


Introduction
Tryptophan (Trp) is an essential amino acid that plays a crucial role in a wide range of physiological processes. It undergoes dynamic co-metabolism by both the host and the resident gut microbiota, resulting in a diverse array of bioactive metabolites with profound impacts on host health (Cervenka et al. Reference Cervenka, Agudelo and Ruas2017). The gut microbiota can convert Trp into a variety of metabolites, including indole, indole–3–lactic acid (ILA), indole–3–propionic acid (IPA), and indole–3–acetic acid (IAA) (Agus et al. Reference Agus, Planchais and Sokol2018; Xue et al. Reference Xue, Li and Zheng2023). Notably, ILA and IPA have been shown to exert beneficial effects, such as maintaining intestinal barrier homeostasis, enhancing intestinal immunity, and preventing metabolic diseases (Gao et al. Reference Gao, Sun and Li2025; Venkatesh et al. Reference Venkatesh, Mukherjee and Wang2014; Xie et al. Reference Xie, Cai and Lu2024; Xue et al. Reference Xue, Chen and Yu2022). Indole is the most abundant Trp metabolite and is generated by microbial tryptophanase activity, primarily from Escherichia coli (Li and Young Reference Li and Young2015). Excessive indole production has been associated with adverse effects, partly because it can be further converted into indoxyl sulfate (IS) and may also promote pathogen persistence or virulence under certain conditions (Darkoh et al. Reference Darkoh, Plants-Paris and Bishoff2019). Given this complex interplay between beneficial and deleterious Trp metabolites, identifying dietary strategies that can selectively channel microbial Trp catabolism toward the production of protective derivatives has become a major research focus (Li et al. Reference Li, Zheng and Zou2024; Qi et al. Reference Qi, Li and Yu2022; Sinha et al. Reference Sinha, Laursen and Brinck2024).
A number of studies have demonstrated that dietary fiber intake is well established to decrease intestinal transit time, which prevents the shift of colonic metabolic substrates from carbohydrate to protein and eventually reduces harmful protein-derived metabolites (Roager et al. Reference Roager, Hansen and Bahl2016). However, recent studies indicate that dietary fiber in the colon is able to directly remodel microbial amino acid metabolism, in particular Trp microbial metabolism (Sinha et al. Reference Sinha, Laursen and Brinck2024). An association analysis across five cohorts showed that higher dietary fiber intake was positively associated with increased IPA levels and negatively associated with IS concentrations (Qi et al. Reference Qi, Li and Yu2022). Dietary fiber, such as wheat bran and pectin, could modulate gut microbiota composition and microbial metabolism pathways, thereby shifting Trp catabolism toward the generation of health-promoting indole derivatives, such as ILA and IPA (Sinha et al. Reference Sinha, Laursen and Brinck2024; Yan et al. Reference Yan, Shi and Liu2023). Inulin, as a typical soluble dietary fiber, is a functional plant polysaccharide that can resist the digestive enzymes of mammals (Zong et al. Reference Zong, Liu and Deehan2024). An increasing number of studies have demonstrated that inulin’s ability to selectively enrich specific microbial taxa and alter the intestinal microenvironment is a key mechanism underlying its prebiotic properties (Han et al. Reference Han, Nam and Xu2021; He et al. Reference He, Shaoyong and Chen2025; Vandeputte et al. Reference Vandeputte, Falony and Vieira-Silva2017). For instance, research has shown that inulin can remodel the intestinal epithelium through interactions between the gut microbiota and immune cells (Corrêa et al. Reference Corrêa, Castro and Fachi2023). In addition, inulin has been reported to effectively limit the rise in gut microbiota-derived indole levels in peritoneal dialysis patients by modulating indole-producing bacterial populations (Li et al. Reference Li, Xiong and Zhao2020). Despite these findings, it remains unclear whether inulin directly modulates Trp metabolism and what specific microbial or host mechanisms are involved in this regulatory process.
To address these gaps, this study explores the effects of inulin supplementation, in combination with dietary Trp, on microbial Trp metabolism and gut barrier function under physiological conditions. We use a 2 × 2 dietary design in healthy mice, investigating whether the specific Trp metabolites produced by the intestinal community are affected by the availability of the substrate as well as by the presence of inulin in the gut environment, and whether this metabolic shift is associated with short-chain fatty acids (SCFAs) production, activation of barrier-related signaling pathways, and improved colonic epithelial features. Our findings provide novel evidence that inulin not only reshapes the gut microbial community structure but also reprograms microbial Trp catabolism toward the production of health-promoting indole derivatives, particularly IPA. This metabolic shift is further associated with increased SCFAs production and enhanced intestinal barrier integrity. These findings reveal a previously underexplored mechanism by which fermentable dietary fiber modulates host-microbiota metabolic interactions to promote gut homeostasis.
Materials and methods
Experimental animals
The study was conducted using 5-week-old male ICR mice, which were purchased from the Laboratory Animal Centre of Zhejiang University. Prior to the experiment, the mice were housed for 1 week to allow for acclimatization. Throughout the experimental period, mice were maintained under standardized conditions (22 ± 2°C, 50 ± 10% humidity, and a 12-hour light/dark cycle) with ad libitum access to food and water. The mice were fed a diet containing either 2 g/kg or 16 g/kg Trp, with or without the addition of 50 g/kg inulin, for a duration of 2 weeks. A total of 32 mice were used in this study and were randomly assigned into four experimental groups (n = 8 per group). The sample size was determined based on previous literature reporting similar dietary intervention studies in mice (Murga-Garrido et al. Reference Murga-Garrido, Hong and Cross2021). The dosing strategy was based on previous studies reporting their tolerability and physiological relevance in healthy murine models (Sinha et al. Reference Sinha, Laursen and Brinck2024). The specific diet compositions are described in Supplementary Table 1. Inulin was manufactured as a single lot and was provided by BENEO, with an average DP ≥ 23, a content of about 100%, and a sweetness level of 0% (Orafti® HP, Belgium). All mice were housed in the breeding room with specific pathogen-free (SPF) conditions at the Zhejiang University Animal Facility. The animal experiment was approved by the Laboratory Animal Welfare and Ethics Review Committee of Zhejiang University (Approval No. ZJU20240928) and conducted in accordance with ethical standards. During the experiment, feed intake was recorded. At the end of the experiment, mice were euthanized, and samples were collected immediately. Blood samples were collected by cardiac puncture, kept at room temperature for 30 minutes, and centrifuged at 3,000 × g for 15 minutes at 4℃ to obtain serum. Fresh fecal samples were collected and immediately snap-frozen in liquid nitrogen, then stored at −80℃. The entire colon was excised, and its length was measured to evaluate colonic injury. Approximately 0.5 cm segments of non-compressed intestinal tissues, liver, and kidney were collected and fixed in 4% paraformaldehyde for subsequent hematoxylin and eosin (H&E) staining. The remaining intestinal tissues, intestinal contents, and liver tissues were collected into cryotubes, snap-frozen in liquid nitrogen, and stored at −80℃ for further analysis.
IL-22 level detection
The levels of interleukin-22 (IL-22) were determined by enzyme-linked immunosorbent assay (ELISA) using corresponding kits, which were purchased from YOBIBIO (Cat: U96-3250E, Shanghai, China), according to the manufacturer’s instructions.
Histological analysis of tissue
The fresh tissue of liver, kidney, and colonic tissues was fixed in 4% paraformaldehyde, dehydrated, transparentized, paraffin-coated, and consequently segmented and stained through H&E (Cat: C0105M, Beyotime, China) and Alcian blue-periodic acid-Schiff (AB-PAS) according to the method described by Shaoyong et al. (Reference Shaoyong, Jin and Jiang2023). Immunohistochemistry was performed on 4 μm colon sections using an anti-rabbit Ki-67 primary antibody (Cat: A20018; ABclonal; 1:500), followed by incubation with an HRP-conjugated goat anti-rabbit secondary antibody (Cat: GB23303, Servicebio, 1:200) (Qiao et al. Reference Qiao, Yang and Deng2025). Representative photomicrographs were captured using a Leica DMIL microscope equipped with a DFC450C digital camera (Leica Microsystems, Germany).
RNA extraction and quantitative real-time PCR
Total RNA was extracted from the collected tissues using RNAiso Plus (Takara, Japan) according to the manufacturer’s instructions. The qPCR reactions were set up using the TB Green Premix Ex Taq II (Cat: RR820Q, TaKara, Japan) kit on a High Throughput Quantitative Fluorescent PCR Instrument (Bio-Rad, USA). The relative mRNA expression levels were determined using the 2−ΔΔCT method. The primer sequences for the amplification of each target gene are listed in Supplementary Table 2.
Western blot analysis
Total protein was extracted from colonic tissues using radioimmunoprecipitation assay buffer supplemented with 1% phenylmethylsulfonyl fluoride (Cat: BL1426A, Biosharp, China). Protein concentrations were determined with a BCA assay kit (Cat: A65453, Thermo, USA). Equivalent amounts of the extracted proteins were separated by SDS-PAGE electrophoresis and then electroblotted onto PVDF membranes (Millipore, Bedford, USA). Membranes were blocked with 5% bovine serum albumin solution to minimize nonspecific binding, followed by overnight incubation at 4°C with the following primary antibodies: aryl hydrocarbon receptor (AhR; Cat: AF6278, Affinity, 1:1500), UDP-glucuronosyltransferase 1-1 (Ugt1a1) (Cat: A1359, ABclonal, 1:2000), Cytochrome P450 1A1 (Cyp1a1) (Cat: CPA1307, Cohesionbio, 1:750). After washing with TBST buffer, membranes were incubated with goat anti-rabbit IgG-HRP secondary antibodies (Cat: ab7090, Abcam, 1:2000) at room temperature for 1 hour. The protein bands were visualized using an ECL assay kit (Cat: P0018M, Beyotime, China).
