



Social media summary
Purebred dogs show different dependence on human behaviour and improvement during social learning.
1. Introduction
Dogs are domesticated animals that have very successfully inhabited an anthropogenic niche. They are present in every culture (Serpell, Reference Serpell2017), reaching a staggering 1 billion-strong population worldwide, according to some estimations (Coppinger & Coppinger, Reference Coppinger and Coppinger2021). Although most of them are ‘free-ranging dogs’ (Range & Marshall-Pescini, Reference Range and Marshall-Pescini2022b), even these animals rely on humans for food and shelter (Boitani et al., Reference Boitani, Ciucci and Ortolani2007). However, people who live in the WEIRD (‘Western, Educated, Industrialized, Rich, Democratic’) countries (Chira et al., Reference Chira, Kirby, Epperlein and Bräuer2023) are more familiar with companion and working dogs (Sykes et al., Reference Sykes, Beirne, Horowitz, Jones, Kalof, Karlsson, Larson, Litwak, McDonald, Murphy, Pemberton, Promislow, Rowan, Stahl, Tehrani, Tourigny, Wynne, Strauss and Larson2020) that clearly belong to someone (the ‘owner’, or an organization). Dogs who are kept for their services, or for leisure activities (‘family dogs’), can be considered as the ‘perfect companion’ for humans in a biological/evolutionary sense (Pongrácz & Dobos, Reference Pongrácz and Dobos2023). They can initiate and maintain mutually enjoyable interactions with their owners (Bradshaw et al., Reference Bradshaw, Pullen and Rooney2015); they do not go feral easily (i.e., they do not need to be restricted to stay with us – Boitani & Ciucci, Reference Boitani and Ciucci1995), and after proper socialization, dogs express their inherited capacity to show these behavioural features (Miklósi & Topál, Reference Miklósi and Topál2013).
From an ethological point of view, the most interesting aspect of dogs’ behaviour is those socio-cognitive traits that enabled them to maintain their human-dependent status through the formation of a beneficial relationship and cooperation with humans (Range & Marshall-Pescini, Reference Range and Marshall-Pescini2022a; Topál et al., Reference Topál, Miklósi, Gácsi, Dóka, Pongrácz, Kubinyi and Csányi2009b). The most often mentioned fundamental synchronizing mechanisms (Kubinyi et al., Reference Kubinyi, Viranyi and Miklósi2007; Topál et al., Reference Topál, Miklósi, Gácsi, Dóka, Pongrácz, Kubinyi and Csányi2009b) between dogs and humans are dog-owner attachment (Topál et al., Reference Topál, Miklósi, Csányi and Dóka1998), ability to communicate both as receivers and signallers (e.g., Kaminski & Nitzschner, Reference Kaminski and Nitzschner2013; Pongrácz, Reference Pongrácz2017), rule learning (Hertel et al., Reference Hertel, Kaminski and Tomasello2014; Topál et al., Reference Topál, Gergely, Erdőhegyi, Csibra and Miklósi2009a), and social learning (Pongrácz, Reference Pongrácz, Kaminski and Marshall-Pescini2014). At the same time, several studies highlight the role of proximate factors that fine-tune dogs’ human-compatible behaviours, such as the effects of upbringing (Harvey et al., Reference Harvey, Craigon, Blythe, England and Asher2016), training (Lazarowski et al., Reference Lazarowski, Singletary, Rogers and Waggoner2021), or housing (D’Aniello & Scandurra, Reference D’Aniello and Scandurra2016). To make the picture even more complex, in addition to the species-level genetic effects of domestication and the individual-level effects of ontogeny and learning, dogs have undergone specific processes of artificial selection, which resulted in landraces and dog breeds (e.g., Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017; Talenti et al., Reference Talenti, Dreger, Frattini, Polli, Marelli, Harris, Crepaldi, Cocco, Hogan, Bigi, Caniglia, Parker, Pagnacco, Ostrander and Crepaldi2018). The hundreds of morphologically and often functionally different dog breeds can offer a unique opportunity to understand the factors and pathways of selection behind highly variable dog behaviour.
Dog breeds offer an obvious grouping variable to researchers when they investigate various cognitive (e.g., Gnanadesikan et al., Reference Gnanadesikan, Hare, Snyder-Mackler and MacLean2020) and behavioural phenotypes (e.g., Serpell & Duffy, Reference Serpell and Duffy2014). Morrill et al. (Reference Morrill, Hekman, Li, McClure, Logan, Goodman, Karlsson, Dong, Alonso, Carmichael, Snyder-Mackler, Alonso, Noh, Johnson, Koltookian, Lieu, Megquier, Swofford, Turner-maier and Karlsson2022) found no evidence for behavioural genotypes that would be unique in any dog breed; however, they described more general behavioural traits (e.g., ‘biddability’) that had detectable genetic background in breeds that were selected for close cooperation with humans (e.g., retrievers, sheepdogs). Although, according to our current knowledge, there is no exclusive genotype behind any dog breed’s behavioural traits, the various behaviours can be expressed with different intensities across dog breeds (e.g., Mehrkam & Wynne, Reference Mehrkam and Wynne2014). There are several publications that assessed dozens of the most commonly available dog breeds regarding such traits as human-directed aggressive behaviour (Mikkola et al., Reference Mikkola, Salonen, Puurunen, Hakanen, Sulkama, Araujo and Lohi2021); problem solving (Junttila et al., Reference Junttila, Valros, Mäki, Väätäjä, Reunanen and Tiira2022), boldness (Starling et al., Reference Starling, Branson, Thomson and McGreevy2013), or trainability (Serpell & Hsu, Reference Serpell and Hsu2005). As useful as these explorative studies can be in the detection of some of the targeted behaviours across large databases (e.g., Asp et al., Reference Asp, Fikse, Nilsson and Strandberg2015), convenience sampling of dog breeds usually suffers from poor explanatory power regarding the found behavioural variance. These surveys necessarily target the most popular dog breeds at a given location and time (e.g., Tonoike et al., Reference Tonoike, Nagasawa, Mogi, Serpell, Ohtsuki and Kikusui2015), without the opportunity of forming biologically relevant hypotheses (for a review, see Pongrácz & Dobos, Reference Pongrácz and Dobos2025a).
When researchers directly focus on the behaviour of a few, ‘handpicked’ dog breeds, these can be chosen as the representatives of a larger group of breeds (three breeds, according to their skull shape: Ujfalussy et al., Reference Ujfalussy, Bognár, Molnár, Miklósi and Kubinyi2023; and another three breeds, according to their prey-related behaviour: Udell et al., Reference Udell, Ewald, Dorey and Wynne2014). A serious limitation of these studies originates from the skewed sampling of breeds, because the delicate but often significant breed-typical characteristics of behaviour can make it difficult to extrapolate the results to represent a whole group of breeds.
As a solution to the aforementioned problems, ecologically valid investigation of breed-based behavioural comparisons can be organized around overarching grouping categories (Pongrácz & Dobos, Reference Pongrácz and Dobos2025a), such as ancestry (the genetic distance from the hypothetical common ancestor of dogs), or functional selection (which resulted in a few main types of dog breeds, such as cooperative or independent working dogs). Based on the between-breed genetic distance cladograms (e.g., Dutrow et al., Reference Dutrow, Serpell and Ostrander2022; Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017), researchers found differences between the so-called ‘primitive’ and more derived breed clades’ behavioural phenotypes (e.g., Hansen Wheat et al., Reference Hansen Wheat, Fitzpatrick, Rogell and Temrin2019). Regarding the effects of functional breed selection, cooperative and independent working dog breeds were found to be differently responsive to human attention (Heberlein et al., Reference Heberlein, Manser and Turner2017; Pongrácz & Lugosi, Reference Pongrácz and Lugosi2024) and communication (Dobos et al., Reference Dobos, Lugosi and Pongrácz2025; Dobos & Pongrácz, Reference Dobos and Pongrácz2024; Gácsi et al., Reference Gácsi, McGreevy, Kara and Miklósi2009). Furthermore, the two functional breed types were found to utilize differently the opportunity to learn from humans (Dobos & Pongrácz, Reference Dobos and Pongrácz2023) and conspecific demonstrators (Lugosi et al., Reference Lugosi, Udvarhelyi-tóth, Dobos and Pongrácz2024). Ethological experiments based on both ancestry-based and functional clustering of dog breeds provided solid and ecologically valid explanations for the phenotypical variability of dogs’ behaviour. However, if we are interested in the more specific characteristics of, and differences between, dog breeds’ behaviour, the previously mentioned grouping systems may fail to provide a fine enough resolution for the researcher.
Ancestry and work function-based groupings necessarily blend finer details of dogs’ behaviour that otherwise may cause noticeable differences among those breeds that were sorted together into a given genetic clade, or functional breed type (Pongrácz & Dobos, Reference Pongrácz and Dobos2025a). For example, many sighthound breeds (such as the Greyhound, Whippet, or Borzoi) and herding breeds that originate from the British Isles (such as the Border Collie, Sheltie, Collie), are in close genetic relationship (Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017); however, they were bred for very different functions (to pursue and kill the fleeing prey on their own vs. herding/directing/collecting livestock on command without harm). Gundogs (such as the retrievers, pointers, spaniels, and setters), and herding dogs of various geographical origins, belong to multiple separate genetic clades (Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017); however, they have been similarly selected for working in close cooperation with their handlers (Gácsi et al., Reference Gácsi, McGreevy, Kara and Miklósi2009). Still, their actual tasks show vast differences (i.e., gundogs signal, flush, and retrieve game to the hunter, while herding dogs move livestock). As there are dozens of highly refined and task-specific working dog sub-types, it is also expected that their human-directed behavioural traits would be different (e.g., Pongrácz et al., Reference Pongrácz, Miklósi, Vida and Csányi2005). It is therefore likely that we would miss something important regarding dog breeds’ behaviour when we apply one of the overarching and otherwise biologically relevant clustering systems (e.g., ancestry-based, or functional selection-related groups).
