


Introduction
Applying radiocarbon analysis in forensic investigations such as paper document age estimates or the determination of the amount of biogenic carbon in plastic polymers (e.g., Hüls et al. 2017; Telloli et al. Reference Telloli, Rizzo, Canducci and Bartolomei2019), rely on an up-to-date reference data set of 14C data over the recent decades. Traditionally, tree-ring- and atmospheric-14C data such as the recent compilation of Hua et al. (Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022) are used in these investigations, and here we propose to add and use 14C-data from archived seasonal growing plant tissues which should provide valuable isotopic information linked to atmospheric carbon isotopic composition. Seasonal growing plant tissue such as crop seeds contain carbon metabolized in a short-term period and also allow an extension into the future by adding measurements from recent harvests.
The Leibniz Institute of Plant Genetics and Crop Plant Research (IPK) in Gatersleben in Germany hosts an extensive collection of plant species and crops, primarily of cereals and other crops, from 1946 until today. Radiocarbon measurements of seasonal wheat seeds with alternating winter and spring growth habits, cultivated from 1946 to 2020, have been reported previously (Hüls et al. Reference Hüls, Börner and Hamann2021). Recently, additional 14C studies have utilized herbarium collections (Carbone et al. Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023) and crop archives (Dunbar et al. Reference Dunbar, Scott and Tripney2024), covering annual data from 1910 to 2021 and three-year intervals from 1852 to 2020, respectively.
This study extends the IPK Gatersleben radiocarbon dataset of wheat from 2020 to 2025 and complements this data set by adding measurements from soybean (Glycine max (L.) Merr.), which has been cultivated under comparable conditions to wheat at the IPK experimental fields since 1946. Soybeans have a different growth period than wheat, offering complementary radiocarbon data. Additionally, δ1 3C composition of wheat was determined for comparison with other biogenic and atmospheric carbon isotope compositions (14C, 13C).
Material and methods
Material
The IPK collection (see Figure 1) comprises 150,000 accessions from 3212 plant species and 776 genera, including wild and primitive forms, landraces, and both historical and modern cultivars, primarily of cereals but also other crops (Börner and Khlestkina Reference Börner and Khlestkina2019; Börner et al. Reference Börner, Khlestkina, Chebotar, Nagel, Arif, Neumann, Kobiljski, Lohwasser and Röder2012). The institute and its agricultural plots are located east of the Harz mountains in an area with average annual air temperature of 9 °C at 2 m height and average annual precipitation of 41 mm (source: https://wetter.ipk-gatersleben.de/apex/f?p=182:10:6480178410756 with calculated annual mean from monthly data between 1953 and 2026).
Location of the IPK (tagged with a flag) in Germany.

Figure 1. Long description
A map of Germany highlighting key cities and the location of the IPK. The map includes major cities such as Kiel, Hamburg, Hannover, Berlin, and Frankfurt. The IPK is marked with a flag symbol. The map also shows the surrounding regions of Europe in a lighter shade, providing geographical context. A compass rose indicating north is present in the bottom right corner, and a scale bar showing 100 kilometers is included for distance reference.
Wheat (Triticum aestivum L.) and soybean (Glycine max (L.) Merr.) have been grown since 1946 in the IPK experimental fields (2 m2 plots) in Gatersleben (latitude 51°49'19.74''N, longitude 11°17'11.80''E, 110.5 m a.s.l.). Using standard agronomic practices, seasonal seed harvests from these long-term cultivation plots were used for radiocarbon analyses.
Wheat samples consist of winter and spring varieties. Winter wheat was sown in October and harvested in the beginning of July, incorporating atmospheric carbon metabolized from April to June. Spring wheat was sown in March or April (depending on seasonal weather conditions) and harvested at the beginning of August and thus should reflect carbon fixation from May to July. The dates mid-May, i.e., May 15th (15-5-1946/2025) and mid-June, i.e., June 15th (15-6-1946/2025) for winter- and spring wheat variants, respectively, are used as reference (mid) points defining the carbon fixation periods mirrored in the plant tissue radiocarbon content.
Likewise, soybeans, which are sown around mid-May, flowered by late June or early July. The plants and seeds developed from July to September, metabolizing atmospheric CO2 during this period, with the date August 15 serving as the reference point. Soybean is harvested in late September or early October.
