



Impact statement
Edaphic habitat islands formed by special soils with extreme chemical or physical properties frequently contain rare species and unique biotic communities that are priorities for conservation. However, uncertainty in the degree of stability or change that can be expected in these communities as environmental conditions change (e.g., shifting precipitation patterns) complicates the development of conservation strategies. For example, divergent perspectives are that these communities should resist change by being stress-tolerant, or instead, they could be especially susceptible to change by already occupying habitats with extreme conditions. Our multi-decade study on special soils associated with gypsum in the Mojave Desert (Nevada, USA) revealed that portions of the flora (e.g., shrubs and the gypsum-associated perennial forb, Enceliopsis argophylla) were stable, despite climatic variability including drought encompassed by our three measurement years (2008, 2020, and 2025). Meanwhile, most perennial forbs had high turnover among our study sites through time, including two species of special conservation status (Anulocaulis leiosolenus and Arctomecon californica) that declined in abundance. From a standpoint of conservation of these unique communities, our results suggest stability of the shrub component and perennial community structure (e.g., distribution of cover among species, as transient species when turnover occurred were usually replaced by other transients to maintain community structure) by resisting change despite climatic variation. Declines in the two special-status species, however, suggest that further work to understand their persistence mechanisms (e.g., seed dormancy) and long-term dynamics could aid in differentiating cyclic population fluctuations from directional declines. More broadly, our results aid in understanding uncertainty in how communities on special soils may persist or change by showing that high degrees of stability and turnover can occur simultaneously across decades.
Introduction
Characterized by unusual chemical or physical properties, patchy distribution and limited extent, special soils form edaphic islands supporting endemic species and unique communities (Damschen et al., Reference Damschen, Harrison and Grace2010; Escudero et al., Reference Escudero, Palacio, Maestre and Luzuriaga2015; Ottaviani et al., Reference Ottaviani, Méndez-Castro, Conti, Zelený, Chytrý, Doležal, Jandová, Altman and Klimešová2022). Examples of special soils include those derived from serpentine, gypsum, carbonates and quartz, among others, along with rock outcrops with shallow soils (Cartwright, Reference Cartwright2019). The edaphic islands these special soils form within a landscape matrix of more typical soils support an estimated 20,000–40,000 vascular plant species that are edaphic specialists, or 5–10% of the global vascular flora (Corlett and Tomlinson, Reference Corlett and Tomlinson2020). Edaphic specialist plants are often considered to have unusually high stress tolerance or specialized adaptations to the unique habitats of edaphic islands (Palacio et al., Reference Palacio, Escudero, Montserrat-Martí, Maestro, Milla and Albert2007; Corlett and Tomlinson, Reference Corlett and Tomlinson2020). Accompanying the often rare and conservation-priority specialist species, unique soils can harbor distinctive genotypes of more widespread species, anomalous communities increasing beta diversity, and diverse ecological interactions such as plant–pollinator relationships, adding to the conservation significance of special soil habitats (Hulshof and Spasojevic, Reference Hulshof and Spasojevic2020).
Despite the conservation-priority status of many communities on special soils, several authors have noted that uncertainty is high regarding how these communities may respond to changing environmental conditions (e.g., drought) and that studies of long-term change in communities on special soils are rare and a research priority (Damschen et al., Reference Damschen, Harrison and Grace2010; Fernandez-Going et al., Reference Fernandez-Going, Anacker and Harrison2012; Sánchez et al., Reference Sánchez, Alonso-Valiente, Albert and Escudero2017; Coppi et al., Reference Coppi, Lazzaro and Selvi2022). One hypothesis is that communities on special soils should be relatively stable based on factors such as their species being stress-tolerant (and thus resistant or resilient to environmental changes) and that special soils are often infertile, reducing productivity to temper the magnitude and rate of change (Grime et al., Reference Grime, Fridley, Askew, Thompson, Hodgson and Bennett2008). Alternatively, communities on special soils are hypothesized to be highly dynamic and sensitive to changing environmental conditions (e.g., climatic changes or nutrient deposition) because special soils are already stressful environments at extremes (Damschen et al., Reference Damschen, Harrison, Ackerly, Fernandez-Going and Anacker2012). Thus, either an intensification or alleviation of stress could induce substantial change, as alleviating stress in the habitat may not be positive for edaphic specialists (Cartwright, Reference Cartwright2019). This raises the possibility that different groups of edaphic specialists and subsets of more widespread species able to inhabit special soils (albeit often at lower abundance) may exhibit different temporal changes on unique soils. An additional uncertainty for the degree of stability or change is that special soils are frequently sparsely vegetated. This low plant cover could interact with, or amplify effects of, environmental changes such as climatic warming or drought (Harrison et al., Reference Harrison, Damschen, Fernandez-Going, Eskelinen and Copeland2015). Further understanding the degree of stability or change in communities on edaphic islands could be considered particularly important during environmental change. Suitable habitat for edaphic specialists may not exist in other climatic regions, or species may be unable to disperse to suitable soils through matrix habitat (Damschen et al., Reference Damschen, Harrison, Ackerly, Fernandez-Going and Anacker2012; Corlett and Tomlinson, Reference Corlett and Tomlinson2020).