If the goal is to investigate the behavioural characteristics of several dog breeds, and we would like to avoid the loss of finer details because of the prefixed clustering by function or ancestry, while at the same time we would like to apply ecologically valid hypotheses, we would need a different approach. Accordingly, those investigations could provide a solution, which established dimensions (or ‘traits’) based on several behavioural elements and then used these dimensions to sort large numbers of subjects into clusters or place them along scales/axes. For example, regarding dog-owner attachment, Topál et al. (Reference Topál, Miklósi, Csányi and Dóka1998) created such clusters that described the various attachment styles of dogs, like those seen in the case of human infant-mother attachment. Ben-Michael et al. (Reference Ben-Michael, Vossen, Felling and Peters1997) used a questionnaire with owner-reported problematic dog behaviours, and with hierarchical cluster analysis, they created eight types of dogs with behavioural problems. The investigation of personality traits in dogs also uses a behaviour-based approach, in which the subjects can eventually be characterized with scores from the various trait scales (e.g., Salonen et al., Reference Salonen, Mikkola, Niskanen, Hakanen, Sulkama, Puurunen and Lohi2023). Some researchers used a similar approach to create breed behavioural profiles, based on dimensions such as ‘trainability’, ‘boldness’, ‘reactivity’ and ‘aggression’ – resulting from questionnaires (Hart & Hart, Reference Hart and Hart1985; Turcsán et al., Reference Turcsán, Kubinyi and Miklósi2011). Finally, the possibly largest and longest ongoing endeavour of behaviour-based assessment of breed and breeding-line profiling is the Swedish Dog Mentality Assessment (DMA), which resulted in several personality trait-like dimensions that describe the general suitability of dogs for various working tasks (e.g., Svartberg, Reference Svartberg2006; Svartberg & Forkman, Reference Svartberg and Forkman2002).
In our current research, we aimed to develop a novel, behaviour-based grouping system for dogs that would be biologically relevant for a better understanding of the background of dog-human interactions. Our goal was to use a novel approach that could help us to organize and understand the subtleties of many dozens of working dog breeds’ behaviour, independent of their functional or ancestry-based designations. Our chosen model scenario was social learning from a human demonstrator during a spatial problem-solving task (i.e., in a detour test, for a review, see Pongrácz & Dobos, Reference Pongrácz and Dobos2025c). This task has been proven to be an especially fruitful behavioural paradigm for the investigation of dog-human interactions in a communication- and attention-based context (e.g., Dobos & Pongrácz, Reference Dobos and Pongrácz2023, Reference Dobos and Pongrácz2024; Pongrácz & Dobos, Reference Pongrácz and Dobos2025b). We hypothesized that, partly depending on their breed-specific characteristics, dogs will show different levels of attention (which is considered to be an important factor in social learning – Dobos & Pongrácz, Reference Dobos and Pongrácz2024; Pongrácz et al., Reference Pongrácz, Miklósi, Timár-geng and Csányi2004), and improvement of efficiency (which is considered to be the main measure of the effect of social learning). We predicted that apart from the effect of their ancestry and functional selection, the more specific breed-typical behavioural traits would also influence their reliance and dependence on human behaviour in an observational learning scenario.
2. Materials and methods
2.1. Ethics approval and consent to participate
We conducted the research in accordance with the Hungarian regulations on animal experimentation and guidelines for use of animals in research, as outlined by the Association for the Study of Animal Behaviour. The fully reward-based and non-invasive methodology was reviewed and approved by the Animal Welfare Committee of Eötvös Loránd University (Certificate number ELTE-AWC-013/2023). Companion dogs were always tested in the presence of their owners. Informed consent was obtained from the dog owners before involving their dogs in the study. We informed the owners that they could interrupt or stop the experiments at any time they felt that their dog experienced high levels of stress. (This never happened.)
2.2. Dog participants
For this study, we analysed the data of N = 187 privately owned, adult, purebred dogs. At the time of their testing, dogs had to be older than 1 year, and they belonged to the so-called working breeds, which were originally selected for various performance tasks (e.g., hunting, herding, livestock guarding, sledge pulling, etc.). We did not test any ‘toy’ breeds, which had no other purpose besides being a companion animal (e.g., Chihuahua, Bichon Havanese). Breed functions were assessed based on the original breed standards. The subjects were originally tested in one of the following three studies: (Dobos & Pongrácz, Reference Dobos and Pongrácz2023, Reference Dobos and Pongrácz2024; Pongrácz and Veres, Reference Pongrácz and Veres2025). For this study, apart from the above-mentioned demographic measures, the selection of dogs from the unified subject pool of previous studies was based on the following criteria: (1) the subject had to be tested with a V-shaped fence (thus, we excluded those subjects who were tested with a straight barrier in Pongrácz and Veres, 2025); (2) the subject had to be tested with a human demonstrator (thus, we excluded the ‘no-demo’ control subjects from Dobos and Pongrácz (Reference Dobos and Pongrácz2023) and Pongrácz and Veres (2025)); and (3) the demonstration had to happen with ostensive verbal attention eliciting by the experimenter (thus, we excluded the subjects who received ‘neutral intonation’ demonstration in Dobos and Pongrácz (Reference Dobos and Pongrácz2024)).
2.3. Testing method in brief (further details are given in Dobos & Pongrácz, Reference Dobos and Pongrácz2023)
Here we only describe those details of the original experiments, which are relevant from the aspect of the current research. A video clip helps to visualize the experimental procedure: https://doi.org/10.6084/m9.figshare.31839121
Dogs were tested outdoors in a spatial (detour) task, where we provided a demonstration of the task to see whether the various working dog breeds would learn via observation from the experimenter. All tests were conducted between September 2022 and December 2023. The demonstrators were two young (21 years old) women, who performed as demonstrators in the case of N = 78 and N = 109 subjects, respectively.
The experimental device was a V-shaped transparent fence (both ‘wings’ were 3 m long and 1 m high), made from a light steel frame with wire mesh stretched across it. The fence was firmly inserted into the ground; thus, dogs could not dig under it (Supplementary Figure S1). The angle of the fence was set to 80 degrees.
Each dog was tested only once in a single session of three trials. Companion dogs were recruited via social media advertisements, and they arrived at the testing location with their owners. We explained the testing protocol to the owners, and they gave their informed consent for the handling of the collected experimental data. The tests were video recorded for later data extraction.
Each test consisted of three 1-min-long trials. In each trial, the dog had to obtain the reward from the inner corner of the V-shaped fence. In most cases, the reward was the favourite food of the dog, based on the advice of the owner, for the best motivation effect; however, in a very few cases, the dog could only be motivated with toys, so we used the favourite toy as a reward in these instances. To succeed in this task, the dog had to detour the fence from the outside. At the beginning, the dog and its owner positioned themselves at the starting point, which was 2 m from the tip of the fence’s corner. The owner kept the dog at the starting point by holding its collar or harness. In Trial 1, the experimenter conspicuously placed the reward at the inner corner by leaning over the fence from the front. Then the experimenter returned to the starting point and stood one step behind the owner. On her signal, the owner released the dog, who had maximum 1 min to obtain the reward. Whether the dog detoured the fence successfully or failed to do so, it was returned to the starting point, and the next trial started.
At the beginning of Trials 2 and 3, the experimenter demonstrated the detour, while the owner stayed with the dog at the starting point. Throughout her walk around the fence, the experimenter kept the dog’s attention on herself by showing the reward in her hand and saying the dog’s name, as well as attention-grabbing phrases (such as ‘Look at me!’, ‘Here I go!’) in an ostensive tone. When the experimenter reached the inner corner, she conspicuously placed the reward again on the ground, then showed her empty hands towards the dog; finally, she walked out from behind the fence along the other wing, then returned to the starting point. After her arrival, the dog was released, and it could attempt to obtain the reward. The experimenter did the demonstration from the opposite direction in Trial 2 and Trial 3. The owners were allowed to encourage their dogs during each trial, but they had to stay at the starting point, and we disallowed hand and verbal signals, which would indicate/encourage the dog how it should perform the detour.
2.4. Behavioural coding
Based on the video footage, behavioural elements were extracted with BORIS software (© Olivier Friard and Marco Gamba). The description and units of those behaviours, which were used for the data analysis in this research, are shown in Supplementary Table S1.
The reliability of behavioural coding was assessed via comparison of the original coding with the results of a second, independent coder, who processed the video files of 13 dogs. Based on Spearman correlation analysis, the coding was reliable in the case of each behavioural parameter. Spearman’s rho – Reward latency: R(39) = 0.998; P < 0.001; Looking at the owner frequency: R(39) = 0.956; P < 0.001; Demonstration duration: R(26) = 0.958; P < 0.001); Demonstration watching duration: R(26) = 0.920; P < 0.001).
We used the following derived behavioural variables for further analysis: latency improvement, where we focused on whether the dogs showed a consistently improving detour performance from trial to trial. This parameter was calculated via division of Trial 2 latency by Trial 1 latency (Trial 2 Latency Improvement); and via division of Trial 3 latency by Trial 2 latency (Trial 3 Latency Improvement). Furthermore, we calculated the average of the three trials’ looking at the owner frequencies (Average Owner Looking). Finally, we also used the relative duration of watching the demonstration in Trials 2 and 3, which was the total duration while the dog looked at the demonstrator from the beginning till the end of her walk, divided by the demonstration’s duration. The demonstrations have been performed in a highly consistent and steady manner, with only a minimal variation in their length. The duration of demonstrations did not differ significantly between Trials 2 and 3 (GEE, Wald Chi2(1) = 0.296; P = 0.587; Means: Trial 2 = 15.58s, 95%CI: 15.12–16.04; Trial 3 = 15.49, 95%CI: 14.99–15.48).
