



Introduction
CRC is one of the leading causes of cancer-related morbidity and mortality worldwide, highlighting the urgent need for innovative therapeutic strategies (Ref. Reference Nguyen and Shanmugan1). Multimodal therapy, integrating surgery, chemotherapy, radiotherapy and targeted agents, has emerged as a cornerstone of CRC management, significantly improving survival rates and reducing local recurrence by addressing the heterogeneous nature of the disease through complementary mechanisms (Ref. Reference Wiegering, Isbert, Dietz, Kunzmann, Ackermann, Kerscher, Maeder, Flentje, Schlegel, Reibetanz, Germer and Klein2). Recent advances in molecular biology and microbiome research have unveiled intricate relationships between gut microbiota, genetic regulation and cellular homeostasis, suggesting a multifaceted approach to CRC management (Refs Reference Roy, Cao, Singh, Fang, Yang and Wei3, Reference Rahimi-Kolour, Eshaghi, Shams, Sanjabi, Nobili, Raeisi, Sadeghi, Nazemalhosseini-Mojarad and Fatemi4).
Probiotics, defined as live microorganisms that confer health benefits to the host, have garnered attention for their role in modulating the composition and functionality of the gut microbiota (Ref. Reference Mafe, Iruoghene Edo, Akpoghelie, Gaaz, Yousif, Zainulabdeen, Isoje, Igbuku, Opiti, Garba, Essaghah, Ahmed and Umar5). Specific strains, such as Lactobacillus (L.) and Bifidobacterium (B.), have demonstrated anti-inflammatory effects and the ability to enhance gut barrier integrity (Ref. Reference Smolinska, Popescu and Zemelka-Wiacek6). These mechanisms are fundamental in CRC, where dysbiosis – a microbial imbalance – has been implicated in tumourigenesis (Ref. Reference He, Qi, Shu, Ding, Zeng, Wen, Xiong and Deng7). Additionally, probiotics may influence the immune response by promoting the production of short-chain fatty acids (SCFAs), which possess anti-carcinogenic properties and can inhibit tumour cell proliferation (Ref. Reference Ramesh, Srinivasan, Subbarayan, Chauhan, Krishnamoorthy, Kumar, Krishnan and Shrestha8).
Non-coding RNAs (ncRNAs), particularly miRNAs and lncRNAs, are emerging as key regulators of gene expression in CRC. These molecules modulate cellular processes such as cell proliferation, apoptosis and metastasis (Ref. Reference Farzam, Eslami, Jafarizadeh, Alamdari, Dabbaghipour, Nobari and Baradaran9). Dysregulation of specific ncRNAs has been associated with CRC progression, making them potential biomarkers for diagnosis and prognosis (Ref. Reference Tripathi and Jain10). Furthermore, ncRNAs can interact with signalling pathways involved in autophagy, a cellular process that maintains homeostasis by degrading damaged organelles and proteins (Ref. Reference Liu and Zhang11).
Autophagy is a highly conserved lysosome-dependent catabolic pathway that is critical in maintaining cellular homeostasis, function and development. It recycles damaged molecules and organelles, allowing cells to adapt to stress conditions such as nutrient deprivation, hypoxia and oxidative stress (Ref. Reference Russell, Yuan and Guan12). Autophagy plays a dual role in cancer: it can suppress tumour initiation by removing damaged cellular components, but it may also promote the survival of established tumours under stress conditions (Ref. Reference Tan, Luo, Li, Ling, Wei, Wang and Wang13). Because of this paradoxical role, researchers have increasingly focused on autophagy as a therapeutic target. Both inhibitors and activators of this pathway have been explored, ranging from repurposed drugs such as chloroquine, initially applied in malaria and autoimmune disorders, to mTOR inhibitors like rapamycin. However, their clinical use is limited by considerable side effects (Refs Reference Liu, Yang, Zou and Qu14, Reference Levy, Towers and Thorburn15). In addition, several naturally occurring compounds, including spermidine and trehalose, have been suggested as potential autophagy modulators. Still, their precise mechanisms of action and long-term safety profiles remain to be clarified (Refs Reference Mizunoe, Kobayashi, Sudo, Watanabe, Yasukawa, Natori, Hoshino, Negishi, Okita, Komatsu and Higami16, Reference Büttner, Broeskamp, Sommer, Markaki, Habernig, Alavian-Ghavanini, Carmona-Gutierrez, Eisenberg, Michael, Kroemer, Tavernarakis, Sigrist and Madeo17). Despite these pharmacological advances, there is growing interest in microbial-based approaches, particularly probiotics, which may influence autophagic pathways through physiologically relevant mechanisms that merit further investigation regarding their safety profile compared to synthetic agents (Refs Reference Lai, Tsai, Kuo, Ho, Day, -y, Chung, Huang, Wang, Liao and Huang18, Reference Wu, Wang, Zou, Wang, Sun, Fu, Wang, Wang, Xu and Li19, Reference Yang, Li, Tye, Chen, Lu, He, Zhou and Xiao20).
The intricate relationship between autophagy, probiotics and ncRNAs adds another layer of complexity to CRC management. Recent studies suggest probiotics can modulate autophagic activity, suppressing tumour growth (Ref. Reference Thoda and Touraki21). Compared to conventional pharmacological agents, probiotics and their derivatives may represent a safer and more cost-effective strategy for modulating autophagy, with fewer adverse effects and simpler formulations (Refs Reference Papadimitriou, Zoumpopoulou, Foligné, Alexandraki, Kazou, Pot and Tsakalidou22, Reference Stavropoulou and Bezirtzoglou23). However, despite increasing evidence, the precise molecular mechanisms through which probiotics regulate autophagy via ncRNA-mediated pathways remain largely unclear, representing a critical gap for future investigation.
Additionally, ncRNAs have been shown to regulate autophagy-related genes, influencing cellular responses to therapeutic interventions (Ref. Reference Jasim, Almajidi, Al-Rashidi, Hjazi, Ahmad, Alawadi, Alwaily, Alsaab, Haslany and Hameed24). The synergistic interplay among these three components – probiotics, ncRNAs and autophagy – highlights a promising avenue for developing precision-based therapeutic strategies to enhance treatment efficacy and improve patient outcomes.
To further refine these personalized approaches, increasing attention is being directed towards biological sex as a potential determinant of therapeutic success. It is well established that CRC exhibits significant sex-based dimorphism, with clinical observations indicating that women have higher rates of both right-sided tumours and microsatellite instability (MSI)-high tumours compared to men (Refs Reference Baraibar, Ros, Saoudi, Salvà, García, Castells, Tabernero and Élez25, Reference Choi, Kim, Nam, Kim, Song, Na and Kang26). Beyond these epidemiological patterns, emerging evidence suggests the existence of a microgenderome, in which sex hormones, particularly oestrogen, may influence gut microbial diversity and intestinal barrier integrity (Refs Reference Vemuri, Sylvia, Klein, Forster, Plebanski, Eri and Flanagan27, Reference Kaliannan, Robertson, Murphy, Stanton, Kang, Wang, Hao, Bhan and Kang28). While this review primarily aims to elucidate the fundamental and relatively sex-neutral molecular crosstalk within the probiotic–ncRNA–autophagy axis, we acknowledge that these sex-dependent variables represent a critical frontier. Integrating such dimorphic factors into the proposed mechanistic framework will be essential for the future experimental validation and clinical translation of precision-based microbial therapies.
This narrative review synthesizes the intricate crosstalk within the probiotic–ncRNA–autophagy triad in CRC management, moving beyond descriptive roles to elucidate the underlying molecular mechanisms and their potential therapeutic implications. By integrating multi-omics insights from microbiology and molecular oncology, we provide a comprehensive framework aimed at bridging microbial modulation with epigenetic regulation, offering a new perspective for precision medicine in CRC treatment.
CRC pathogenesis: molecular and cellular mechanisms
CRC develops through the progressive accumulation of genetic and epigenetic alterations in the colonic and rectal epithelium, transitioning from normal mucosa to adenoma and invasive carcinoma (Ref. Reference Cao, Tian, Deng, Yang and Chen29). This progression, primarily characterized by the adenoma-carcinoma sequence and the serrated polyp pathway, is now recognized as a dynamic process heavily influenced by the gut microbial and metabolic landscape.
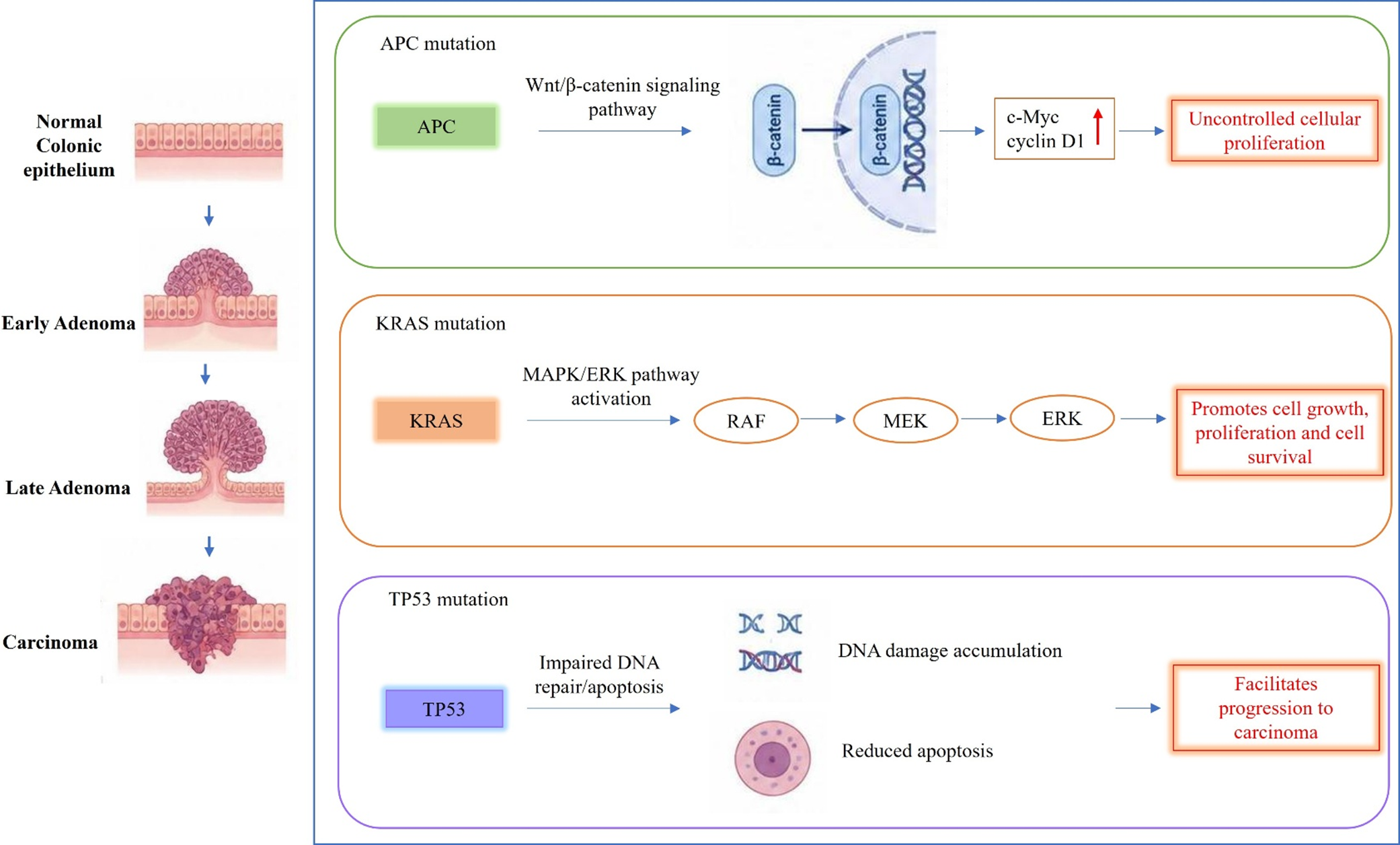
The adenoma-carcinoma sequence, which accounts for approximately 80% of CRC cases, is initiated by loss-of-function mutations in the Adenomatous Polyposis Coli (APC) gene, a key tumour suppressor (Figure 1) (Ref. Reference Li, Geng, Luo, Wang, Mo, Luo, Wang, Song, Sheng and Xu30). These mutations activate the Wnt/β-catenin signalling pathway, resulting in β-catenin stabilization and nuclear translocation. This process upregulates oncogenes such as c-Myc and cyclin D1, driving uncontrolled cellular proliferation (Ref. Reference Goh, Ji, Phang, Gu, Nair, Wei, Ansari and Liew31). Subsequent mutations in KRAS activate the MAPK/ERK pathway, further promoting cell growth and survival. Additionally, TP53 mutations impair DNA repair and apoptosis, facilitating progression to carcinoma (Ref. Reference Xu, Ren, Wang, Zhang and Lu32). This pathway is predominantly associated with chromosomal instability (CIN), characterized by numerical and structural chromosomal aberrations, loss of heterozygosity and deletion of tumour suppressor genes. It is observed in approximately 70% of sporadic CRC cases (Ref. Reference Chen, Agustinus, Li, Dibona and Bakhoum33).