Here, we examined multi-decade change in plant communities affiliated with gypsum, a special soil with unique properties (e.g., abnormally high sulfur and calcium content and presence of physical surface crusts) and that harbors endemic and conservation-priority species globally (Escudero et al., Reference Escudero, Palacio, Maestre and Luzuriaga2015; Cera et al., Reference Cera, Montserrat-Marti and Palacio2023). We asked the following: Have species composition, diversity and structure (e.g., vegetative cover, shrub density) of gypsum-associated plant communities changed through time? What is the degree of temporal stability or turnover in species of different growth forms (e.g., forb, shrub) and association with gypsum soil?
Methods
Study area
We performed the study in Clark County, Nevada (USA), in the Mojave Desert (Supplementary Figure S1). This is a hot desert with a landscape matrix predominately of Larrea tridentata–Ambrosia dumosa shrubland (Meyer, Reference Meyer1986; Abella et al., Reference Abella, Prengaman, Embrey, Schmid, Newton and Merkler2012). On the western perimeter of our study area, the Las Vegas, Nevada, airport weather station (663 m in elevation) recorded an average of 10 cm/year of precipitation (60% falling in the winter–spring growing season from November through April) and daily average temperature ranges of 1–14 °C in January and 25–40 °C in July (1948–2025 records; National Oceanic and Atmospheric Administration, National Centers for Environmental Information, Asheville, NC). Our 10 study sites were chosen because they contained conservation-priority plant species affiliated with gypsum soil and were on public lands administered by the National Park Service (Lake Mead National Recreation Area) and the U.S. Bureau of Land Management (Abella et al., Reference Abella, Chiquoine and Vanier2013; Figure 1). Each site, ~ 1–5 ha in size, encompassed habitat for the native perennial forbs A. leiosolenus and A. californica, both listed as rare, special-status species by the Bureau of Land Management in Nevada and with additional conservation status under a Clark County Multi-Species Habitat Conservation Plan. A. californica is further listed as critically endangered by the State of Nevada and was reviewed for listing (but not listed in part from protections afforded by the conservation plan) nationally under the U.S. Endangered Species Act (U.S. Fish and Wildlife Service, 2024). The species are of conservation priority owing to their geographically restricted and patchy distribution on gypsum-affiliated soil in the northeastern Mojave Desert, habitat loss (e.g., expansion of urban Las Vegas, inundation by the filling of Lake Mead after completion of the Hoover Dam in the 1930s) and potential degradation of remaining habitat through various disturbances (Stosich et al., Reference Stosich, DeFalco and Scoles-Sciulla2022; Bailey et al., Reference Bailey, Pereira, Sion, Kobelt, Gentilcore, Antoninka and Bowker2023). All of our study sites were on public lands under protection management with minimal known disturbance, aside from a possibility of occasional feral livestock (e.g., Equus asinus) passing through.
Repeat photos from the same locations showing plant communities among three measurement years within an 18-year period on four example plots associated with gypsum soils, Mojave Desert, Nevada, USA. Photos were taken in spring (mid-April to early May) by SRA in 2008 and by University of Nevada, Las Vegas staff in 2020 and 2025.

Data collection
In each of the 10 sites, we randomly located a 10 m × 10 m (0.01 ha) plot. Plots spanned an extent of 59 km, averaged 21 km apart, were within gypsum-affiliated soil map units averaging 838 ha in size and were separated from each other by matrix habitat predominately of L. tridentata-A. dumosa shrubland (Supplementary Table S1, Supplementary Figure S1). Elevation among plots ranged from 383–634 m, and topography was flat to rolling with an average slope gradient of 4%. Parent material of soils among plots was alluvium or residuum primarily derived from gypsum and including limestone, sandstone or siltstone. At the great group level of U.S. soil taxonomy, soils were mapped as Petrogypsids, Haplogypsids, Calciargids, Torriorthents and unclassified gypsum badlands (Lato et al., Reference Lato, Merkler, Lugo and Harrington2006). Surface soils (0–5 cm mineral soil, with little to no O horizon) had concentrations of 1–6% sulfur, 820–1456 μg/g calcium and 0.2–3.4 μg/g magnesium among plots, detailed in Supplementary Table S1.