2.5. Statistical analysis
We used IBM SPSS.29 for analysing the data. Significance level was 0.05 in all cases. First, we ran Principal Component Analysis, based on correlations between variables with varimax rotation, to assess whether there were significant associations among the behavioural parameters. The number of PCA components was chosen using the break point of the scree plot (see Cattell, Reference Cattell1966). If a behaviour had loading < 0.25, it was automatically disregarded. To further simplify the components, we applied a backward elimination approach, excluding step-by-step those behaviours that had low loading (<0.5), or contributed to more than one component with similar absolute loading (this approach is commonly used in PCA analysis, e.g., Cadima & Jolliffe, Reference Cadima and Jolliffe2001). Cronbach’s alpha was calculated to assess the internal consistency of the final extracted factors and to test the repeatability of the measurement (DeVellis & Thorpe, Reference DeVellis and Thorpe2021).
We found two components with the PCA (see Supplementary Table S2), which together explained 78.048 percent of the total variation. Both components were reliable according to the Cronbach-alpha analysis (Component 1 ‘Latency Improvement’: Cronbach’s alpha = 0.736; Component 2 ‘Relative Demo Watching’: Cronbach’s alpha = 0.668). From among the behavioural elements, Average Owner Looking has been excluded from the components because of its too low factor loading.
With the calculated factor scores of the two components, we ran a hierarchical cluster analysis using Ward’s algorithm. We used the resulting agglomeration schedule for deciding the number of clusters, where we applied the ‘elbow method’ to find the steepest jump of distances between the neighbouring coefficients. We identified 6 clusters, and based on this, we ran an additional K-means clustering to finalize the membership of subjects in the clusters.
Finally, we performed General Linear Models with Tukey post hoc tests to find out whether the clusters can be characterized with significantly different Latency Improvement (in short ‘Improvement’) and Relative Demo Watching (in short ‘Attention’) values. We also added to the models the dogs’ housing conditions (indoor-only vs. indoor-outdoor keeping [none of the dogs tested were only kept outdoors]), their training history (none, trained only at home, one course at dog school, regular dog school, private trainer, specific work/sport training), breed type (cooperative/independent) and ancestry groups (according to Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017; Pongrácz & Dobos, Reference Pongrácz and Dobos2025b) as fixed factors.
3. Results
3.1. Descriptive analysis of the clusters
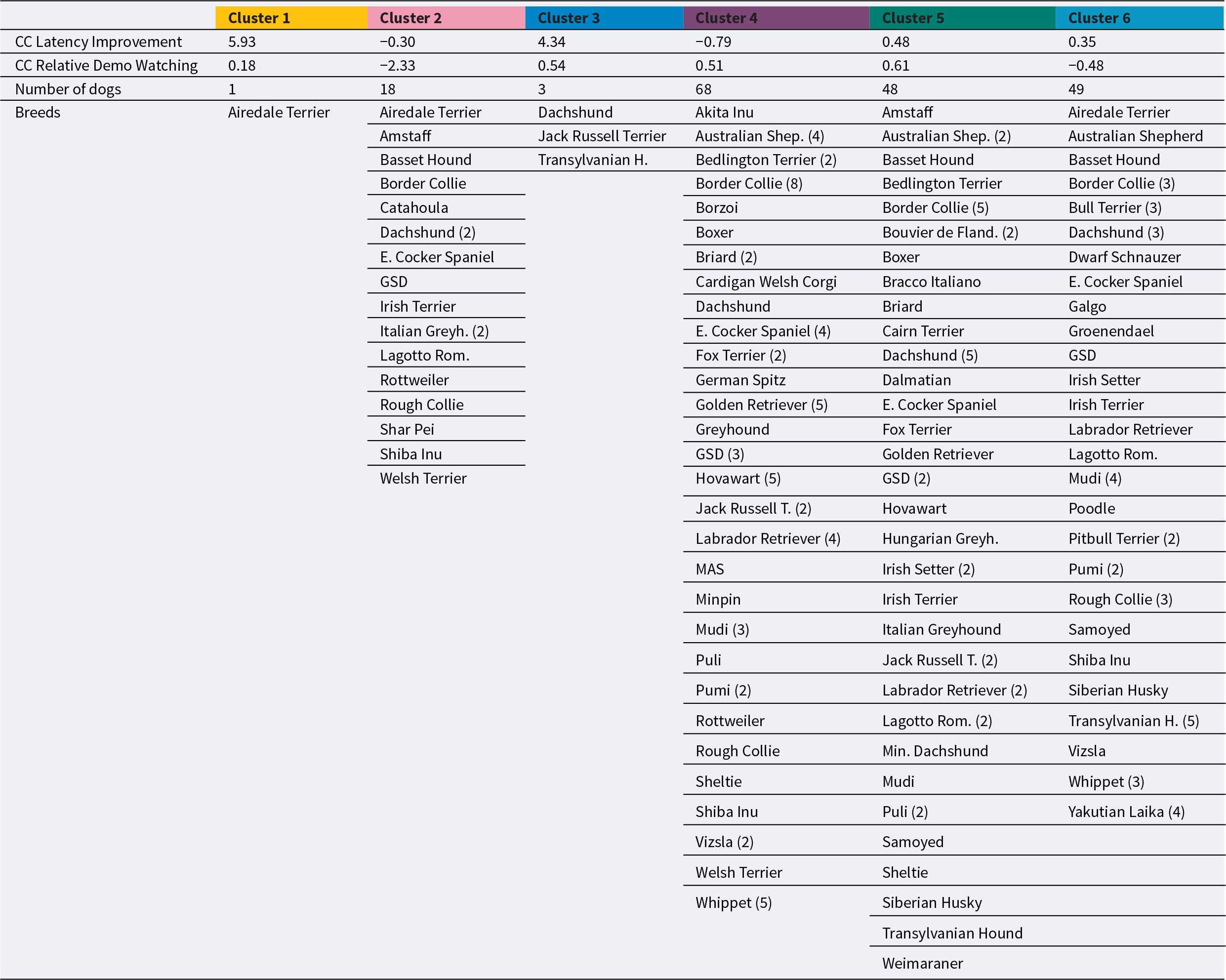
Table 1 shows the structure of the clusters we have identified.
Final cluster centres (CC) according to the K-means procedure, and dogs belonging to the 6 clusters. Amstaff: American Staffordshire Terrier; GSD: German Shepherd Dog; MAS: Miniature American Shepherd; Minpin: Miniature Pinscher. The background colours in the first row correspond with the colours used for the individual clusters in Figure 1

Table 1 Long description
The table lists six K-means clusters with two cluster-centre measures: latency improvement and relative demo watching, plus the number of dogs and breed membership per cluster. Latency improvement is highest in Cluster 1 at 5.93, followed by Cluster 3 at 4.34; Clusters 2 and 4 are below zero at minus 0.30 and minus 0.79. Relative demo watching is lowest in Cluster 2 at minus 2.33 and highest in Cluster 5 at 0.61; Clusters 3 and 4 are also positive at 0.54 and 0.51, while Cluster 6 is slightly below zero at minus 0.48. Cluster sizes vary widely: Cluster 1 has 1 dog, Cluster 3 has 3, Cluster 2 has 18, and the largest groups are Cluster 4 with 68 dogs, Cluster 6 with 49, and Cluster 5 with 48. Breed lists show many breeds per cluster, with repeated representation of herding and working types such as Border Collie, German Shepherd Dog, and Australian Shepherd across multiple clusters. Because clusters differ greatly in size, the cluster-centre values for very small clusters, especially Cluster 1 and Cluster 3, should be interpreted cautiously.
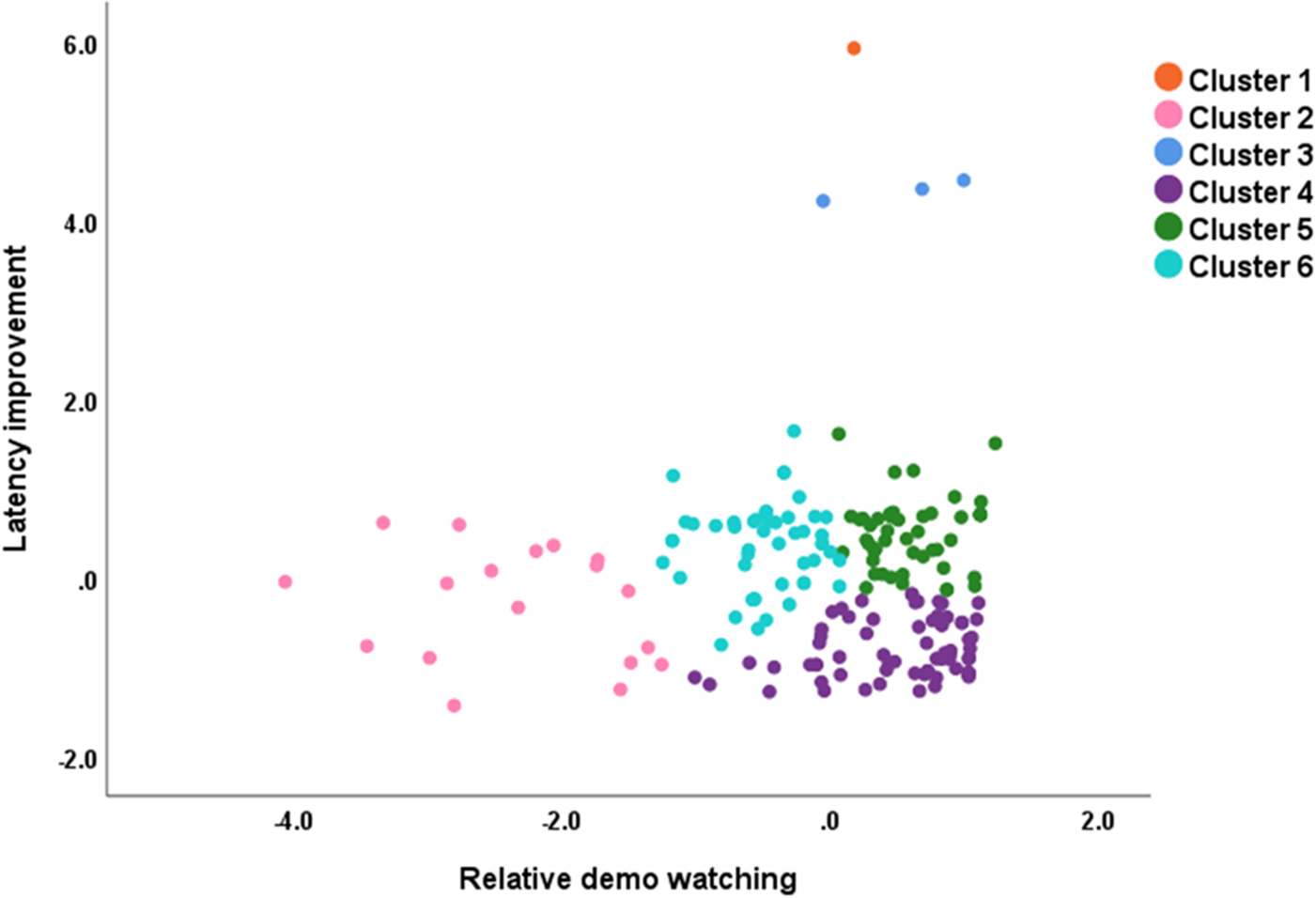
We show the overall positioning of the six clusters in Figure 1.