Wheat and soybeans were stored in glass tubes under controlled conditions, maintained at 22.5 °C ± 3 °C and a relative humidity of 45–50%.
Methods
Single wheat grain and soybean were washed with acetone for 30 min in an ultrasonic bath, decanted, and dried subsequently at 60 °C in an oven overnight. Afterwards, prepared material was frozen in liquid nitrogen and powdered, i.e., homogenized, using pestle and mortar. Between 6 and 7 mg of powdered material was flame-sealed together with precombusted cupric oxide (™ Merck, CuO with a core of Cu2O, about 99.986% purity) and silver capsules (99.99% purity) in quartz-ampoules and combusted at 900 °C. CO2 was cryogenically purified and graphitized with catalytic Fe and H2 at 600 °C for 14C analysis. Unpublished monitoring of graphitization parameters (pressure readings of reduction components CO2 and H2) by MH indicate an overall reduction efficiency of 99 ± 0.05% (N=1518).
Radiocarbon measurements were conducted at the Leibniz-Labor with a HVEE 3MV Tandetron 4130 accelerator mass spectrometer (AMS). Simultaneously measured 14C/12C and 13C/12C are compared to Oxalic Acid II, and 14C is normalized to δ13C of –25‰ (Stuiver and Polach 1977). Blank correction is based on modelled 14C distribution vs. sample size of Alfa graphite (Nadeau et al. Reference Nadeau, Grootes, Schleicher, Hasselberg, Rieck and Bitterling1998; Nadeau and Grootes Reference Nadeau and Grootes2013). Radiocarbon concentration is given in F14C (fraction modern, Reimer et al. Reference Reimer, Brown and Reimer2004) and Δ14C by correcting for the decay of the measurement standard and samples (Stenström et al. Reference Stenström, Skog, Georgiadou, Genberg and Johansson2011; Stuiver and Polach 1977) (
 ${\Delta ^{14}}C = (({F^{14}}C*{e^{\lambda \left( {1950 - x} \right)}}) - 1)*1000$
;
${\Delta ^{14}}C = (({F^{14}}C*{e^{\lambda \left( {1950 - x} \right)}}) - 1)*1000$
;
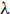 $\lambda $
=1/8267, x=year of seed growth). Reported 14C uncertainty takes into account the uncertainty of 14C/12C, uncertainty of fractionation correction, and uncertainty of applied blank correction.
$\lambda $
=1/8267, x=year of seed growth). Reported 14C uncertainty takes into account the uncertainty of 14C/12C, uncertainty of fractionation correction, and uncertainty of applied blank correction.
Two subsamples of seasonal wheat and soy seeds were measured in different batches. For soy, duplicate samples of the seasons 1946–1967 and 2000–2025 were analyzed. Variance-weighted means were calculated using
 $\bar x = {{\sum\limits_{i = 1}^n {{{{x_i}} \over {\sigma _i^2}}} } \over {\sum\limits_{i = 1}^n {{1 \over {\sigma _i^2}}} }}\;{\rm{and}}\;\sigma _{\bar x}^2 = {1 \over {\sum\limits_{i = 1}^n {{1 \over {\sigma _i^2}}} }}$
$\bar x = {{\sum\limits_{i = 1}^n {{{{x_i}} \over {\sigma _i^2}}} } \over {\sum\limits_{i = 1}^n {{1 \over {\sigma _i^2}}} }}\;{\rm{and}}\;\sigma _{\bar x}^2 = {1 \over {\sum\limits_{i = 1}^n {{1 \over {\sigma _i^2}}} }}$
with
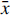 $\bar x$
being weighted mean F14C or Δ14C, and
$\bar x$
being weighted mean F14C or Δ14C, and
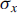 ${\sigma _x}$
being the error of the weighted mean (Ward and Wilson Reference Ward and Wilson1978). When replicate measurements were significantly different from each other (i.e., differences > 2σ), additional third measurements were conducted to improve the agreement of all individual measurements or for detecting outliers.
${\sigma _x}$
being the error of the weighted mean (Ward and Wilson Reference Ward and Wilson1978). When replicate measurements were significantly different from each other (i.e., differences > 2σ), additional third measurements were conducted to improve the agreement of all individual measurements or for detecting outliers.
