


‘Through the storm we reach the shore,
You give it all, but I want more,
And I’m waiting for you,
With or without you’
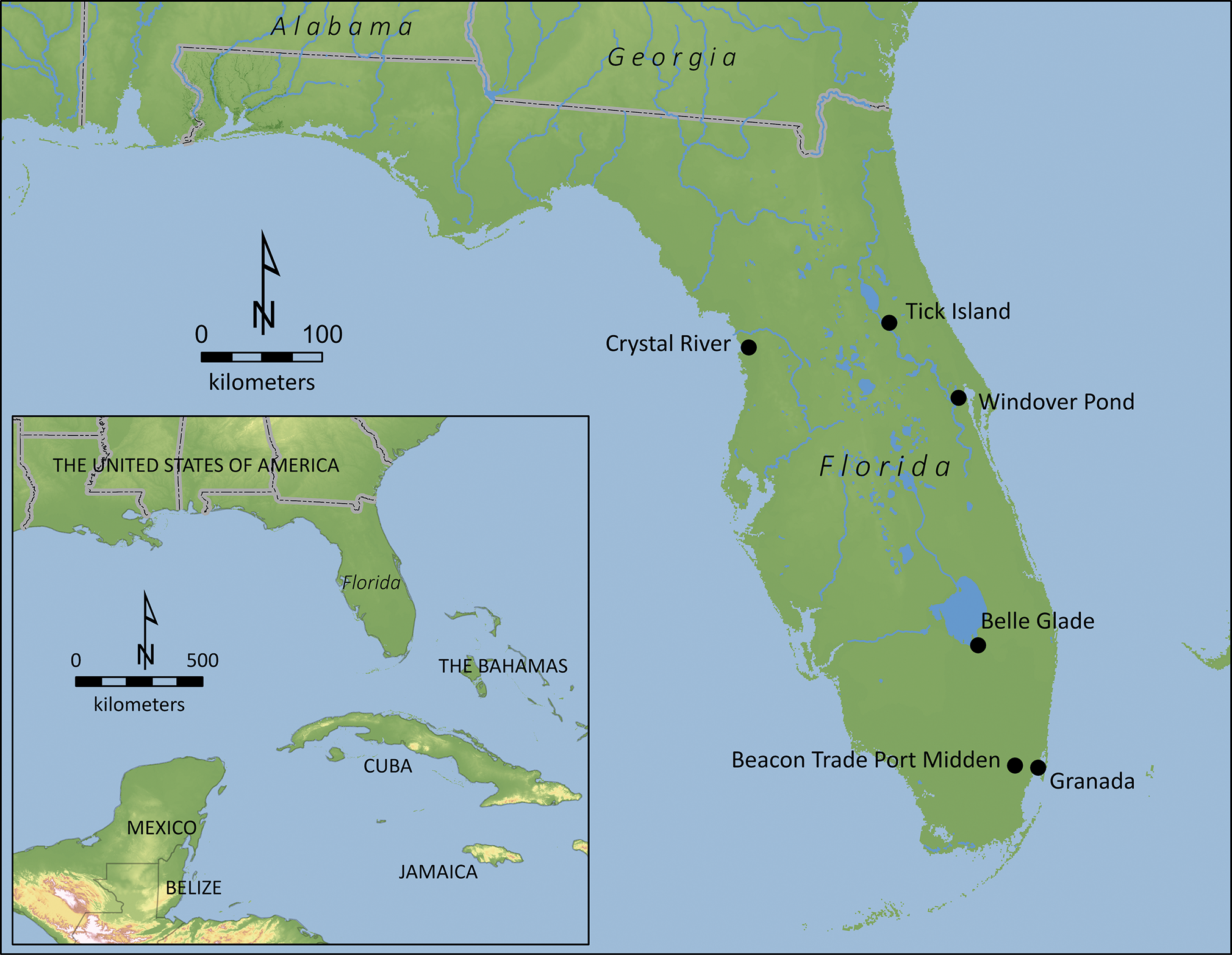
(Bono, 1987)The ‘animal turn’ (Anderson Reference Anderson1997) in the social sciences has generated new understandings of human-animal relations present and past, moving our focus from human mastery over non-human animals (henceforth simply ‘animals’) as economic or symbolic resources to the recognition of animals as agentive beings engaged in mutually constitutive relationships with people (Hill Reference Hill2013). Archaeological zoontologies (Whitridge Reference Whitridge, Baltus and Bareis2017; Wolfe Reference Wolfe and Wolfe2003) have tended to focus on animals with whom people of the past were regularly entangled, either in their everyday lives as companion and work animals—such as cats, dogs and horses (Argent Reference Argent2010; Fadum & Gruber Reference Fadum, Gruber, Recht and Tsouparopoulou2021; Losey et al. Reference Losey, Bazaliiskii and Garvie-Lok2011)—or at least seasonally as favoured prey—like deer, bears, swans and dugongs (Brittain & Overton Reference Brittain and Overton2013; Conneller Reference Conneller2004; Hamilakis & Overton Reference Hamilakis and Overton2013; Losey et al. Reference Losey, Bazaliiskii, Lieverse and Watts2013; McNiven Reference McNiven2010; McNiven & Feldman Reference McNiven and Feldman2003; Whitridge Reference Whitridge, Baltus and Bareis2017). In contrast, I focus on an archaeological case study representing a more ‘eventful’ form of human–animal relations: irregular and infrequent ‘encounters’ between Indigenous peoples and Florida manatees (Trichechus manatus latirostis) in precolonial Florida, USA (Fig. 1).