16S rRNA sequencing of fecal bacteria
Bacterial DNA was extracted from the fecal samples using a DNA isolation kit (DP328, TIANGEN) following the manufacturer’s instructions. The bacterial 16S rRNA genes were then amplified using the universal primers 27F (5′-AGRGTTYGATYMTGGCTCAG-3′) and 1492R (5′-RGYTACCTTGTTACGACTT-3′). The PCR amplification was carried out as reported by Zhang et al. (Reference Zhang, Yang and Zheng2021). The amplified products were then purified and pooled for paired-end sequencing on the Illumina MiSeq platform (Illumina, USA) by Majorbio (Shanghai, China) following standard protocols. The individual real biological sequences, referred to as amplicon sequence variants (ASVs), along with their frequencies. An ASV table was generated for abundance information and further analysis on MicrobiomeAnalyst 2.0. To assess taxa diversity and evenness, Shannon’s index, Chao1, and observed richness were calculated. The variability among sample groups was explored using principal coordinate analysis (PCoA) based on UniFrac distances. Statistical differences between microbial communities were tested using DESeq2 (Love et al. Reference Love, Huber and Anders2014). LEfSe analysis was conducted to identify ASVs exhibiting significant differences among experimental groups, using a linear discriminant analysis score threshold of 3.5 and a significance level of 0.05 for both the Kruskal–Wallis and Wilcoxon rank-sum tests (Segata et al. Reference Segata, Izard and Waldron2011).
Quantification of SCFAs in colonic contents
SCFAs were measured as previously described by Jin et al. (Reference Jin, Kalainy and Baskota2019). The SCFAs analysis was performed using an Agilent GC-MS 5975 equipped with a DB-WAX column (30 cm × 0.32 mm × 0.5 μm) (Agilent, CA, USA). A standard curve was used to quantify SCFAs, including acetate, propionate, isobutyrate, butyrate, isovalerate, and valerate.
Quantification of indole derivatives in serum and feces by UPLC-MS/MS in MRM mode
Trp catabolites from colon contents and serum samples were quantified by ultra-performance liquid chromatography coupled with tandem mass spectrometry in multiple reaction monitoring mode (UPLC-MS/MS in MRM mode). For the fecal samples, approximately 25 mg of accurately weighed material was transferred to 2 mL microcentrifuge tubes. Ice-cold 80% methanol (v/v, pre-chilled to −20°C) was added to each tube in a volume of 300 μL. The samples were then homogenized using a cryogenic grinder (Servicebio, China) at 50 Hz for 6 minutes at −10°C. This was followed by 40 kHz ultrasonication for 30 minutes at 5°C to further disrupt the samples. The samples were incubated at −20°C for 30 minutes to precipitate proteins, and the supernatants were collected after centrifugation at 13,000 × g for 15 minutes.
For the serum samples, 100 μL aliquots were transferred to 2 mL microcentrifuge tubes, and 300 μL of ice-cold methanol was added. The samples were vortex-mixed for 30 seconds, followed by the same steps of ultrasonic treatment, static incubation, and centrifugation as described for the fecal samples. The instrument used was a Thermo Scientific Dionex UltiMate 3000 UHPLC system (Thermo Fisher Scientific, USA). Chromatographic separation was conducted on an Acquity UPLC BEH C18 column (100 × 2.1 mm, 1.7 μm) maintained at 40°C. The mobile phase comprised 0.1% formic acid in water (solvent A) and 0.1% formic acid in methanol (solvent B), delivered at a flow rate of 0.4 mL/min. The gradient elution was programmed as follows: 10% B from 0 to 0.5 minutes, ramping to 95% B from 0.5 to 3 minutes, and returning to 10% B from 3 to 5 minutes. MS analysis was conducted using an electrospray ionization source in positive ion mode with MRM for detection. High-purity nitrogen was used as the nebulizing and drying gas, while high-purity argon was used as the collision gas. Instrumental parameters such as spray voltage and collision energy were optimized for maximum sensitivity. Details of the monitored ion pairs, cone voltages, and collision energies are provided in Supplementary Table 3. A sample volume of 8 μL was injected via a thermostated autosampler set at 4°C.
Statistical analysis
All data were presented as the mean ± standard error of the mean (SEM). Statistical differences among groups were analyzed using GraphPad Prism (GraphPad Software, CA, USA). Normality and homogeneity of variance were assessed prior to analysis. For data meeting the assumptions of normality and equal variance, two-way ANOVA was used to evaluate the main effects of dietary Trp level, inulin supplementation, and their interaction, followed by Fisher’s least significant difference test for multiple comparisons. For data that did not satisfy the assumptions for parametric analysis, differences among groups were analyzed using the Kruskal–Wallis test followed by Dunn’s multiple-comparisons test. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and NS means no significance. Adobe Illustrator software was used for drawing.
Results
Dietary administration of Trp and inulin induced no significant histopathological alterations in major Trp-metabolizing organs
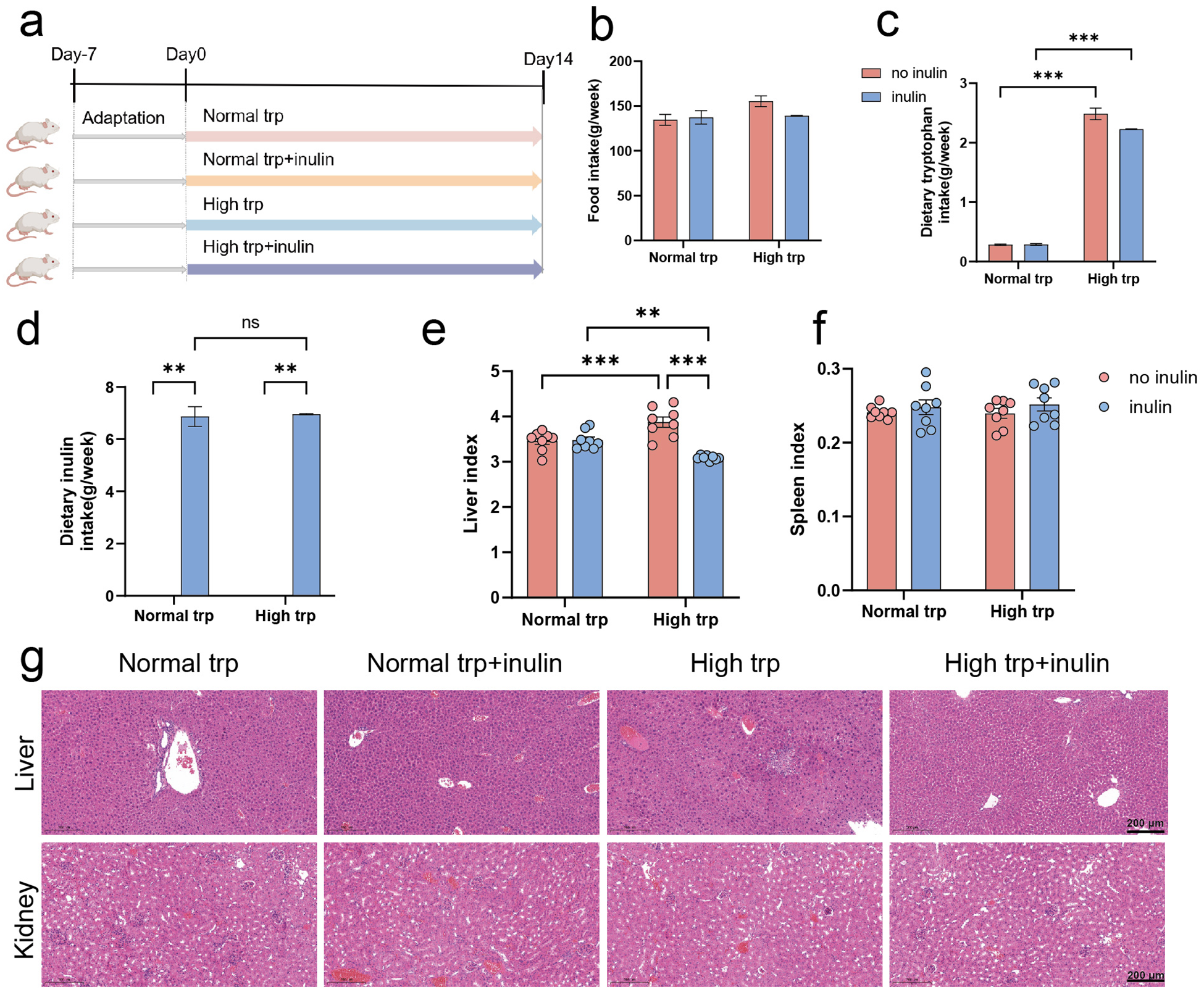
To ensure the safe and effective delivery of Trp to the colonic environment for its biological functions, while mitigating potential systemic metabolic diversion, we examined the histopathological impacts of a dietary intervention regimen on major Trp -metabolizing organs. To address this concern, four groups of SPF mice were fed for 2 weeks either a normal (2 g/kg) diet or a high-Trp (16 g/kg) diet, containing either no inulin or 50 g/kg inulin (Fig. 1a). Importantly, this dietary intervention strategy did not affect the total food intake (P > 0.05) (Fig. 1b). Quantitative analysis confirmed that the actual Trp intake in Trp-supplementation group reached 8–10 times the baseline levels of normal Trp group (P < 0.001) (Fig. 1c), while inulin intake was as expected in inulin-supplementation group (Fig. 1d).