The six clusters, according to the latency improvement and relative demo watching values of the individual dogs. The breeds that belong to the individual clusters are listed in Table 1. In the case of latency improvement, smaller values mean that the dogs made faster detours towards the end of the test.

Cluster 1 + Cluster 3: These four dogs (Airedale Terrier, Jack Russell Terrier, Dachshund, and Transylvanian Hound) showed high relative demo-watching but could not improve at all – on the contrary, their latencies became even longer along the trials. Cluster 2: This cluster is characterized by minimal relative demo-watching, but in general good latency improvement. There are 16 various dog breeds in the cluster, mainly (12) of the independent working type. Cluster 4: These dogs are characterized by high relative demo-watching and the best improvement in latency. This is the largest cluster (68 dogs), dominated by cooperative breeds, while independent working dogs (N = 13) were rare in this cluster. Cluster 5: Dogs in this cluster also watched the demo a lot but improved their latencies somewhat less than Cluster 4. This large (48 dogs) cluster can be characterized as a slightly ‘lower’ performing variant of Cluster 4. The ratio of independent (20) and cooperative working dogs (28) was more balanced. Cluster 6: Moderate amount of relative demo-watching and only modest latency improvement characterized this cluster. This is also a large (49 dogs) cluster, where there were slightly more independent working dogs (27) than cooperative ones (22).
Primitive/Basal breeds (e.g., Shiba Inu, Siberian Husky, Yakutian Laika) were scattered across most clusters, but they were the most frequent in Clusters 2 and 6, with modest-to-low relative demo-watching but rather effective learning. The best-performing dogs (i.e., they showed the most effective social learning) concentrated in Cluster 4. It mainly consisted of cooperative working gundogs (such as the Golden Retriever and English Cocker Spaniel), modern utility breeds (GSD, Belgian herding dogs, Hovawart), herding dogs (e.g., Border Collie, Mudi), and an independent sighthound (Whippet). Scent hounds and earth dogs (Dachshunds, Terriers) were also scattered across the clusters.
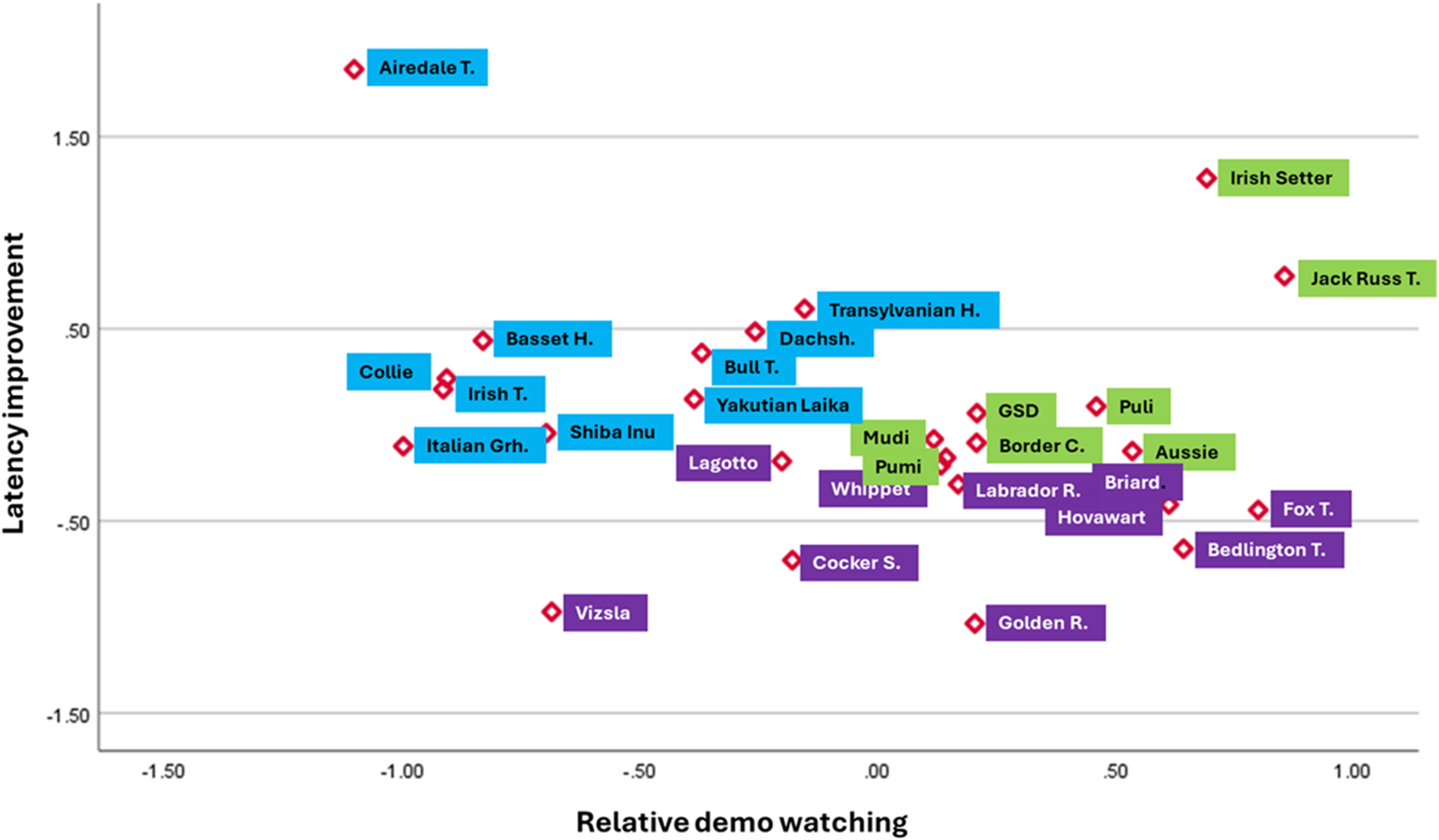
When we made an order of performance among the clusters in the social learning task, it included Clusters 6, 5, and 4, where we found that the ratio of cooperative dogs in the individual clusters was similarly increasing as the detour latency decreased and relative demo-watching increased. Figure 2 provides a closer look at the positioning of those breeds that had at least three dogs in our sample.
Dog breeds within Cluster 4 (dark purple), Cluster 5 (green), and Cluster 6 (light blue). Breed average coordinates were calculated for each breed that had at least three representatives in our sample. In the case of latency improvement, smaller values mean that the dogs made faster detours towards the end of the test.

Figure 2 Long description
The scatter plot displays latency improvement on the vertical axis and relative demo watching on the horizontal axis. The horizontal axis ranges from negative 0.50 to 1.00, while the vertical axis ranges from negative 0.50 to 1.50. Each point represents a dog breed, with labels such as Alredale T., Irish Setter and Jack Russel T. Clusters are color-coded: Cluster 4 in dark purple, Cluster 5 in green and Cluster 6 in light blue. Notable outliers include Alredale T. and Jack Russel T., which are positioned away from the main cluster. The plot suggests a relationship between demo watching and latency improvement, with breeds in Cluster 6 showing higher latency improvement. The distribution of points indicates varying levels of demo watching across breeds, with some breeds demonstrating faster detours as demo watching increases.