We inventoried vascular plants near the peak spring growing season between mid-April and early May in 2008, 2020 and 2025. These measurement years represented a range of precipitation in the winter–spring, hydrological-year growing season. Using the Las Vegas Airport station, precipitation from November of the prior calendar year through April of the current year was 59% of average in 2008, 180% in 2020 and 25% in 2025. Some annual plants emerged in the below-average precipitation 2008, mass germination occurred in the wet 2020, and no annuals emerged on our plots in the dry 2025. Thus, the temporal context of our study represents moderate, good and poor years in terms of annual plant emergence as a contingency factor for variability among years. Meanwhile, directional trends may be detectable for the longer-lived perennials with maximum life spans of individuals exceeding 100 years (e.g., A. dumosa, Atriplex confertifolia and Ephedra torreyana; Bowers et al., Reference Bowers, Webb and Rondeau1995).
In each plot each year, we categorized aerial cover of live plants by species using cover classes, assigning 0.01% for trace and the class midpoint to convert to percent cover (Peet et al., Reference Peet, Wentworth and White1998). We also counted the number of individuals on each plot for the two special-status species (A. leiosolenus and A. californica) and adults (≥ 15 cm tall and branching present) for four medium–large shrub species (A. dumosa, E. torreyana, L. tridentata and Psorothamnus fremontii). After we observed dieback or leaf loss of perennials following severe mid-2020 through mid-2022 drought (Supplementary Figure S2), we recorded average dieback of each perennial species on each plot in 2025. Dieback was the percent of a plant’s canopy (recorded to the nearest 5% and ranging from 0–100%) in vertical projection that had dead leaves, dead branches or leafless branches. Plants with 100% dieback were not necessarily dead, such as the drought-deciduous A. dumosa, which can leaf out following unknown numbers of leaf-off years (McAuliffe and Hamerlynck, Reference McAuliffe and Hamerlynck2010). Species taxonomy and classification by growth form (e.g., perennial forb) and nativity to the U.S. follow Natural Resources Conservation Service (2025).
Data analysis
To address our study questions, we performed multivariate community analyses in PC-ORD 7.11 and univariate analyses in PAST 5.2.2. We examined variation among years (2008, 2020 and 2025) in multivariate species composition (relative cover as cover of speciesi/∑ cover of all species on a plot) for vascular plants (annuals and perennials; no biennials were present) and for perennials only using nonmetric multidimensional scaling (Sørensen distance, random starting coordinates and 250 runs with real and randomized data). We tested the null hypothesis of no difference in species composition across years using blocked multi-response permutation procedures (Euclidean distance), including plots as repeated-measures blocks.
Next, we calculated a series of univariate variables (Supplementary Table S2): native cover (divided into annuals and perennials) and species richness (100 m2), non-native cover (all were annuals), density (plants/ha) of the two special-status species and the four major shrub species, the Shannon and Simpson diversity indices (Kunakh et al. Reference Kunakh, Volkova, Tutova and Zhukov2023; based on percent cover and computed in PC-ORD 7.11), community evenness (Shannon diversity/ln(species richness)), and cover of native species classified according to their typical distributional affiliation with gypsum. This classification, based on Meyer (Reference Meyer1986) also in the northeastern Mojave Desert, included gypsophile (closely associated with gypsum and soils intermixed with gypsum), gypsocline (principally associated with gypsum soil but also other unique soils and are not regionally dominant species) and gypsovag (regionally common on gypsum and non-gypsum soil). We compared means of univariate variables among years using repeated measures (plots as subjects), permutational analysis of variance (9,999 permutations), followed by pairwise comparisons (Bonferroni-adjusted sequentially) when P < 0.05. For dieback measured in 2025, we calculated means and standard errors of means by species.
We assessed change in species occurrences and community structure by calculating turnover and species rank-abundance curves. Following Milberg and Hansson (Reference Milberg and Hansson1993), we calculated turnover for species based on presence/absence on each plot between 2008 and 2020 and between 2020 and 2025. Turnover, ranging from 0–100%, was calculated as: ((number of gains and losses)/(number of gains, losses, and persistence instances)) × 100. Based on cover for each species averaged across the 10 plots, for each year we calculated a community rank-abundance curve in PAST 5.2.2 modeled using the geometric series (Matthews and Whittaker Reference Matthews and Whittaker2014). After log-transforming cover to enable linear regression of species rank-abundance relationships, we used analysis of covariance to compare slopes of the regressions to test if community structure changed among years (Fattorini, Reference Fattorini2005).