A schematic illustration of the adenoma-carcinoma sequence in CRC. The figure highlights the sequential involvement of APC, KRAS and TP53 mutations and their associated signalling pathways during tumour progression. Figure created using BioRender.com.

Figure 1. Long description
The diagram is divided into a vertical anatomical column on the left and three horizontal molecular pathway panels on the right.
Anatomical Progression (Left Column):
* Top: Normal Colonic epithelium showing a flat layer of cells.
* Second: Early Adenoma showing a small protrusion of cells.
* Third: Late Adenoma showing a larger, mushroom-shaped growth.
* Bottom: Carcinoma showing an invasive, irregular mass penetrating the tissue layers.
Molecular Pathways (Right Panels):
1. A P C mutation (Green panel): A P C leads to the Wnt/beta-catenin signaling pathway. An arrow shows beta-catenin moving into a cell nucleus containing D N A. This leads to c-Myc and cyclin D 1 with a red upward arrow, resulting in Uncontrolled cellular proliferation.
2. K R A S mutation (Orange panel): K R A S leads to M A P K/E R K pathway activation. A linear sequence of arrows connects R A F to M E K to E R K. This results in Promotes cell growth, proliferation and cell survival.
3. T P 5 3 mutation (Purple panel): T P 5 3 leads to Impaired D N A repair/apoptosis. This is illustrated by D N A damage accumulation (shown as broken D N A strands) and Reduced apoptosis (shown as a shrinking cell). This facilitates progression to carcinoma.
The serrated polyp pathway, comprising 15%–20% of CRC cases, involves hyperplastic polyps and sessile serrated adenomas. This pathway is frequently associated with BRAF V600E mutations and the CpG Island Methylator Phenotype (CIMP) (Ref. Reference Wang, Xu, Hu, Li, Yao, Han, Zhang and Qu34). It leads to MSI due to defective DNA mismatch repair (MMR), primarily through epigenetic silencing of MLH1 or germline mutations in MMR genes (MLH1, MSH2, MSH6, PMS2) (Ref. Reference Schöniger and Rüschoff35). MSI-high (MSI-H) tumours, which are prevalent in Lynch syndrome (95%) and approximately 20% of sporadic cases, are typically right-sided, exhibit a high mutational burden and are more immunogenic, potentially benefiting from immunotherapy (Refs Reference Grigorie, Potlog and Alexandrescu36, Reference Palmeri, Mehnert, Silk, Jabbour, Ganesan, Popli, Riedlinger, Stephenson, de Meritens, Leiser, Mayer, Chan, Spencer, Girda, Malhotra, Chan, Subbiah and Groisberg37).
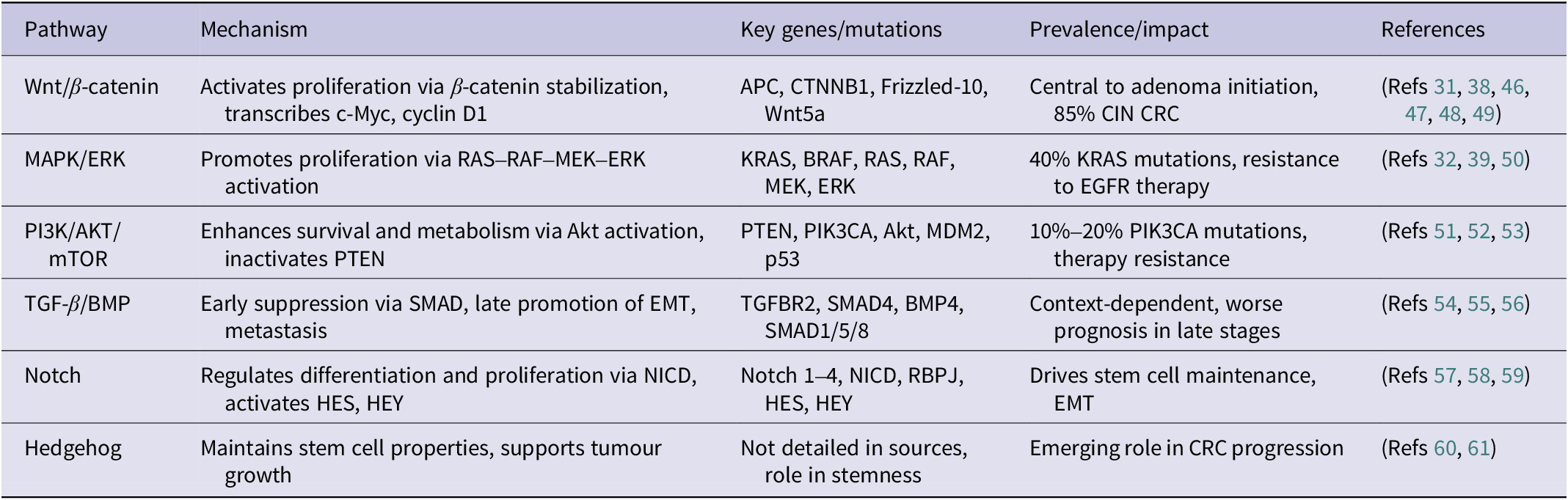
Several signalling cascades play a central role in CRC pathogenesis, as detailed in Table 1. The Wnt/β-catenin pathway is particularly critical, with miR-135 and miR-155 targeting APC. Therapeutic interventions include Porcupine inhibitors (e.g., LGK974 and ETC-159) and β-catenin degraders (e.g., PRI-724) (Ref. Reference Díaz, Martín and Gentili38). The MAPK/ERK pathway, characterized by KRAS mutations in ~40% of cases, is targeted by inhibitors such as sotorasib (Ref. Reference Dillon, Lopez, Lin, Sales, Perets and Jain39). The PI3K/AKT/mTOR pathway, mutated in 10%–18% of cases, is addressed by drugs like idelalisib (Ref. Reference Machado, Machado, Pessoa, Barreto, Gadelha, DDS, Ribeiro, Lopes, Moraesfilho, Moraes, Khayat and Moreira-Nunes40). CIN, affecting 70% of CRC cases, results in widespread genomic alterations. In contrast, MSI, driven by MMR deficiency, leads to a high neoantigen load that enhances immunogenicity (Refs Reference Kim and Bodmer41, Reference Amodio, Vitiello, Bardelli and Germano42), while the CIMP, involving hypermethylation of CpG islands, silences tumour suppressor genes such as p16 and MGMT. CIMP often co-occurs with BRAF mutations and is associated with a poorer prognosis (Ref. Reference Fennell, Kane, Liu, McKeone, Fernando, Su, Bond, Jamieson, Dumenil, Patch, Kazakoff, Pearson, Waddell, Leggett and Whitehall43). Right-sided CRCs, which are associated with MSI-H, CIMP and BRAF mutations, generally have worse outcomes, except for MSI-H tumours, which respond well to immunotherapy agents like pembrolizumab and nivolumab (Ref. Reference Han, Chok, Peh, Ho, Tan, Koo, Tan and Ong44). In contrast, left-sided CRCs, linked to CIN and TP53/KRAS mutations, typically have a better prognosis (Ref. Reference Gavrić, Hodžić, Sarajlić, Salibašić, Bajramagić, Dizdarević and Kulović45). The Consensus Molecular Subtypes (CMS) further classify CRC into four groups: CMS1 (14%, MSI Immune, BRAF mutations, 73% 5-year survival), CMS2 (37%, Canonical, TP53, EGFR, 77% 5-year survival), CMS3 (23%, Metabolic, KRAS mutations, 75% 5-year survival) and CMS4 (13%, Mesenchymal subtype, associated with worse prognosis) (Ref. Reference Gavrić, Hodžić, Sarajlić, Salibašić, Bajramagić, Dizdarević and Kulović45). CRC pathogenesis is complex; however, the individual and synergistic effects of probiotics, ncRNAs and autophagy may offer novel therapeutic strategies for CRC by targeting these interconnected pathways to improve treatment outcomes.
Key signalling pathways involved in CRC pathogenesis

Table 1. Long description
The table consists of five columns and six rows of data.
* Row 1: Wnt/beta-catenin pathway. Mechanism: Activates proliferation via beta-catenin stabilization, transcribes c-M y c, cyclin D 1. Key genes: A P C, C T N N B 1, Frizzled-10, Wnt5a. Impact: Central to adenoma initiation, 85% C I N C R C.