Between 175 and 225 µg of individual wheat and soy material was sealed in tin capsules for IRMS δ1 3C analysis. Samples were analyzed in duplicate, and for every 6th sample two different reference materials for isotopic normalization were run. A multi-point calibration procedure was applied for δ1 3C calculations (Coplen et al. Reference Coplen, Brand, Gehre, Gröning, Meijer, Toman and Verkouteren2006). δ1 3C values are normalized using reference material, e.g., IAEA-CH3 cellulose, IAEA-601 benzoic acid, and IAEA-CH6 sucrose. δ1 3C results are reported in ‰ on the VPDB scale (Coplen et al. Reference Coplen, Brand, Gehre, Gröning, Meijer, Toman and Verkouteren2006).
A Flash IRMS IsoLink elemental analyzer (Thermo Fisher Scientific Inc.) was employed for sample combustion and gas purification. Combusted gases were carried by high-purity helium, and CO2 was purified in the elemental analyzer and separated chromatographically from other combustion gases using a packed GC column. The elemental analyzer was interfaced via a ConFlo IV open-split to a DELTA™ Q isotope ratio mass spectrometer (Thermo Fisher Scientific Inc.).
Discrimination against 13C (Δ13C) or 14C (ΔΔ14C) with respect to their carbon source (e.g., atmospheric CO2) can be calculated based on the following formula:
 ${\Delta ^{13}}{\rm{C}}\; = \;{{{{{\delta ^{13}}{{\rm{C}}_{{\rm{source}}}}} \over {1000}}\; - \;{{{\delta ^{13}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}} \over {1 + \;{{{\delta ^{13}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}}}{\rm{*}}1000\;\left( {{\rm{after}}\;{\rm{Farquhar}}\;{\rm{et}}\;{\rm{al}}.\;1989} \right)$
${\Delta ^{13}}{\rm{C}}\; = \;{{{{{\delta ^{13}}{{\rm{C}}_{{\rm{source}}}}} \over {1000}}\; - \;{{{\delta ^{13}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}} \over {1 + \;{{{\delta ^{13}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}}}{\rm{*}}1000\;\left( {{\rm{after}}\;{\rm{Farquhar}}\;{\rm{et}}\;{\rm{al}}.\;1989} \right)$
 $\Delta {\Delta ^{14}}{{C}}\; = \;{{{{{\Delta ^{14}}{{\rm{C}}_{{\rm{source}}}}} \over {1000}}\; - \;{{{\Delta ^{14}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}} \over {1 + \;{{{\Delta ^{14}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}}}{\rm{*}}1000\quad \;\left( {{\rm{after}}\;{\rm{Farquhar}}\;{\rm{et}}\;{\rm{al}}.\;1989} \right)$
$\Delta {\Delta ^{14}}{{C}}\; = \;{{{{{\Delta ^{14}}{{\rm{C}}_{{\rm{source}}}}} \over {1000}}\; - \;{{{\Delta ^{14}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}} \over {1 + \;{{{\Delta ^{14}}{{\rm{C}}_{{\rm{plant}}}}} \over {1000}}}}{\rm{*}}1000\quad \;\left( {{\rm{after}}\;{\rm{Farquhar}}\;{\rm{et}}\;{\rm{al}}.\;1989} \right)$
In contrast to δ13C and Δ14C, Δ13C and ΔΔ14C allow the investigation into biological processes (Farquhar and Lloyd 1993). For calculation, δ13Catmosphere was taken from Graven et al. (Reference Graven, Allison, Etheridge, Hammer, Keeling, Levin, Meijer, Rubino, Tans, Trudinger, Vaughn and White2017) and Δ14Catmosphere (i.e., Δ14Csource) from NHZ1 summer (Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022).
Results
Development of wheat and soybean radiocarbon composition (F14C) during the bomb-spike
Gatersleben plant 14C (see Figure 2 and Table 1 in supplement) lies below the modern standard concentration of 1 F14C before 1955 and increase subsequently towards a first maxima in 1959 (ca.1.6 F14C), resulting from the increasing number (and energy) of atmospheric nuclear tests (UNSCEAR 2000; Bergkvist and Ferm Reference Bergkvist and Ferm2000; Prăvălie Reference Prăvălie2014). Atmospheric nuclear test activity paused in 1959, and only 3 tests with low energy yield in 1960 caused a first 14Catm reduction (ca.<1.24 F14C in 1960 and 1961, respectively). Resuming nuclear tests in the 2nd half of 1961 again caused increased 14Catm concentrations as documented by 1962 spring/early summer biomass 14C concentration. Evolution of 14C concentrations in seeds during the early phase of the bomb-spike compare reasonably well to the Trondheim pine tree-ring tissue measurements of the same period (Hüls et al. Reference Hüls, Börner and Hamann2021; Svarva et al. Reference Svarva, Grootes, Seiler, Stene, Thun, Vaernes and Nadeau2019).