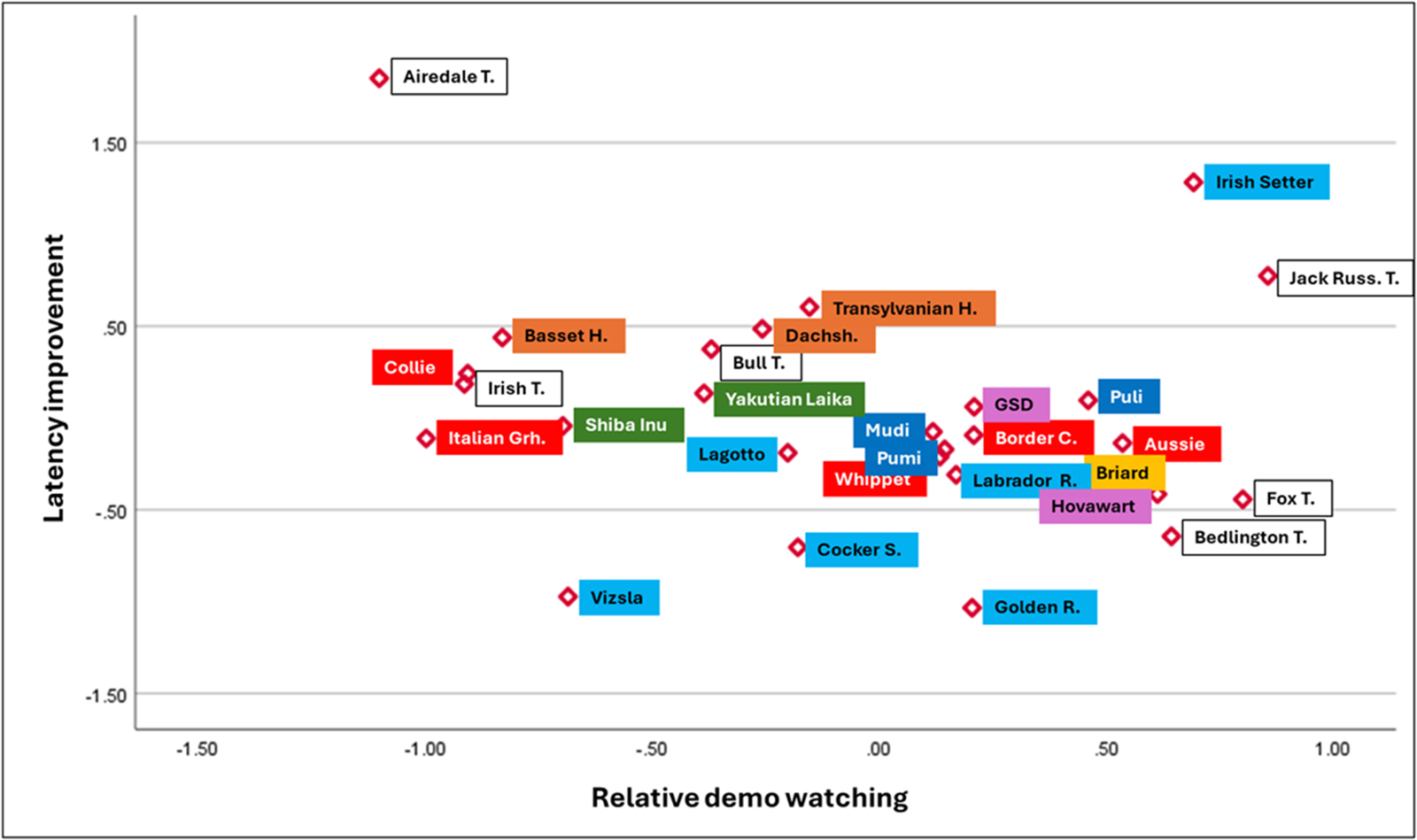
Finally, we examined the positioning of those breeds in the clusters that were represented by three or more dogs in our sample and sorted the breeds according to their place on the ancestry cladogram by Parker et al. (Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017) (Fig. 3). Terriers were scattered across Clusters 4, 5, and 6, enumerating both excellently and poorly performing breeds according to their detour latencies and relative duration of watching the human demonstrator. Scent hounds did not pay too much attention to the demonstrator and showed weak latency improvement; however, most gundogs were among the best performers in our sample. Herding dogs (British and Continental breeds alike) and modern utility breeds (such as the German Shepherd Dog) showed high values in relative demo-watching but did not improve their detour latencies as much as the gundogs.
Dog breed averages in Clusters 4, 5, and 6, according to the breeds’ location in the cladogram of Parker et al. (Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017). Primitive breeds (Clade A): green; Hungarian herding dogs (Clade G): dark blue; Terriers (Clade L): white; German Shepherd Dog (Clade M): purple; Scent hounds (Clade O): Orange; Gundogs (Clades P + Q + R): light blue; Briard and Belgian Sheepdogs (Clade S): yellow; British herding dogs and sighthounds (Clade T): red. In the case of latency improvement, smaller values mean that the dogs made faster detours towards the end of the test.

Figure 3 Long description
The scatter plot displays latency improvement on the vertical axis and relative demo watching on the horizontal axis. The plot includes various dog breeds, each represented by a labeled point. Notable outliers include Airedale Terrier and Vizsla, which are positioned far from the main cluster of breeds. The main cluster of breeds is centered around the origin, indicating a mix of latency improvement and demo watching. The legend uses color coding to categorize breeds: primitive breeds, Hungarian herding dogs, terriers, German Shepherd Dog, scent hounds, gundogs, Briard and Belgian Sheepdogs and British herding dogs and sighthounds. The plot illustrates the relationship between the two variables, showing that some breeds have higher latency improvement with less demo watching, while others show the opposite trend.
3.2. The association among clusters, breed type, ancestry groups, and behavioural parameters
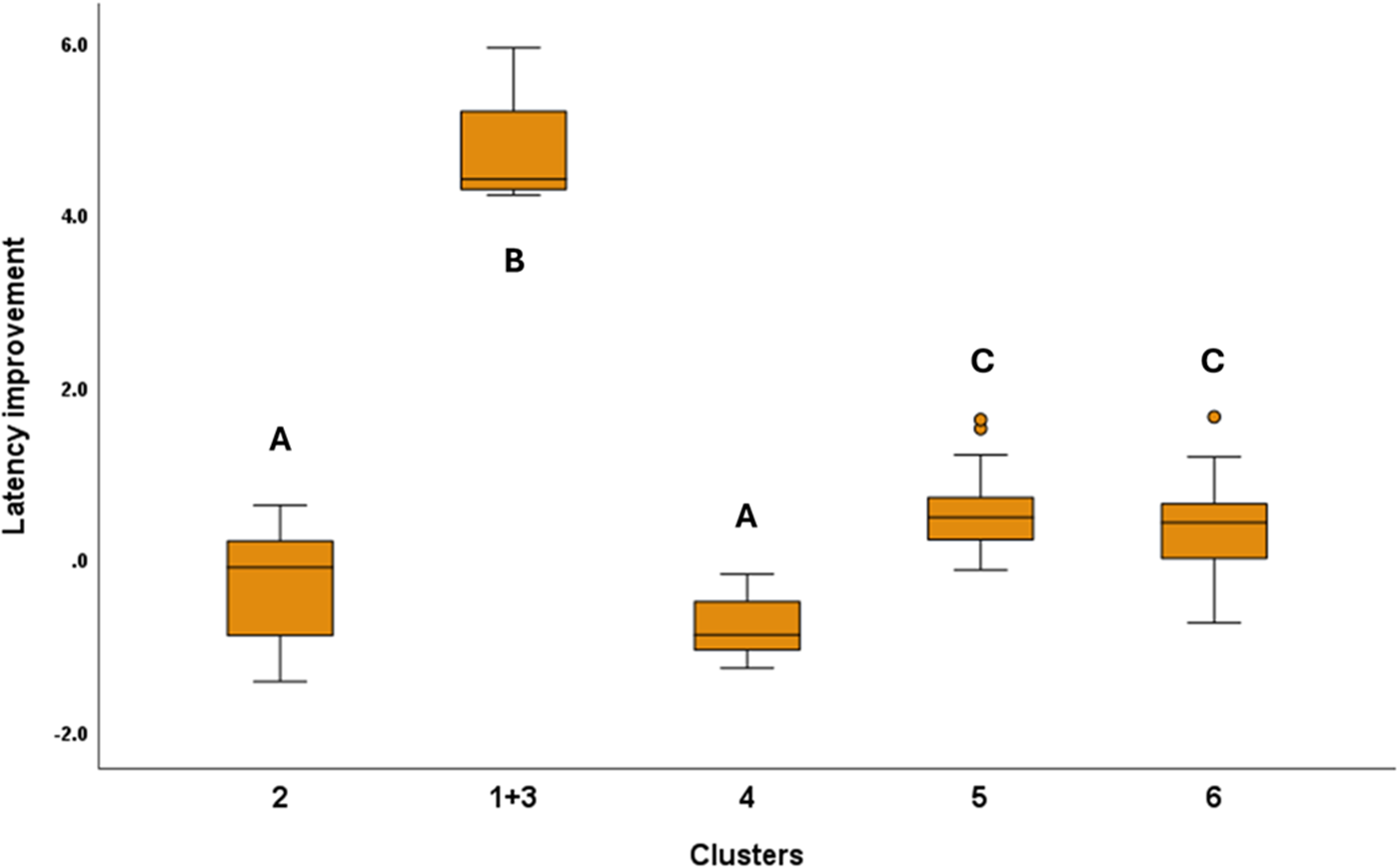
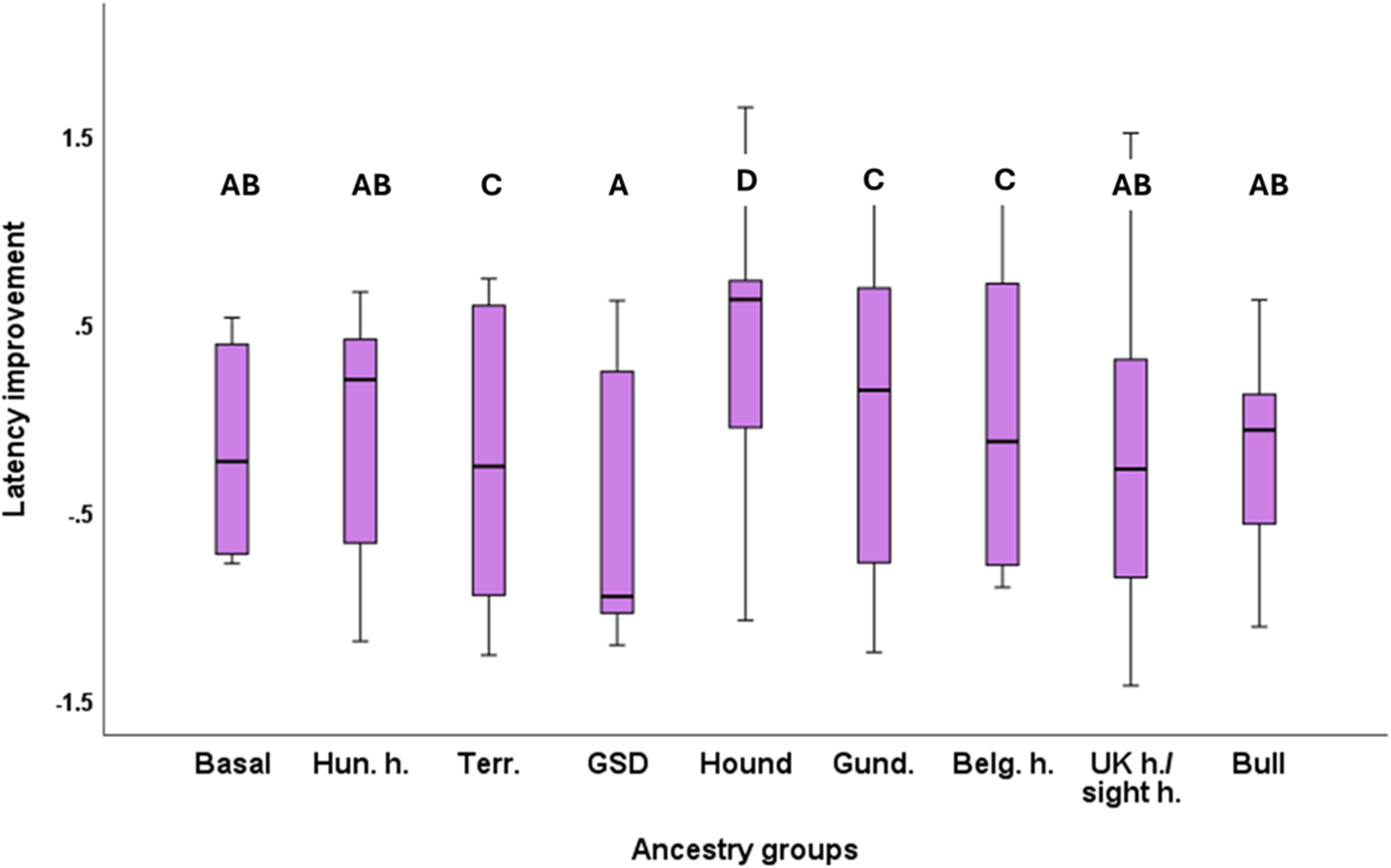
