not significantly different, while treatments with different letters differ significantly. Comparisons are made within each plant group category rather than across categories. The 2023 cover data (A) were assessed in July 2023, at 1 yr after the initial herbicide application (July 2022) and following one mowing event. The 2024 cover data (B) were assessed in July 2024, approximately 2 yr after the initial (July 2022) and 1 yr after second (July 2023) herbicide and mowing application.
Treatment effects on grass biomass were not consistent between years. In 2023, grass biomass differed significantly among treatments (ANOVA: F(3, 44) = 3.61, P = 0.0205), and pairwise contrasts indicate one significant comparison, the Aminopyralid + metsulfuron-methyl treatment had lower grass biomass than the Control (P = 0.012). In 2024, grass biomass also showed a significant treatment effect (Kruskal–Wallis χ2 = 9.13, df = 3, P = 0.0277), but no pairwise comparisons were significant (all P > 0.05). The Combination treatment maintained grass biomass levels comparable to the Control and Aminopyralid + metsulfuron-methyl treatments. This may have reflected reduced competition from forbs and other broadleaf weeds following Aminopyralid + metsulfuron-methyl application, allowing grasses to persist despite mowing. This pattern was consistent with findings from Maron and Jefferies (Reference Maron and Jefferies2001), who reported that mowing and biomass removal significantly reduced aboveground biomass while increasing belowground biomass, likely due to growth limitation from the removal of meristems during cutting. Their study also suggested that mowing suppressed annual grass growth and enhanced the establishment of both native and exotic forbs. Herbicide treatments alone have been shown to maintain native cool-season grasses (Aulakh Reference Aulakh2020; Lym et al. Reference Lym, Becker, Moechnig, Halstvedt and Peterson2017), and combining with mowing has also been reported to improve control of competitive species and support native grass establishment (Robertson et al. Reference Robertson, Hickman, Harmoney and Leslie2013). However, the absence of a detectable mowing effect in our study may reflect the timing and frequency of mowing. Weekly mowing across growing seasons has been shown to facilitate forb establishment in grass-dominated communities without reducing grass biomass (Williams et al. Reference Williams, Jackson and Smith2007). Our treatments were only mowed once annually.
Total biomass was significantly affected by treatments in both years, although the strength and pattern of responses differed. In 2023, total biomass differed significantly among treatments (ANOVA: F(3, 44) = 22.58, P < 0.001, Figure 1A). Control and Mow treatments had the highest biomass, while Aminopyralid + metsulfuron-methyl and Combination treatments showed large reductions. Total biomass was significantly lower in Aminopyralid + metsulfuron-methyl (P < 0.001) and Combination (P < 0.001) relative to Control, and similarly lower than Mow (P = 0.002 and P < 0.001, respectively). In 2024, total biomass was significantly affected by treatment (Kruskal–Wallis: χ2 = 9.48, df = 3, P = 0.0236; Figure 1B). Pairwise Wilcoxon tests indicated that total biomass in the Mow treatment was significantly lower than in Control (P = 0.0039), while other comparisons were not significant. The reduction in total biomass in Mow (mean = 45.1 g 0.125 m− 2) compared with Control (79.5 g 0.125 m− 2) reflected the pattern observed in grass biomass, which comprised most of the total mass.
Litter responded strongly to treatments in both years, but in opposite directions. In 2023 (ANOVA: F(3, 44) = 10.88, P < 0.001, Figure 1A), Aminopyralid + metsulfuron-methyl plots accumulated the highest litter, while the Combination treatment had the lowest. Post hoc tests showed that litter biomass in Mow and Combination was significantly lower than in the Aminopyralid + metsulfuron-methyl treatment (both P < 0.001). In 2024, treatment effects remained significant (Welch’s ANOVA: F(3, 23.58) = 5.59, P = 0.0048, Figure 1B), but the pattern reversed, with the Combination treatment showing substantially lower litter biomass than Control (mean = 5.68 vs. 19.70 g 0.125 m− 2; P < 0.001), while all other pairwise comparisons were nonsignificant. These results suggest that the Combination of mowing and aminopyralid + metsulfuron-methyl substantially reduced litter biomass, which may alter soil microclimate conditions and affect vegetation recovery in restored plots.