Results
In total, on the 10 plots for the three measurement years (2008, 2020 and 2025), we recorded 65 plant species, including 37 annuals (33 native, 89%) and 28 perennials (all native). We recorded 35 (16 annual, 19 perennial) species in the below-average precipitation year of 2008, 56 (32 annual, 24 perennial) in the wet 2020 and 22 (all perennial, as annuals did not emerge) species in the dry 2025 (Table 1, Supplementary Table S3). Of the perennial species, three were gypsophiles, classified as closely associated with gypsum soil, including the forbs A. leiosolenus and E. argophylla and the shrub Petalonyx parryi. Another perennial, the forb A. californica, was a gypsocline principally associated with gypsum. Among native annuals, two forbs were gypsophiles (Phacelia palmeri and Psathyrotes pilifera) and three were gypsoclines (Camissonia multijuga, Eriogonum insigne and Phacelia pulchella).
Variation in cover, frequency and turnover within an 18-year period including three measurement years (2008, 2020 and 2025) for 28 native perennial species among 10 gypsum-affiliated sites in the Mojave Desert, Nevada, USA

Note: In parentheses after species names, species are classified based on their association with gypsum following Meyer (Reference Meyer1986): gypsophile (gp): tightly associated with gypsum; gypsocline (gc): principally associated with gypsum but also inhabits other unique substrates and is not a regional dominant in the Mojave Desert; and gypsovag (gv): common on gypsum and non-gypsum soils. Species without designations were not classified by Meyer (Reference Meyer1986). Cover is aerial cover, averaged for 10 plots. Frequency is out of 10 plots (− signifies species is absent). Gain, loss and persist represent the number of plots (out of 10) colonized, vacated or consistently inhabited between study years. Turnover is calculated as: ((number of gains and losses)/(number of gains, losses and persistence instances)) × 100 (Milberg and Hansson Reference Milberg and Hansson1993). Cover, frequency and turnover for the 37 annual species detected during the study are in Supplementary Table S3.
In ordinations of community composition based on relative cover for all species and for only perennials, no consistent directional change was evident within the 18-year period (Figure 2). Instead, most plots exhibited minimal change or no clear trajectory among measurement years. Consistent with the ordinations, inferential permutational analysis revealed no significant change (P > 0.39) in species composition among years (Figure 2). Sørensen similarity of each plot to itself through time averaged at least 53% among plots for all comparisons, including 2008:2020, 2008:2025 and 2020:2025.