For this analysis, we merged Cluster 1 with Cluster 3 because Cluster 1 had only one dog in it, and both clusters had similar final cluster centres (see Table 1). Latency Improvement had significant association with the Clusters (F4,148 = 176.06; P < 0.001). According to the Tukey post hoc test, Clusters 2 and 4 improved the most, while the merged Cluster 1 + 3 showed the poorest detour performance. Clusters 5 and 6 were in between, and these did not differ in their latency improvements (Fig. 4). We also found a significant association between Latency Improvement and Ancestry Groups (F8,148 = 3.290; P = 0.0027). According to the Tukey post hoc test, Scent hounds (Clade O) showed the lowest improvement of the detour latency, while German Shepherd Dogs and Hovawarts (Clade M) performed the best (Fig. 5). Housing (F1,148 = 1.142; P = 0.287), training history (F5,148 = 2.232; P = 0.054) and breed type (F1,148 = 0.052; P = 0.820), did not have significant association with Latency Improvement.
The significant association between latency improvement and the clusters. For this analysis, we merged Clusters 1 and 3 because of their low subject numbers and similar performance. Significant group differences are marked with different letters. In the case of latency improvement, smaller values mean that the dogs made faster detours towards the end of the test.

The significant association between latency improvement and the dog breeds’ ancestry groups. Significant group differences are marked with different letters. In the case of latency improvement, smaller values mean that the dogs made faster detours towards the end of the test. Hun. H. = Hungarian herding breeds; Terr. = Terriers; GSD = German Shepherd Dog (+ Hovawart); Gund. = Gun dogs; Belg. H. = Belgian herding dogs and Briard; UK h. = British herding dogs; sight h. = Sight hounds; Bull = Bull type dogs.

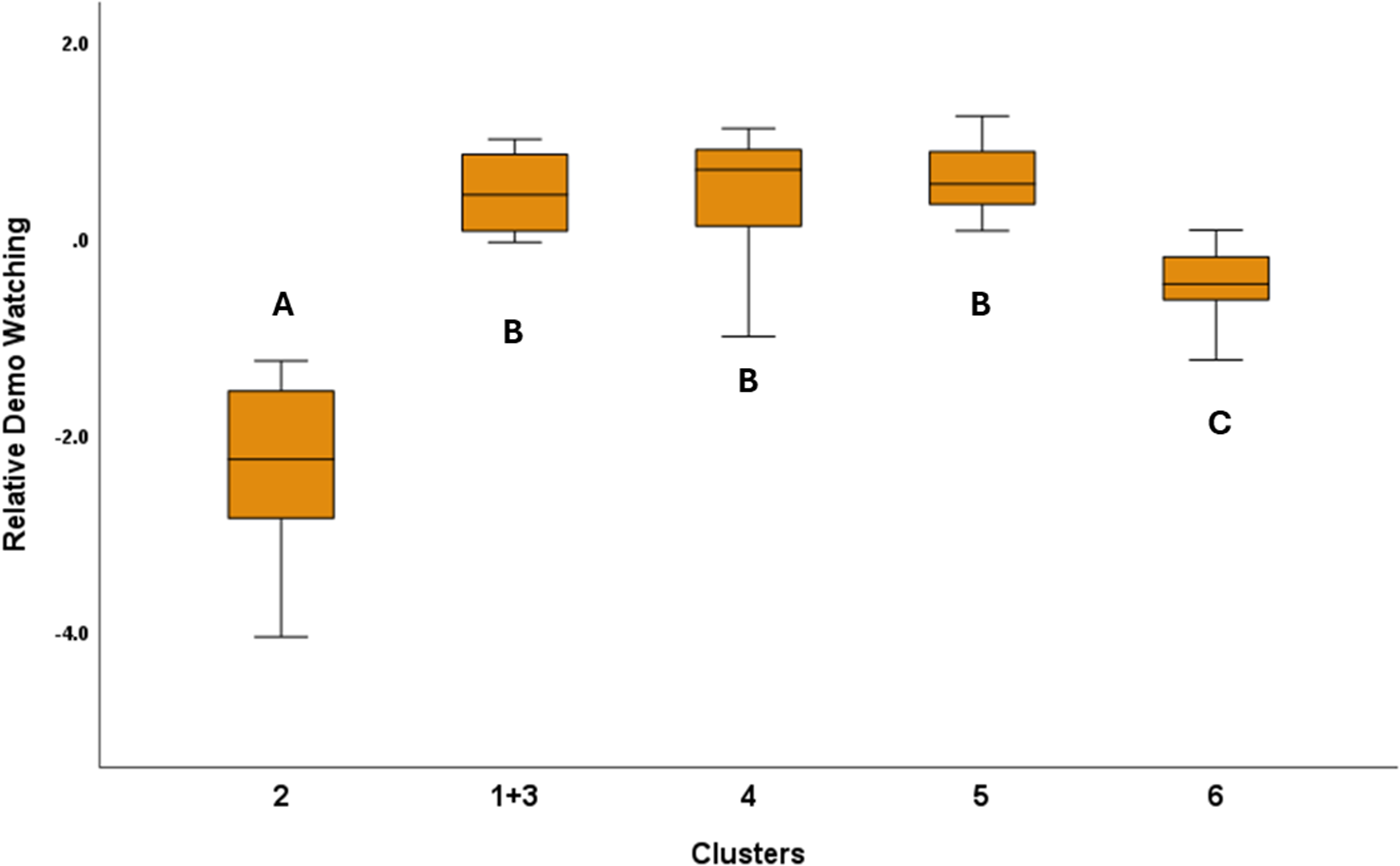
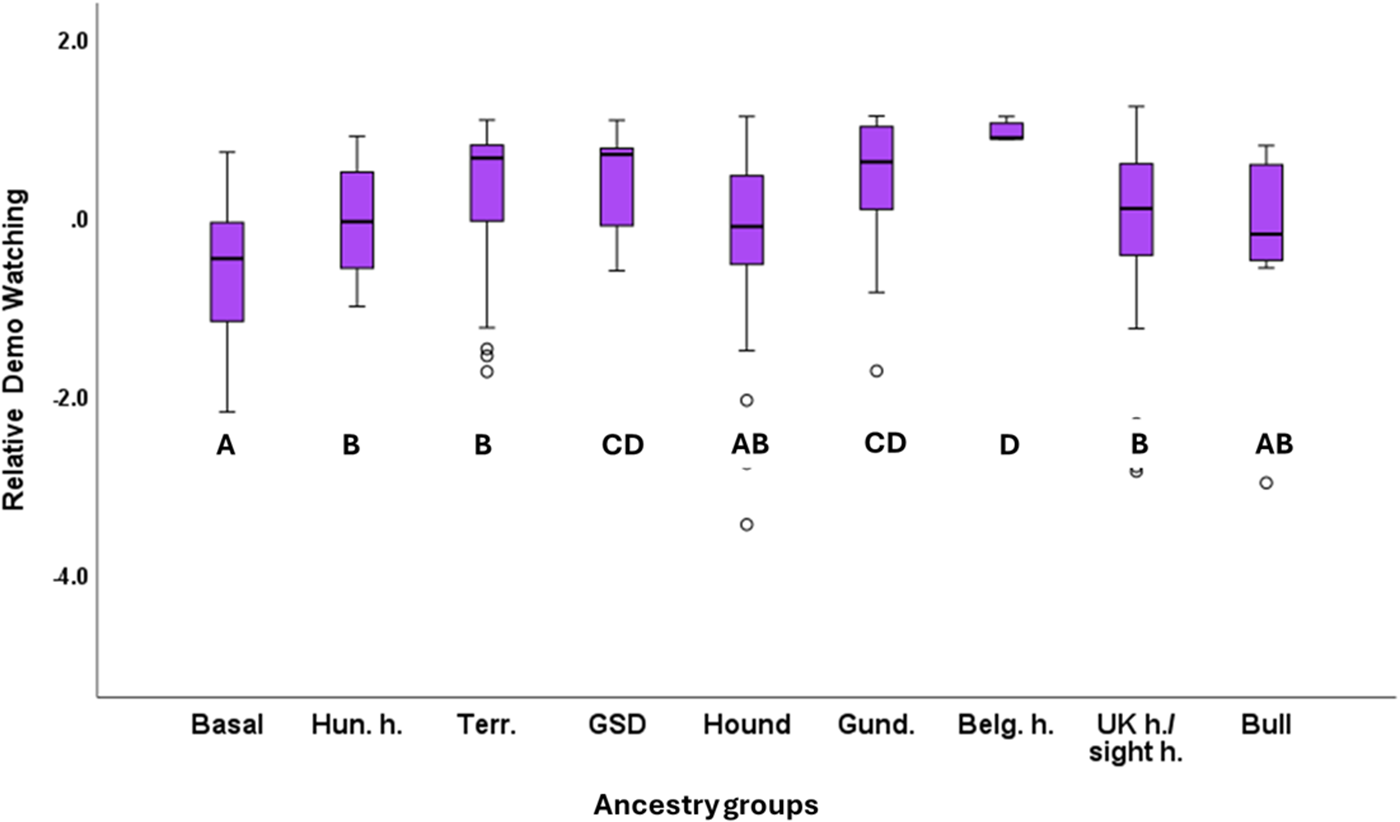
In the case of Relative Demo Watching, we found a strong significant association with the Clusters (F4,148 = 115.064; P < 0.001). According to the post hoc test, dogs in Cluster 2 had the lowest relative demo-watching, and Cluster 6 had the second shortest relative demo-watching values. The other clusters had significantly higher relative demo-watching, and they did not differ from each other (Fig. 6). Ancestry Groups also had a significant association with Relative Demo Watching (F8,148 = 2.056; P = 0.044). According to the post hoc tests, Belgian sheepdogs had the highest relative demo-watching, while the primitive/basal dog had the shortest relative demo-watching. Scent hounds and bull-type dogs also had lower relative demo-watching, while the gundogs and the German Shepherd dog showed higher values of relative demo-watching (Fig. 7). Housing (F1,148 = 0.489; P = 0.486), training history (F5,148 = 1.142; P = 0.341) and Breed Type (F1,148 = 1.206; P = 0.274) did not associate significantly with Relative Demo Watching.