Tryptophan and inulin supplementation do not induce histopathological alterations in the liver and kidney. (a) Schematic diagram of the experiment. (b) Food intake. (c) Dietary tryptophan intake. (d) Dietary inulin intake. (e) Liver index. (f) Spleen index. (g) Representative H&E-stained sections of liver and kidney tissues (scale bar: 200 μm). Data are presented as mean ± SEM. N = 8 biologically independent animals per group. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 1 Long description
The infographic presents the effects of tryptophan and inulin supplementation on mice. (a) Schematic diagram of the experiment shows four groups: Normal trp, Normal trp+inulin, High trp and High trp+inulin, over a 14-day period. (b) Bar graph of food intake shows no significant difference between groups. (c) Bar graph of dietary tryptophan intake indicates significant increases in the High trp groups. (d) Bar graph of dietary inulin intake shows expected levels in inulin-supplemented groups, with no significance in Normal trp. (e) Bar graph of liver index shows significant differences, especially in High trp groups. (f) Bar graph of spleen index shows no significant differences. (g) H&E-stained sections of liver and kidney tissues are shown for each group, with a scale bar of 200 μm. Data are presented as mean ± SEM, with statistical significance indicated by asterisks: < 0.05, < 0.01, < 0.001 and ns for no significance.
We further examined changes in the liver and kidney, which are the primary organs involved in Trp metabolism and immune function (Ronen et al. Reference Ronen, Gross and Ben-Shachar1996). As shown in Fig. 1e, under the no-inulin condition, mice fed a high-Trp diet exhibited a significantly higher liver index compared with those fed a normal Trp diet (P < 0.001). Notably, inulin supplementation significantly reduced the liver index in the high-Trp diet group (P < 0.001), whereas no obvious effect was observed under the normal-Trp condition (P > 0.05) (Fig. 1e). No significant differences were observed in spleen index across each group (P > 0.05), indicating no evident splenic enlargement or inflammation (Fig. 1f). H&E staining revealed extensive hepatocellular vacuolar degeneration around the central vein, portal area, and within the hepatic parenchyma in the high-Trp diet group, accompanied by mild infiltration of lymphocytes and granulocytes. In contrast, no such histopathological alterations were observed in the group receiving both high-Trp and inulin diet mice. These findings further supported the changes observed in the liver index. Kidney sections displayed well-organized renal parenchyma, with preserved glomerular and tubular morphology across all experimental conditions. No signs of tubular degeneration, interstitial fibrosis, or inflammatory cell infiltration were evident (Fig. 1g). High Trp intake may affect hepatic lipid metabolism, whereas inulin supplementation exerted a protective effect under high-Trp conditions.
Trp and inulin synergistically support colonic epithelial structure and barrier-related features
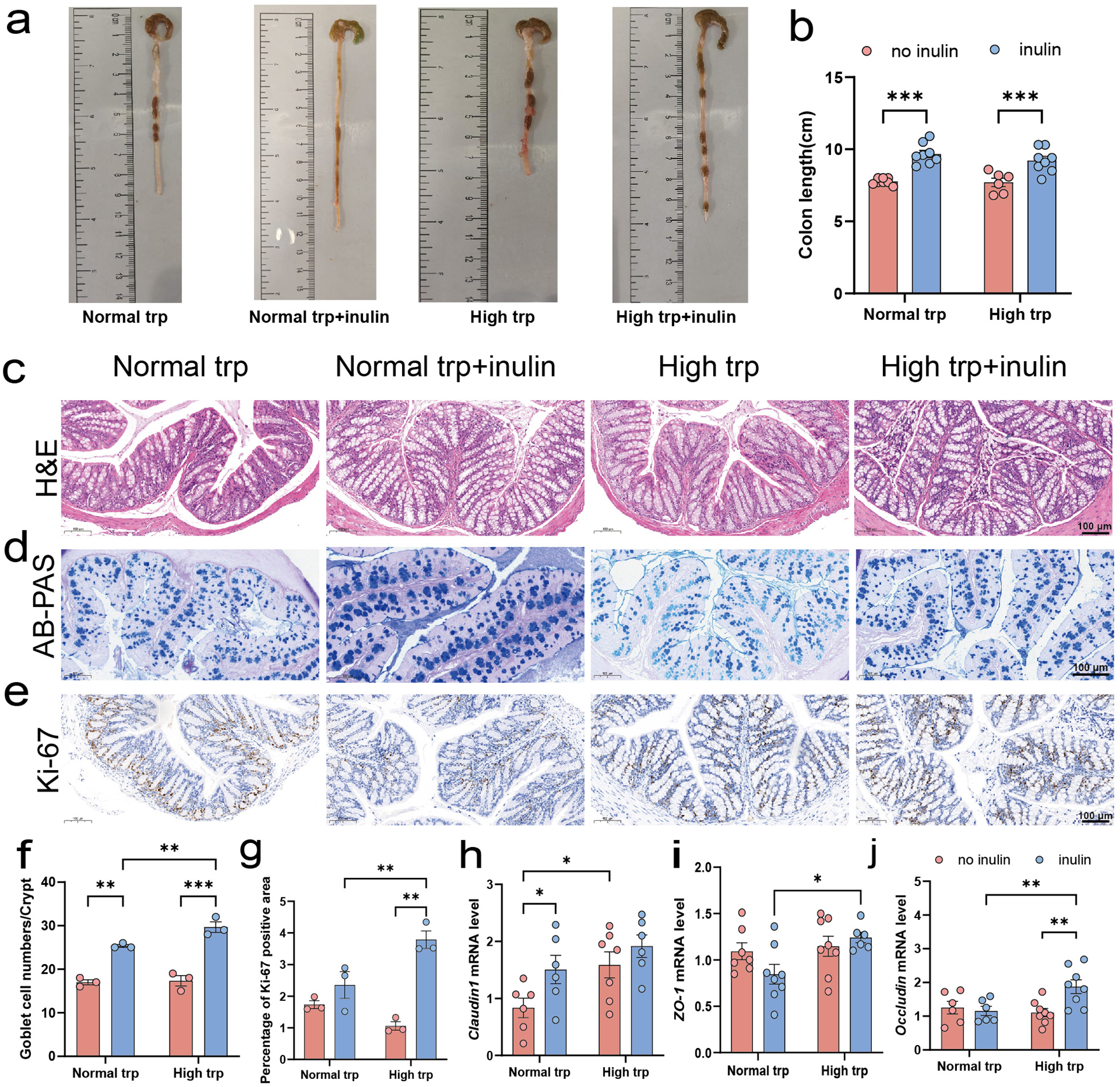
Given that Trp indole-related metabolism, along with inulin fermentation, predominantly occurs in the distal intestinal segments, we further investigated structural and functional changes specifically in the colon. Macroscopically, inulin supplementation significantly increased colonic length under both normal- and high-Trp diets (P < 0.001), while elevating dietary Trp alone did not markedly alter colon length (Fig. 2a, b). Moreover, H&E staining showed that inulin intake was associated with a more compact colonic architecture (Fig. 2c). AB-PAS staining further showed that inulin markedly increased goblet cell numbers per crypt under both Trp conditions, with a greater goblet cell abundance observed in the high-Trp and inulin diet group compared with the normal-Trp and inulin diet group (P < 0.01) (Fig. 2d, f). In the high-Trp diet group, the sections of the colon after inulin intervention showed a higher number of Ki-67 positive areas. Furthermore, under the condition of an inulin diet, increasing dietary Trp markedly increased Ki-67 positive area (P < 0.01) (Fig. 2e, g). At the molecular level, inulin increased the expression of Claudin1 under normal Trp condition (P < 0.05) (Fig. 2h) and a significant elevation in Occludin expression under high-Trp diet condition (P < 0.01) (Fig. 2j). Both ZO-1 and Occludin expression were elevated by high-Trp in the inulin condition (P < 0.05) (Fig. 2i, j). Collectively, these data indicate that inulin improves colonic architecture, mucus barrier features, and epithelial renewal, with more pronounced benefits under high-Trp condition.