Radiocarbon concentration in Gatersleben plant tissue (Triticum aestivum L., wheat; Glycine max (L.) Merr., soybean), grown during 1946–2025. Dashed line corresponds to monthly 14C of the NHZ1 compilation (Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022).

Figure 2. Long description
The line graph presents radiocarbon concentration in plant tissue from 1946 to 2025. The x-axis represents the year of growth, ranging from 1940 to 2030, while the y-axis indicates the F 14 C values, ranging from 0.90 to 1.90. The graph includes three data lines: a dashed line for the monthly NHZ1 compilation, green triangles for Triticum aestivum L. (wheat), and purple circles for Glycine max (L.) Merr. (soybean). The data shows a significant increase in radiocarbon concentration around the mid-1960s, peaking at approximately 1.90 F 14 C, followed by a gradual decline to around 1.00 F 14 C by 2025. The wheat and soybean data closely follow the NHZ1 compilation trend. All values are approximated.
Maximum nuclear tests in late 1962 caused increased and maximum atmospheric 14C in 1963 as seen in the Northern Hemisphere 1 compilation of Δ14C: 945‰, 1.9478 F14C (Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022) and Gatersleben biomass 14C by late summer 1963 (Δ14C 951‰, 1.9543 F14C). The Limited Test Ban Treaty (LTBT or partial test ban treaty) was signed August 5, 1963.
Excess atmospheric 14C concentration decreased as atmospheric 14CO2 was sequestered in the natural sinks such as the ocean and the biosphere. Recorded Gatersleben plant 14C decreased rapidly by about 76‰/yr until 1968 (Δ14C: 550–570‰, 1.55–1.57 F14C; see Figure 2). Biomass 14C decreased between 1970 and 1977 by ca. 28 ‰/yr. Between 1977 and 2009, annual biomass—(and from that deduced atmospheric—) 14C reduction seem to follow an exponential function with a “time-constant” of T1/2 ≈ 11yr.
Gatersleben plant 14C reached the modern reference level of 1 in 2019 and lies below the reference level in 2025 (Figure 2, F14Cwheat= 0.9924 ± 0.0023, F14Csoy= 0.9959 ± 0.0024, Δ14Cwheat = –16.57 ± 2.28, Δ14Csoy = –13.10 ± 2.45‰). Recently, more plant-tissue 14C timeseries using samples from herbarium collections and seed vaults were published. Carbone et al. (Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023) used several different annual plant specimens grown on the Southern Colorado Plateau around Flagstaff, Arizona, USA, collected in the Deaver Herbarium at the Northern University of Arizona in Flagstaff, for the period 1910–2015, and complemented by harvesting living plants between 2015 and 2021. Dunbar et al. (Reference Dunbar, Scott and Tripney2024) analyzed carbon isotopes (13C & 14C) in single barley grains (Hordeum vulgare L.), collected from the archive of the Long-Term Experiments (LTEs) Hoosfield Spring Barley at Rothamsted Research (Harpenden, Hertfordshire, UK), representing the growth season March-September between 1852 and 2020 and a temporal resolution of 3 years. The Colorado Plateau lies in the Northern Hemisphere Zone 2, while Rothamsted and Gatersleben are located within Northern Hemisphere Zone 1 (NHZ1, NHZ2; Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022).
A comparison of Gatersleben, Colorado Plateau, and Rothamsted plant 14C is given in Figure 3 in four time slices (A: 1945–1955, B: 1955–1970, C: 1970–2000, and D: 2000–2025). Pre-bomb plant-14C for all three series are below the monthly NHZ1 and II reference data. After 1955, measured 14C concentrations in all plant tissues start to increase above the modern reference level around 1955–1956, with peak concentrations measured in Rothamsted barley and Gatersleben seeds in mid-1963. Colorado Plateau plant 14C peaked in 1964. A delayed rise of plant 14C pre-1964 compared to NHZ2 was noted by Carbone et al. (Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023), not observed in Gaterseleben or Rothamsted seeds compared to NH1 data.