Nonmetric multidimensional scaling ordinations of species composition (relative cover) of all vascular plant species and only the 28 perennial species within an 18-year period in plant communities associated with gypsum soils in the Mojave Desert, Nevada, USA. In each ordination, each point is a plot (n = 10), with its temporal change among measurement years (2008, 2020 and 2025) connected by lines. Centroids represent the average species composition of each year. The ordination for all species represented 68% of the variance in the first two axes (58% for Axis 1 and 10% for Axis 2). The ordination for perennials represented 74% of the variance (60% for Axis 1 and 14% for Axis 2). The inset tables show tests of the null hypothesis of no difference in species composition (relative cover) among years using blocked multi-response permutation procedures (BMRPP), along with the average similarity of each repeated measures subject (plot) to itself through time.

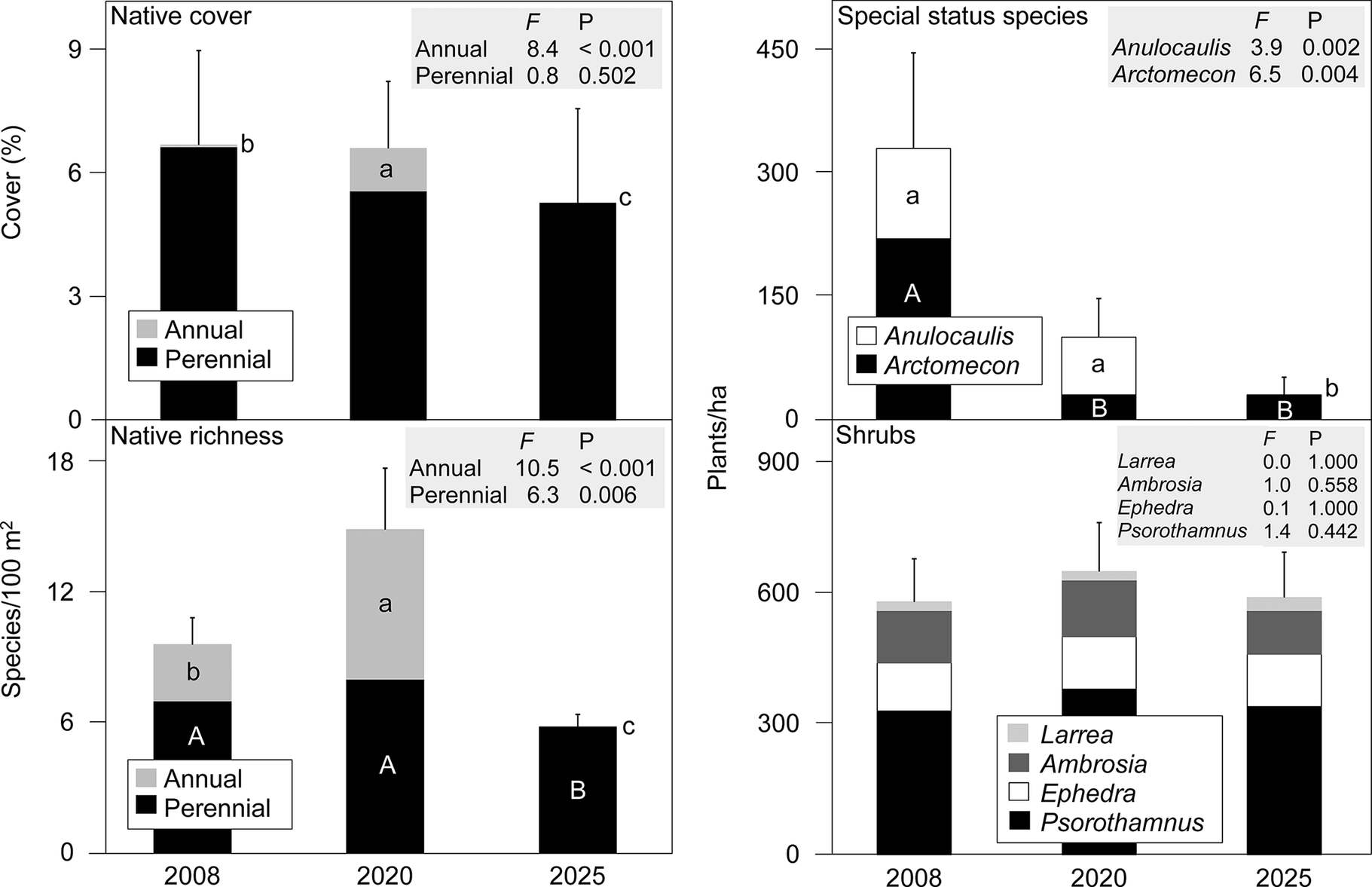
Univariate, native plant community characteristics differed in their degree of variation among years (Figure 3). Perennial plant cover did not change significantly across years while perennial species richness was lowest in 2025. Native annual cover and richness were highest in 2020, intermediate in 2008 and absent in 2025. Density of the two special-status species, A. leiosolenus and A. californica, in total exceeded an average of 300 plants/ha in 2008 but subsequently declined. After averaging 110 plants/ha in 2008 and 70/ha in 2020, no live A. leiosolenus inhabited our plots in 2025. A. californica declined in density by 86% between 2008 and 2020 and remained unchanged at 30 plants/ha between 2020 and 2025. Density of live, adult shrubs was nearly stable, and none of the four large shrub species displayed significant temporal variation. In 2025 when we recorded dieback, dieback averaged 50% (SEM = 3%) across perennial species among the 10 plots. Among the dominant shrubs, P. fremontii exhibited less than half the dieback at 24% than any other shrub species, all of which had ≥48% dieback.
Temporal variation in characteristics of plant communities associated with gypsum soils among three measurement years within an 18-year period in the Mojave Desert, Nevada, USA. Bars are means, and error bars are +1 standard error for mean totals. Sets of uppercase and lowercase letters display separate pairwise comparisons (Bonferroni-adjusted sequentially) of means within categories of repeated measures; permutational analysis of variance (pseudo-F 2,18 and P shown for each variable) had P < 0.05. Species are abbreviated by genus and include A. leiosolenus, A. californica, L. tridentata, A. dumosa, E. torreyana and P. fremontii.

In addition to those in Figure 3, other univariate community metrics similarly showed varying degrees of temporal change among study years (Supplementary Table S4). Cover of non-native plants (all of which were annuals) followed the same pattern as native annuals, being highest in 2020, intermediate in 2008 and absent in 2025. Community evenness did not differ among years, while diversity indices were lower in 2025 (which lacked annuals) than in either 2008 or 2020. Among categories of native plant species classified according to edaphic specialization, cover of gypsophiles, closely affiliated with gypsum, and gypsovags, regionally common on gypsum and non-gypsum soil, did not vary significantly among years. However, cover of gypsoclines (principally associated with gypsum but also other unique soils) differed among all years, being highest in 2020 (mostly via annuals), intermediate in 2008 and lowest in 2025.