* Row 2: M A P K/E R K pathway. Mechanism: Promotes proliferation via R A S-R A F-M E K-E R K activation. Key genes: K R A S, B R A F, R A S, R A F, M E K, E R K. Impact: 40% K R A S mutations, resistance to E G F R therapy.
* Row 3: P I 3 K/A K T/m T O R pathway. Mechanism: Enhances survival and metabolism via Akt activation, inactivates P T E N. Key genes: P T E N, P I K 3 C A, Akt, M D M 2, p 53. Impact: 10% to 20% P I K 3 C A mutations, therapy resistance.
* Row 4: T G F-beta/B M P pathway. Mechanism: Early suppression via S M A D, late promotion of E M T, metastasis. Key genes: T G F B R 2, S M A D 4, B M P 4, S M A D 1/5/8. Impact: Context-dependent, worse prognosis in late stages.
* Row 5: Notch pathway. Mechanism: Regulates differentiation and proliferation via N I C D, activates H E S, H E Y. Key genes: Notch 1 to 4, N I C D, R B P J, H E S, H E Y. Impact: Drives stem cell maintenance, E M T.
* Row 6: Hedgehog pathway. Mechanism: Maintains stem cell properties, supports tumour growth. Key genes: Not detailed in sources. Impact: Emerging role in C R C progression.
Abbreviations: APC: adenomatous polyposis coli; CIN: chromosomal instability; CRC: colorectal cancer; MAPK: mitogen-activated protein kinase; ERK: extracellular signal-regulated kinase; EGFR: epidermal growth factor receptor; PI3K: phosphoinositide 3-kinase; mTOR: mammalian target of rapamycin; PTEN: phosphatase and tensin homolog; TGF-β: transforming growth factor beta; BMP: bone morphogenetic protein; SMAD: suppressor of mothers against decapentaplegic; EMT: epithelial–mesenchymal transition; NICD: intracellular domain of notch.
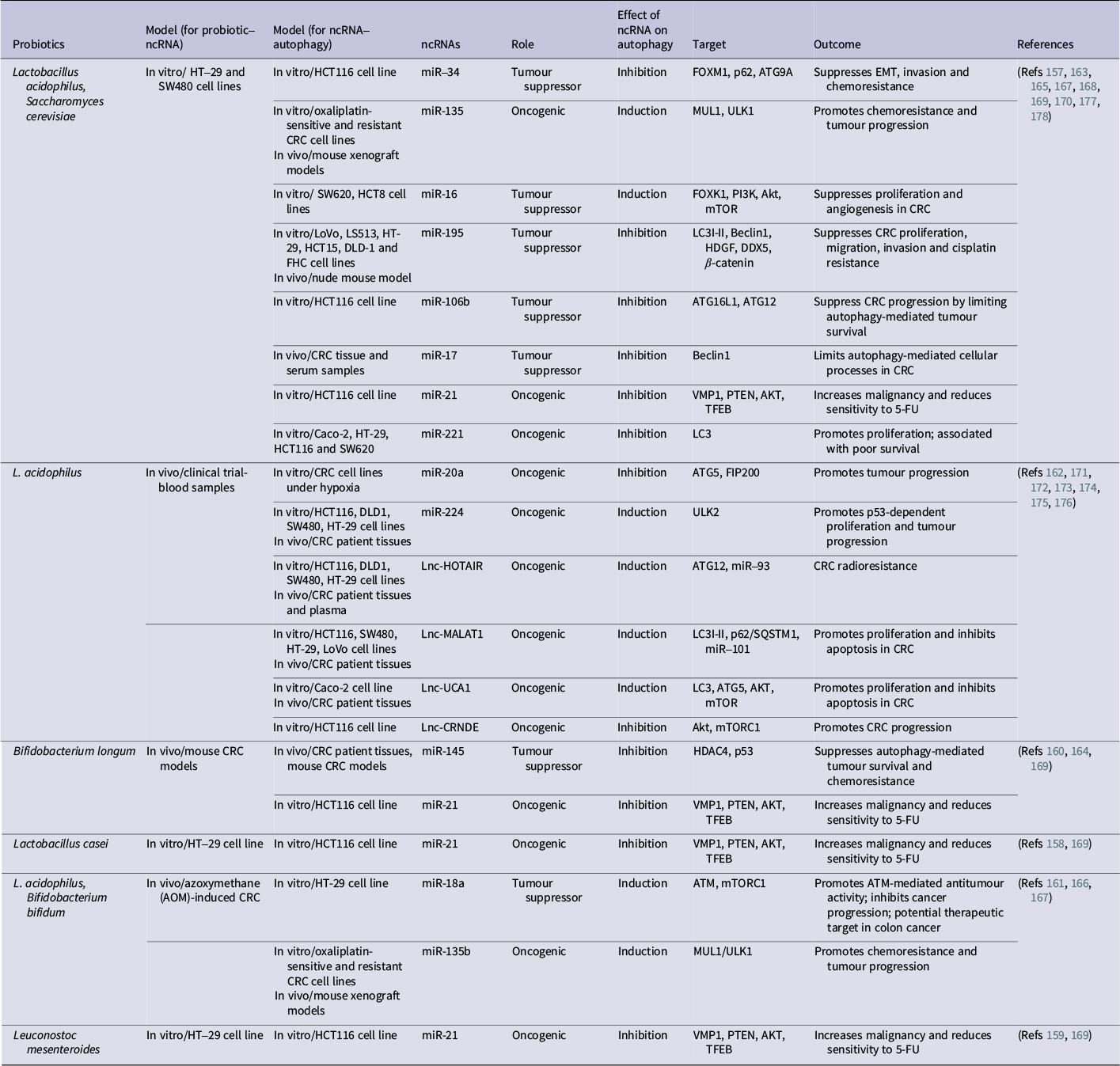
The role of ncRNAs associated with probiotics in CRC, focusing on autophagy

Table 2. Long description
The table details the interactions between specific probiotics and non-coding R N As (n c R N As) in colorectal cancer (C R C) models.
- Lactobacillus acidophilus and Saccharomyces cerevisiae: Associated with multiple m i Rs. m i R 34, m i R 16, m i R 195, m i R 106b, and m i R 17 act as tumor suppressors, while m i R 135, m i R 21, and m i R 221 are oncogenic. Targets include F O X M 1, U L K 1, P I 3 K, Beclin 1, and A T G 12. Outcomes range from suppressing E M T and chemoresistance to promoting tumor progression.
- Lactobacillus acidophilus (alone): Linked to oncogenic factors like m i R 20a, m i R 224, and long non-coding R N As (L n c R N As) including H O T A I R, M A L A T 1, U C A 1, and C R N D E. These generally induce autophagy (except m i R 20a and L n c C R N D E which inhibit it) to promote tumor progression, radioresistance, and inhibit apoptosis.
- Bifidobacterium longum: Associated with tumor suppressor m i R 145 (inhibits autophagy via H D A C 4) and oncogenic m i R 21 (inhibits autophagy via V M P 1).
- Lactobacillus casei and Leuconostoc mesenteroides: Both are linked to oncogenic m i R 21, which inhibits autophagy and increases malignancy.
- Lactobacillus acidophilus and Bifidobacterium bifidum: Associated with tumor suppressor m i R 18a (induces autophagy via A T M and m T O R C 1) and oncogenic m i R 135b (induces autophagy via M U L 1 and U L K 1).
Recent advances in the knowledge of CRC pathogenesis have elucidated key molecular pathways, enabling targeted therapies such as sotorasib for KRAS mutations and pembrolizumab for MSI-H tumours, particularly improving outcomes for specific subtypes. However, an incomplete understanding of epigenetic mechanisms, such as the role of the CIMP in therapy resistance and the lack of longitudinal studies to validate CMS classifications across diverse populations, limits broader clinical application. These gaps highlight the need for innovative therapeutic strategies targeting unexplored pathways and encourage further research into integrative approaches for precision medicine in CRC.
Microbial interventions for CRC management
Recent research has explored the therapeutic potential of probiotics, prebiotics, synbiotics and postbiotics in modulating the gut microenvironment to prevent and manage CRC (Ref. Reference Cheraghpour, Fatemi, Shadnoush, Talebi, Tierling and Bermúdez-Humarán62). However, the success of these interventions in reshaping the microbial landscape is significantly dictated by the host’s biological sex and environmental background. Building upon the sex-based dimorphism mentioned earlier, empirical evidence indicates that women typically exhibit a higher abundance of Akkermansia muciniphila and Lactobacillus species, which contribute to a more robust intestinal barrier, whereas Bacteroides and Prevotella are frequently more dominant in the male gut (Ref. Reference Yong-Sung, Tatsuya and Byung-Yong63). These variations are not merely taxonomic but functional; oestrogen-driven metabolic pathways influence the production of SCFAs, meaning that the same prebiotic or probiotic strain may yield different concentrations of anti-inflammatory metabolites depending on the host’s hormonal milieu (Refs Reference Lephart and Naftolin64, Reference Wang, Shi, Zheng, Zhou, Mi, Wu and Feng65). Beyond biological sex, dietary habits associated with different regions further influence these outcomes; for instance, the ‘Western diet’ common in the United States and Europe often fosters a pro-inflammatory microbiota, while traditional diets in the Far East and Middle East, rich in fermented foods and fibres, may enhance the efficacy of probiotic-based precision medicine (Refs Reference Malesza, Malesza, Walkowiak, Mussin, Walkowiak, Aringazina, Bartkowiak-Wieczorek and Mądry66, Reference Dimidi, Cox, Rossi and Whelan67). Moreover, the distinct microbial enterotypes observed in these regions are often shaped by a combination of traditional dietary patterns and environmental exposures that differ from the high-urbanization stressors of Western societies. These cultural and lifestyle-specific factors contribute to a higher prevalence of beneficial taxa, such as certain lactic acid bacteria, which have been shown to synergize with microbial interventions to suppress the pro-tumourigenic inflammatory markers associated with CRC (Ref. Reference Garavaglia, Vallino, Ferraresi, Visciglia, Amoruso, Pane, Munteanu and Isidoro68). Understanding these regional lifestyle–microbiota interactions is essential for tailoring synbiotic therapies to the host’s baseline ecological state. Consequently, sex-specific and regional microbial signatures have emerged as critical predictors for clinical outcomes, reinforcing the necessity of adopting stratified protocols when dosing synbiotics or selecting postbiotic derivatives.