Overview and comparison of 14C concentrations measured in plant tissues from Gatersleben/Germany, Rothamsted/UK (Dunbar et al. Reference Dunbar, Scott and Tripney2024), and the Colorado Plateau/USA (Carbone et al. Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023), in time-slices from 1940 to 2025, in comparison to the monthly 14C of NHZ 1 and 2 compilations (Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022). (A) period between 1946 to 1955, (B) period between 1955 to 1970, (C) period between 1970 to 2000, (D) period between 2000 and today.

Figure 3. Long description
The image contains four line graphs labeled A, B, C, and D, each representing different time periods from 1940 to 2025. The graphs compare 14C concentrations measured in plant tissues from Gatersleben/Germany, Rothamsted/UK, and the Colorado Plateau/USA. Graph A covers the period between 1946 to 1955, Graph B from 1955 to 1970, Graph C from 1970 to 2000, and Graph D from 2000 to the present. Each graph includes data points for Rothamsted barley, Colorado Plateau plants, Gatersleben wheat, and Gatersleben soil, along with monthly NHZ1 and NHZ2 compilations. The x-axis represents the year of growth, and the y-axis represents the 14C concentration. The graphs show trends and variations in 14C levels over the specified periods, with different symbols and colors representing the various data sets. All values are approximated.
After the LTBT in 1963, atmospheric and biomass 14C decreased. Gatersleben and Colorado Plateau plants 14C follow NHZ1&2 concentrations closely. Between 1966 and 1983, the Rothamsted barley record give elevated 14C concentrations with respect to NHZ1, indicating enriched atmospheric 14CO2 from the weapons testing reaching this locality as speculated by Dunbar et al. (Reference Dunbar, Scott and Tripney2024). With eastern flowing airmasses, passing western Europe’s industrialized regions, atmospheric CO2 reaching Gatersleben may get depleted by fossil, 14C-free emissions, which would explain lower Gatersleben seed 14C concentrations. Elevated 14C concentrations in Rothamsted barley are also observed between 2005 and 2016 compared to NHZ1, while Gatersleben seeds give depleted 14C during 2005–2010 and 2017–2019. Carbone et al. (Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023) speculated about an influence of increased wildfires to be responsible for elevated Colorado Plateau plants 14C in 2015–2021.
Evolution of wheat stable carbon isotope composition (δ13C) over the bomb-spike
Stable carbon isotopes were analyzed on Gatersleben wheat seeds from the 1946–2022 seasons and compared to barley seeds δ13C from Rothamsted (Dunbar et al. Reference Dunbar, Scott and Tripney2024; see Figure 4A). The comparison between wheat and barley is not intended to be a detailed examination of 13C composition between species because of physiological differences and different growth conditions due to climate and soil composition, although both plants follow the C3 photosynthetic pathway. However, it is believed to be useful for a generalized view of the stable carbon isotope evolution in the environment during the recent past.
(A) δ13C of Gatersleben wheat (Triticum aestivum) and δ13C of Rothamsted barley (Hordeum vulgare). (B) Global atmospheric δ13C compiled by Graven et al. Reference Graven, Allison, Etheridge, Hammer, Keeling, Levin, Meijer, Rubino, Tans, Trudinger, Vaughn and White2017. (C) 14C discrimination ΔΔ14C (after Farquhar et al. Reference Farquhar, Ehleringer and Hubic1989) of Gatersleben wheat using summer NHZ1 14C data as source. (D) 13C discrimination Δ13C (after Farquhar et al. Reference Farquhar, Ehleringer and Hubic1989) of Gatersleben wheat using global δ13C compiled by Graven et al. (Reference Graven, Allison, Etheridge, Hammer, Keeling, Levin, Meijer, Rubino, Tans, Trudinger, Vaughn and White2017) as source. (E) and (F) local average April-July precipitation and temperature at 2 m ground level from IPK Gatersleben weather station (https://wetter.ipk-gatersleben.de/).