Location of Florida and sites mentioned in the text.
There is a logic to the archaeological inclination to focus on familiar animals, beyond the practical reasoning that the taxa most commonly companioned or consumed are also those most likely to be represented in faunal assemblages. More broadly, as Oma (Reference Oma2006, 201) describes, proximity is a key component of human–animal relations; the concurrence in time and space that accompanies human relations with domestic animals provides embodied sensory and tactile experiences that may be key to deeper interspecies relationships and the human understanding of an animal as ‘an individual, an agent, and a subject’.
Still, as Bille et al. (Reference Bille, Hastrup, Flohr Sørensen, Bille, Hastrup and Flohr Sørensen2010, 11) note in their call for an ‘anthropology of absence’, there is a difference between simple spatial proximity (‘being near’) and ‘presence’, as well as between ‘being away from’ and ‘absence’. Hill (Reference Hill2013, 126) argues that hunter-gatherers, while perhaps lacking the proximity afforded by living with domesticated animals, form relationships with animals through ‘tracking, traveling, dreaming, listening, [and] observing’. Haraway (Reference Haraway2008, 205) observes that ‘agential cuts’ are made in contact zones with complicated histories and geographies. Drawing from this, Metcalf (Reference Metcalf2008, 115) argues that it is possible for humans and animals to develop ‘intimacy without proximity’. As an example, he describes humans and grizzly bears as companion species of a sort given their extended, shared histories, imbricated political boundaries, and overlapping ecological niches—among other entanglements.
In addition, as Watson and Warkentin (Reference Watson and Warkentin2013, 2) note in regard to relational spaces, proximity operates in both ‘physical and emotional realms’, and embodiment involves not only immediate sensory experience but also the longer-term memories that may be generated by encounters. Consistent with this, studies suggest that modern human encounters with wild animals—however fleeting—often produce experiences that people find transformative and unforgettable (Curtin Reference Curtin2009). For example, human experiences of swimming with wild dolphins are reported to affect feelings of numinosity (Rames Reference Rames2016)—Jung’s (Reference Jung and Hull1966, 229) concept of the thrill of encountering the divine—and ‘peak experiences’—Maslow’s (Reference Maslow1976, 59–68) term for transcendent moments of joy that stand above other life experiences (Curtin Reference Curtin2006; DeMares Reference DeMares2000; DeMares & Krycka Reference DeMares and Krycka1998). On the other hand, human encounters with some animals, such as large carnivores, may invoke very different emotions and experiences; in addition to the above-mentioned work of grizzly–human encounters by Metcalf (Reference Metcalf2008), Collard (Reference Collard2012, 34–5) and Lowther (Reference Lowther2013) describe how encounters with cougars in the forests of the Pacific Northwest leave people feeling insecure and unsafe long after the moment of encounter. Of course, not every encounter necessarily produces a strong affective or cognitive shift; encounters may be essentially neutral rather than clearly positive or negative. Proximity and embodied experience are key to the way people perceive and remember encounters, but so are a variety of other factors, including the rarity of the animal species, whether one is seeing an animal for the first time, and the level of surprise and danger involved (Curtin Reference Curtin2010).