Treatment Effects on Plant Community Composition
Plant species composition responses to treatments were more distinct in 2023 compared with 2024. In 2023, PERMANOVA indicated that treatments affected community structure (pseudo-F(3, 44) = 5.44, P = 0.001) with all treatment pairs differing significantly (all P < 0.004) except the Aminopyralid + metsulfuron-methyl and Combination (P = 0.061). In 2024, treatment effects persisted but were less pronounced (pseudo-F(3, 44) = 3.43, P = 0.001), with only two significant pairwise contrasts (Mow vs. Combination and Control vs. Aminopyralid + metsulfuron-methyl; both P = 0.001). These results indicated that mowing maintained a community composition similar to the Control, both dominated by introduced and noxious species, while the two treatments with aminopyralid + metsulfuron-methyl shifted composition by suppressing invasive species and promoting native grass establishment (Table 2). The similarity between the Aminopyralid + metsulfuron-methyl and Combination treatments suggested that mowing did not change composition beyond the effect of Aminopyralid + metsulfuron-methyl application alone. The similarity between the Aminopyralid + metsulfuron-methyl and Combination treatments is more likely explained by the strong herbicide effect, which eliminated forbs in both treatments, resulting in grass-dominated communities regardless of mowing. The lack of mowing effectiveness may have been due to the specific mowing strategy used, including factors such as equipment type, mowing season, height, and frequency (Butler et al. Reference Butler, Brouder, Johnson and Gibson2013; Pirchio et al. Reference Pirchio, Fontanelli, Frasconi, Martelloni, Raffaelli, Peruzzi and Grossi2018; Rinella et al. Reference Rinella, Jacobs, Sheley and Borkowski2001). The persistence of introduced grasses under mowing was likely due to their origin as agronomic forage species, which are well adapted to repeated defoliation and able to recover quickly after mowing (Ferraro and Oesterheld Reference Ferraro and Oesterheld2002).
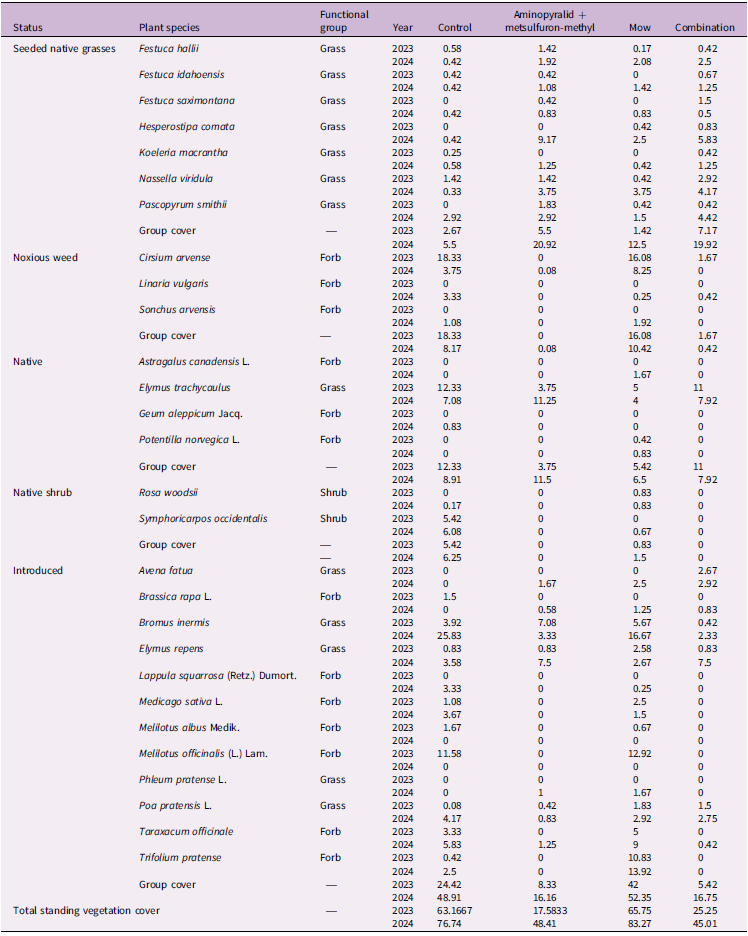
Percent cover of plant species by treatment, grouped by status and functional group in 2023 and 2024 in Edmonton, Alberta, Canada. a
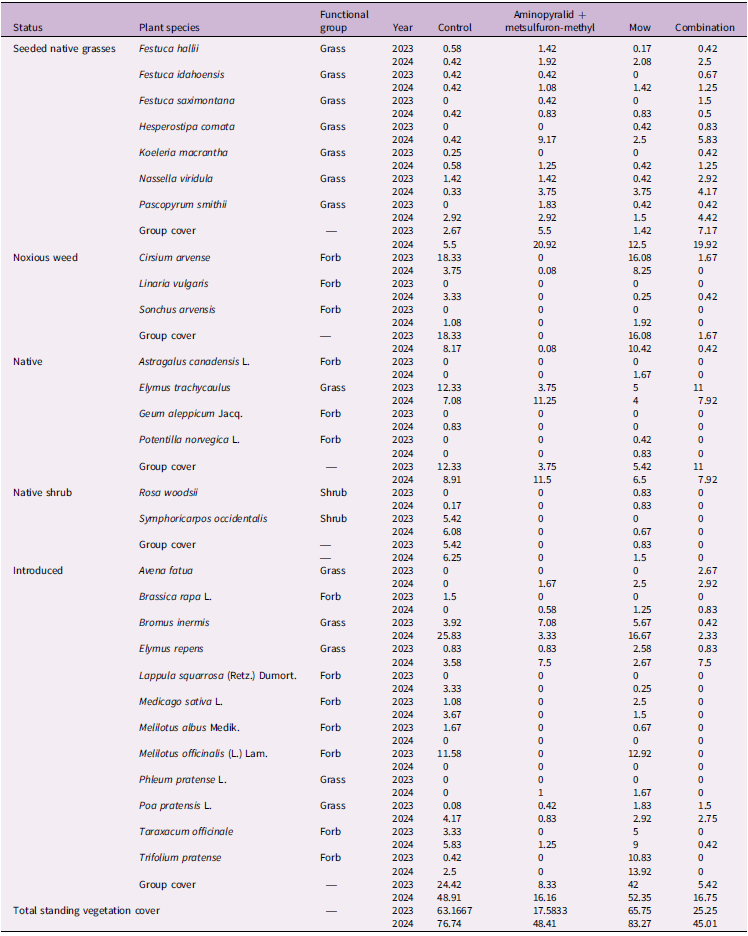
a Values represent the mean percent cover across all plots within each treatment for each species. The final row presents total mean standing vegetation cover per treatment. Cover was sampled in July 2023 and July 2024 before the annual treatment applications; therefore, the 2023 values represent vegetation responses 1 yr after the initial 2022 treatment, and the 2024 values represent vegetation responses 2 yr after the initial 2022 treatment and 1 yr after the 2023 treatment.