Figure 4. Long description
The image contains six graphs. The first graph shows the carbon isotope ratio delta 13C of Gatersleben wheat and Rothamsted barley over time. The second graph displays the global atmospheric delta 13C compiled by Graven et al. 2017. The third graph presents the 14C discrimination delta delta 14C of Gatersleben wheat using summer NHZ1 14C data as a source. The fourth graph illustrates the 13C discrimination delta 13C of Gatersleben wheat using global delta 13C compiled by Graven et al. 2017 as a source. The fifth graph shows local average April-July precipitation, and the sixth graph depicts local average April-July temperature at 2 meters ground level from the IPK Gatersleben weather station. All values are approximated.
Gatersleben wheat show a decrease in 13C concentrations, from between –24‰ and –25‰ before 1950 to between –26‰ and –27‰ in 2023, caused by the ongoing fossil carbon emission (e.g., Suess Reference Suess1955) with a generally lighter isotopic composition. The general decreasing trend of about 2‰/century as depicted from wheat 13C (see Figure 4) during 1946–1990 apparently increased from the early 1990s onwards to about 15‰/century. After 2015, measured δ13Cwheat concentrations appear constant at ca. –27‰ to –28‰. Rothamsted barley seed give slightly more depleted 13C between 1946 and 1990 at ca. –25‰ to –26‰, decreasing after 1990 towards –27‰ to –28‰. Between 1947 and 1970, barley δ13C are ca. 1‰ more depleted compared to wheat δ13C. Between 1970 and 1990, the difference seems to be smaller at ca. 0.8‰, and after 1995 the difference appears on average 0.5‰ but with a larger internal barley 13C variability.
Compared to global atmospheric δ13C as compiled by Graven (2017) with an enhanced rate in δ13C decrease after 1960, Gatersleben wheat and Rothamsted barley show an increasing δ13C decline only after 1990. Increasing use of petrol products such as kerosene and natural gas for energy and heat production may explain the increased δ13Cbiomass decline as suggested by Dunbar et al. (Reference Dunbar, Scott and Tripney2024).
Carbon isotope discrimination
Figure 4(C, D) gives the 14C and 13C discrimination of Gatersleben seeds against the atmospheric source, i.e., NHZ1 summer 14C and atmospheric 13C (Graven et al. Reference Graven, Allison, Etheridge, Hammer, Keeling, Levin, Meijer, Rubino, Tans, Trudinger, Vaughn and White2017; Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022).
During the early and maximum phase of the bomb-pulse, calculated ∆∆14CGatersleben show a considerable large variability of ca. 15‰ (seed14C - NHZ1summer_14C) between 1955 and 1980 (see Figure 4C). We speculate that the larger 14C differences between summer NHZ1 and Gatersleben plants during that period may indicate local 14C differences within the atmosphere due to spatially heterogeneous isotopic composition within the Northern Hemisphere as has been recently described for the tropical South America (Ancapichún et al. Reference Ancapichún, De Pol-Holz, Christie, Santos, Collado-Fabbri, Garreaud, Lambert, Orfanoz-Cheuquelaf, Rojas, Southon, Turnbull and Creasman2021). Large differences are also observed in Colorado Plateau plant 14C (s.o.) and may indicate similar atmospheric processes. Clearly, more seasonal 14C data and an expanded geographic distribution would be needed to constrain temporal delays in certain carbon sinks.
From about 1980 onwards, Gatersleben plant 14C values are closer to summer NHZ1 concentrations with ∆∆14C varying between –5‰ to 10‰ (see Figure 4), indicating on average a fossil carbon concentration of about 1% when compared to the supposedly clean Jungfraujoch atmospheric 14C record (Emmenegger et al. Reference Emmenegger, Leuenberger and Steinbacher2020; Hammer and Levin Reference Hammer and Levin2017; Hüls et al. Reference Hüls, Börner and Hamann2021).