Colonic morphological and barrier-associated changes in response to tryptophan and inulin supplementation. (a) Macroscopic images of mouse colons. (b) Colonic length (cm). (c) Hematoxylin and eosin-stained sections of colon (scale bars: 100 μm). (d) Alcian blue-periodic acid Schiff-stained colon sections (scale bars: 100 μm). (e) Immunohistochemical staining to detect the expression level of Ki-67 (scale bars: 100 μm). (f) Quantification of goblet cell density (cells per crypt). (g) Quantitative analysis of the Ki–67 positive area. (h–j) Relative mRNA expression levels of intestinal tight junction proteins: (h) Claudin1, (i) ZO-1, and (j) Occludin (normalized to β-actin). Data are presented as mean ± SEM. N = 6–8 biologically independent animals per group. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 2 Long description
This scientific figure illustrates colonic morphological and barrier-associated changes in response to tryptophan and inulin supplementation. Top row (a-b): Macroscopic images of mouse colons and a bar graph showing colonic length in centimeters. Inulin increases colon length significantly under both normal and high tryptophan conditions. Middle rows (c-e): Histological sections stained with H&E, AB-PAS and Ki-67. Inulin increases goblet cell numbers and Ki-67 positive areas, indicating enhanced cell proliferation. Bottom row (f-j): Bar graphs showing goblet cell density per crypt, Ki-67 positive area and mRNA expression levels of Claudin1, ZO-1 and Occludin. Inulin elevates Claudin1 and Occludin expression, especially under high tryptophan conditions. Data are presented as mean ± SEM, with statistical significance indicated by asterisks. The overall conclusion is that inulin improves colonic architecture and function, with more pronounced effects under high tryptophan conditions.
Inulin modulates fecal SCFAs profiles
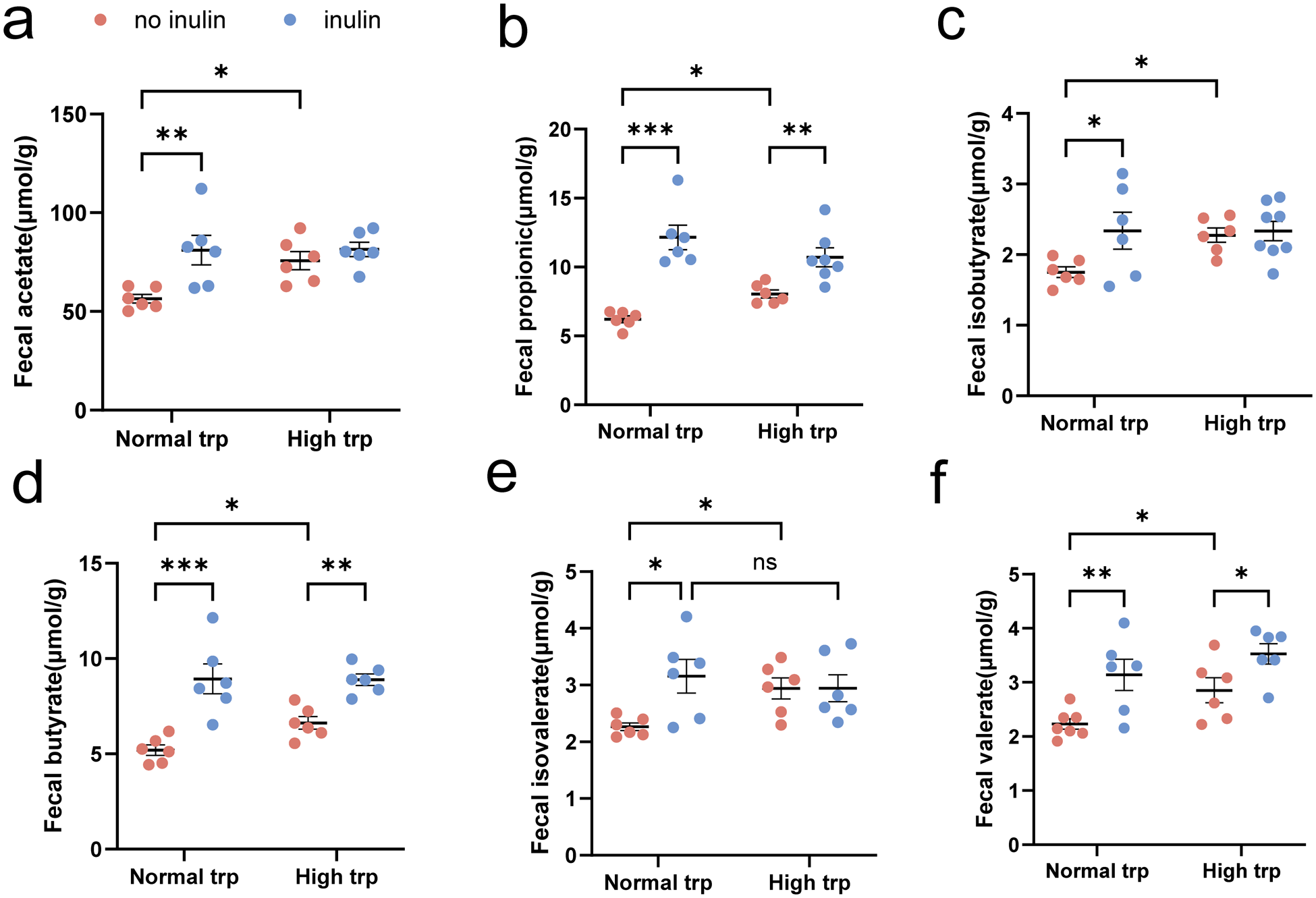
Previous studies have demonstrated that gut microbiota can degrade dietary fiber into SCFAs to exert pleiotropic physiological effects, including modulation of immune responses and maintenance of epithelial barrier integrity. We quantified fecal SCFAs concentrations across experimental groups (Fig. 3). Under normal-Trp diet condition, inulin supplementation markedly elevated all SCFAs, including acetate, propionate, isobutyrate, butyrate, isovalerate, and valerate (P < 0.05) (Fig. 3a–f). Under high-Trp conditions, however, the effect of inulin was more selective, remaining significantly elevated SCFAs only in propionate, butyrate, and valerate (P < 0.05) (Fig. 3b, d, f). In addition, in the absence of inulin, high-Trp treatment significantly increased acetate, propionate, isobutyrate, butyrate, isovalerate, and valerate compared with the normal-Trp diet (P < 0.05) (Fig. 3a–f).
Effect of tryptophan-inulin co-intervention on SCFAs levels of fecal contents. (a–f) Fecal contents concentrations of SCFAs: (a) acetate, (b) propionate, (c) isobutyrate, (d) butyrate, (e) isovalerate, and (f) valerate. Data expressed as mean ± SEM. N = 6–8 biologically independent animals per group. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 3 Long description
The image contains six scatter plots labeled a to f, each showing fecal SCFA concentrations under normal and high tryptophan conditions with and without inulin. Each plot has the x-axis labeled as Normal trp and High trp and the y-axis labeled with the specific SCFA in micromole per gram. A legend indicates no inulin and inulin. Plot a shows fecal acetate levels. Under normal trp, inulin increases acetate levels significantly compared to no inulin. Under high trp, acetate levels are higher than normal trp, with inulin showing a slight increase. Plot b displays fecal propionate levels. Inulin significantly increases propionate under both normal and high trp conditions. High trp shows higher levels than normal trp. Plot c illustrates fecal isobutyrate levels. Inulin increases isobutyrate under normal trp, with a smaller increase under high trp. Plot d shows fecal butyrate levels. Inulin significantly increases butyrate under both conditions, with high trp showing higher levels than normal trp. Plot e presents fecal isovalerate levels. Inulin increases isovalerate under normal trp, with no significant change under high trp. Plot f shows fecal valerate levels. Inulin increases valerate under both conditions, with high trp showing higher levels than normal trp. Each plot includes mean lines with error bars and significance brackets indicating statistical differences. The legend differentiates no inulin and inulin using distinct markers.