Wilson (Reference Wilson, Böhm and Ullrich2019, 25–6) describes a human–animal encounter as ‘a particular event of relation’ (emphasis in original), or an ‘unexpected moment’ with the potential to affect ‘shock’ and ‘rupture’. This framing draws from colonial and post-colonial studies, where ‘encounter’ is understood as an ‘engagement across difference’ (Faier & Rofel Reference Faier and Rofel2014, 363). In human–animal studies, ‘difference’ is often framed around encounters with rare, unusual, or dangerous animals (e.g. Blue & Alexander Reference Blue, Alexander, Gillespie and Collard2015; Gibbs & Warren Reference Gibbs and Warren2014; Keul Reference Keul2013; Lorimer Reference Lorimer2014; Metcalf Reference Metcalf2008; Muntifering et al. Reference Muntifering, Linklater and Naidoo2019; Schänzel & McIntosh Reference Schänzel and McIntosh2000). But this is not necessarily the case; Derrida (Reference Derrida2002) famously relates an unsettling encounter with his cat, and Haraway (Reference Haraway2008, 205) describes an agility training session with her herding dog as a ‘subject-shaping encounter’. Drawing from Deleuze’s (Reference Deleuze and Patton1994, 176) broader understanding of an encounter as something in the world that forces us to think or a sensory experience that affects emotions, Ohrem (Reference Ohrem, Calarco and Ohrem2018, 1) argues that the salient characteristic of human–animal encounters is their ‘potentially transformative influence on thought’, especially as the differences they reveal threaten ‘the workings of conceptual, spatial, and regulatory boundaries’. On the other hand, however, the categorical confusions engendered by human–animal encounters may sometimes be rejected, thus ultimately working to preserve the stability of boundaries (Wilson Reference Wilson, Böhm and Ullrich2019, 28–9; see also Todd & Hynes Reference Todd and Hynes2017, 735).
In the first half of this paper, I review recently published archaeological and historical evidence that the Florida manatee was so uncommon in precolonial Florida that Indigenous peoples would have encountered them eventfully and unexpectedly—perhaps as infrequently as every few hundred years. In the second half of the paper, I consider both the potential transformative, and stabilizing effects of such infrequent encounters—and their intervening absences—for precolonial Native Americans and (to the extent possible) for manatees.
Precolonial human–manatee relations as eventful and unexpected
More than 35 species of the order Sirenia have existed over the last 55 million years, but only four species in two families (Trichechidae and Dugongidae) are still extant (Reep & Bonde Reference Reep and Bonde2021). Sirenian fossils are common in deposits dating to the Oligocene (35–23 mya) and Miocene (23–5 mya) epochs, when much of what is now the Florida peninsula was covered by shallow seas. Fossil Trichechids of Pleistocene (2.5 mya–12,000 ka) age are known from several Florida localities (Domning Reference Domning1982, 604; Morgan Reference Morgan and Emry2002; Morgan & Hulbert Reference Morgan and Hulbert1995; Webb Reference Webb and Webb1974). However, the Florida manatee, a subspecies of the West Indian manatee, experiences cold stress syndrome from prolonged exposure to water below 20°C (Bossart et al. Reference Bossart, Meisner and Rommel2003; Hardy et al. Reference Hardy, Deutsch and Cross2019) and likely could not have survived the periodic episodes of cold air and meltwater that persisted along the Florida peninsula as late as 14,000 ka (Grimm et al. Reference Grimm, Jacobson and Watts1993). Native Americans began living in Florida near the end of the Pleistocene (Halligan et al. Reference Halligan, Waters and Perrotti2016) and Trichechid fossils have been interpreted as showing signs of human predation or butchering (Cumbaa Reference Cumbaa1980; Laist & Reynolds Reference Laist and Reynolds2005, 753–4; Powell & Rathbun Reference Powell and Rathbun1984, 20; Waller Reference Waller1970, 133). However, none are from secure archaeological contexts and Trichechids are not among the taxa (including mastodon, camelids and bison) represented in faunal assemblages from Late Pleistocene archaeological sites (Halligan et al. Reference Halligan, Waters and Perrotti2016; Hoffman Reference Hoffman1983; Smith Reference Smith2020; Reference Smith2022; Webb & Simmons Reference Webb, Simmons and Webb2006).
West Indian manatees likely migrated north to Florida after the last glacial episode (Laist & Reynolds Reference Laist and Reynolds2005, 752–3; Lefebvre et al. Reference Lefebvre, Marmontel, Reid, Woods and Sergile2001, 459; Reep & Bonde Reference Reep and Bonde2021). Although they can tolerate salt water for extended intervals, Florida manatees today live primarily in shallow (<4 m) tidal estuaries (Fig. 2), subsisting on a mix of freshwater, brackish and marine plants (Moore Reference Moore1951, 25; Reep & Bonde Reference Reep and Bonde2021, 44–61). Palaeoenvironmental studies suggest estuarine ecosystems developed in Florda as sea level rose during the middle Holocene around 5000 years ago (Jackson et al. Reference Jackson, Wang and Pluckhahn2023; Jones et al. Reference Jones, Wingard and Stackhouse2019; van Soelen et al. Reference van Soelen, Brooks and Larson2012; Yao & Liu Reference Yao and Liu2017), consistent with archaeological evidence for the earliest shell mounds and middens (Russo Reference Russo1991; Saunders & Russo Reference Saunders and Russo2011; Saunders et al. Reference Saunders, Wrenn, Krebs and Bryant2009)—as well as the earliest known non-fossilized manatee specimen from an archaeological context (as noted below).