Together, the 2 yr show a clear pattern: treatments produced strong, immediate compositional differences in 2023, driven by direct removal of sensitive forbs and early shifts in competition. By 2024, many of these initial effects had diminished as plants recovered through resprouting, seedbank recruitment, and recolonization of disturbed patches. At the same time, dominant perennial grasses, both seeded natives and widespread introduced species, became more abundant across treatments, contributing to greater compositional similarity.
Treatment Effects on Plant Species Richness, Diversity, and Evenness
Establishing plant richness and diversity is a key objective in restoration, as species diversity contributes to ecological function and resilience (Palmer et al. Reference Palmer, Ambrose and Poff1997). In this study, treatments significantly affected species richness, diversity, evenness, and species group responses.
Overall Species Richness and Diversity
Plant species richness and diversity responded similarly to vegetation management in both years. In 2023, species richness (Hill q 0) differed significantly among treatments (Kruskal–Wallis χ2 = 31.49, df = 3, P < 0.001). Post hoc tests showed that richness was significantly lower in both Aminopyralid + metsulfuron-methyl and Combination treatments compared with Control and Mow (all P < 0.01), while Control and Mow did not differ (P = 0.608). Simpson diversity (Hill q 2) also varied significantly (ANOVA: F(3, 44) = 13.98, P < 0.001), with Aminopyralid + metsulfuron-methyl and Combination showing lower diversity than Control (P = 0.0012 and P = 0.0197, respectively), while Mow remained similar to Control (P = 0.354) and higher than both herbicide-based treatments (both P < 0.001). In 2024, richness again differed among treatments (ANOVA: F(3, 44) = 3.42, P = 0.025), showing the same ranking (Mow > Control > Aminopyralid + metsulfuron-methyl ≈ Combination), although only Mow vs. Combination was statistically significant (P = 0.040). Simpson diversity did not differ significantly in 2024 (F(3, 44) = 0.76, P = 0.525), but followed the same numerical pattern, with Mow highest and herbicide-based treatments lowest. Overall, both years demonstrated that herbicide-based vegetation management consistently reduced species richness and diversity, whereas mowing maintained or slightly enhanced plant diversity relative to untreated controls. This is consistent with studies showing that mowing can maintain or increase grassland plant diversity by reducing dominant species and increasing light at the soil surface (Collins et al. Reference Collins, Knapp, Briggs, Blair and Steinauer1998; Valko et al. Reference Valko, Torok, Matus and Tothmeresz2012). A long-term study also supports these patterns: in threatened temperate grasslands, annual mowing increased overall plant species richness for both native and exotic plants, while Shannon diversity responded positively for native species and neutrally for exotic species, resulting in a net increase in total diversity (Smith et al. Reference Smith, Barrett and Milner2018).
Plant community evenness (qE3) showed different treatment responses between years. In 2023, evenness did not differ significantly among treatments (Kruskal–Wallis: χ2 = 5.36, df = 3, P = 0.147). However, in 2024, evenness significantly differed among treatments (ANOVA: F(3, 44) = 3.17, P = 0.034). Mean evenness was highest in the Combination treatment (mean ± SE: 0.814 ± 0.040), followed by Aminopyralid + metsulfuron-methyl (0.730 ± 0.050), Control (0.673 ± 0.052), and Mow (0.619 ± 0.029). Tukey’s honest significant difference test indicated that the Combination treatment had significantly greater evenness than the Mow treatment (P = 0.028), while all other pairwise differences were not significant (P > 0.1). These results suggest that combining mowing with aminopyralid + metsulfuron-methyl application promotes more equitable species abundance distributions than mowing alone. Aminopyralid + metsulfuron-methyl application effectively reduced competitive dominant forb species, promoting evenness, while mowing did not. The high richness observed under the mowing treatment included many weedy or undesirable species (Landschoot et al. Reference Landschoot, Fidanza and Kister2022), which may not align with long-term restoration goals.