Modulation of microbial Trp metabolites by inulin and high-Trp interventions
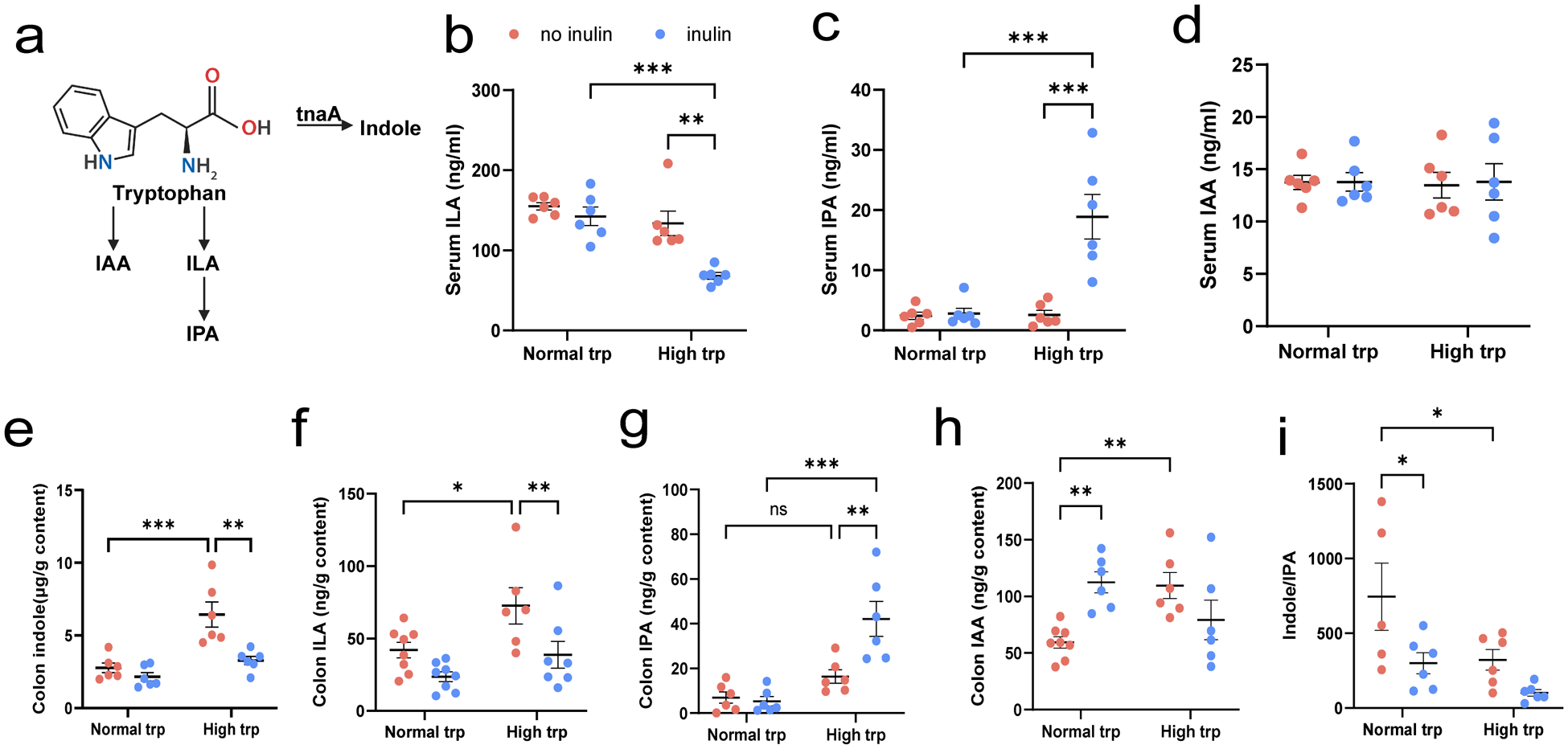
Gut microbial Trp catabolism is shown in (Fig. 4a). To elucidate the potential mechanisms underlying the observed protective effects of the combined high Trp and inulin intervention on the colon, we hypothesized that microbial metabolites derived from Trp catabolism may play a key role in modulating barrier integrity through interactions with mucin secretion and tight junction complexes. To test this hypothesis, we quantified the concentrations of specific Trp metabolites in both the colonic contents and the serum using UPLC-MRM.
Modulation of microbial tryptophan metabolites by high tryptophan and inulin interventions. (a) Schematic diagram of the tryptophan metabolism pathway of gut microbiota. (b–d) Serum concentrations of microbial tryptophan metabolites: (b) ILA, (c) IPA, and (d) IAA. (e–h) Colonic contents concentrations of microbial tryptophan metabolites: (e) indole, (f) ILA, (g) IPA, (h) IAA, and (i) ratio of indole/IPA. Data expressed as mean ± SEM. N = 6–8 biologically independent animals per group. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 4 Long description
The figure consists of multiple panels. Panel (a) is a schematic diagram of the tryptophan metabolism pathway in gut microbiota, showing the conversion of tryptophan to indole, ILA and IPA. Panels (b) to (d) display serum concentrations of microbial tryptophan metabolites: (b) ILA in micrograms per milliliter, (c) IPA in nanograms per milliliter and (d) IAA in milligrams per milliliter. Panels (e) to (h) show colonic contents concentrations: (e) indole, (f) ILA, (g) IPA and (h) IAA, all in micrograms per gram of content. Panel (i) presents the ratio of indole to IPA. Each dot plot compares normal and high tryptophan groups, with and without inulin intervention. Statistical significance is indicated with asterisks: for p less than 0.05, for p less than 0.01 and for p less than 0.001. The data are expressed as mean plus or minus SEM, with 6 to 8 biologically independent animals per group. Key findings include increased serum IPA and ILA levels in the high tryptophan group with inulin and significant changes in colonic indole and ILA concentrations.
The UPLC-MRM chromatograms of standards of ILA, IPA, IAA, and indole are shown in Supplementary Figure 1. The results showed that high-Trp treatment was associated with reduced ILA, and this decrease was further accentuated by inulin under the high-Trp condition (P < 0.01) (Fig. 4b). In contrast, serum IPA content was increased by inulin in the context of high Trp, resulting in markedly higher IPA levels in inulin plus high-Trp diet group compared with high-Trp diet alone or only inulin supplementation group (P < 0.001) (Fig. 4c). Serum IAA remained unchanged across each group (P > 0.05) (Fig. 4d). In colonic contents, high-Trp treatment increased indole accumulation (P < 0.001), whereas inulin supplementation significantly blunted this elevation under high Trp, bringing indole levels close to those observed in the normal-Trp groups (P < 0.01) (Fig. 4e). A similar pattern was observed for colonic ILA, high Trp increased ILA (P < 0.05), but inulin significantly attenuated ILA under high-Trp feeding (P < 0.01) (Fig. 4f). For colonic IPA, high Trp alone did not significantly increase IPA relative to normal Trp in absence of inulin (P > 0.05) (Fig. 4g). However, inulin supplementation robustly elevated IPA under high Trp, which significantly higher than high Trp alone or only inulin supplementation group (P < 0.01) (Fig. 4g). Colonic IAA was increased by inulin under normal-Trp diet and was also elevated by high Trp in the absence of inulin (P < 0.01) (Fig. 4h). Consistently, the indole/IPA ration was reduced by inulin under high-Trp diet; this difference did not reach statistical significance (Fig. 4i), indicating a shift in microbial Trp metabolism away from indole dominance toward increased IPA availability.
Trp and inulin supplementation reshape gut microbial composition
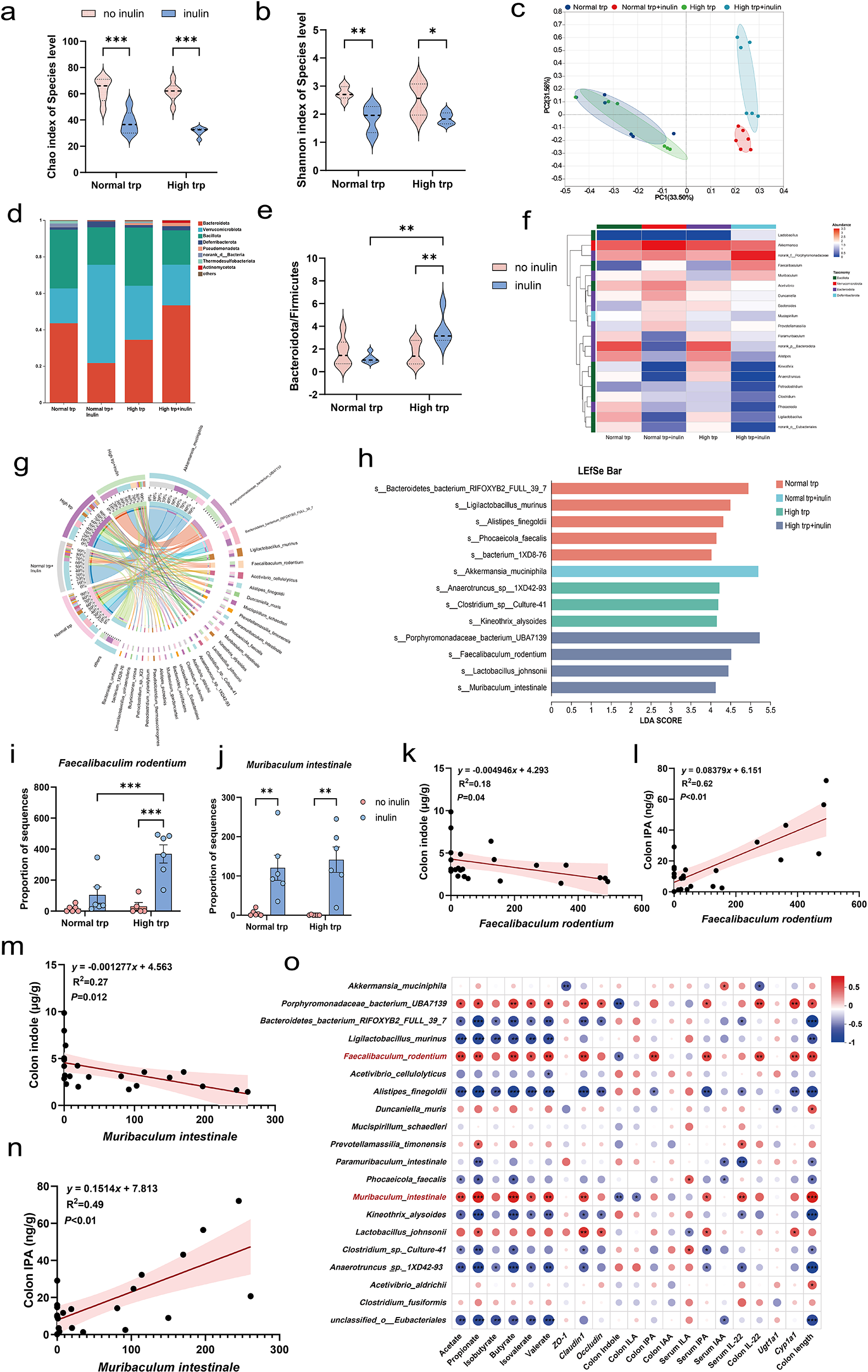
To investigate the specific regulatory effects of the high Trp and inulin co-administration on the gut microbiota, we performed 16S rRNA sequencing. The inclusion of inulin in the diet significantly reduced microbial α-diversity, as evidenced by decreased Chao, and Shannon indices under normal- and high-Trp diet conditions (P < 0.05) (Fig. 5a, b). PCoA further revealed that the inulin supplementation significantly restructured the gut microbial communities, whereas Trp intake alone induced no marked β-diversity shifts (Fig. 5c).