Plant tissue isotopes are determined by the source carbon isotopic composition (13,14CO2_atmosphere) and physiological processes associated with photosynthesis, gas and water exchange. Heat and limited water availability cause closing of leaf stomata, limiting the gas exchange with the atmosphere to preserve moisture and thereby influencing stable carbon isotope composition of plant tissue. As discussed by Wallace et al. (2013), different plant parts show different Δ1 3C values caused by different biochemical compositions, with grains, mainly comprising simple biochemical compounds such as starch, usually having lower Δ1 3C than other plant parts. Figure 4D depicts the discrimination of 13C (Δ13Cwheat) of wheat with respect to atmospheric 13CO2 (Graven et al. Reference Graven, Allison, Etheridge, Hammer, Keeling, Levin, Meijer, Rubino, Tans, Trudinger, Vaughn and White2017). Gatersleben wheat 13C discrimination varies between 15.4‰ and 20.8‰, which is larger than what was observed during growth experiments conducted by Wallace et al. (2013) in Syria on growth plots with (maximum Δ13Cwheat19.36 ‰) and without irrigation (minimum Δ13Cwheat 16.66‰). In addition, these authors observed a rather low correlation of wheat Δ13C and an aridity index. At Gatersleben, experimental growth plots were not irrigated regularly unless during extreme droughts, and less so with cereals and more with soybeans (not shown).
A comparison of Δ13Cwheat with local weather parameters such as the average 2-m height April–July temperature and precipitation (TAMJJ, PAMJJ, see Figure 4(D, F), taken from the local Gatersleben Weather Station (https://meta.icos-cp.eu/objects/X-lXPKZlO4DWX7wncsLQ7akY), indicate no direct correlation with Δ13Cwheat. Average measured Δ13Cwheat values are 17.5‰, close to the threshold referenced in Wallace et al. (2013) for well-watered wheat crops.
Obviously, calculated Δ13Cwheat mirrors measured δ13Cwheat distribution due to the smooth atmospheric δ13C record used and may indicate here that calculated Δ13Cwheat variability is driven by other factors than precipitation and temperature alone. For example, recent greenhouse growth experiments with beach grasses have shown the influences of sea-spray on plant-tissue carbon isotope composition (14C, 13C) from the uptake of 14C-depleted irrigation waters and also increased carbon isotope discrimination related to the DIC (dissolved inorganic carbon) and salt content treatment spray- and irrigation waters (Göhring et al. Reference Göhring, Hüls, Hölzl, Mayr, Strauss and Hamann2025).
Discussion
Seasonally growing plant tissue 14C such as seen in the Gatersleben seeds (wheat and soybean) should mirror atmospheric radiocarbon during the vegetation period, which in turn results from a complex interplay between 14C production in the upper atmosphere, the partitioning into the specific sinks of the biosphere and hydrosphere, and dilution with man-made 14C-free carbon emission. The short-term, inter-annual seasonal variability as seen in reference atmospheric radiocarbon records such as Jungfraujoch (JFJ) (Emmenegger et al. Reference Emmenegger, Leuenberger and Steinbacher2020; Hammer and Levin Reference Hammer and Levin2017), could be caused by cold-season heating and combustion of fossil fuels for energy. Changes in the ocean–atmosphere exchange by varying strength in ocean upwelling (e.g., Graven et al. Reference Graven, Guilderson and Keeling2012), which could release older and 14C depleted CO2 into the atmosphere, could also contribute to atmospheric 14C inventory. For example, since the early 2000s, the North and South Pacific surface ocean became a 14C source as shown by annually precipitated carbonate of living corals of Porites sp. (Andrews et al. Reference Andrews, Siciliano, Potts, Demartini and Covarrubias2016; Wu et al. Reference Wu, Fallon, Cantin and Lough2021), which influences overlying northern and southern hemispheric airmasses.
During the last two decades, visible pauses in the rate of 14C decrease of Gatesleben plants 2008–2009 and 2019–2020 (see Figure 3D) visually coincide with the sunspot minima at the end of the 23rd and 24th solar cycles, respectively (McIntosh et al. Reference McIntosh, Chapman, Leamon, Egeland and Watkins2020), with a larger 14C production due to less solar particle shielding against cosmic particles (Stuiver Reference Stuiver1961; Stuiver and Braziunas Reference Stuiver and Braziunas1993). Also, fossil carbon emission to the atmosphere dropped significantly between 2008 and 2009 (Eurostat 2020), largely caused by the Global Financial Crisis in 2008.