Seeded Species Responses
Seeded species responses showed no significant differences among treatments in either 2023 or 2024 (2023 richness: Kruskal–Wallis χ2 = 5.10, df = 3, P = 0.165; diversity: χ2 = 3.90, df = 3, P = 0.273; cover: χ2 = 5.30, df = 3, P = 0.151; Figure 2A; 2024 richness: χ2 = 4.13, df = 3, P = 0.247; diversity: χ2 = 3.01, df = 3, P = 0.390; cover: χ2 = 6.92, df = 3, P = 0.074; Figure 2B). Although treatment differences in seeded cover were not statistically supported, selective aminopyralid + metsulfuron-methyl suppressed competing vegetation, thereby reducing resource competition and increasing light availability to promote seeded native grass establishment. The target set for this project was to achieve at least 80% total plant cover of native species, most of which were seeded species. In this context, after 2 yr, the current levels of seeded species cover remained below the restoration goal. Despite some improvement under aminopyralid + metsulfuron-methyl–based treatments, seeded species establishment was still limited. The lowest seeded species cover, observed in the Control, highlighted that competition from noxious and introduced species in the seedbank or colonizing the plots inhibited the growth and recruitment of seeded species. Additionally, prior research underscored the importance of both seeding density and seed mix diversity, with high-diversity (35 species) mixes and a seeding rate of 1,366 pure live seed (PLS) m⁻2 shown to achieve the best restoration outcomes (Applestein et al. Reference Applestein, Bakker, Devlin and Hamman2018; Barr et al. Reference Barr, Jonas and Paschke2017). In this project, the seeding density was estimated to be approximately 2,940 PLS m⁻2 based on the known seed mix composition (Table 1) and applied seeding rate (30 kg ha−1). However, because only seven native grass species were included in the mix, the relatively low diversity may have limited overall establishment success despite the higher seeding density. Greater seeding diversity has been suggested to enhance plant community stability and establishment by increasing the likelihood that species can exploit variable conditions across heterogeneous environments (Feng et al. Reference Feng, Zhang, Zhao and Zhang2015). The low establishment of seeded species may also reflect environmental constraints, including limited soil moisture after seeding and precipitation timing, which can strongly influence first-year germination and survival (Denton et al. Reference Denton, Sheley and Hardegree2024; Groves et al. Reference Groves, Bauer and Brudvig2020). At the study site, annual precipitation was ∼327 mm in 2021, 356 mm in 2022, 410 mm in 2023, and 438 mm in 2024, which is below the regional long-term mean of ∼476 mm (Environment and Climate Change Canada 2024). Numerous factors may inhibit germination and seedling survival, including elevated soil temperatures; site preparation and residual competition that influence seedbed conditions; and soil nutrient limitations such as low available phosphorus and cation imbalances in calcium, iron, and manganese (Alstad et al. Reference Alstad, Damschen and Ladwig2018; Espeland et al. Reference Espeland, Hendrickson, Toledo, West and Rand2017; James et al. Reference James, Sheley, Leger, Adler, Hardegree, Gornish and Rinella2019; Newbold et al. Reference Newbold, Knapp and Pile2020). Finally, although our study did not manipulate seeding season, prior work in the Aspen Parkland reports greater seeded species establishment with spring seeding and short-lived mowing benefits accompanied by later reductions in bare ground (Pitchford et al. Reference Pitchford, Dhar and Naeth2024).
Individual Seeded Species Responses
No seeded grass species showed significant differences in cover among treatments in either 2023 or 2024 (Kruskal–Wallis tests; all P ≥ 0.13; Table 2). In 2023, Plains rough fescue [Festuca hallii (Vasey) Piper] (χ2 = 3.723, df = 3, P = 0.289), Idaho fescue (Festuca idahoensis Elmer) (χ2 = 2.044, df = 3, P = 0.563), Rocky Mountain fescue (Festuca saximontana Rydb.) (insufficient data for analysis), needle and thread grass [Hesperostipa comata (Trin. & Rupr.) Barkworth] (χ2 = 2.042, df = 3, P = 0.563), June grass [Koeleria macrantha (Ledeb.) Schult.] (χ2 = 2.043, df = 3, P = 0.563), and N. viridula (χ2 = 1.787, df = 3, P = 0.619) all showed no treatment effect. Western wheatgrass [Pascopyrum smithii (Rydb.) Á. Löve] exhibited a marginal trend (χ2 = 7.029, df = 3, P = 0.083), and was numerically greater in the mowing treatment, but this pattern did not reach statistical significance and was not present in 2024 (χ2 = 0.752, df = 3, P = 0.861). In 2024, F. hallii, F. idahoensis, F. saximontana, H. comata, K. macrantha, P. smithii, and N. viridula again all showed no significant differences among treatments (all P > 0.13). Across both years, seeded native grasses remained uniformly low in cover across all treatments. Although Aminopyralid + metsulfuron-methyl and the Combination treatment reduced competitive vegetation and increased bare ground (discussed later), these changes did not result in greater establishment of any seeded species relative to the Control. The uniformly low cover observed across all seeded grasses indicates that recruitment was likely limited by other environmental factors as discussed earlier or that more time is needed for individual plants to establish.