Tryptophan and inulin supplementation reshape gut microbial composition. (a–b) α-Diversity indices: (a) Chao and (b) Shannon index. (c) PCoA of β-diversity based on Bray–Curtis dissimilarity. (d) Phylum-level taxonomic composition (mean relative abundance >1%). (e) Ratio of Bacteroidota to Firmicutes; (f) Heatmap of genus-level relative abundance (top 20 genera). (g) Circos plot illustrating species-level interactions between experimental groups. (h) LEfSe analysis identifying differentially enriched taxa. (i–j) Relative abundance of (i) F. rodentium and (j) M. intestinale. (k) Negative correlation between F. rodentium abundance and colonic indole levels. (l) Positive correlation between F. rodentium and colonic IPA. (m) Negative correlation between M. intestinale and indole levels. (n) Positive correlation between M. intestinale and IPA levels. (o) Heatmap depicting Spearman rank correlations between microbial taxa showing significant intergroup differences and relevant host factors. Color gradient: red (positive correlation, ρ = 0.8) to blue (negative correlation, ρ = −1). Data expressed as mean ± SEM. N = 6 biologically independent animals per group. Statistical significance is indicated as follows:*P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 5 Long description
The infographic explores the impact of tryptophan and inulin on gut microbiota. Top row: Panels (a-c) show reduced α-diversity indices (Chao, ACE, Shannon) with inulin. Panel (d) illustrates PCoA, indicating distinct group separation. Middle row: Panel (e) displays phylum-level composition, highlighting shifts in Bacteroidota and Firmicutes. Panel (f) shows a decreased Bacteroidota to Firmicutes ratio with inulin. Panel (g) is a heatmap of top 20 genera, showing relative abundance changes. Panel (h) is a Circos plot illustrating species interactions. Panel (i) presents LEfSe analysis, identifying enriched taxa like Lachnospiraceae. Bottom row: Panels (j-k) show specific taxa abundance, with significant increases in Faecalibacterium and Muribaculaceae. Panels (l-o) depict correlations between microbial taxa and host factors, with positive and negative trends. Panel (p) is a heatmap of Spearman correlations, showing significant intergroup differences. Statistical significance is marked by asterisks, with < 0.05, < 0.01, < 0.001.
At the phylum level, the Bacteroidota/Firmicutes ratio was elevated by inulin in the context of high Trp group, which was significantly higher than high Trp alone or only inulin supplementation group (P < 0.01) (Fig. 5d, e). Genus- and species-level analyses further illustrated taxonomic shifts induced by the dietary interventions (Fig. 5f, g). LEfSe identified taxa specifically enriched by the combination diet. The inulin intervention selectively enriched Akkermansia muciniphila, while the high-Trp group exhibited a significant increase in the relative abundances of Anaerotruncus and Kineothrix alysoides taxa. Notably, in the high Trp and inulin combination group, Faecalibaculum rodentium and Muribaculum intestinale were enriched, along with increased abundance of Lactobacillus johnsonii, a species capable of metabolizing Trp into IPA (Fig. 5h-j). We performed linear regression analysis between specific microbial taxa and colonic IPA and indole levels. The results revealed that F. rodentium and M. intestinale were significantly positively correlated with colonic IPA concentrations (P < 0.01), while showing a significant negative correlation with indole levels (P < 0.05) (Fig. 5k-n). Spearman correlation analysis between microbial abundance and host phenotypic markers was presented in (Fig. 5o). Lactobacillus johnsonii exhibited a similar trend. In contrast, the relative abundances of Anaerotruncus and Kineothrix alysoides were positively correlated with indole levels but negatively associated with IPA production (P < 0.05). Additionally, F. rodentium abundance showed a positive correlation with the expression of tight junction-related genes and Cyp1a1 in correlation analyses. The microbiota alterations and metabolomic profiles observed in this study reveal concurrent changes in dietary context, microbial composition, and indole derivative levels, which are associated with epithelial barrier–related markers.
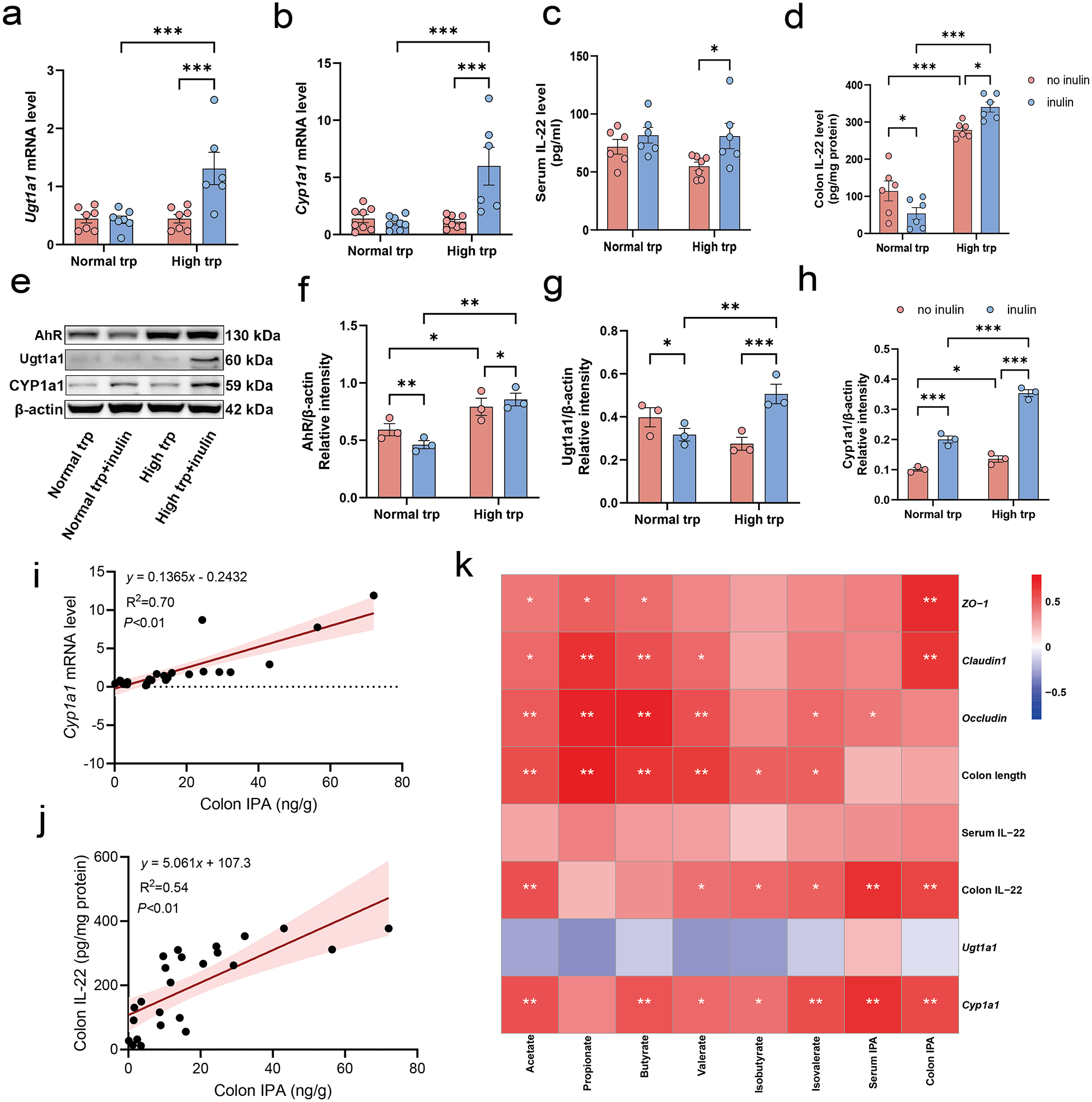
Associations between IPA levels and AhR/PXR/IL-22 pathway gene expression under dietary intervention
The results showed Ugt1a1 and Cyp1a1, which are downstream targets of the pregnane X receptor (PXR) and AhR pathways, respectively, markedly upregulated only when inulin was provided under high-Trp condition (P < 0.001) (Fig. 6a, b). The ELISA assay further showed that inulin significantly increased IL-22 under high Trp (P < 0.05), while the effect was modest under normal Trp (Fig. 6c). In the colon, high-Trp diet robustly increased IL-22 content (P < 0.001). However, inulin reduced colonic IL-22 under normal Trp (P < 0.05), yet further elevated higher IL-22 under high Trp than those between high Trp alone and only inulin supplementation group (P < 0.05) (Fig. 6d). Western blot analysis demonstrated that the combined high-Trp and inulin intervention significantly increased the protein levels of the PXR ligand Ugt1a1, AhR, and its target Cyp1a1 compared to the high-Trp alone or inulin diet alone group (P < 0.05) (Fig. 6e–h). Linear regression analysis revealed that colon IPA levels were significantly positively correlated with Cyp1a1 mRNA levels and colonic IL-22 expression (P < 0.01) (Fig. 6i, j). To explore potential associations between metabolite levels and markers of barrier function, we performed Spearman correlation analysis between IPA, SCFAs, and epithelial indices (Fig. 6k). Notably, colon IPA levels displayed strong positive correlations with colonic IL-22 expression and Cyp1a1 as well as tight junction proteins ZO-1 and Claudin1 (P < 0.01). These associations suggest a potential involvement of the AhR/PXR/IL-22 axis in the barrier-supporting effects observed in the high Trp-inulin group.