Temporal turnover of perennial species was sharply higher for forbs as a group than for shrubs (Table 1). Turnover for the 13 perennial forb species was 56% between 2008 and 2020 and 62% between 2020 and 2025. In comparison, turnover for the 13 shrub species was 23% between both sets of years. Among shrubs, P. fremontii, E. torreyana, A. confertifolia and A. dumosa exhibited no or minimal turnover within the 18-year period. Among perennial forbs, only the gypsophile E. argophylla exhibited no or minimal turnover among years. The special-status species, both forbs, exhibited high turnover between 2020 and 2025. A. californica had 75% turnover and a net loss of one plot, while A. leiosolenus had 100% turnover by vacating all four plots the species had inhabited in 2020. The two cactus species were infrequent and persisted between 2020 and 2025 on the single plots they inhabited. When including all 28 perennial species, turnover at the community level on each plot compared to itself through time averaged 32% (SEM = 6%) between 2008 and 2020 and 44% (SEM = 6%) between 2020 and 2025.
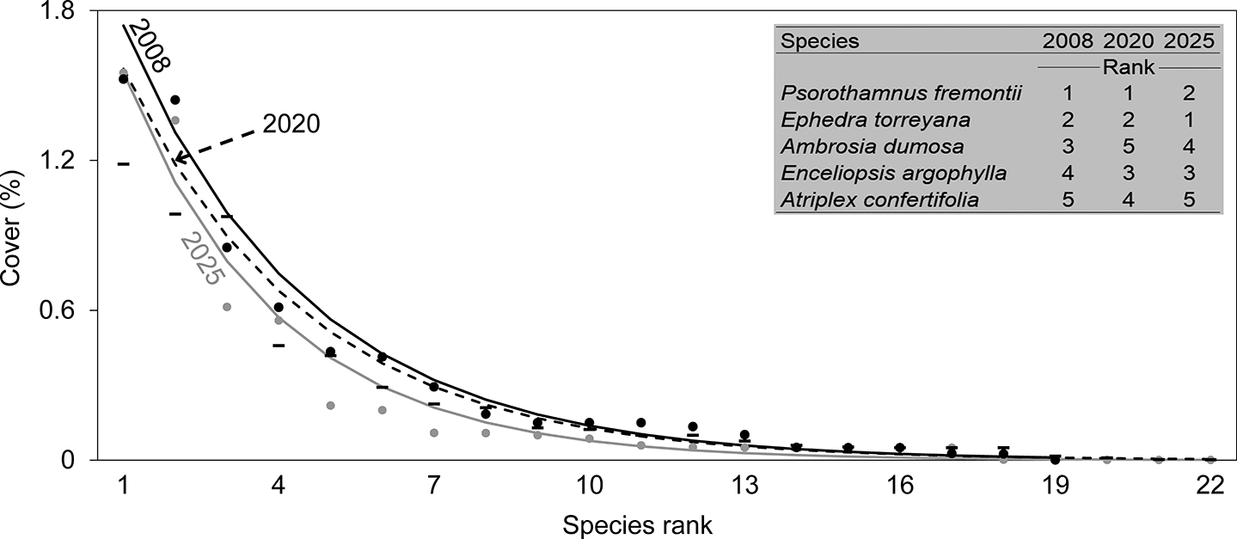
As community compositional turnover was occurring in parts of the perennial flora, the structure of the communities depicted by rank-abundance curves remained stable. Slopes of the log-transformed regressions did not differ among any of the three measurement years within the 18-year period (Figure 4). The top five species (P. fremontii, E. torreyana, A. dumosa, E. argophylla and A. confertifolia) with the most cover in 2008 retained their top five ranking 18 years later in 2025. Species with medium ranks of cover were also often stable in their ranking within the community. For example, Sphaeralcea ambigua varied little with the eighth, sixth and seventh most cover among perennials in 2008, 2020 and 2025. Although species with the least cover were typically transient, they were replaced by approximately equal numbers among years of species with similarly low cover, preserving the structure of the community rank-abundance curves.
Rank-abundance curves for perennial plant communities associated with gypsum soils for three measurement years within an 18-year period in the Mojave Desert, Nevada, USA. Each curve is the average of 10 plots, with points showing the species rank for each year in the same shading of symbols (where dashes represent 2020) as for the curve. The inset table shows the rankings of the five species with the greatest average cover each year (1 = the greatest cover). The k-value for the geometric series curve was 0.246 for 2008 (19 total perennial species present among all 10 plots), 0.244 for 2020 (24 species) and 0.283 (22 species). Note that there can be fewer ranks shown on the x-axis than total species present each year because species could be tied in rank.