These interventions primarily reshape microbial composition and activity, serving as an upstream trigger that influences downstream molecular networks, including epigenetic regulators and autophagic flux. From a mechanistic perspective, probiotics inhibit pathogenic bacteria and reduce inflammation, thereby indirectly modulating pathways such as Wnt/β-catenin, which is often dysregulated in CRC due to chronic inflammation (Ref. Reference Cui, Luo, He, Liu, Lv, Zhang, Su, Zheng, Lu, Wang, Yang, Zhao, Liu, Wang, Zhao, Nie, Jiang, Zhang, Liu, Chen, Cai, Lv, Liu, An, Zhang, Yan, Kelley, Xu, Xu, Liu and Peng69). Si et al. (Ref. Reference Si, Liang, Bugno, Xu, Ding, Yang, Fu, Weichselbaum, Zhao and Wang70) reported that Lactobacillus rhamnosus strains can stimulate antitumour responses by activating the cGAS/STING pathway, thereby enhancing IFN-α/β signalling and cytotoxic CD8+ T-cell activity, emphasizing the contribution of gut microbiota to immune regulation in CRC therapy. Moreover, probiotics improve gut barrier function by upregulating tight junction proteins (e.g., claudin-1, occludin and ZO-1/2) and increasing mucin production, which may reduce cellular stress and indirectly influence survival pathways like PI3K/AKT/mTOR (Ref. Reference Mohseni, Casolaro, Bermúdez-Humarán, Keyvani and Taghinezhad-S71). Cellular models of inflammatory bowel disease have confirmed that B. infantis inhibits the PI3K-Akt–mTOR pathway and promotes Foxp3 expression through PD-L1, suggesting potential links to broader regulatory processes (Refs Reference Stefani, Miricescu, Stanescu-Spinu, Nica, Greabu, Totan and Jinga51, Reference Zhou, Xie and Li72). Shang et al. (Ref. Reference Shang, Liu, Yang, Duan, Xie and Meng73) showed that Bifidobacterium bifidum H3-R2, when combined with Lactococcus lactis KLDS4.0325, triggers apoptosis in colon cancer cells by upregulating caspase-3 and caspase-9, increasing Bax levels and reducing Bcl-2 expression, thereby engaging apoptotic pathways that may intersect with cellular degradation processes, including autophagy, through shared regulatory nodes such as Bcl-2 family proteins and the PI3K/AKT/mTOR axis (Refs Reference Yu, Zhang and Hou74, Reference Yan, Yu, Guo, Zhu, Chen, Li, Zhong and Wang75). A 2021 study on Streptococcus thermophilus showed reduced tumour formation in CRC mouse models through β-galactosidase activity, which inhibits cell proliferation and promotes apoptosis (Ref. Reference Li, Hu, Liu, Zhao, Huang, Liu, Chan, Zhang, Zeng, Coker, Kang, Ng, Zhang, Wong, Gin, Chan, Wu, Yu and Wu76). Tiptiri-Kourpeti et al. (Ref. Reference Tiptiri-Kourpeti, Spyridopoulou, Santarmaki, Aindelis, Tompoulidou, Lamprianidou, Saxami, Ypsilantis, Lampri, Simopoulos, Kotsianidis, Galanis, Kourkoutas, Dimitrellou and Chlichlia77) reported that Lactobacillus casei ATCC 393 exerts antiproliferative and pro-apoptotic effects in vitro on both murine CT-26 and human HT-29 colon carcinoma cell lines, reducing cell viability in a concentration- and time-dependent manner through apoptosis, while in vivo oral administration of live bacteria to BALB/c mice bearing subcutaneous CT-26 tumours resulted in approximately 80% reduction in tumour volume, accompanied by upregulation of TRAIL and downregulation of survivin.
Prebiotics, non-digestible food ingredients that promote the growth and activity of beneficial gut microorganisms, exhibit chemopreventive and anticancer effects in CRC (Ref. Reference Shrifteylik, Maiolini, Dufault, Austin, Subhadra, Lamichhane and Deshmukh78). By fostering probiotic proliferation, they increase the production of SCFAs, such as butyrate, which contribute to anti-inflammatory and anticancer mechanisms potentially tied to epigenetic and metabolic regulation (Ref. Reference Son and Cho79). In vitro research demonstrates that prebiotics can suppress CRC cell proliferation; for instance, Tukenmez et al. (Ref. Reference Tukenmez, Aktas, Aslim and Yavuz80) showed that exopolysaccharides derived from probiotic bacteria exerted antiproliferative effects on HT-29 colon cancer cells by upregulating pro-apoptotic genes (e.g., Bax and Caspase) and downregulating anti-apoptotic genes (e.g., Bcl-2 and Survivin). Qamar et al. (Ref. Reference Qamar, Syed, Nasir, Rehman, Zahid, Liu and Iqbal81) reported that galactooligosaccharides (GOS) from Lactobacillus reuteri reduced aberrant crypt foci in rats dose-dependently, alongside elevated SCFA acid levels and beneficial bacterial populations. Similarly, Fernandez et al. (Refs Reference Fernández, Moreno, Olano, Clemente, Villar and Lombó82, Reference Fernández, Ledesma, Monte, Millán, Costa, de la Fuente, García, Martínez-Camblor, Villar and Lombó83) found that 10% (w/w) GOS-Lu treatment in rats led to a 57.5% reduction in polyps and a 50.4% decrease in mean tumour-affected area, with shifts in gut microbiota including increased Bacteroides and decreased Firmicutes. Ohara et al. (Ref. Reference Ohara and Suzutani84) reported in a pilot clinical trial that a combination of Bifidobacterium longum and fructooligosaccharide (FOS) elevated SCFA concentrations while suppressing Bacteroides fragilis enterotoxin, suggesting a protective role against CRC.
Synbiotics, combining probiotics and prebiotics, provide a synergistic strategy for improving gut health and addressing CRC (Ref. Reference Alam, Shang, Effat, Kanwal, He, Li, Xu, Niu, War and Zhang85). Stene et al. (Ref. Reference Stene, Xu, Fallone de Andrade, Palmquist, Molin, Ahrné, Thorlacius, Johnson and Jeppsson86) indicated that synbiotic interventions may safeguard rectal mucosa by alleviating inflammation and altering mucosa-associated microbiota in rectal cancer patients receiving radiotherapy. Preoperative synbiotic use in CRC patients undergoing colorectal resection has been linked to reduced morbidity, shorter hospital stays and decreased antibiotic requirements (Refs Reference Polakowski, Kato, Preti, Schieferdecker and Ligocki Campos87, Reference Flesch, Tonial, Contu and Damin88). A specific study by Theodoropoulos et al. (Ref. Reference Theodoropoulos, Memos, Peitsidou, Karantanos, Spyropoulos and Zografos89) used synbiotic Forte, containing multiple probiotic strains (Pediococcus pentosaceus 5–33:3, Leuconostoc mesenteroides 32–77:1, Lactobacillus paracasei ssp. paracasei 19 and L. plantarum 2362) and prebiotic fibres (2.5 g inulin, oat bran, pectin and resistant starch), and observed significant improvements in Gastrointestinal Quality of Life Index (GIQLI) scores and a reduction in diarrhoea in postcolectomy CRC patients. Additionally, Moreira et al. (Ref. Reference Moreira, Carriço, Capelas, Pimenta, Santos, Ganhão-Arranhado, Mäkitie and Ravasco90) showed that synbiotics promote beneficial bacteria and SCFAs while reducing pathogenic bacteria, reinforcing their protective potential in CRC. These results imply additive advantages for synbiotics in CRC management. However, additional studies are required to refine composition, dosing and long-term impacts, amid discussions on their benefits relative to probiotics alone.
Postbiotics, metabolic byproducts of probiotics including SCFAs, peptides and other metabolites, represent an emerging therapeutic avenue in CRC by directly influencing host biology through the modulating pathways relevant to cellular homeostasis and therapy responses (Ref. Reference Kumar, Singh, Bee, Rahaman, Kriti, Shivaram, Muztaba, Katari, Ashique, Mishra, Garg and Ashique91). Erfanian et al. (Ref. Reference Erfanian, Safarpour, Tavakoli, Mahdiabadi, Nasseri and Namaei92) reported the anti-proliferative and anti-migratory effects of B. breve and L. rhamnosus postbiotics on HT-29 cells. Postbiotics such as ferrichrome from L. casei have demonstrated selective cytotoxicity against CRC cell lines by inducing apoptosis via the JNK-DDTI3 signalling pathway (Ref. Reference Kvakova, Kamlarova, Stofilova, Benetinova and Bertkova93). A study by Kazmierczak-Siedlecka et al. (Ref. Reference Kaźmierczak-Siedlecka, Daca, Fic, van de Wetering, Folwarski and Makarewicz94), which included 15 CRC patients, found changes in the SCFA pool in stool samples, suggesting a potential role in CRC management. Postbiotics inhibit CRC cell proliferation and induce apoptosis by hyper-activating the Wnt/β-catenin pathway, enhancing the expression of P21waf1 and Bax (components of the p53 pathway) (Refs Reference Xie, Liu, Yu, Qiu, Jiang and Yu95, Reference Al-Khazaleh, Alsherbiny, Münch, Chang and Bhuyan96), downregulating miR-92a and upregulating miR-203, indicating ties to RNA-mediated regulation (Ref. Reference Om, Chand and Kushawaha97). Butyrate, as a postbiotic, has been shown to promote apoptosis by hyper-activating Wnt signalling in DLD-1 and HCT116 cells (Ref. Reference Wu, Wu, He, Wu, Wang and Liu98). Postbiotics also suppress angiogenesis and metastasis by decreasing neuropilin-1 (NRP-1) expression via the NRP-1/VEGF pathway; studies on HCT116, HT-29 and Caco-2 cells have demonstrated reduced NRP-1 levels (Ref. Reference Nenkov, Ma, Gassler and Chen99). By inhibiting glucose metabolism through the GPR109a-AKT pathway, postbiotics can reduce GLUT1 and G6PD abundance, and suppress glycolysis, a hallmark of cancer cells (Ref. Reference Artusa, Calabrone, Mortara, Peri and Bruno100). One study showed that butyrate markedly inhibits glucose transport and glycolysis in CRC cells, reduces membrane GLUT1 and enhances the efficacy of chemotherapy with 5-fluorouracil (5-FU) (Ref. Reference Geng, Yin, Zhang, Gong and Yang101). Postbiotics can also act as histone deacetylase inhibitors (HDACi), influencing gene expression and signalling pathways like ERK and Wnt, influencing tumour cell growth and apoptosis (Refs Reference Eladwy, Alsherbiny, Chang, Fares, Li and Bhuyan102, Reference Kaur, Kaur and Ali103). Although research on postbiotics is still nascent, preliminary evidence suggests their potential as adjunctive therapy in CRC, particularly when combined with conventional treatments. However, safety concerns remain, especially for immunocompromised patients.
Integrating probiotics, prebiotics, synbiotics and postbiotics into CRC management offers a promising approach for personalized medicine. Right-sided CRCs, often associated with MSI-H and CIMP, may benefit from these interventions due to their immunomodulatory effects, potentially enhancing responses to immunotherapy (Refs Reference Mukherji and Weinberg104, Reference Colombo, Concetta, Gebbia, Sambataro, Scandurra and Valerio105). Left-sided CRCs, linked to CIN and TP53/KRAS mutations, may experience reduced chemotherapy side effects through such treatments (Ref. Reference Baran, Ozupek, Tetik, Acar, Bekcioglu and Baskin106).