Diet-induced IPA is associated with gut barrier integrity and related host signaling pathways. (a–b) Relative mRNA expression of PXR and AhR target genes in colonic tissues: (a) Ugt1a1 and (b) Cyp1a1. (c–d) IL-22 concentrations measured by ELISA in (c) serum and (d) colon. (e–h) Representative Western blot images showing expression of key proteins in the AhR/Ugt1a1/Cyp1a1 signaling pathways in colonic tissues, accompanied by a quantitative analysis. (i) The linear correlation between colon IPA concentration and Cyp1a1 gene mRNA expression level. (j) The linear correlation between colon IPA concentration and colon IL-22 level. (k) Correlation heatmap illustrating associations among IPA, SCFAs concentrations, and biomarkers related to AhR/PXR/IL-22 signaling and intestinal barrier function. Data expressed as mean ± SEM. N = 3–8 biologically independent animals per group. Statistical significance is indicated as follows: *P < 0.05, **P < 0.01, ***P < 0.001, and ns means no significance.

Figure 6 Long description
The figure contains eleven panels labeled a through k. Panel a is a bar chart with the x-axis labeled Normal tryptophan and High tryptophan groups and the y-axis labeled Ugt1a1 mRNA level. Four bars are shown per tryptophan condition, split by no inulin and inulin groups. Triple asterisks indicate significance between inulin and no inulin under high tryptophan. Panel b is a bar chart with the same x-axis grouping and the y-axis labeled Cyp1a1 mRNA level. Triple asterisks indicate significance between inulin and no inulin under high tryptophan. Panel c is a bar chart with the x-axis labeled Normal tryptophan and High tryptophan and the y-axis labeled Serum IL-22 level in picograms per milliliter. Single asterisks mark significance between inulin and no inulin under high tryptophan. Panel d is a bar chart with the x-axis labeled Normal tryptophan and High tryptophan and the y-axis labeled Colon IL-22 in picograms per milligram protein. Asterisks mark comparisons between no inulin and inulin within each tryptophan condition and between tryptophan conditions. Triple asterisks appear between normal and high tryptophan no inulin groups. Panel e shows a Western blot image with bands for AhR at approximately 130 kilodaltons, Ugt1a1 at approximately 60 kilodaltons, Cyp1a1 at approximately 60 kilodaltons and beta-actin at approximately 42 kilodaltons. Four lanes correspond to Normal tryptophan, Normal tryptophan plus inulin, High tryptophan and High tryptophan plus inulin. Panel f is a bar chart with the y-axis labeled AhR divided by beta-actin relative intensity, quantifying AhR protein from the blot. Single and double asterisks indicate group differences. Panel g is a bar chart with the y-axis labeled Ugt1a1 divided by beta-actin relative intensity. Single and triple asterisks mark comparisons across groups. Panel h is a bar chart with the y-axis labeled Cyp1a1 divided by beta-actin relative intensity. Single and triple asterisks indicate significance across groups. In panels f through h, the x-axis groups are Normal tryptophan and High tryptophan, each split into no inulin and inulin bars. Panel i is a scatter plot with the x-axis labeled Colon IPA in micrograms per gram, ranging from 0 to 10 and the y-axis labeled Cyp1a1 mRNA level, ranging from approximately minus 10 to 10. A linear regression line is shown with the equation y equals 1.467 times x minus 0.6767, R squared equals 0.53 and P less than 0.01. Panel j is a scatter plot with the x-axis labeled Colon IPA in micrograms per gram, ranging from 0 to 10 and the y-axis labeled Colon IL-22 in picograms per milligram protein, ranging from approximately 0 to above 200. A linear regression line is shown with the equation y equals 31.7 times x plus 117.2, R squared equals 0.43 and P less than 0.01. Panel k is a correlation heatmap. The rows and columns are labeled Acetate, Propionate, Butyrate, Valerate, Isovalerate, Colon IPA, Serum IL-22, Colon IL-22, Ugt1a1 and Cyp1a1. The columns are labeled ZO-1, Claudin1, Occludin, Colon length, Serum IL-22, Colon IL-22, Ugt1a1 and Cyp1a1. The color scale ranges from minus 1 to plus 1, where darker shading indicates stronger correlation. Single and double asterisks within cells indicate statistical significance at less than 0.05 and less than 0.01 respectively. Strong positive correlations marked with double asterisks are visible between Colon IPA and Colon IL-22, Cyp1a1, ZO-1 and Claudin1.
Discussion
The major finding in our study showed that supplementation with Trp alone in the diet significantly elevated indole content, but co-supplementation with inulin altered the metabolic consequences of increased dietary Trp availability, which produced more IPA and less indole. Rather than simply increasing total Trp-derived metabolites, the combined intervention was associated with a shift in microbial Trp catabolism away from indole and toward IPA, together with improved goblet cell density, enhanced Ki-67 staining, upregulated tight junction-related genes, and higher expression of AhR/PXR/IL-22-related markers. These data support the concept that the physiological effect of dietary Trp depends on the surrounding fermentative environment created by co-administered fiber.
In agreement with our model, indole production has previously been shown to be inhibited in the presence of starch during batch fermentation using human fecal slurries (Smith and Macfarlane Reference Smith and Macfarlane1997), and pectin and inulin supplementation have been reported to decrease indole and increase ILA, IAA, and IPA accumulation in the medium of a batch culture inoculated with human feces (Huang et al. Reference Huang, Boekhorst and Fogliano2023). Sinha et al. further demonstrated, in defined microbial communities and in vivo, that pectin reduced indole and promoted ILA and IPA, and proposed a mechanism whereby fiber degradation releases fermentable sugars that repress tnaA expression in E. coli, thereby limiting Trp-to-indole conversion and potentially preserving luminal Trp for alternative microbial metabolic fates (Sinha et al. Reference Sinha, Laursen and Brinck2024). Although this mechanism was established using pectin-based systems rather than inulin directly, it may also be relevant to inulin because inulin is readily fermented by gut microbiota (Jung et al. Reference Jung, Bae and Song2025). In our study, inulin supplementation increased SCFAs levels under both normal and high Trp conditions, consistent with enhanced carbohydrate fermentation in the colon. In addition, the fiber-dependent rerouting of Trp metabolism may also be linked to the SCFAs content (Yang et al. Reference Yang, Su and Lv2023). This interpretation is supported by recent evidence that mildly acidic pH. The elevation of SCFAs content can reduce the pH level in the colon, which also strongly represses tnaA expression in E.coli and reduces indole level, accompanied by higher IPA level (Brinck et al. Reference Brinck, Laursen and Pedersen2026). Notably, in our study, while high Trp feeding increased SCFAs, the overall Trp metabolite profile still favored indole production, suggesting that SCFAs alone may not fully explain the rerouting of microbial Trp metabolism. Under inulin supplementation, IPA remained significantly higher in the high Trp group despite comparable SCFAs levels between normal and high Trp conditions, supporting the possibility that fermentable carbohydrate availability, rather than SCFAs abundance, may play a more prominent role in shaping microbial Trp metabolic routing.
Dietary fiber intake is associated with alterations in gut microbiota composition that coincide with increased production of IPA. Circulating IPA levels are positively associated with gut microbiome diversity and specific microbial taxa, indicating that gut microbiota composition is a major correlate of inter-individual variation in IPA (de Mello et al. Reference de Mello, Paananen and Lindstrom2017; Qi et al. Reference Qi, Li and Yu2022; Yamanaka Reference Yamanaka2020). LEfSe analysis identified F. rodentium, M. intestinale, and L. johnsonii as signature taxa in the combination group. Conversely, the high-Trp group was enriched in Anaerotruncus, Kineothrix, and Clostridium spp., with Anaerotruncus being particularly notable due to its ability to convert glucose and mannose into indole (Lau et al. Reference Lau, Woo and Woo2006). Strikingly, this indole-producing genus was not detected as a dominant taxon in the combination group, indicating that inulin supplementation was associated with a reduced abundance of bacteria linked to indole production. We observed a positive correlation between F. rodentium and M. intestinale with IPA concentration. The observed taxonomic shifts, including the marked dominance of F. rodentium in the combination group, align with prior reports demonstrating the capacity of this species to enhance intestinal immune surveillance, promote crypt cell proliferation, and accelerate epithelial turnover rates in murine models (Cao et al. Reference Cao, Bae and Villarreal2022). Furthermore, the reduction in indole levels in the combination group diminished the production of its nephrotoxic derivative, IS, which is consistent with the reported ability of L. johnsonii to ameliorate chronic kidney disease progression by attenuating uremic toxin accumulation (Miao et al. Reference Miao, Liu and Wang2024; Zhang et al. Reference Zhang, Nie and Sun2024). Faecalibaculum spp. have been associated with increased levels of AhR-activating metabolites and amelioration of metabolic dysregulation in murine models of colitis, obesity, and steatohepatitis (Liao et al. Reference Liao, Cao and Zhao2023; Pan et al. Reference Pan, Yang and Kulyar2025; Wang et al. Reference Wang, Hu and Wu2025). Similarly, Muribaculum abundance strongly positively correlated with IPA levels (Huang et al. Reference Huang, Liu and Wang2022; Wang et al. Reference Wang, Han and Liu2023). L. johnsonii has been reported to encode tryptophanase and aromatic amino acid transporters, which may be relevant to microbial Trp metabolism (Teixeira et al. Reference Teixeira, Harrison and da Silva2022). The replacement of indole-producing Anaerotruncus with these beneficial taxa in the combination group highlights the microbial specificity of inulin’s modulatory effects.