The significant association between relative demo watching and the clusters. For this analysis, we merged Clusters 1 and 3 because of their low subject numbers and similar performance. Significant group differences are marked with different letters.

The significant association between relative demo watching and the ancestry groups. Significant group differences are marked with different letters. Hun. H. = Hungarian herding breeds; Terr. = Terriers; GSD = German Shepherd Dog (+ Hovawart); Gund. = Gun dogs; Belg. H. = Belgian herding dogs and Briard; UK h. = British herding dogs; sight h. = Sight hounds; Bull = Bull type dogs.

5. Discussion
In this study, our main goal was to use a behaviour-based approach to identify biologically relevant differences in one of the main synchronizing mechanisms between dogs and humans: social learning. We used a new approach to examine the behaviour of dogs in a spatial problem-solving task. Instead of applying one of the well-known grouping factors in dog breed-related research (e.g., functional breed selection or genetic relatedness), we identified two composite behavioural dimensions: attention and improvement, which were of fundamental importance to the dogs’ ability to learn from a human demonstrator. These behavioural dimensions can be considered as belonging to a dependence-independence scale between dogs and humans. The attention dimension was characterized by the relative duration of watching the demonstrator during the execution of a detour around a V-shaped fence. Improvement shows the speed of reaching the reward, which indicates the individual’s ability to effectively learn from the demonstrator. Of course, improvement could also be achieved via individual (trial-and-error) learning by some subjects; however, in other studies where dogs had to perform repeated trials without demonstration, the subjects did not show significant improvement along the trials (e.g., Dobos & Pongrácz, Reference Dobos and Pongrácz2023; Pongrácz et al., Reference Pongrácz, Miklósi, Timár-geng and Csányi2004). By comparing individuals with the help of these two behavioural dimensions, we could for the first time assess the social learning capacity of dogs without masking potential breed-related differences by sorting them into functional or ancestry-based groups.
With cluster analysis, we formed six distinguishable clusters of dogs that participated in the social learning experiments. These clusters showed significant association with the two dimensions: the relative duration of watching the demonstration and improvement in the latency of reaching the reward. There were a few dogs (Clusters 1 and 3) that seemingly paid a lot of attention to the demonstration (based on their high relative demo-watching values); however, they could not learn at all how to make a detour effectively and faster along the trials. These dogs were highly attentive towards human activity and communication; however, their performance did not show improvement as a result. We can hypothesize that these dogs watched the demonstrator not necessarily with an interest in the solution of the problem, but as a sign of giving up on a difficult task. This human-dependent behaviour was found to be very typical in dogs who were facing an ‘unsolvable task’ (e.g., Hirschi et al., Reference Hirschi, Mazzini and Riemer2022; Pongrácz & Lugosi, Reference Pongrácz and Lugosi2024). Dogs in Cluster 2 had very low relative demo-watching; however, they detoured the fence increasingly faster along the trials, indicating that this group looked less likely at the demonstrator but still improved their performance. In this cluster, most subjects belonged to the independent working dog breeds, which have a lesser need for interaction with their owner during the original type of work they were selected for. Therefore, the resourcefulness of these dogs might be connected to artificial selection by humans. Earlier, it was found that effective learning from the human demonstrator was in positive association with the relative duration of demo watching (e.g., Dobos & Pongrácz, Reference Dobos and Pongrácz2024). Interestingly, in Cluster 2, most breeds were represented by only one dog, which highlights that effective social learning with low interest in the demonstration can be a somewhat exceptional phenomenon. As we found no effect of the training history and housing conditions on the relative duration of watching the demonstrator, these environmental factors most probably did not contribute relevantly to the paradoxically good performance of the dogs in Cluster 2. Dogs in Cluster 5 represented a transitional level of reliance and dependence on human help in the social learning task. These dogs had medium-level relative demo-watching and learned the task with moderate success. Finally, Clusters 4 and 6 included the best learners who showed equally high attention to the demonstration. Interestingly, we found an almost balanced number of cooperative and independent working breeds in these clusters. Contrary to previous considerations about independent breeds’ low levels of interest in human communication (e.g., Dobos et al., Reference Dobos, Lugosi and Pongrácz2025; Dobos & Pongrácz, Reference Dobos and Pongrácz2024; Gácsi et al., Reference Gácsi, McGreevy, Kara and Miklósi2009), in these clusters, representatives of both functional breed groups were highly attentive towards the human demonstration and were able to improve thanks to the provided information. Clusters 4 and 6 segregated based on slightly different improvements of detour latencies, which highlight the importance of finer details of breed-typical behaviours in connection with the requirements of the detour task. One could hypothesize that maybe some dogs could relate the positioning of the reward behind an obstacle as being ‘forbade’ to go for it, resulting in lower performance in the test. This was unlikely as the whole experimental procedure (placement of the reward and the subsequent problem-solving phase) included the constant positive attention eliciting and encouraging from both human partners (demonstrator and owner) towards the dogs.
Regarding the individual dog breeds, our results revealed two typical arrangements in connection with the behavioural dimensions. Some breeds were impossible to characterize by a typical behaviour according to our clustering paradigm and were scattered across the clusters (e.g., Dachshunds). Some breeds, represented by relatively higher numbers of individuals in our sample (Australian Shepherd, Border Collie, Mudi, Transylvanian Hound and Whippet), typically appeared in more than one of the behaviour-based clusters. This indicates a considerable within-breed variability of our behavioural dimensions, which supports the importance of investigating other than breed-based clustering systems in the case of canine cognition and behaviour (see also Morrill et al., Reference Morrill, Hekman, Li, McClure, Logan, Goodman, Karlsson, Dong, Alonso, Carmichael, Snyder-Mackler, Alonso, Noh, Johnson, Koltookian, Lieu, Megquier, Swofford, Turner-maier and Karlsson2022). Sources of relevant individual (i.e., non-breed dependent) differences in being attentive towards human behaviour could be the selection of within-breed working and show-lines (Cavalli et al., Reference Cavalli, Carballo, Dzik and Bentosela2020), housing conditions (e.g., D’Aniello & Scandurra, Reference D’Aniello and Scandurra2016; Topál et al., Reference Topál, Miklósi and Csányi1997), and the training level of the dog (Lazarowski et al., Reference Lazarowski, Singletary, Rogers and Waggoner2021).
Contrarily, some breeds showed a compact disposition along the two behavioural variables (e.g., Transylvanian Hound, Yakutian Laika). This indicates that in this particular social learning paradigm, some breeds had more predictable behaviour than others did. Their capacity to learn from an unfamiliar human might be more dependent on genetically determined traits (i.e., affected by domestication and artificial selection), which are probably less modified by lifetime differences (e.g., training). Similarly, there were groups of otherwise closely related breeds, such as the terriers, that were scattered around the clusters. Interestingly, some terrier breeds (Fox Terrier, Bedlington Terrier, and Jack Russell Terrier) showed the highest attention, from among all the tested subjects, to the human demonstration. These results may highlight differences within ancestry groups, which are specific to the selection history of some included breeds. For example, terriers that excelled in paying attention to the demonstrator may have been subjected to intense directional selection where not only their independent vermin-exterminating drive, but also high controllability by the handler, was also preferred. We should again remember that training history did not have a significant association with any of our behavioural dimensions; thus, the excellent performance of these terriers was most probably not related to their training. Except for a few dogs that showed high relative demo-watching but could not improve their detour performance, breeds were more widely dispersed throughout the relative demo-watching axis; meanwhile, there was less difference in their latency improvement. There might be a threshold for the minimum duration of attention that is necessary for effective learning from the demonstrator. Below this threshold, the dogs do not show effective improvement, as it was also shown in studies with no demonstration control groups (Dobos & Pongrácz, Reference Dobos and Pongrácz2023; Pongrácz & Dobos, Reference Pongrácz and Dobos2025c). Above this threshold, the dogs can show considerable within- and between-breed variance in their dependence on the demonstration. This can be affected by breed function, ancestry, or other individual differences, including experience.