Discussion
Climate context
The data indicate that both turnover and stability occurred simultaneously in different parts of the perennial plant community within the 18-year study period, which spanned high climatic variability and was generally dry. Based on the Palmer Drought Severity Index, our study began following the exceptionally wet 2004–2005 and subsequent dry conditions before and including our initial 2008 sampling (Supplementary Figure S2). After the wet 2004–2005, of the 20 years from 2006–2025, 65% were in drought (35% moderate–severe drought) during the November through April winter growing season and 80% were in drought (50% moderate–severe) during the warm season from May through October. Our spring 2020 sampling corresponded with a respite from dry conditions, as summer 2019 and the November 2019 through April 2020 growing season were wet. Following this, severe drought commenced from mid-2020 to mid-2022. Near-average conditions occurred in 2023–2024 before drought again characterized our 2025 sampling when no annuals emerged on our plots. While our study lacked a continuous time series of annual sampling, our study did encompass three different years spanning a range of current-year moisture conditions and key years within multiyear trends. For example, the sampling in 2008, while in a dry current year, may have still reflected a legacy of the exceptionally wet 2004–2005. The 2020 sampling portrays a brief wet period during a protracted dry period. The 2025 sampling represents a current dry year following a two-year, severe drought ending only two years earlier. Overall, our 18-year study period is consistent with the increasing prevalence of drought in North American deserts in recent decades (Soulé and Knapp, Reference Soulé and Knapp2024), highlighting the likely importance of understanding community dynamics during these drier conditions.
Turnover and stability in perennial forbs, including special-status species
Although the stable E. argophylla was an exception, turnover in perennial forbs as a group was over twice as high as for shrubs. While maximum life spans are not available for all species, prior research indicates that many perennial forb species in North American hot deserts have maximum life spans of years to a few decades, compared with the many decades to centuries for shrubs (Goldberg and Turner, Reference Goldberg and Turner1986; Bowers et al., Reference Bowers, Webb and Rondeau1995). For example, the herbaceous perennial S. ambigua, inhabiting 40% of our plots in 2008 and 20% in 2025, may have a typical maximum life span of only 8 years (Drake et al., Reference Drake, Esque, Nussear, DeFalco, Scoles-Sciulla, Modlin and Medica2015). One of the special-status species, A. californica, has a reported maximum life span of 6 years (de Queiroz and Meyer, Reference de Queiroz and Meyer2023). Moreover, a 16-year demographic study of its congener, Arctomecon humilis, also on gypsum soil, indicated that seedlings lived an average of 2.6 years and even those surviving to 1 year had an average total life span of only 4.6 years (Harper and Van Buren, Reference Harper and Van Buren2004). The shorter life spans of forbs, necessitating new recruitment of individuals in irregularly occurring favorable years, is likely associated with the high turnover in this group (Bowers et al., Reference Bowers, Turner and Burgess2004; Harper and Van Buren, Reference Harper and Van Buren2004).
The two special-status species, the perennial forbs A. leiosolenus and A. californica, both exhibited declines in all abundance metrics (cover, frequency and density) during our study. Although little autecological published data are available for A. leiosolenus, data for the more well-studied A. californica suggest that cycles of protracted population declines and replenishments characterize the species’ life history (de Queiroz and Meyer, Reference de Queiroz and Meyer2023). A. californica forms a persistent soil seed bank (Megill et al., Reference Megill, Walker, Vanier and Johnson2011; Abella et al., Reference Abella, Chiquoine and Vanier2013; de Queiroz and Meyer, Reference de Queiroz and Meyer2023). A seed burial experiment on gypsum soil in the Mojave Desert revealed that without new inputs, the A. californica seed bank could persist 20 years until complete depletion (de Queiroz and Meyer, Reference de Queiroz and Meyer2023). Moreover, the species may have multiple seed dormancy mechanisms, which remain poorly understood but are suspected to induce a variety of bet-hedging strategies for seed persistence across variable weather years (Abella et al., Reference Abella, Chiquoine and Vanier2013; Pereira et al., Reference Pereira, Chiquoine, Larranaga and Abella2021; de Queiroz and Meyer, Reference de Queiroz and Meyer2023). These seed burial and germination data are reinforced by observations that A. californica has reappeared at sites after 15 years of absence (Stosich et al., Reference Stosich, DeFalco and Scoles-Sciulla2022). Collectively, the literature for A. californica and its congener (A. humilis) indicates that one or more good recruitment years can reverse population declines (Harper and Van Buren, Reference Harper and Van Buren2004), suggesting that A. californica has well-developed persistence mechanisms, including a soil seed bank that could last our entire 18-year study. However, we agree with Bailey et al. (Reference Bailey, Pereira, Sion, Kobelt, Gentilcore, Antoninka and Bowker2023) that long-term demographic monitoring of A. californica populations is likely needed to differentiate fluctuation cycles from directional declines, a common challenge with rare plant conservation generally (Lindell et al., Reference Lindell, Ehrlén and Dahlgren2022).