Noxious Weed Responses
The pattern of noxious weed responses to treatments in 2023 and 2024 was very similar: herbicide-based treatments (Aminopyralid + metsulfuron-methyl and Combination) consistently reduced noxious weed richness and cover relative to Control and Mow, while Simpson diversity showed little variation. Noxious weed species richness differed significantly among treatments in both years (2023: Kruskal–Wallis χ2 = 24.79, df = 3, P < 0.001; 2024: χ2 = 19.34, df = 3, P < 0.001; Table 2). In 2023, post hoc tests showed significant richness differences for Control vs. Aminopyralid + metsulfuron-methyl, Control vs. Combination, Aminopyralid + metsulfuron-methyl vs. Mow, and Mow vs. Combination (all P < 0.05). In 2024, richness in Aminopyralid + metsulfuron-methyl and Combination remained significantly lower than in Control (P = 0.029), while other pairwise contrasts were not significant. Noxious weed cover also varied significantly among treatments in both years (2023: Kruskal–Wallis χ2 = 25.51, df = 3, P < 0.001; Figure 2A; 2024: χ2 = 20.69, df = 3, P < 0.001; Figure 2B), with post hoc tests confirming that both herbicide-based treatments had significantly lower cover than Control and Mow in 2023 (P < 0.05) and significantly lower cover than both Control (P = 0.0318 and P = 0.0395) and Mow (P = 0.0066 and P = 0.0086) in 2024. Simpson diversity (Hill q 2) could not be meaningfully assessed in 2023 due to dominance by a single species (C. arvense) and did not differ significantly in 2024 (χ2 = 1.28, df = 3, P = 0.733; Table 2). These results indicate that the aminopyralid + metsulfuron-methyl program (ClearView®, 230 g product ha−1, applied in 2022 and 2023) was highly effective at suppressing the targeted noxious forbs in our plots, with some plots reaching near-zero values for both richness and cover in the year of application.
Mowing alone allowed persistence of all noxious weed species, C. arvense, L. vulgaris, and S. arvensis (Table 2). Although the suppression of aggressive noxious species such as C. arvense aligned with restoration goals, aminopyralid + metsulfuron-methyl–based treatments also resulted in considerable reductions in total vegetation cover and litter accumulation (Table 2; Figure 1A and 1B). This led to increased bare-ground exposure, which may have increased erosion risk, disrupted nutrient cycling, and degraded soil quality (Edwards and Pimentel Reference Edwards and Pimentel1989; Facelli and Pickett Reference Facelli and Pickett1991; Lajtha et al. Reference Lajtha, Bowden and Nadelhoffer2014; Rose et al. Reference Rose, Cavagnaro, Scanlan, Rose, Vancov, Kimber and Van Zwieten2016). Management strategies should therefore identify ways to maintain ground cover, for example, by combining herbicide application with mulch or rapid revegetation strategies (Feng et al. Reference Feng, Zhang, Zhao and Zhang2015; Marble Reference Marble2015). Additionally, precision herbicide application methods, such as spot/patch spraying with lower herbicide rates or shielded application, may reduce nontarget impacts and maintain protective ground cover (Smith et al. Reference Smith, Kanjithanda, Hayashi, French and Milner2023). Future efforts may also benefit from integrated approaches, such as pairing targeted herbicide with selective clipping removal to enhance native plant establishment while mitigating negative ecosystem effects.
Native Plant Species Responses
Only two native shrub species, Woods’ rose (Rosa woodsii Lindl.) and western snowberry (Symphoricarpos occidentalis Hook.), were recorded at very low abundance, which prevented meaningful calculation of Simpson diversity (Hill q 2) in both years due to zero-inflation, identical values, or insufficient variation. In 2023, native shrub richness (Kruskal–Wallis χ2 = 6.41, df = 3, P = 0.093) and cover (Kruskal–Wallis χ2 = 6.67, df = 3, P = 0.083) did not differ significantly among treatments, indicating no detectable response to vegetation management. In 2024, however, native shrub cover differed significantly among treatments (Kruskal–Wallis χ2 = 14.47, df = 3, P = 0.0023). Post hoc Nemenyi comparisons showed that shrubs were completely absent in both the Aminopyralid + metsulfuron-methyl and Combination treatments, resulting in significantly lower cover than the Control (P = 0.025), while Mow did not differ from Control (P = 0.851). The stronger treatment effects observed in 2024 likely reflect an additive impact of the repeat Aminopyralid + metsulfuron-methyl application, which further suppressed R. woodsii and S. occidentalis beyond what was detectable in the first year of treatment. Aminopyralid and metsulfuron have been used to control S. occidentalis by up to 97% (Sbatella et al. Reference Sbatella, Wilson and Sleugh2011) and Rosaspp. by up to 86% (Enloe and Lauer Reference Enloe and Lauer2016). This represented a potential downside to the use of herbicides in restoration efforts, as native shrubs could contribute to the restoration of vegetation structure and provide important ecosystem services, such as support for pollinators (Gilgert and Vaughan Reference Gilgert and Vaughan2011; Ruiz-Jaen and Aide Reference Ruiz-Jaen and Aide2005).