However, the field faces challenges, including variability in strain selection, dosage and patient-specific responses, necessitating large-scale, randomized, double-blind clinical trials to validate the safety and efficacy. This foundational role of microbial interventions in modulating pathways and immune responses positions them as potential bridges to molecular regulatory processes, setting the stage for exploring integrated strategies in CRC therapy.
Role of ncRNAs in CRC pathogenesis
The ncRNAs are RNA molecules that do not encode proteins but play critical regulatory roles in gene expression through mechanisms such as transcriptional regulation, posttranscriptional modification and epigenetic modulation (Ref. Reference George, Subramanian and Supriya107). Among ncRNAs, miRNAs, typically 20–22 nucleotides long, regulate gene expression by binding to target mRNAs, leading to their degradation or translational repression (Ref. Reference Seyhan108). The lncRNAs, longer than 200 nucleotides, exert their effects through chromatin remodelling, transcriptional regulation and interactions with other RNAs or proteins (Ref. Reference Ferrer and Dimitrova109). In CRC, recent studies have elucidated the roles of miRNAs and lncRNAs in tumourigenesis, including cell proliferation, apoptosis, invasion, metastasis and immune evasion (Ref. Reference Ma, Liu, Roopashree, Kazmi, Jasim, Phaninder Vinay, Fateh, Yang, Rajabivahid, Dehghani-Ghorbi and Akhavan110). NcRNAs are increasingly recognized as diagnostic biomarkers and therapeutic targets in CRC, as they modulate key cellular pathways and contribute to maintaining cellular homeostasis.
miRNAs
MiRNAs are crucial regulators in CRC, influencing tumour progression and immune modulation through interactions with target mRNAs (Ref. Reference Zare, Yaghoubi, Khoshnazar, Jafari Dargahlou, Machhar, Zheng, Duijf and Mansoori111). In vitro studies have shown that miR-15b-5p enhances T-cell infiltration by sequestering PD-L1, thereby improving T-cell responses (Ref. Reference Zabeti Touchaei and Vahidi112). In contrast, miR-148a-3p impairs CD8+ T cell-mediated immune attacks by silencing the CANX/MHC-I pathway, which is essential for antigen presentation (Ref. Reference Zabeti Touchaei and Vahidi113). Furthermore, miR-27a represses MHC I cell surface exposure, contributing to poor prognosis in CRC (Ref. Reference Colangelo, Polcaro, Ziccardi, Pucci, Muccillo, Galgani, Fucci, Milone, Budillon, Santopaolo, Votino, Pancione, Piepoli, Mazzoccoli, Binaschi, Bigioni, Maggi, Fassan, Laudanna, Matarese, Sabatino and Colantuoni114). Other miRNAs, such as miR-29b-3p, promote epithelial–mesenchymal transition (EMT) and enhance migration and invasion by targeting the H19/miR-29b-3p/PGRN axis (Ref. Reference Ding, Li, Zhao, Li, Yang and Zhang115), whereas miR-138-5p is involved in angiogenesis and immune regulation, promoting EMT through the CCAT1/miR-138-5p/HMGA1 axis (Ref. Reference Gao, Li, Liu, Zhang, Gao and Wang116). Dysregulation of miRNAs like miR-141 and miR-216a has been associated with tumour occurrence and metastasis, making them potential prognostic factors (Ref. Reference Klicka, Grzywa, Klinke, Mielniczuk and Włodarski117). Additionally, miRNAs are involved in therapy resistance; for example, miR-675-5p supports hypoxia-induced drug resistance by disrupting apoptotic signals, targeting pro-caspase-3 and enhancing 5-FU resistance (Ref. Reference Zichittella, Barreca, Cordaro, Corrado, Alessandro and Conigliaro118), while miR-27b-3p sensitizes CRC cells to oxaliplatin by inhibiting autophagy through downregulation of ATG10 (Ref. Reference Yejia, Jin, Jinjun, Dan, Yufeng, Aiping, Chen, Yao, He and Pu119). These studies demonstrate the dual roles of miRNAs in CRC, either promoting or inhibiting antitumour immune responses and therapy efficacy.
LncRNAs
LncRNAs play a pivotal role in CRC pathogenesis by influencing multiple cellular processes through diverse mechanisms (Ref. Reference Alshahrani, Al-Hadeithi, Almalki, Malviya, Hjazi, Mustafa, Alawady, Alsaalamy, Joshi and Alkhafaji120). Both human and cellular studies showed that MIR22HG acts as a tumour suppressor by facilitating immunotherapy, likely through the interaction with the TGFβ/SMAD signalling pathway, which correlates with increased CD8A expression and CD8+ T-cell infiltration (Refs Reference Xu, Shao, Song, Xie, Zhou, Yin, Ding, Zou, Li and Zhang121, Reference Al-Kateb, Heidarzadehpilehrood, Pirhoushiaran, Abdollahzadeh, Saffari, Majidzadeh-A, Layeghi and Modarressi122). Conversely, the lncRNA LINC00657 negatively correlates with CD8+ T-cell infiltration, suggesting a role in immune evasion (Ref. Reference Li, Wang, Liu, Lv, Man, Liang and Qiao123). A comprehensive bioinformatics analysis of 568 CRC patient samples from The Cancer Genome Atlas (TCGA) database indicated that lower expression of lncRNA like MIR4435-HG correlates with increased infiltration of plasma cells and resting memory CD4+ T cells. In contrast, higher expression levels of MIR4435-HG are linked to greater infiltration of neutrophils and follicular helper T cells (Ref. Reference Chen, Song, Li, Zhang, Lin, Sun, Wang, Liu, Guo and Yu124). Other notable lncRNAs include H19, which promotes CRC cell proliferation and chemoresistance by sponging miR-181a-5p, activating the Wnt/β-catenin signalling pathway (Refs Reference Yuan, Tao, Zhang, Zhang, Sun and Wu125, Reference Li, Wang, Liu, Gemingnuer, Wang and Meng126), and HOTAIR, involved in immune evasion by interacting with the PRC2 complex and modulating PD-L1 expression (Ref. Reference Xie, Xia, Jiao, Zhao, Wang, Lin, Xing, Li, Lu, Zhong, Miao, Zhou, Qian, Wang, Zhang, Gu, Chu and Liu127). Additionally, NEAT1 contributes to immune escape and cancer cell metabolism by regulating the miR-101-3p/ZEB1/PD-L1 axis (Ref. Reference Yin and Wang128). These findings illustrate the complex interplay between lncRNAs and immune responses, underscoring their significance in tumour progression, which is relevant to cellular homeostasis.
The dysregulation of ncRNAs in CRC has significant clinical implications, offering opportunities for non-invasive biomarkers and therapeutic interventions. Clinical studies have identified that circulating lncRNAs and miRNAs can be used as diagnostic tools for early detection and prognosis (Ref. Reference Lotfi, Kholghi, Golab, Mohammadi and Barati129). For example, lncRNAs such as CRNDE and ATB have been detected in plasma and are associated with CRC progression (Ref. Reference Saadh, Menon, Verma, Prasad, Allela, Mahdi, Ahmad and Husseen130). Similarly, miRNAs like miR-15b-5p and miR-148a-3p are potential biomarkers for monitoring immune responses. The therapeutic potential of targeting ncRNAs is being explored, with strategies such as miRNA mimics, inhibitors and lncRNA-targeted therapies showing promise in preclinical studies (Ref. Reference Robinson, Bertsch, Leithner, Stiegler, Olschewski and Hrzenjak131). Recent research by Kousar et al. (Ref. Reference Kousar, Ahmad, Abduh, Kanwal, Shah, Naseer and Anjum132) suggests that upregulating miR-15b-5p or downregulating miR-148a-3p could enhance immunotherapy efficacy; however, challenges such as developing effective delivery systems, ensuring specificity and minimizing off-target effects remain.
Identifying ncRNAs as key regulators of CRC progression and immune modulation has paved the way for innovative diagnostic and therapeutic strategies, particularly their roles as non-invasive biomarkers and targets for enhancing immunotherapy efficacy. However, the predominance of preclinical data, variability in ncRNA expression across CRC subtypes and challenges in achieving targeted delivery with minimal off-target effects limit their immediate clinical application. Future research should focus on large-scale clinical validation and the development of precise delivery systems to unlock the full therapeutic potential of ncRNAs in personalized CRC management.
Autophagy’s impact on CRC
Autophagy, a highly conserved cellular process involving the degradation and recycling of damaged organelles and proteins. While initially characterized as a tumour-suppressive mechanism, a growing body of evidence reveals a ‘double-edged sword’ phenomenon where autophagy can either inhibit or promote CRC progression depending on the tumour stage, genetic background and microenvironmental stress (Ref. Reference Rajendran and Oon133). At the mechanistic level, autophagy progresses through initiation, elongation and maturation, generating double-membrane autophagosomes that capture cellular components for lysosomal degradation and nutrient recycling (Refs Reference Yang, Klionsky, Levine, Yoshimori and Deretic134, Reference Li, He and Ma135, Reference Mostowy136, Reference Debnath, Gammoh and Ryan137). This process is orchestrated by autophagy-related proteins (Atgs) and regulated by AMPK and mTOR pathways (Refs Reference Kroemer, Mariño and Levine138, Reference Park, Lee and Kim139). Under nutrient or energy stress conditions, AMPK suppresses mTORC1 and activates the ULK1 complex to initiate phagophore formation (Refs Reference Park, Lee and Kim139, Reference Park, Jung, Seo, Otto, Grunwald, Kim, Moriarity, Kim, Starker, Nho, Voytas and Kim140). Subsequent steps, including LC3 lipidation and membrane expansion, lead to autophagosome maturation and lysosome fusion. Selective clearance of damaged proteins and organelles is mediated by adaptor proteins such as SQSTM1, thereby linking autophagy to cellular quality control and metabolic adaptation (Ref. Reference Russell, Yuan and Guan12).