The host readouts further support the biological relevance of this metabolic shift. IPA and SCFAs were positively associated with markers related to epithelial barrier integrity, while the combination intervention increased colonic Cyp1a1 and Ugt1a1 expression and elevated IL-22 levels, findings that are consistent with activation of barrier-supportive pathways linked to microbial indole derivatives (Xie et al. Reference Xie, Cai and Lu2024). Given the positive correlations between colonic IPA and both Cyp1a1 and IL-22, IPA may represent one mechanistic link between diet-induced microbial remodeling and improved mucosal homeostasis. At the same time, SCFAs likely contribute complementary protective effects, as these metabolites are well known to support epithelial energy metabolism, mucosal immunity, and barrier maintenance (Yang et al. Reference Yang, Jiang and Zhou2025). Thus, the beneficial phenotype observed in the combination group may reflect coordinated actions of multiple microbiota-derived metabolites rather than a single effector alone.
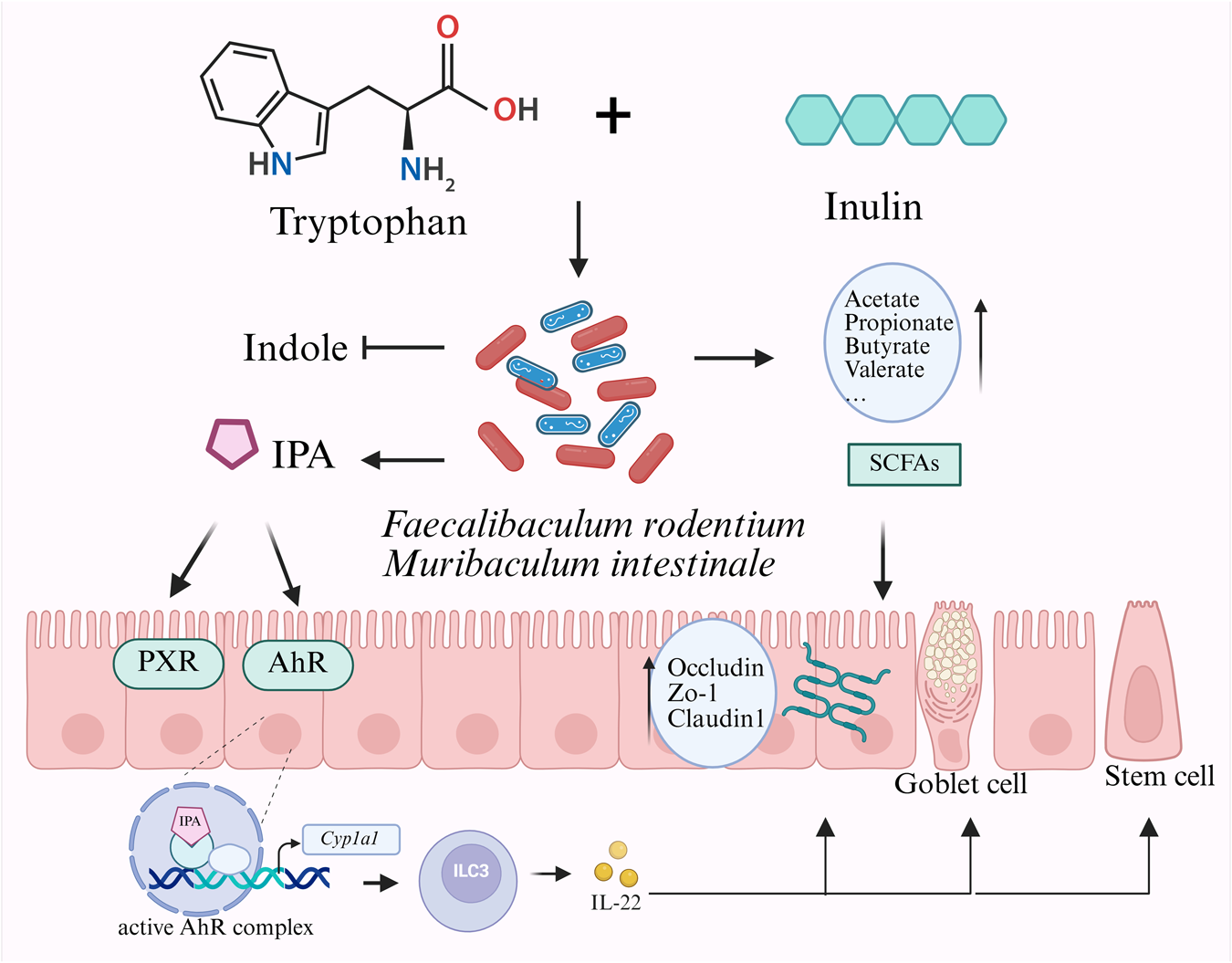
In conclusion, our findings suggest that dietary inulin is associated with modulation of microbial Trp catabolism toward IPA production and is related to improved epithelial barrier markers in healthy mice (Fig. 7). This study provides the concept that dietary fiber and amino acid interactions can reshape the gut metabolic landscape in favor of protective metabolites. Further investigations in inflammatory conditions and human cohorts are warranted to determine the therapeutic potential of this diet-microbiota-metabolite axis.
Interactive effects of dietary tryptophan and inulin on gut microbiota–mediated tryptophan metabolism and intestinal health. Dietary co-supplementation with tryptophan and inulin was associated with increased abundance of F. rodentium and M. intestinale, microbial taxa linked to enhanced IPA production and reduced indole levels. IPA and SCFAs showed positive correlations with key host markers related to the AhR/PXR/IL-22 axis and intestinal barrier function, suggesting potential microbiota-metabolite-host interactions contributing to gut homeostasis.

Figure 7 Long description
The diagram illustrates the interaction between tryptophan and inulin in the gut. Tryptophan and inulin are shown entering the gut microbiota, represented by various bacteria. This interaction leads to the production of indole and indole-3-propionic acid (IPA). The bacteria Faecalibaculum rodentium and Muribaculum intestinale are highlighted. Short-chain fatty acids (SCFAs) such as acetate, propionate, butyrate and valerate are produced. These compounds influence the intestinal epithelium, affecting receptors like PXR and AhR. An active AhR complex is shown interacting with Cyp1a1, leading to the production of IL-22. This pathway impacts the intestinal barrier, involving proteins like Occludin, Zo-1 and Claudin1 and affects goblet and stem cells.
Conclusion
In summary, our study demonstrates that the combined dietary supplementation of Trp and inulin is associated with coordinated shifts in gut microbiota composition and its metabolic outputs under physiological conditions. This combinatorial intervention was linked to altered Trp metabolism, favoring the accumulation of IPA over indole, along with enrichment of SCFAs such as butyrate and propionate. These microbial-derived metabolites showed positive associations with intestinal barrier-related indicators, including tight junction gene expression and goblet cell density, suggesting a potential contribution to mucosal homeostasis. Our study provides foundational evidence for the rational design of functional foods that target host-microbiota crosstalk through specific dietary components.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/anr.2026.10043.
Data availability statement
The 16S rRNA sequencing data from mouse feces have been deposited in the SRA database (https://www.ncbi.nlm.nih.gov/geo/) under accession number PRJNA1275593. The raw data and processed data required to reproduce these findings are available from the corresponding author upon request.
Acknowledgements
This research was supported by the fund from National Key Research and Development Program of China (2023YFD1301101, 2022YFD1300505), National Natural Science Foundation of China (32372889), Zhejiang Provincial Natural Science Foundation Program of China (LZ25C170001), Fundamental Research Funds for the Central Universities (226-2024-00080), Postdoctoral Fellowship Program of CPSF (GZB20240642), China Postdoctoral Science Foundation (2025M770268), Agriculture Research System of China (CARS-35), and National Center of Technology Innovation for Pigs.
Author contributions
Yan Wang: conceptualization, data curation, methodology, writing – original draft; Yanzhe Zhang: methodology, data curation; Yanli Chen: methodology; Siyu Wei: writing – review & editing; Yalin Liu: methodology; Yizhen Wang: supervision; Lei Qiao: methodology, writing – review & editing; Mingliang Jin: funding acquisition, supervision, writing – review & editing.
Conflicts of interest
The authors declare that they have no competing interests.



 Open access
Open access