The pattern of native species response to vegetation management in 2023 and 2024 was identical, with no significant treatment effects on richness, Simpson diversity, or cover in either year (2023 richness: Kruskal–Wallis χ2 = 4.29, df = 3, P = 0.231; diversity: χ2 = 1.85, df = 3, P = 0.605; cover: χ2 = 7.39, df = 3, P = 0.060, Figure 2A; 2024 richness: χ2 = 4.07, df = 3, P = 0.254; diversity: χ2 = 2.30, df = 3, P = 0.512; cover: χ2 = 3.06, df = 3, P = 0.383, Figure 2B). Although the native species showed no treatment effects, individual native species exhibited year-to-year variation in cover. For example, E. trachycaulus had its highest cover in the Control treatment in 2023, whereas in 2024 its highest cover occurred in the Aminopyralid + metsulfuron-methyl treatment (Table 2). Although E. trachycaulus was not included in the seeded mix, its presence in the commercial lot (≈33 seeds 25 g−1) indicates a low-level, non-target native contaminant that subsequently established and is reported here as an unintended but ecologically desirable native recruit. This species is well known for its ability to establish rapidly following disturbance and to expand aggressively in newly opened microsites (Tilley et al. Reference Tilley, Ogle, St John, Holzworth, Crowder and Majerus2006).
Introduced Species Responses
Introduced species showed consistent responses to vegetation management across both years, with Aminopyralid + metsulfuron-methyl–based treatments consistently reducing their abundance. This was indicated by significant treatment effects on introduced richness, diversity, and cover in both years (2023 richness: Kruskal–Wallis χ2 = 33.26, df = 3, P < 0.001; 2023 Simpson diversity: χ2 = 24.16, df = 3, P < 0.001; 2023 cover: χ2 = 28.03, df = 3, P = 3.58 × 10⁻6; 2024 richness: ANOVA F(3, 44) = 7.03, P < 0.001; 2024 Simpson diversity: χ2 = 1.28, df = 3, P = 0.733; 2024 cover: ANOVA F(3, 44) = 20.47, P < 0.001; Figures 2 and 3). Post hoc Nemenyi and Tukey tests showed that introduced richness and cover were significantly lower in the Aminopyralid + metsulfuron-methyl and Combination treatments compared with Control and Mow (all P < 0.01), while Mow did not differ from Control in 2024 (P = 0.95). This indicates that mowing alone was less effective than herbicide-based treatments at suppressing introduced species. Although aminopyralid and metsulfuron were primarily used for broadleaf weed control, this combination has also demonstrated high efficacy in controlling certain annual invasive grass species, achieving 68% to 88% control (Flessner et al. Reference Flessner, Lassiter and Bamber2019). However, in our study, differential tolerance among introduced grasses was observed: perennial grasses such as B. inermis were effectively suppressed under aminopyralid + metsulfuron-methyl treatments, while some annual grasses, such as wild oat (Avena fatua L.), persisted. Avena fatua occurred under the Aminopyralid + metsulfuron-methyl (1.67% cover), Mow (2.50%), and Combination (2.92%) treatments but was absent in the Control, suggesting it exploited newly opened niches created by aminopyralid + metsulfuron-methyl application or mowing disturbance (Table 2).