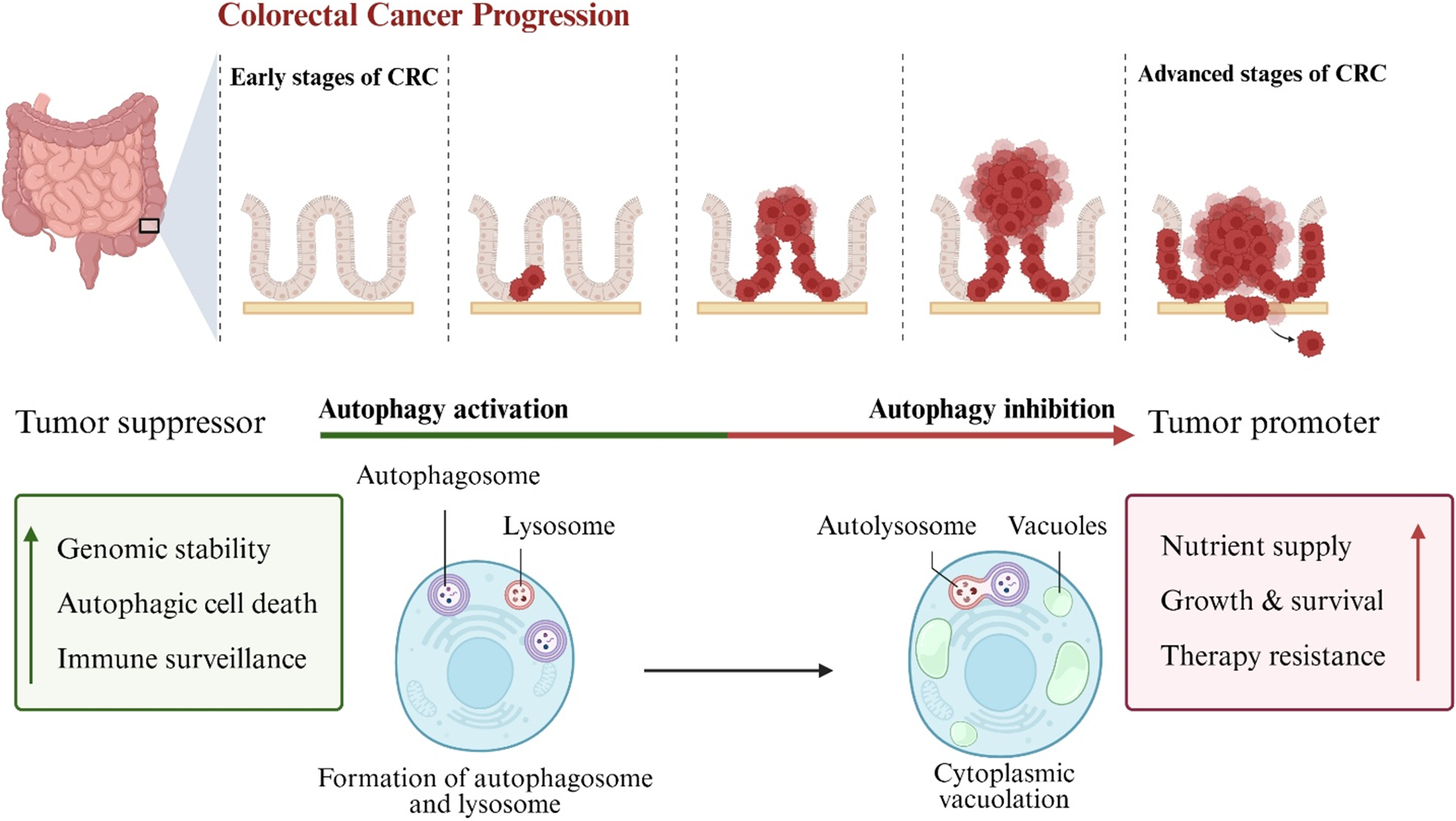
This catabolic process is essential for maintaining cellular homeostasis, but the impact on CRC varies across disease stages. In the early stages of CRC, autophagy functions as a tumour suppressor by maintaining genomic stability, inducing autophagic cell death and enhancing immune surveillance through clearing damaged cellular components (Figure 2) (Ref. Reference Liu and Zhang11). For example, autophagy helps eliminate inflammation-promoting compounds and signals, protecting against cancer onset (Ref. Reference Vafadar, Tajbakhsh, Hosseinpour-Soleimani, Savardshtaki and Hashempur141). Conversely, in advanced CRC, autophagy can promote tumour progression by providing essential nutrients to cancer cells under metabolic stress conditions such as hypoxia and nutrient deprivation, thereby supporting tumour growth, survival and resistance to therapy (Ref. Reference Devenport, Singhal, Radyk, Taranto, Kerk, Chen, Goyert, Jain, das, Oravecz-Wilson, Zhang, Greenson, Chen, Soleimanpour, Reddy, Lyssiotis and Shah142). Elia et al. highlighted how autophagy interacts with the tumour microenvironment (TME), influencing immune responses, hypoxia and oxidative stress. They suggested its modulation as a potential adjuvant strategy to enhance CRC therapy across diverse molecular subtypes (Ref. Reference Elia and Haigis143). The complexity of autophagy is further emphasized by its ability to shape the TME, where it can either suppress or promote tumourigenesis depending on the cellular context, making it a challenging yet promising target for therapeutic intervention (Ref. Reference Maes, Rubio, Garg and Agostinis144).
Schematic overview of the dual role of autophagy in colorectal cancer (CRC) progression. The upper panel illustrates the progression of CRC from early to advanced stages, highlighting tumour growth and invasion. The lower panel depicts the balance between autophagy activation and inhibition in tumour cells. Autophagy activation supports tumour suppression by promoting genomic stability, autophagic cell death and immune surveillance through autophagosome and lysosome formation. Conversely, autophagy inhibition facilitates tumour promotion by enhancing nutrient supply, tumour cell growth and survival and therapy resistance, accompanied by cytoplasmic vacuolation. This dynamic shift in autophagy function reflects its complex role in CRC development and therapy response. Figure created using BioRender.com.

Figure 2. Long description
The diagram is divided into two horizontal sections.
Upper Panel: Colorectal Cancer Progression.
From left to right, it shows an anatomical diagram of the large intestine with a highlighted section of the colon wall. Five stages follow, separated by dashed lines.
1. Early stages of C R C: Normal intestinal crypt structure.
2. Initial tumor growth: A small cluster of red cells appears in the crypt.
3. Intermediate growth: The red tumor cells expand within the crypt.
4. Advanced stages of C R C: A large mass of red tumor cells protrudes outward.
5. Invasion: Tumor cells break through the basement membrane and invade surrounding tissue.
Lower Panel: Autophagy Dynamics.
A central horizontal arrow transitions from green (Autophagy activation) on the left to red (Autophagy inhibition) on the right.
Left Side (Tumor Suppressor):
A green box lists Genomic stability, Autophagic cell death, and Immune surveillance with an upward green arrow. Below this, a cell diagram shows the formation of autophagosome and lysosome. Labels point to a double-membraned autophagosome containing cellular debris and a smaller lysosome.
Right Side (Tumor Promoter):
A red box lists Nutrient supply, Growth and survival, and Therapy resistance with an upward red arrow. A black arrow points from the first cell to a second cell showing cytoplasmic vacuolation. This cell contains an autolysosome (the fusion of an autophagosome and lysosome) and several large, clear vacuoles.
Recent human and animal studies have advanced our understanding of the potential role of autophagy in CRC, with a focus on molecular mechanisms and clinical implications. A single-arm phase II clinical trial evaluated the autophagy inhibitor hydroxychloroquine combined with FOLFOX and bevacizumab in 37 patients with metastatic CRC. The trial demonstrated increased levels of the autophagy marker LC3 and reported an overall response rate (ORR) of 68% with an 11% complete response (CR) rate (Ref. Reference O’Hara, Karasic, Vasilevskaya, Redlinger, Loaiza-Bonilla, Teitelbaum, Giantonio, Damjanov, Reiss, Rosen, Heitjan, Troxel, Amaravadi and O’Dwyer145). In comparison, the study by Saltz et al. (Ref. Reference Saltz, Clarke, Díaz-Rubio, Scheithauer, Figer, Wong, Koski, Lichinitser, Yang, Rivera, Couture, Sirzén and Cassidy146) reported an ORR of 47% for the same regimen of FOLFOX and bevacizumab, and showed that the addition of bevacizumab did not further improve response rates. Although a significant improvement in overall survival was not observed in the hydroxychloroquine trial, the high clinical response rate suggests that targeting autophagy can enhance the efficacy of standard chemotherapy. Sasaki et al. (Ref. Reference Sasaki, Tsuno, Sunami, Kawai, Hongo, Hiyoshi, Kaneko, Murono, Tada, Nirei, Takahashi and Kitayama147) demonstrated that inhibiting autophagy can enhance chemotherapy efficacy, as combining chloroquine with 5-FU in CRC cell lines significantly increased anti-proliferative effects and helped overcome therapy resistance. Qureshi-Baig et al. (Ref. Reference Qureshi-Baig, Kuhn, Viry, Pozdeev, Schmitz, Rodriguez, Ullmann, Koncina, Nurmik, Frasquilho, Nazarov, Zuegel, Boulmont, Karapetyan, Antunes, Val, Mittelbronn, Janji, Haan and Letellier148) demonstrated that hypoxia-induced autophagy drives CRC initiation and progression via the PRKC/PKC-EZR pathway, highlighting its role in tumour adaptation to hypoxic conditions. This study used mouse models to show how autophagy supports tumour growth under stress, with findings suggesting potential targets for intervention. Additionally, Li et al. (Ref. Reference Li, Song, Zhu, Aziz, Zhou, Zhang, Xu, Feng, Chen, Wang and Jin149) demonstrated that targeting metabolism through glutaminolysis inhibition is synergistically enhanced by autophagy inactivation and asparagine depletion. This study revealed a synthetic lethal relationship among these pathways, where CRC cells activate autophagy as a prosurvival mechanism to compensate for metabolic stress induced by glutamine starvation. The simultaneous blockade of these metabolic and recycling routes significantly reduced tumour growth in animal models, highlighting that the therapeutic potential of autophagy modulation is closely linked to the broader metabolic reprogramming of colon cancer cells. Left-sided CRCs associated with CIN and TP53/KRAS mutations may benefit from autophagy-targeted strategies to reduce therapy resistance (Ref. Reference Skarkova, Kralova, Vitovcova and Rudolf150). However, the field faces challenges, including variability in patient responses, the need for effective delivery systems and minimizing off-target effects. These findings confirm autophagy’s role as a therapeutic target, though challenges remain in optimizing clinical applications.
Autophagy research has introduced innovative strategies, such as combining inhibitors like chloroquine with chemotherapy to overcome resistance, offering potential for personalized CRC treatments. However, the dual role of autophagy complicates clinical applications, as limited human trials and variable patient responses present significant hurdles. Further studies are necessary to clarify context-specific effects and to develop targeted modulators that balance autophagy’s tumour-suppressive and tumour-promoting functions.
Interplay among probiotics, autophagy and non-coding RNAs
The core of our hypothesis rests on the existence of a dynamic mechanistic crosstalk where probiotics and their metabolites serve as upstream cues that reprogramme the ncRNA landscape to precisely calibrate autophagic flux. While previous studies have often examined these components in isolation, we propose an integrated signalling axis where ncRNAs act as the functional bridge between microbial interventions and the cellular degradative machinery.