A 14C decrease as seen in Gatersleben seeds in 2010 is also observed in maize leaves collected in the northern Netherlands in 2010 and 2011, with ∆∆14CJFJ-leaves of about 6‰ (Bozhinova et al. Reference Bozhinova, Palstra, van der Molen, Krol, Meijer and Peters2016), and in 2011 samples of Hungarian acacia honey, giving ∆∆14CJFJ-honey of about 10‰ (Varga et al. Reference Varga, Sajtos, Gajdos, Jull, Molnár and Baranyai2020).
Despite an average fossil-fuel carbon contribution of ca. 1% (Hüls et al. Reference Hüls, Börner and Hamann2021), we believe that this record as well as other plant-based 14C records as shown above, could be useful in forensic investigations such as paper document age estimates (Huels et al. Reference Huels, Pensold and Pigorsch2017; Pigorsch et al. Reference Pigorsch, Kießler, Hüls and Meinl2020) or the determination of the bio-carbon content in fuels (e.g., Culp et al. Reference Culp, Cherkinsky and Ravi Prasad2014).
Conclusion
Analysis of seasonally growing annual plants, photosynthesizing and metabolizing over a short period, may enable a better temporal resolution of the 14C record as compared to records from perennial plants such as tree-rings. Radiocarbon measurements of seasonal cultivated and archived seeds (wheat: Triticum aestivum L., soy: Glycine max (L.) Merr.) from the IPK Gatersleben track plant tissue 14C contents during the growth seasons between 1946 and 2025. Wheat samples consist of winter or spring type with growth periods between April–July and May–August, respectively. Soy on the other hand, is sown around mid-May and harvested End September–early October.
Gatersleben seed 14C concentrations follows datasets such as the Northern Hemisphere (NHZ1) record (e.g., Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022), i.e., with a steep increase in 14C between 1955–1964, followed by a steep reduction in atmospheric radiocarbon concentration after the LTBT caused by the partitioning into the biosphere and hydrosphere. The rate of reduction in atmospheric 14C slows down significantly in following decades. During recent decades, the reduction in atmospheric 14C concentration appeared discontinuous with sudden drops. Recent (2020–2025) measured Gatersleben seed 14C concentrations vary around F14C of 0.9959 to 0.9924 and Δ14C of –13.10 to –16.57‰.
The Gatersleben seed 14C record could indicate a fossil carbon contribution around 1%. Measured δ13C of Gatersleben wheat compares reasonably well to the recently published record of barley δ13C from the Rothamsted Institute in the UK, showing a sharp intensification of the decline in δ13C after 1990, probably caused by a changed δ13C composition of fossil carbon emissions (perhaps related to increasing fractions of natural gas relative to coal) as also suggested by Dunbar et al. (Reference Dunbar, Scott and Tripney2024).
A comparison of different 14C time series over the last 80 years from the Colorado Plateau/USA, located within NHZ2 (Carbone et al. Reference Carbone, Ayers, Ebert, Munson, Schuur and Richardson2023), England (Dunbar et al. Reference Dunbar, Scott and Tripney2024), and Germany (Gatersleben seed), both in NHZ1, reveal discrepancies with reference data, e.g., Northern Hemisphere Zone 1 & 2, and could indicate heterogenous atmospheric 14C composition from the admixture of different air masses. For example, pre-bomb plant tissue 14C of Gatersleben wheat and soybeans, Rothamsted barley as well as Colorado plateau plants, give lower 14C compared to monthly NHZ1 and NHZ2 data (Hua et al. Reference Hua, Turnbull, Santos, Rakowski, Ancapichún, Pol-Holz, Hammer, Lehman, Levin, Miller, Palmer Jonathan and Turney2022), and Rothamsted barley show higher 14C concentrations during the 1970s and during 2000–2010.
Overall, Gatersleben plant and atmospheric reference 14C differences appear to be driven by large concentration variability during peak nuclear testing phase and atmospheric air mass variability. After 1980, observed 14C differences are probably driven by regional fossil-fuel derived 14CO2 emissions and the dilution of atmospheric 14C with fossil, 14C-free carbon, superimposed on the natural atmospheric 14C-decline from carbon uptake in natural sinks (biosphere, ocean).
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/RDC.2026.10225
Acknowledgments
The authors would like to thank the technical staff of the Leibniz-Laboratory for their commitment in sample handling and the technical staff of IPK Gatersleben involved in handling and storing of seed samples. We appreciated the constructive comments of reviewers and the associate editor helping to shape the manuscript.







 Open access
Open access