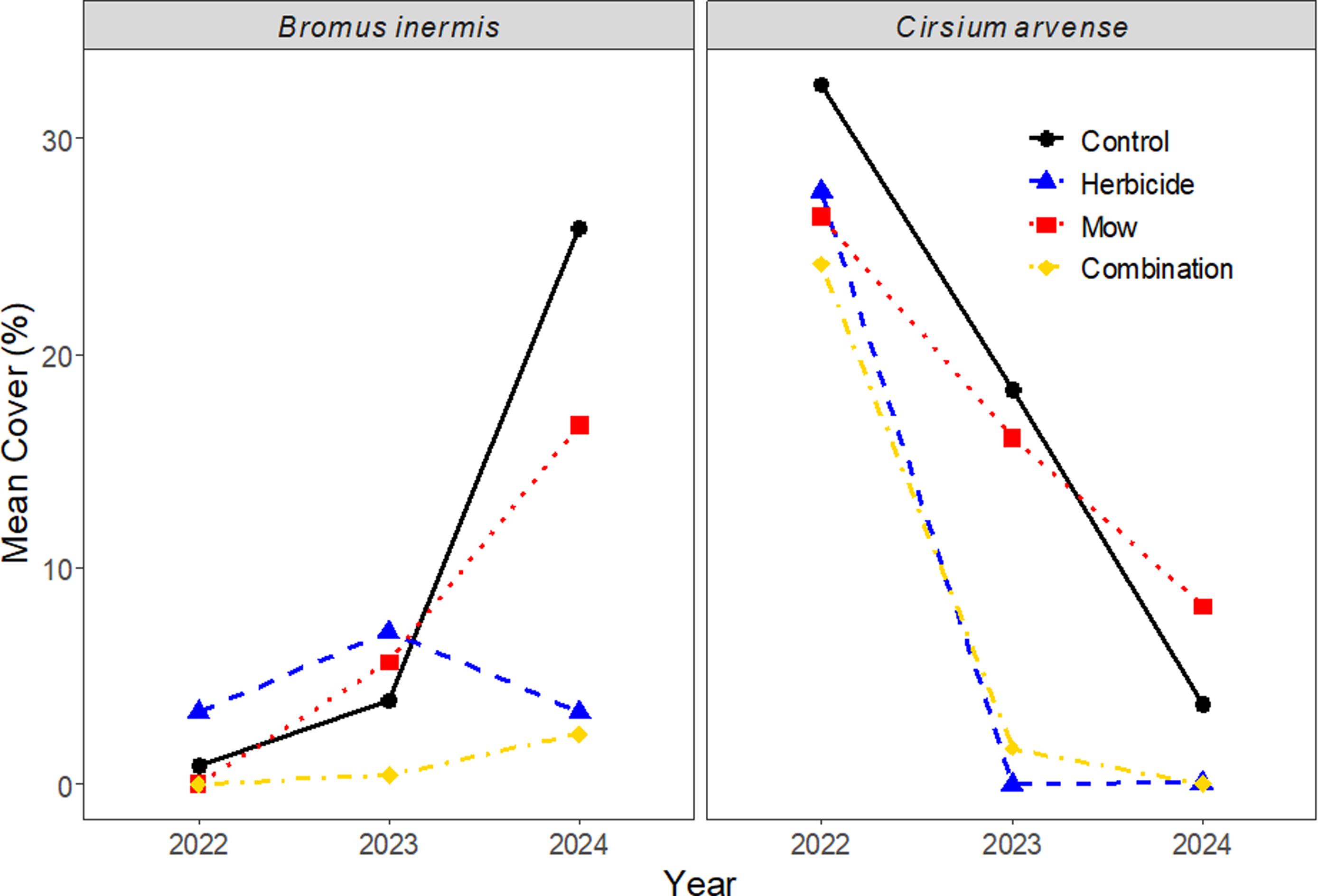
Mean predicted cover of Bromus inermis and Cirsium arvense across 3 yr (2022–2024) under four vegetation-management treatments in Edmonton, Alberta, Canada: Control (no treatment), Herbicide (aminopyralid + metsulfuron-methyl), Mow, and Combination (mowing + herbicide). Values represent model-estimated means from zero-inflated negative binomial mixed-effects models with plot as a random effect.

In contrast, mowing retained many introduced grasses and forbs, which increased both cover and species richness relative to the other treatments. This pattern was consistent with findings from other studies, particularly those that employed repeated or long-term mowing regimes (Abu-Dieyeh and Watson Reference Abu-Dieyeh and Watson2005; Landschoot et al. Reference Landschoot, Fidanza and Kister2022; Maron and Jefferies Reference Maron and Jefferies2001). Quackgrass [Elymus repens (L.) Gould] cover was higher under Aminopyralid + metsulfuron-methyl (7.50%) and Combination (7.50%) treatments compared with the Control (3.58%) and Mow (2.67%). This pattern likely reflects herbicide selectivity; as an acetolactate synthase inhibitor, metsulfuron provides poor control of E. repens (1%) but effectively suppresses broadleaf weeds such as C. arvense (86%) (Sprague et al. Reference Sprague, Frasier and Penner1999). Consequently, E. repens persistence or reestablishment following herbicide application is expected. Dandelion (Taraxacum officinale F.H. Wigg.) and red clover (Trifolium pratense L.), which are disturbance adapted and have rapid regrowth, had high cover under Mow (9.00% and 13.92%, respectively) but were nearly absent under Aminopyralid + metsulfuron-methyl and Combination treatments.
Year-to-Year Dynamics of Bromus inermis and Cirsium arvense Responses
Bromus inermis cover responded to vegetation management over time. Zero-inflated negative binomial models indicated significant effects (treatment: χ2 = 53.30, df = 7, P < 0.001; year: χ2 = 67.91, df = 6, P < 0.001; treatment by year: χ2 = 38.62, df = 6, P < 0.001; Figure 3). In 2022, because treatments had not yet been applied, the B. inermis cover values recorded that year reflect natural pretreatment variation, with low cover likely created by site preparation. Treatment effects became clear after management was implemented: in 2023, B. inermis cover increased in the Control, Aminopyralid + metsulfuron-methyl, and Mow treatments but remained strongly suppressed in the Combination treatment (0.41%), and pairwise contrasts showed significantly lower cover in the Combination treatment relative to Control (P = 0.032). By 2024, B. inermis cover increased substantially in Control and Mow plots while remaining low under Aminopyralid plus metsulfuron-methyl and the Combination treatment, with Tukey-adjusted comparisons indicating that the Combination treatment had 99% lower cover than Control (P = 0.0321) and 97% lower than Mow (P = 0.029). Together, these results demonstrate that Aminopyralid + metsulfuron-methyl and the Combination treatment consistently suppressed B. inermis, whereas untreated and mowed plots experienced marked increases in cover following treatment application.