Manatee. State Archives of Florida, Florida Memory. (https://www.floridamemory.com/items/show/271514).

The current population of manatees in Florida is estimated between 8000 and 12,000 (Gowan et al. Reference Gowan, Edwards and Krzystan2023). Although historical population levels are not well understood, this is likely far more manatees than at any point in Florida’s recent—or even more distant—past. The earliest aerial censuses in the 1970s put the total population of Florida and adjacent areas at around 700–800, distributed mainly in central and southern Florida (Hartman Reference Hartman1974; Irvine & Campbell Reference Irvine and Campbell1978). These observations are consistent with anecdotal accounts from the 1940s–50s (e.g. Kyle Reference Kyle1941; Moore Reference Moore1951). There is a common assumption that manatees may have been more common in Florida before human predation in the nineteenth and early twentieth centuries (e.g. Best & Hunt Reference Best and Hunt2020, 2–3; Carr & Milstein Reference Carr and Milstein2018; Jackson Reference Jackson2001, 5413; Laist & Reynolds Reference Laist and Reynolds2005, 740; Powell & Rathbun Reference Powell and Rathbun1984, 20). However, only a few studies have considered evidence for pre-modern manatee populations. Cumbaa (Reference Cumbaa1980, 9), based on a review of archaeological evidence, concluded that precolonial sea-cow populations were low and that Indigenous exploitation of manatees was ‘negligible’. O’Shea (Reference O’Shea, Odom, Riddleberger and Ozier1988, 190), drawing from Cumbaa and a review of historical sources, agreed that manatees had probably never been ‘extremely abundant’ in Florida, but suggested this was due to hunting pressure from both Indigenous peoples and later, colonial-era settlers.
Pluckhahn and Thulman (Reference Pluckhahn and Thulman2024) recently completed a more comprehensive review of historical baselines for the Florida manatee based on archaeological and archival sources. Their systematic sample consisted of 67 reports of archaeological investigations that included formal analysis of vertebrate fauna, with a combined total number of individual specimens (NISP) of nearly 2 million and total estimated minimum number of individuals (MNI) of almost 30,000. None of these reports described the identification of unfossilized manatee specimens.
Pluckhahn and Thulman (Reference Pluckhahn and Thulman2024) also surveyed the archaeological report database for opportunistic identifications of manatee bones, typically consisting of specimens modified into tools or ornaments (generally reported separately from other faunal remains) as well as unmodified bones tentatively identified as manatee without a formal zooarchaeological analysis. The opportunistic survey identified 12 reports of manatee or possible remains from archaeological sites, nine of which refer to sites dating to the precolonial era. However, three of the nine reported precolonial occurrences consist of fossils that likely predate human settlement of Florida; these fossils were likely collected by precolonial and later peoples, either as curiosities or as raw materials for the production of tools and ornaments (lithics suitable for flaking or grinding are uncommon across much of the Florida peninsula).
Figure 1 shows the locations of the six remaining precolonial sites with reported manatee bones. Three of these cases lack secure taxonomic identification or context (or both). Cumbaa (Reference Cumbaa1980, 9) described the identification of manatee bone in the faunal assemblage from the Tick Island site but provides no further information; Jahn’s (Reference Jahn1978, table 3) faunal analysis did not identify manatee. A ‘possible’ manatee bone was found at the Beacon Trade Port Midden (Janus Research, Inc. 1997, 27). Finally, there are reports of a ‘possible’ manatee bone tool at the Crystal River site (Bullen Reference Bullen1955, 30); reports of additional manatee specimens from this or nearby sites lack secure context (Cumbaa Reference Cumbaa1980, 9; Ellis et al. Reference Ellis, Dean and Martin2003, 10).