Based strictly on the ancestry grouping, the best-performing dogs with the most effective social learning ability were the closely related German Shepherd Dogs and Hovawarts. This result fits with our earlier findings (Pongrácz & Dobos, Reference Pongrácz and Dobos2025b), where it was found that this ancestry group excels in a detour task if they are provided with a demonstration. However, in this current research, by applying cluster analysis on the combined data of social learning and paying attention to the demonstration, we found that there were two larger, complex groups of dog breeds among the most effective learners. Besides the previously mentioned two breeds, Cluster 4 is mainly comprised of the cooperatively working gundogs (such as the Golden Retriever and English Cocker Spaniel), herding dogs (e.g., Border Collie, Mudi, Belgian sheepdogs), and an independently working sighthound (Whippet). According to our new, behaviour-based approach, this cluster contained the most human-dependent breeds as they also showed the highest relative demo-watching values. Although functional breed type (independent vs. cooperative) did not associate significantly with any of the behavioural dimensions based on the whole sample, Cluster 4 contained mostly cooperative working breeds. The cooperative working dogs’ high performance in social learning was confirmed in earlier studies (e.g., Dobos & Pongrácz, Reference Dobos and Pongrácz2023, Reference Dobos and Pongrácz2024). In the case of some sighthound breeds (e.g., the Whippet), attention to human communication and demonstration can be connected to their close genetic relationship with the highly dependent British herding dog breeds (Parker et al., Reference Parker, Dreger, Rimbault, Davis, Mullen, Carpintero-ramirez and Ostrander2017; Pongrácz & Dobos, Reference Pongrácz and Dobos2025b). The other excellently performing group was Cluster 2, which contained an interesting selection of single representatives of many, mostly independent working dog breeds. Apart from its superb latency improvement, this cluster was the opposite of Cluster 4, as Cluster 2 had the lowest relative demo-watching values.
Interestingly, Cluster 4, where most gundogs were concentrated, performed significantly better in the social learning task than Clusters 5 and 6, where most of the herding dogs were sorted. So far, earlier studies that utilized the detour paradigm with human demonstration could not separate these breeds based on their latency improvement. Dobos and Pongrácz (Reference Dobos and Pongrácz2023, Reference Dobos and Pongrácz2024) concluded that as both gundog and herding dog breeds belong to the cooperative type, similar high performance and relative demo-watching values should be expected from them. Other investigations, using different experimental paradigms (e.g., following human pointing signals: Gácsi et al. (Reference Gácsi, McGreevy, Kara and Miklósi2009)), or analysing overarching genetic traits (e.g., ‘biddability’: Morrill et al. (Reference Morrill, Hekman, Li, McClure, Logan, Goodman, Karlsson, Dong, Alonso, Carmichael, Snyder-Mackler, Alonso, Noh, Johnson, Koltookian, Lieu, Megquier, Swofford, Turner-maier and Karlsson2022)) also found gundogs and herding dogs similarly close in these socio-cognitive traits. Turcsán et al. (Reference Turcsán, Kubinyi and Miklósi2011), based on a large-scale questionnaire study, placed herding and hunting dogs into different clusters based on their trainability and boldness; however, these results are not directly comparable with our analysis, because Turcsán and colleagues did not distinguish between cooperative gundogs and independent hunting dogs (scent- and sighthounds). Therefore, our new approach has, for the first time, found differences between the human-dependent behaviour (social learning) of two working dog types, which are otherwise equally highly watchful observers of human demonstration and verbal communication (Dobos et al., Reference Dobos, Lugosi and Pongrácz2025; Dobos & Pongrácz, Reference Dobos and Pongrácz2024). This new result indicates that finer, behaviour-based analyses can reveal relevant differences between dog breeds’ (or rather breed types’) behaviour, which otherwise might remain hidden when more general grouping variables are used. Gundogs were bred not only to work cooperatively with their owner and rely on their utterances, but also to find and retrieve prey on command (Delgado et al., Reference Delgado, Stella, Croney and Serpell2024; Lazarowski et al., Reference Lazarowski, Waggoner, Krichbaum, Singletary, Haney, Rogers and Angle2020). At the same time, herding dogs have been selected for directing livestock without physically attacking or grabbing them (Lasserre et al., Reference Lasserre, Ducreux, Chassier, Joly, Cacheux, Le Morzadec, Dayde-Fonda and Gilbert2024). As the detour task includes a target (the reward) that the dog must reach and touch (i.e., eat), this paradigm might be more suitable for the gundogs to learn from a demonstrator than for herding dogs that must follow the commands of the owner without fetching and without hurting the flock. Moreover, the motivation for food might also be higher in the case of the gundogs, according to studies on risk factors associated with canine obesity (e.g., Such & German, Reference Such and German2015; White et al., Reference White, McBride, Redhead and Bishop2007). From this aspect, it is important to notice the effect of directional selection behind different types of working tasks. These may require specifically refined behavioural traits (e.g., grabbing, retrieving) or enhanced motivations (e.g., towards food), which might result in differences at the socio-cognitive level of the breeds. Our results suggest that there might be breed-related differences that come to light if the approach to investigating dogs’ behaviour does not include wider background variables.
Among the limitations of the study, we can mention that as the dogs’ detour latencies in Trial 1 showed considerable variability, thus, dogs who made a fast detour in Trial 1, in theory they had limited option for further improvement. The limited sample size did not allow us to exclude these dogs from analysis. However, we found that the majority of the fast initial detourers still managed to steadily improve from trial to trial; thus, their inclusion probably did not bias the results. Another limitation of our study was that most of the examined breeds were represented by only a few individuals; thus, in these instances the breed-level generalizability of the results was low. However, the collected data in the social learning paradigm still allowed us to compare many breeds that were selected for different types of work with humans. On the other hand, our sample was limited to cooperative or independent working dog breeds and did not include breeds that were primarily selected for companion animal purposes (e.g., various toy breeds, such as the Bichon Havanese or the Pug). It would, however, be interesting to analyse companion breeds regarding their human dependence in a social learning task, as these breeds were found to be more dependent than working breeds in other interactive scenarios (eye contact: Bognár et al., Reference Bognár, Szabó, Deés and Kubinyi2021; ‘unsolvable’ task: Ujfalussy et al., Reference Ujfalussy, Bognár, Molnár, Miklósi and Kubinyi2023).
6. Conclusions
By comparing the three aspects of how social learning in the various dogs can be examined, we can conclude that genetic distance-based ancestry (Pongrácz & Dobos, Reference Pongrácz and Dobos2025b), functional breed selection (Dobos & Pongrácz, Reference Dobos and Pongrácz2023, Reference Dobos and Pongrácz2024), and the use of behavioural dimensions can be equally appropriate methodologies to investigate and interpret dogs’ behaviour in social learning. In our current study, functional breed types did not have any association with the behavioural dimensions. However, it is important to emphasize that functional breed selection is not an invalid grouping factor. Contrarily, our new, behaviour-based clustering analysis showed that we have the potential to disentangle the different, dependence-based behaviours shown in a social learning scenario, with a more detail-oriented touch. By deepening our focus on the biologically relevant aspects of dogs’ behaviour, the cooperative–independent breed comparison can be further enhanced, and we will have a better explanatory insight into the characteristic breed-typical behaviours.
Importantly, we found that dog breeds can show considerable variance in their socio-cognitive traits that can even overshadow between-breed differences in a context of human-dependent attention and social learning. The true source of this variability remains an exciting research opportunity for the future; however, it would require a much higher sample size from the individual breeds than this current study.
Our study also showed that making generalized assumptions about the behaviour of a dog breed solely based on the breed type can cause people to overlook breeds that would not typically be considered for a task. Biologically relevant testing paradigms can help enthusiasts and professionals to fine-tune their choices without restricting the selection to a few popular breeds.
Supplementary material.
The supplementary material for this article can be found at https://doi.org/10.1017/ehs.2026.10057
Acknowledgements
The authors are thankful to Prof. József Topál for his seminal suggestions on the idea of this research, to Blanka Veres for helping with the tests, and to Celeste R. Pongrácz for English proofreading the manuscript.
Author contributions
P.P.: Conceptualization, methodology, formal analysis, resources, data curation, writing – original draft preparation, writing – review and editing, visualization, supervision. P.D.: Investigation, validation, resources, data curation, writing – review and editing, project administration. Both authors read and approved the final manuscript.
Financial support.
Péter Pongrácz and Petra Dobos were supported by the Hungarian National Research, Development and Innovation Office (NKFIH, Grant # K143077), and by the Morris Animal Foundation (D25CA-039). Petra Dobos was supported by the University Research Fellowships (EKÖP-25-3-I-ELTE-147) of the Ministry for Innovation and Technology.
Competing interests
The authors declare that they have no competing interests.
Research transparency and reproducibility interest
All data generated or analysed during this study are included in this published article, and its supplementary information can be accessed at Mendeley Data [Pongrácz, Péter (2026), ‘Dog breed behavior, dependence and reliance dimensions,’ Mendeley Data, V1, doi: 10.17632/24vr3gk8c5.1].



 Open access
Open access