Direct evidence supports the modulation of autophagy by probiotics in CRC. Probiotics and their metabolites, such as SCFAs, regulate protective autophagy, contributing to antitumour effects through a complex network, as reviewed by Sadeghloo et al. (Ref. Reference Sadeghloo, Nabavi-Rad, Zali, Klionsky and Yadegar151). Several studies have highlighted that probiotics can induce or inhibit autophagy, leading to antitumour effects. For instance, a surface-layer protein derived from Lactobacillus acidophilus NCFM induced autophagic cell death in HCT116 human colon cancer cells, as evidenced by increased expression of Beclin-1 and LC3-II, alongside degradation of p62. This modulation suppressed cell proliferation, suggesting a mechanism for reducing CRC cell viability (Ref. Reference Wang, Cheng, Zhang, Xu, Zhang and Lu152). Similarly, native strains of Lactobacillus and Bifidobacterium, including Bifidobacterium bifidum 1001, Bifidobacterium longum 1044, B. infantis 1015 and 1063, L. plantarum 42, Lacticaseibacillus rhamnosus 195, L. brevis 205 and Limosilactobacillus reuteri 100, upregulated autophagy-related genes such as PIK3C3, ATG14, Beclin-1, PIK3R4, ATG5, ATG16, ATG7 and ATG3 in HT-29 human colon cancer cells, exerting anti-inflammatory effects under inflammatory conditions (Ref. Reference Jouriani, Torkamaneh, Torfeh, Ashrafian, Aghamohammad and Rohani153). In contrast, metabolites from L. plantarum KCTC 3108 inhibited autophagy in Caco-2 human colonic cells by downregulating ATG9A, ATG16L1, ATG5, Beclin-1 and LC3 I/II, which correlated with decreased cell viability and potential anticancer effects (Ref. Reference Jeong, Kim, Park, Lee, Lee, Hlaing, Yoo, Rhee and Im154). Animal models further corroborate these findings, demonstrating therapeutic benefits in CRC. Ligilactobacillus salivarius LZZAY01, administered in a colitis-associated cancer (CAC) model using AOM/DSS-treated C57BL/6j mice, induced autophagy through elevated Beclin-1 and LC3II/I expression, leading to reduced tumour numbers, suppressed colon cancer cell growth and improved gut microbiota. Complementary in vitro experiments with CT-26, HCT-116 and SW-620 cell lines reinforced these observations by showing enhanced autophagy and apoptosis (Ref. Reference Yang, Li, An, Chen, Zhao, Cui, Zhang, Lu, Tian, Bao and Zhao155).
Similarly, probiotics directly influence the expression of ncRNAs, which regulate autophagy and thereby affect CRC development and progression (Refs Reference Garavaglia, Vallino, Ferraresi, Visciglia, Amoruso, Pane, Munteanu and Isidoro68, Reference Soudha, Prasad and Panicker156). Evidence from cellular models indicates that probiotic metabolites can reprogramme ncRNA expression profiles in CRC cells. In one study, supernatants derived from L. acidophilus and Saccharomyces cerevisiae enhanced the levels of tumour-suppressor miRNAs (miR-34, miR-135, miR-25, miR-16, miR-195, miR-27, miR-98 and let-7) while reducing oncogenic miRNAs (miR-9, miR-106b, miR-17, miR-21, miR-155 and miR-221) in HT-29 and SW-480 human colon cancer cells. This dual modulation of ncRNAs was associated with diminished cellular proliferation and suggested a possible reversal of CRC progression (Ref. Reference Saffar, Larypoor and Torbati157). Similarly, Lactobacillus casei downregulated miR-21 in HT-29 human colon cancer cells, inducing a time- and dose-dependent apoptosis (Ref. Reference Hosseinzadeh, Bahadori, Ghorbani and Mohammadimehr158). In another study, Leuconostoc mesenteroides downregulated miR-21 and miR-200b in HT-29 cells, promoting apoptosis and hindering inflammation and cell survival (Ref. Reference Vahed, Barzegari, Saadat, Goreyshi and Omidi159). Administration of B. longum in a murine model of colitis-associated CRC induced by DSS and DMH resulted in downregulation of oncogenic miRNAs (miR-21a and miR-155) and upregulation of tumour suppressor miRNAs (miR-145 and miR-15a). These molecular changes were associated with reduced aberrant crypt foci and increased necrosis and fibrosis in colonic tissue (Ref. Reference Fahmy, Gamal-Eldeen, El-Hussieny, Raafat, Mehanna, Talaat and Shaaban160). Likewise, L. acidophilus and B. bifidum downregulated miR-135b and miR-155 while upregulating miR-26b and miR-18a in an AOM-induced colon cancer model in BALB/c mice, contributing to improved colon cancer treatment outcomes (Ref. Reference Heydari, Rahaie, Alizadeh, Agah, Khalighfard and Bahmani161). In a randomized clinical trial involving patients with non-metastatic rectal cancer, Khodaii et al. (Ref. Reference Khodaii, Mehrabani Natanzi, Khalighfard, Ghandian Zanjan, Gharghi, Khori, Amiriani, Rahimkhani and Alizadeh162) reported that supplementation with L. acidophilus positively modulated the lncRNA–miRNA–mRNA regulatory network. Specifically, the probiotic consumption upregulated tumour suppressor miRNAs (miR-548 ac, miR-378a, miR-34a, miR-601 and miR-133b) and tumour suppressor lncRNAs (LincRNA-P21). It also downregulated oncogenic miRNAs (miR-21, miR-20a, miR-20b, miR-424, miR-1244, miR-135b and miR-224) and lncRNAs (CCAT1, LOC152578, UCA1, CRNDE, PVT1, MALAT1, XLOC_000303, XLOC_006844, BCAR4 and HOTAIR). These findings suggest potential novel approaches for disease monitoring and therapy.
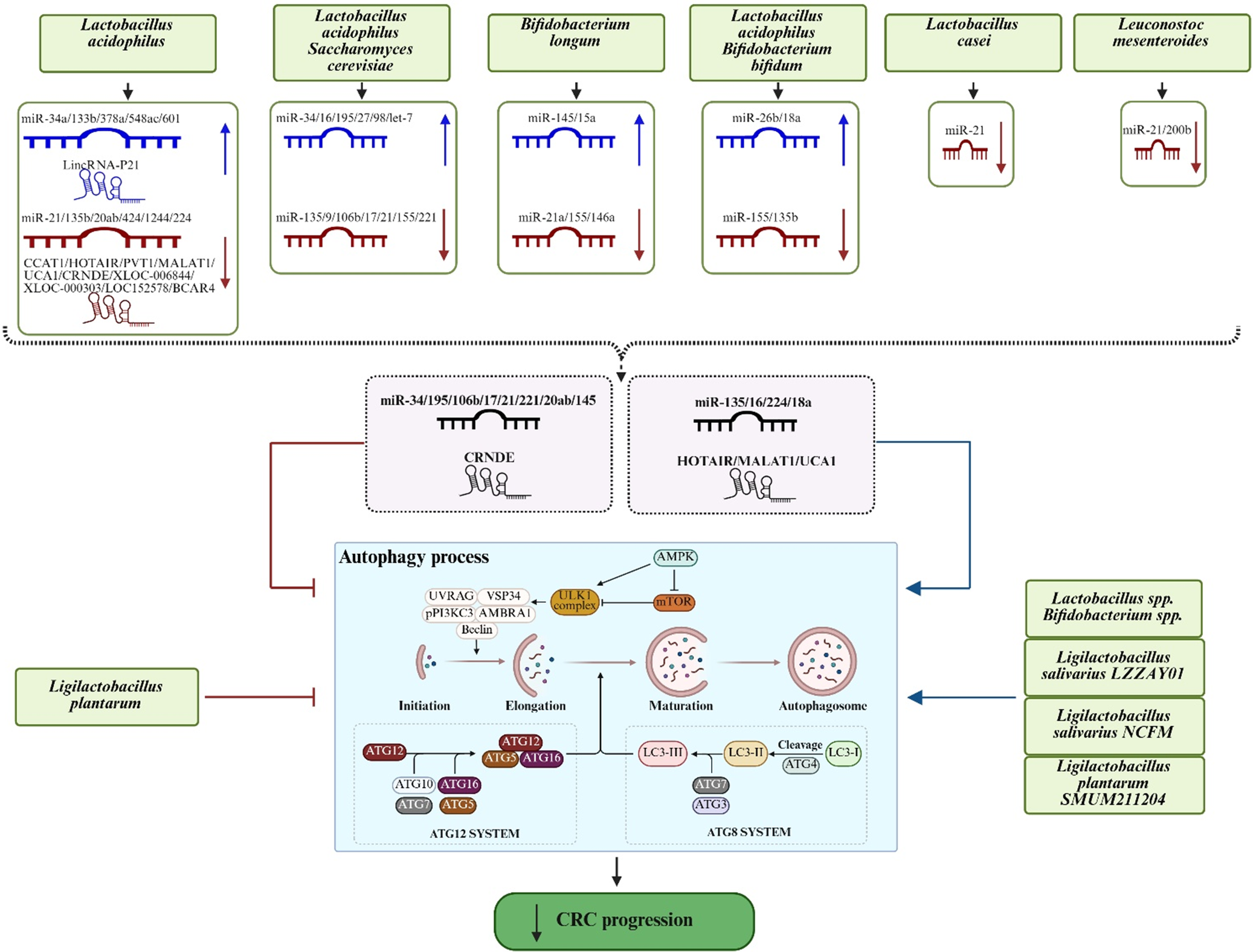
Building on this collective evidence, we hypothesize that probiotics exert a multi-layered influence on CRC evolution by acting through diverse molecular levels. Beyond direct modulation of autophagy-related pathways, probiotics may indirectly reprogramme the cellular landscape by reshaping ncRNA networks – including miRNAs that regulate antioxidant enzymes – which in turn fine-tune autophagic flux and metabolic redox balance. We propose that this integrated microbial–epigenetic–autophagic axis governs tumour dynamics and therapy resistance, where ncRNAs serve as versatile rheostats translating microbial signals into systemic cellular responses (Figure 3 and Table 2). While few studies have simultaneously mapped all three components, the functional overlap between probiotic-targeted ncRNAs and the regulatory machinery of both oxidative stress and autophagy provides a compelling conceptual foundation for this expanded mechanism.
Conceptual framework illustrating the interplay among probiotics, ncRNAs and autophagy in CRC. Solid arrows indicate reported effects of probiotics on autophagy or ncRNA expression. The dashed arrows highlight a proposed pathway whereby probiotics may indirectly modulate autophagy through ncRNA regulation. The ncRNAs shown in the dashed box represent molecules that have been reported both to be regulated by probiotics and independently implicated in autophagy regulation in CRC. Figure created using BioRender.com.

Figure 3. Long description
The diagram is organized into four vertical levels.