This leaves just three sites for which there are secure claims for one or more unfossilized manatee specimens from precolonial contexts. The earliest consists of a manatee rib bone modified for use as an atlatl handle found in the commingled burial of subadults at the Windover site, an Early (8000–6000 bce) to Middle (6000–3000 bce) Archaic period mortuary pond (Penders Reference Penders and Doran2002, 101); Penders suggested that—owing to the specimen’s large size—it may have served a ‘nontechnical purpose’. Next, Willey (Reference Willey1949, 43) identified a ‘large paddle-shaped implement’ manufactured from the scapula of a large animal—‘possibly a manatee’—from the Belle Glade site, dating to the Woodland period (1000 bce to 1000 ce); a photo of the artifact suggests that the taxonomic identification is likely accurate and that the specimen was not fossilized. Finally, two manatee rib bones modified for use as plummet-pendants are reported for the later Mississippian period (1000–1500 ce) occupation at the Granada site (Richardson & Pohl Reference Richardson, Pohl and Griffin1982, 129); the report does not indicate whether the bones were fossilized, but the photos suggest this was not the case. In sum, for the more than 13,000 years of Indigenous precolonial occupation of Florida, Pluckhahn and Thulman (Reference Pluckhahn and Thulman2024) were able to confidently identify only four manatee specimens from three precolonial archaeological sites; claims of manatee bones at three additional sites cannot be verified. This paucity is striking, considering the evidence for manatee exploitation by precolonial Indigenous peoples elsewhere in the circum-Caribbean (Bradley Reference Bradley1983; Colten et al. Reference Colten, Newman and Worthington2009, 79; Colten & Worthington Reference Colten and Worthington2019; Götz et al. Reference Götz, Valadez Azúa, Rodríguez Galicia, Muñoz, Götz and Ramos2014; McKillop Reference McKillop1984; Reference McKillop1985; Quitmyer Reference Quitmyer2003; Roca & Iglesias Reference Roca, Iglesias, Muñoz, Götz and Roca2014; Wilson Reference Wilson2007, 34, 141; Wing Reference Wing1973; Wing et al. Reference Wing, Hoffman and Ray1968).
Were this scarcity limited to the precolonial era, one might consider the possibility that the Indigenous peoples avoided killing or consuming manatees or perhaps took special precautions with the disposal of the remains of butchered sea cows. However, Pluckhahn and Thulman (Reference Pluckhahn and Thulman2024) also noted that manatees were not mentioned in historical accounts of Florida prior to the late 1700s, and only sparingly in the century that followed. Manatees are also not represented in faunal assemblages associated with colonial-era Spanish and British settlements in Florida (Pluckhahn & Thulman Reference Pluckhahn and Thulman2024), although they are reported for similar contexts in the circum-Caribbean (Baughman Reference Baughman1946, 235; Deagan & Cruxent Reference Deagan and Cruxent2002, 145–6; Deagan & Reitz Reference Deagan, Reitz and Deagan1995, table 9.3).
Comparison with the archaeological occurrence of several other animal species bolsters the interpretation that manatees were uncommon in Florida prior to the modern era. For example, faunal remains of Delphinidae spp. (dolphins and pilot whales) have been identified at more than 35 precolonial or early historic sites (Pluckhahn Reference Pluckhahn2025b), despite being much more difficult to hunt than manatees. Faunal remains of the Florida panther (Puma concolor cougar) have been identified from at least 15 sites (Wheeler Reference Wheeler2011, 140), despite its nocturnal and elusive behaviour. The occurrence of manatee specimens is more consistent with those of two animal taxa which were likely only occasional visitors to Florida or had very limited numbers and range before going extinct in the 1800s: the Great Auk (Pinguinus impennis), a flightless bird restricted mainly to the Northern Atlantic (identified at five sites) (Brodkorb Reference Brodkorb1960; Fradkin Reference Fradkin1980, 113; Hay Reference Hay1902; Kligerman Reference Kligerman2025, 45; McLean Reference McLean2019, 73; Weigel Reference Weigel1958) and the Caribbean Monk Seal (Monachus tropicalis), probably limited mainly to the Caribbean and extreme southern Florida (represented on six or seven sites) (Cockrell Reference Cockrell1970, 5; Cumbaa Reference Cumbaa1971, 6; DuChemin Reference DuChemin2018; Fradkin Reference Fradkin1976; Goggin & Sommer Reference Goggin and Sommer1949, 76; Quitmyer & Kennedy Reference Quitmyer and Kennedy2001, 16; Webb et al. Reference Webb, Wilson, Hale and Carr1993; Wing & Loucks Reference Wing, Loucks and Griffin1982).