Cirsium arvense showed declines in cover over time, in that zero-inflated negative binomial models indicated significant effects (treatment: χ2 = 32.63, df = 7, P < 0.001; year: χ2 = 109.77, df = 6, P < 0.001; treatment by year: χ2 = 26.93, df = 6, P < 0.001). In 2022, before Aminopyralid + metsulfuron-methyl and Mow treatments were applied, C. arvense cover was uniformly high across plots, reflecting natural pretreatment variation. In 2023, C. arvense cover remained high in Control and Mow plots but was nearly eliminated in plots with Aminopyralid + metsulfuron-methyl and Combination treatments, with pairwise contrasts indicating significantly lower cover than Control (P = 0.0179 and P = 0.0211) and Mow (P = 0.0035 and P = 0.0343), respectively. By 2024, C. arvense was almost completely absent in both Aminopyralid + metsulfuron-methyl and Combination plots, while persisting but lower in Control (4.35%) and Mow (8.11%). Pairwise contrasts indicated significantly lower cover in Aminopyralid + metsulfuron-methyl (P = 0.0179 vs. Control; P = 0.0035 vs. Mow) and Combination (P = 0.021 vs. Control; P = 0.006 vs. Mow). Overall, these results demonstrate that Aminopyralid + metsulfuron-methyl, applied alone or paired with mowing, was highly effective at suppressing C. arvense, whereas Control and Mow plots maintained substantial cover through 2023 and 2024.
The contrasting temporal responses of B. inermis and C. arvense demonstrate the necessity of species-specific strategies and sustained management in grassland restoration. Bromus inermis, a deeply rhizomatous C3 grass, exhibited rapid regrowth under annual mowing and untreated conditions, indicating strong vegetative resilience and the ability to exploit open canopy space once disturbance ceased. Similar recovery patterns have been reported elsewhere, where B. inermis persistence was attributed to extensive belowground bud banks and rapid clonal spread following defoliation (Dyer et al. Reference Dyer, Turner and Seastedt1991). In contrast, C. arvense showed a sharp and sustained decline following the initial treatment year, with cover reduced by more than 80% across all treatments. This pattern aligns with field studies reporting that aminopyralid or metsulfuron suppresses clonal thistles (Cirsiumspp.) when followed by seeding of competitive vegetation (Enloe et al. Reference Enloe, Lym, Wilson, Westra, Nissen, Beck and Halstvedt2007). The timing and rate of herbicide application are essential for effective control of C. arvense. Studies show that the most effective periods are the bud to early-bloom stage in late spring and the fall regrowth period when plants form basal rosettes, with fall treatments often providing the strongest and most persistent suppression (Hunter Reference Hunter1996; Beck and Sebastian Reference Beck and Sebastian2000; Government of Saskatchewan n.d.; Jacobs et al., Reference Jacobs, Sciegienka and Menalled2006). These windows coincide with growth stages in which carbohydrates are actively moving downward into the root system. Because systemic herbicides follow the same translocation pathways as photosynthesis, products such as the aminopyralid + metsulfuron mixture used in our study are carried more efficiently into the plant’s extensive creeping roots at these times. This improves root injury and reduces the likelihood of subsequent shoot emergence.
Collectively, these results underscore that management efficacy depends on life-history traits: rhizomatous grasses like B. inermis require high-frequency repeated or integrated control to limit belowground regeneration, whereas clonal forbs such as C. arvense can be effectively suppressed through short-term chemical intervention when followed by competitive plant establishment.
Our findings demonstrate the ecological trade-offs of aminopyralid + metsulfuron-methyl use in Aspen Parkland restoration. Treatments with aminopyralid + metsulfuron-methyl effectively suppressed noxious weeds and supported native species recovery but also reduced structural diversity by eliminating shrubs. Selective application approaches, combined with posttreatment reseeding or protective measures for shrubs, could have helped balance these outcomes. Including species with early-establishing or disturbance-tolerant traits could have further improved establishment success.
Acknowledgments
We thank Sam Bennett, Ping Nixon-Hermansen, L. Shea Giesbrecht, Batbaatar Amgaa, Danielle Perri, Kelly Wu, Rae-Anne Martin, and Sarah Johnson for their help in the field; Gary Braithwaite for arranging herbicide treatments; and Joshua Mcilravey, Nathaniel Papay, and Enoch Ruey for facilitating site access.
Funding
We are grateful for financial support from MITACS, EPCOR Water Services Inc., the Alberta Conservation Association, and Future Energy Systems.
Competing interests
The authors declare no conflicts of interest.

 Open access
Open access