1. Top Level: Six green boxes list probiotic species including Lactobacillus acidophilus, Saccharomyces cerevisiae, Bifidobacterium longum, Bifidobacterium bifidum, Lactobacillus casei, and Leuconostoc mesenteroides. Arrows point down to specific n c R N A regulation panels.
2. Second Level: Panels show blue (upregulated) and red (downregulated) m i R N A and linc R N A strands. For example, Lactobacillus acidophilus upregulates m i R-34 a and Linc R N A-P 21 while downregulating m i R-21 and C C A T 1. A dashed box in the center contains m i R-34, m i R-135, C R N D E, and H O T A I R, representing molecules regulated by probiotics that also impact autophagy.
3. Third Level: A large central box titled Autophagy process.
- Initiation and Elongation: Shows complexes involving U V R A G, V S P 34, Beclin, and A M B R A 1.
- Maturation and Autophagosome: Shows the transition from L C 3-I to L C 3-I I.
- Regulatory Systems: Below the main flow are the A T G 12 S Y S T E M (A T G 12, A T G 5, A T G 16, A T G 10, A T G 7) and A T G 8 S Y S T E M (L C 3, A T G 4, A T G 7, A T G 3).
- External Modulators: Ligilactobacillus plantarum is shown with inhibitory lines toward the autophagy box, while other species like Ligilactobacillus salivarius point to it with activating blue arrows.
4. Bottom Level: A final green oval with a downward arrow indicates that these combined effects lead to a decrease in C R C progression.
Emerging evidence supports this hypothesis, as several tumour-suppressor ncRNAs upregulated by probiotics modulate autophagy in CRC. For instance, miR-34a inhibits autophagy by targeting key regulators, reducing autophagic flux, decreasing EMT and heightening sensitivity to chemotherapeutics like 5-FU in CRC cells (Ref. Reference Huang, Kaller and Hermeking163). Probiotic-induced elevation of miR-34a, as observed in clinical trials, may thus suppress protective autophagy in advanced CRC, promoting apoptosis and limiting tumour proliferation. Similarly, miR-145 can target histone deacetylase 4 (HDAC4) to activate p53, thereby inhibiting autophagy and enhancing 5-FU sensitivity via the ATF4/miR-145/HDAC4/p53 axis (Ref. Reference Zhao, Chen, Zhang, Ma, Hu and Xu164). MicroRNA-16-5p, upregulated by probiotics, suppresses proliferation and angiogenesis in CRC by targeting FOXK1 and inhibiting the PI3K/Akt/mTOR pathway, potentially inducing tumour-suppressive autophagy (Ref. Reference Huang, Xu, Ke, Pan, Ai, Xie, Lan, Hu and Wu165). The miR-18a, elevated in probiotic-treated models, promotes autophagy through upregulation of ataxia telangiectasia mutated (ATM) and inhibition of mTORC1, facilitating autophagolysosomal degradation of oncogenic proteins like heterogeneous nuclear ribonucleoprotein A1 (hnRNP A1) and inducing apoptosis in CRC cells (Ref. Reference Qased, Yi, Liang, Ma, Qiao and Liu166). These tumour-suppressive miRNAs suggest that probiotics may fine-tune autophagy to impair CRC cell survival and resistance mechanisms.
Conversely, probiotics downregulate a range of oncogenic ncRNAs involved in autophagy regulation, which could potentially counteract their autophagy-promoting effects and contribute to the inhibition of CRC progression. For instance, suppression of miR-135 prevents the activation of protective autophagy through the MUL1/ULK1 pathway, potentially reducing oxaliplatin resistance (Ref. Reference Wang, Wang, Zhang, Deng, Liu, Liu, Li, Bai, Ning, Wang, Ge and Ba167). The miR-106b, an autophagy inhibitor targeting ATG16L1 and ATG12, is suppressed by probiotics, which could restore autophagic activity in a context-dependent manner to prevent tumour adaptation (Ref. Reference Zhai, Wu, Chuang and Kwon168). Oncogenic miR-21 forms an autoregulatory loop with vacuole membrane protein-1 (VMP1) via the PTEN/AKT/TFEB pathway, inhibiting autophagy (Ref. Reference Wang, Peng, Zeng, Zhang, Liu, Jiang, Lu and Zou169); however, probiotic-mediated reduction of miR-21 may alleviate this suppression, redirecting autophagy towards antitumour effects. The miR-221, targeted for downregulation by probiotics, could inhibit autophagy by binding tumour protein 53-induced nuclear protein 1 (TP53INP1), and its reversal could enhance autophagic cell death (Ref. Reference Liao, Li, Ye, Zeng, Li, Pu, Ding, He and Huang170). Other oncogenic miRNAs, such as miR-20a and miR-224, may exhibit contrasting effects on autophagy. The miR-20a is inhibited via the ATG5/FIP200 pathway under hypoxic conditions, whereas miR-224 is induced through the p53/ULK2 axis. Accordingly, their attenuation could diminish autophagy-mediated tumour progression (Refs Reference Che, Wang, Huang, Zhang, Zhao, Zhang and Yuan171, Reference Le Mei, Zheng, Liu, Li, Lim, Qi, Zhao and Wang172).
Among lncRNAs, probiotic-induced downregulation of HOTAIR, MALAT1, UCA1 and CRNDE may also contribute to therapeutic benefits. Reduced HOTAIR disrupts its sponging of miR-93, leading to decreased ATG12 expression and autophagy inhibition, thereby improving radiosensitivity (Ref. Reference Liu, Chen, Chen, Liu, Gu, Fan and Ge173). Lower MALAT1 levels free miR-101, limiting autophagy activation (Ref. Reference Si, Yang, Ge, Yu, Yao, Sun, Ren and Ding174). Similarly, knockdown of UCA1 and CRNDE attenuates autophagy through AKT/mTOR suppression, enhancing apoptosis and drug sensitivity (Refs Reference Yang, Wang, Tao, Li, Cao and Yang175, Reference Song, Li, Liang, Zhuo, Wang and Dai176).
The synergistic interplay among probiotics, autophagy and ncRNAs emerges as a pioneering framework for CRC therapy, enabling a multifaceted approach to modulating tumour dynamics through integrated regulation of immune responses, epigenetic modifications and cellular homeostasis. The unique potential of this triad lies in its ability to bridge microbial influences with molecular and cellular processes, offering avenues to enhance overall treatment efficacy and overcome complex resistance mechanisms in CRC. However, key limitations persist, including the underrepresentation of human clinical evidence, inconsistencies in microbial and molecular responses across patient demographics and technical hurdles in ensuring targeted integration without unintended systemic impacts. To realize this potential, rigorous, large-scale clinical investigations are imperative to delineate optimal combinations and dosages, while exploring how this triad can be adapted to varying CRC profiles. Such advancements could redefine precision oncology, raising intriguing questions about how these interactions might extend to other cancers or personalized regimens.
Conclusion
The evidence synthesized in this review establishes probiotics as pivotal upstream regulators within a complex mechanistic crosstalk involving ncRNAs and autophagy in CRC. By systematically reprogramming ncRNA expression profiles and subsequently recalibrating autophagy-related genes, probiotics exert a multi-layered influence on tumour proliferation, inflammation and chemoresistance. These interconnected signalling networks suggest that probiotics do not merely act as supplements but as orchestrators of the cellular degradative machinery, providing a robust molecular rationale for their integration into precision-based CRC management.
Future research should transition from descriptive studies to mechanistically driven clinical trials that evaluate the synergy between specific probiotic strains and standard chemotherapeutic agents. A primary focus should be on deciphering the ncRNA–autophagy interface to identify novel, non-invasive biomarkers that can predict treatment response. Furthermore, stratifying patient cohorts based on CMS will be essential to optimize probiotic protocols and address inter-individual variability. Unlocking the full potential of this microbial–epigenetic–autophagy axis will pave the way for a new era of personalized oncology, where integrative therapies are tailored to the unique molecular landscape of each CRC patient.
Abbreviations
- AMPK
-
AMP-activated protein kinase
- APC
-
adenomatous polyposis coli
- ATG
-
autophagy-related gene/protein
- ATM
-
ataxia telangiectasia mutated
- Bax
-
Bcl-2-associated X protein
- Bcl-2
-
B-cell lymphoma 2
- CIMP
-
CpG island methylator phenotype
- CIN
-
chromosomal instability
- CMS
-
consensus molecular subtypes
- CRC
-
colorectal cancer
- DMH
-
1,2-dimethylhydrazine
- EGFR
-
epidermal growth factor receptor
- FOLFOX
-
5-fluorouracil, leucovorin, and oxaliplatin (chemotherapy regimen)
- FOXK1
-
forkhead box protein K1
- G6PD
-
glucose-6-phosphate dehydrogenase
- GIQLI
-
Gastrointestinal Quality of Life Index
- GLUT1
-
glucose transporter 1
- HDAC
-
histone deacetylase
- HDACi
-
histone deacetylase inhibitors
- IFN
-
interferon
- IL
-
interleukin
- LC3
-
microtubule-associated proteins 1A/1B light chain 3B
- lncRNAs
-
long non-coding RNAs
- MALAT1
-
metastasis associated lung adenocarcinoma transcript 1
- MAPK/ERK
-
mitogen-activated protein kinase/extracellular signal-regulated kinase pathway
- MHC-I
-
major histocompatibility complex class I
- miRNAs
-
microRNAs
- MSI
-
microsatellite instability
- mTOR
-
mammalian target of rapamycin
- ncRNAs
-
non-coding RNAs
- NRP-1
-
neuropilin-1
- PD-L1
-
programmed death-ligand 1
- PI3K
-
phosphoinositide 3-kinase
- PRC2
-
polycomb repressive complex 2
- SCFAs
-
short-chain fatty acids
- SMAD
-
mothers against decapentaplegic homolog
- SQSTM1
-
sequestosome 1 (p62)
- TGFβ
-
transforming growth factor beta
- TNF-α
-
tumour necrosis factor alpha
- ULK1
-
unc-51 like autophagy activating kinase 1
- VEGF
-
vascular endothelial growth factor
- VMP1
-
vacuole membrane protein-1
- Wnt
-
wingless-related integration site (signalling pathway)
- ZO-1/2
-
zonula occludens-1/2.
Data availability statement
No datasets were generated or analysed during the current study.
Acknowledgements
We used ChatGPT-5 to assist with paraphrasing and improving the clarity of the English language in this manuscript; no scientific content was generated by the tool.
Author contribution
N.F. prepared the initial draft manuscript and contributed ideas for the design of figures and tables. Z.S. conceptualized the study, contributed to manuscript writing and critically revised the final version. A.M.A. provided critical manuscript editing and offered key suggestions for figures and tables. A.S. assisted with manuscript editing and revisions. All authors have seen and approved the final version of the manuscript and the author list.
Funding statement
The authors received no funding for this article.
Competing interests
The authors declare that they have no competing interests.

 Open access
Open access