One possibility is that manatees were not present at all in precolonial Florida and these few archaeological specimens represent material acquired through trade with Indigenous peoples elsewhere in the Caribbean. The fact that the better-documented specimens from precolonial archaeological contexts in Florida were fashioned into tools or ornaments may support this conjecture. Pluckhahn and Thulman (Reference Pluckhahn and Thulman2024) note that the form of the ‘paddle-shaped implement’ from the Belle Glade site (Willey Reference Willey1949, 43) resembles vomiting spatulas made of manatee bone from Taino sites in the Caribbean (e.g. National Museum of Jamaica 2024; Roca & Iglesias Reference Roca, Iglesias, Muñoz, Götz and Roca2014). However, claims for direct connections between Florida and the Antilles based on artifact similarities have been rightly criticized for lack of rigour (e.g. Bullen Reference Bullen1974; Griffin Reference Griffin1943, 87; Knight & Worth Reference Knight, Worth, Cordell and Mitchem2021).
Alternatively, manatees—either living or dead—may infrequently have washed ashore in Florida as they were displaced northward by strong winds and currents. Historically, manatee carcasses occasionally washed ashore in areas of northern Florida, attracting attention from residents who were unaccustomed to seeing them (Pensacola Daily News 1893). In the recent past, living manatees have been found up the Atlantic Coast as far as Rhode Island (Associated Press 2023) and up the Mississippi River as far as Tennessee (Tampa Tribune 2006), although they eventually succumb to cold stress at such northern latitudes.
More likely, manatees only occasionally extended their range north from the Caribbean to the Florida peninsula, owing to their susceptibility to cold stress and the episodic cooling associated with the Little Ice Age that began in the 1200s and continued to the 1800s (Pluckhahn & Thulman Reference Pluckhahn and Thulman2024). Isotope studies of corals, ostracods and planktonic foraminifera suggest sea surface temperatures in the Caribbean were 2–3 C° cooler at times during this interval (Cronin et al. Reference Cronin, Wingard and Dwyer2012, 257; DeLong et al. Reference DeLong, Flannery and Poore2014, 416; Nyberg et al. Reference Nyberg, Malmgren, Kuijpers and Winter2002, 35; Winter et al. Reference Winter, Ishioroshi and Watanabe2000) and sixteenth-century Spanish colonial accounts of Florida describe freezes that would be considered anomalous today (Hudson Reference Hudson1997, 190; White Reference White2018, 40–41, 68). The manatee’s expansion in numbers and range over the last century are probably owing to natural and anthropogenic climate change, the creation of warm-water refugia such as power plants, and changing human attitudes (Pluckhahn Reference Pluckhahn2025a; Pluckhahn & Thulman Reference Pluckhahn and Thulman2024).
Assuming the latter hypothesis is correct, current archaeological evidence suggests encounters between manatees and people in the precolonial era were extremely rare—perhaps only once every one or two millennia. Granting the fragmentary nature of the archaeological record, as well as sampling biases, we might imagine that such encounters were slightly more common—perhaps of the order of once every few centuries. Even at this rate, however, human and manatee encounters must have been unexpected.
Precolonial human–manatee encounters as transformative or stabilizing
Beyond their seeming infrequency, human–manatee encounters had the potential to be unsettling for Indigenous peoples because of the profound differences sea cows embody. Manatees, along with some other members of the order Sirenia, are the only herbivorous marine mammals. Unlike other marine mammals, but like many terrestrial ungulates, they have prehensile upper lips (for grasping food) (Reep & Bonde Reference Reep and Bonde2021, 50) and ‘rolling dentition’ (where new teeth form in the back of the jaw and move forward as old molars wear down and fall out) (Reep & Bonde Reference Reep and Bonde2021, 59). Unlike most mammals other than primates (as well as beavers, opossums, elephants and pinnipeds), manatees have fingernails (Reep & Bonde Reference Reep and Bonde2021, 51). Virtually alone among mammals, manatees lack eyelids and eyelashes (although they have nictitating membranes) and have six cervical vertebrae rather than seven (which requires them to turn their entire body to look behind them) (National Oceanographic and Atmospheric Administration 2025).
Indigenous peoples of precolonial Florida may also—or alternatively—have been unsettled by the manatee’s similarities to themselves; Sirenians are often considered anomalous because of their perceived anthropomorphic features. For the Torres Strait Islanders, McNiven (Reference McNiven and Watts2013, 110) describes a ‘blurred and fluid divide between humans and dugongs’ founded on an ontology of ‘shared and permeable personhood’ (McNiven Reference McNiven2010, 225). The Lio of the Flores Islanders refer to dugongs with a term that translates as ‘dugong people fish’, evincing their belief that the animals are ‘half human and half fish’ (Forth Reference Forth2020; Reference Forth2021). In the Maya area, manatee bones were sometimes fashioned into musical instruments and anthropomorphic figurines (Saint-Victor Reference Saint-Victor2024, 66–7), the latter sometimes interpreted as representations of the transformation of sea cows into humans (O’Neil Reference O’Neil2002, 80–81). Western societies of the modern era have also often seen Sirenians as anomalously human-like, as epitomized by the presumed (but poorly documented) historical association of manatee with mermaids (see Ritvo Reference Ritvo1997, 181).
These anomalous and anthropomorphic qualities made Sirenians the focus of special ritual attention for Indigenous peoples around the world (e.g. depositing them in shrines, employing them as charms or other ritual paraphernalia) (e.g. Forth Reference Forth2020; Reference Forth2021; Loveland Reference Loveland, Helms and Loveland1976; McKillop Reference McKillop1985; McNiven Reference McNiven2010; Reference McNiven and Watts2013; McNiven & Feldman Reference McNiven and Feldman2003; Méry et al. Reference Méry, Charpentier, Auxiette and Pelle2009; Saint-Victor Reference Saint-Victor2024). But if the manatee’s rarity of occurrence and curiosity of appearance made encounters with them a source of shock and rupture for the precolonial Indigenous peoples of Florida, the archaeological evidence for this is limited at best. Perhaps consistent with the attribution of special ritual status for sea cows, the better-documented archaeological specimens consist exclusively of tools or ornaments, as noted above. However, there are no definitive depictions of manatees in the extensive range of zoomorphic figures represented in wood, stone, bone, shell and ceramic art from precolonial Florida (see Wheeler Reference Wheeler1996). Footnote 1
In contrast, there are a few depictions of dolphins from precolonial archaeological contexts in Florida (Pluckhahn Reference Pluckhahn2025a; Wheeler Reference Wheeler2011). Elsewhere, I have suggested that Indigenous peoples in the region may have fished cooperatively with dolphins for mullet (Mugil spp.), as documented in similar environments elsewhere in the world (Daura-Jorge et al. Reference Daura-Jorge, Cantor and ingram2012; Neil Reference Neil2002; Simões-Lopes et al. Reference Simões-Lopes, Daura-Jorge and Cantor2016), based on the form of these artistic renderings and the fishing-related functions of the artifacts on which they appear (Pluckhahn Reference Pluckhahn2025a). I further suggested the possibility that dolphins may have been understood as sentient beings, based on depictions with eyes of exaggerated prominence—a convention that Wheeler (Reference Wheeler1996, 179) relates to an Indigenous belief that one of the three souls resided in the pupil of the eye.
Depictions of panthers are even more common in the art of precolonial Florida, occurring across artifacts of diverse form, function and materials (Wheeler Reference Wheeler2011, 143–7). The panther figures are typically naturalistic but sometimes exhibit anthropomorphic features (Bell Reference Bell2021, 62; Wheeler Reference Wheeler2011, 150). This diversity suggests the significance of panthers for Indigenous peoples of Florida may have been more variable, possibly including functions as clan progenitors, guardians and symbols of hunting prowess or curing (Wheeler Reference Wheeler2011, 151).
The absence of any artistic renderings of manatees is thus striking. Perhaps manatees were so rare, and considered so unusual by the Indigenous peoples of precolonial Florida, that they were understood in a way analogous to modern-day cryptids—animals of legend (such as sea monsters and sasquatches), sometimes of seeming supernatural character, whose existence is contested or questioned—as Forth (Reference Forth2021) has argued for the manner in which the contemporary Lio understand dugongs. Elsewhere in the southeast United States, at least in the Mississippian period, Indigenous people imagined and celebrated supernatural ‘monsters’ such as water cougars and horned serpents in artistic form (Knight Reference Knight and Galloway1989). But, although the precolonial Indigenous peoples of Florida occasionally depicted mythological creatures such as horned alligators (Wheeler Reference Wheeler1996, 152–3, 180–81), they seem to have preferred naturalistic portrayals of well-known animals. As Boyd (Reference Boyd2017, 310) observes, although every form of life creates itself in relation to others, this may rest on either the acknowledgement, or the erasure, of other existences; perhaps the omission of manatees from symbolic representation was a means by which the Indigenous precolonial peoples erased their encounters with sea cows from individual and collective memory.
Given the ambiguity of the archaeological record, we also cannot rule out the opposite possibility: Indigenous peoples of precolonial Florida may have managed to incorporate manatees into existing ontologies, avoiding any major structural disruptions. For example, the Seminole Indians of the late eighteenth and nineteenth centuries referred to manatees with a term said to translate as ‘big beaver’ (Bartram Reference Bartram1988), suggesting they may have conceived of sea cows as a sort of hypertrophic version of a better-known and beloved animal (see Williams & Jones Reference Williams, Jones, Pluckhahn and Ethridge2006). As Todd and Hynes (Reference Todd and Hynes2017, 735) observe, where encounters with animals can be reconfigured from a sense of difference to one of recognition, it may work to preserve, rather than unsettle ‘the stability of essences, categories, attributes and ideals’ (see also Wilson Reference Wilson, Böhm and Ullrich2019, 28–9).
Encounters may upset not only taxonomies, but also geographies. Barua (Reference Barua2015, 268) has described human-animal encounters as ‘an interweaving of trails, tracks and paths’ among heterogenous beings that territorialize in their own terms. Encounters with manatees may have precipitated a sort of ‘cartographic inversion’ for precolonial peoples of Florida, by alerting (or reminding) them of the presence or potential of unknown lands and seas.
Unfortunately, the archaeology record is also ambiguous with respect to how manatees conceived and remembered their encounters with precolonial Indigenous peoples. Scientific understanding of manatee cognition is underdeveloped, but recent studies suggest sea cows show relatively advanced abilities for learning, including cognitive spatial mapping and long-term memory (Bauer & Reep Reference Bauer and Reep2022; Henaut et al. Reference Henaut, Charles and Delfour2022). Perhaps the paucity of manatee bones on precolonial Florida archaeological sites represents the wariness of sea cows to engage with people, given memories of previous encounters. As Recht and Tsouparopoulou (Reference Recht, Tsouparopoulou, Recht and Tsouparopoulou2021:9) observe, we need more studies of animal agency in archaeology; however, for Florida manatees, and arguably for most of the animals in the ancient world, this will remain a challenge.
Conclusion
Human–manatee encounters are common in contemporary Florida. On cold days, human spectators gather to observe aggregations of manatees at springs, power plants and other warm-water refuges. Around 100,000 people annually visit the city of Crystal River to see or swim with sea cows (Sorice et al. Reference Sorice, Shafer and Ditton2006). Captive Florida manatees attract visitors to zoos and aquariums within the state and well beyond (Henaut & Delfour Reference Henaut and Delfour2023). Likenesses of sea cows adorn licence plates, mailboxes, T-shirts, stickers and plush toys. Of course, not all encounters are so positive. Although manatees manoeuvre to avoid watercraft (to the extent they are able), boat collisions account for one-quarter of reported manatee deaths (Rycyk et al. Reference Rycyk, Deutsch and Barlas2018). Over half of the encounters between human swimmers and manatees include some form of harassment by the former; sea cows are more likely than people to break off the interaction (King & Heinen Reference King and Heinen2004).
Archaeological evidence suggests that the precolonial Indigenous peoples encountered manatees much more eventfully than contemporary Floridians, probably as infrequently as every few generations (and perhaps far less often). Nevertheless, as Bille et al. (Reference Bille, Hastrup, Flohr Sørensen, Bille, Hastrup and Flohr Sørensen2010, 4) observe, that which is materially absent may still influences people’s experience of the material world. Regular concurrence in time and space may facilitate the establishment of deeper human–animal relationships (Oma Reference Oma2006, 201) but it is not a prerequisite for emotional connections, as evidenced by the attachments many modern Floridians feel for sea cows. But if precolonial peoples and sea cows established any sort of ‘intimacy without proximity’ (Metcalf Reference Metcalf2008, 115) there is little archaeological evidence for this, apart from a few manatee bones fashioned into tools and ornaments.
Unexpected and eventful encounters across difference still have the potential to cause shock and rupture (Wilson Reference Wilson, Böhm and Ullrich2019, 25–6) and, given the manatee’s many unusual characteristics of form and behaviour, we can imagine that the precolonial inhabitants of Florida found them as unsettling and anomalous as many people elsewhere in the world. Again, however, if sea cows were afforded any special ontological status, the archaeological record is largely silent on the matter. Perhaps sea cows were considered so discordant with existing ontologies that rendering them in symbolic form would have been a fraught endeavour; on the other hand, perhaps they were easily incorporated into existing categories of being, as the historic Seminole seem to have done by conceptualizing manatees as a ‘big beaver’.
At a minimum, the case of manatee and human interaction in precolonial Florida is a reminder that relational spaces of the past were not limited to those animals that are commonly found in middens. Haraway (Reference Haraway2008, 244), in describing humanness as an ongoing social processes involving encounters with other beings, famously emphasized that ‘becoming is always becoming with’—but the opposite may also be true: becoming is sometimes becoming both with and without.
Acknowledgements
An earlier version of this paper was presented at the meeting of the Society for American Archaeology in 2025; I thank Jessica Jenkins and John Henshaw for the invitation. This article benefited from the constructive comments of two anonymous reviewers.



 Open access
Open access