From a conservation perspective, we highlight additional considerations for the two special-status species. The 20-year period since 2005 with only a single year (2020) with the winter–spring growing season ranked as moist by the Palmer Drought Severity Index is the second longest such period (the first encompassing mid-century drought from 1942–1979) in the 130-year drought record back to 1895 (Supplementary Figure S2). On our plots, the wet 2020 growing season was insufficient to forestall the ongoing decline in both species of aboveground plant abundance between 2020 and 2025. While viable A. californica seeds can persist 20 years in the soil (de Queiroz and Meyer, Reference de Queiroz and Meyer2023), the 18-year dry period of our study could nevertheless have reduced A. californica seed banks to curtail recruitment capacity in future favorable years. Additionally, floral resources could be affected by drought that are provided by other plant species surrounding the special-status species, with poorly understood influences on pollination and seed production (Chanprame et al., Reference Chanprame, Meidt, Griswold, Wilson and Graham2024). On the other hand, A. californica does not appear to depend on nurse plants for recruitment like many other desert perennials do, reducing the sensitivity of this species to nurse plant mortality (Abella et al., Reference Abella, Chiquoine and Vanier2013; Hartung et al., Reference Hartung, Rominger and Meyer2023).
By 2025, A. leiosolenus was absent from all of our study plots, and this species’ autecology and potential persistence mechanisms are not well known compared with A. californica. In a prior seed bank study in gypsum soil in the study area, we did not detect seeds of A. leiosolenus (even in samples below and within 1 m of mature plants) in the soil using either extraction or emergence assays (Abella et al., Reference Abella, Chiquoine and Vanier2013). Further research integrating soil seed bank and germination ecology with long-term demographic monitoring of both special-status species may further aid in understanding population dynamics to support conservation strategies for these species (e.g., distinguishing transient, cyclical declines from directional, persistent declines, or whether persistence in some locations of the landscape can facilitate recruitment elsewhere).
Stability in the shrub component
Aside from shrub species that inhabited only one or a few plots, shrubs were quite stable in multiple metrics, including similar cover, density and frequency among years and low or no turnover in occurrences among plots. The common, medium-to-large shrubs A. dumosa, A. confertifolia, E. torreyana and P. fremontii exemplified this stability of minimally varying cover and low turnover. Available from the literature, maximum life spans of individuals of the former three species exceed 100+ years (Bowers et al., Reference Bowers, Webb and Rondeau1995). With their long life spans, it may seem unsurprising that these shrubs were the most stable. However, considerable mortality and turnover have occurred in these and other shrubs in the Mojave and Sonoran deserts on non-gypsum soils, such as after prior droughts (e.g., Beatley, Reference Beatley1979; Miriti et al., Reference Miriti, Rodríguez-Buriticá, Wright and Howe2007; McAuliffe and Hamerlynck, Reference McAuliffe and Hamerlynck2010; Brown et al., Reference Brown, Rodriguez Buritica, Goldberg, Reichenbacher, Venable, Webb and Wilder2024). Excepting P. fremontii, which exhibited the least dieback, shrubs in our study had heavy dieback in 2025, which may correlate with the severe mid-2020–2022 drought and dry 2025, but they remained alive with little change in density. Whether the minimal dieback in P. fremontii relates to it being a legume with symbiotic nitrogen fixation aiding drought tolerance is unclear and warrants further research.
P. fremontii may be one of the most important, stable, foundational shrubs on these gypsum-affiliated soils. This stability could have multiple implications for ecosystem functions. For instance, Psorothamnus spp. are potentially important secondary nectar sources for a bee (Perdita meconis) considered for listing under the U.S. Endangered Species Act and that pollinates the special-status A. californica (Chanprame et al., Reference Chanprame, Meidt, Griswold, Wilson and Graham2024).
Overall stability in communities and edaphic specialists on special soils
Analyses of community composition, structure and diversity suggested a high degree of stability in these gypsum-associated soils within the 18-year period. Aside from a few metrics such as species diversity being high in the wet year of 2020 with prevalent annuals, most metrics did not vary among years. This was exemplified by no significant difference in community composition among years, with general stability in shrubs and turnover in many forbs but with directional change absent. Temporally stable rank-abundance curves revealed that when turnover occurred in more transient perennial forb species, they were replaced by other transient perennials. Thus, rather than directional temporal change, spatial turnover may predominate in the overall perennial forb component. Future research could map plants across broader areas to potentially partition spatial and temporal variation in these communities. In particular locations such as our plots, these communities may fit a core-transient species model of communities (Grime, Reference Grime1998; Abella et al., Reference Abella, Guida, Roberts, Norman and Holland2019). In this framework, a component of the flora is frequently occurring and stable (core species, such as the shrub P. fremontii and forb E. argophylla in our study) while another component is transient. In our study, this transient component was not just annuals but also much of the perennial forb component, excepting E. argophylla.
The classification of species’ associations with gypsum soil was not strongly related to stability or persistence. For example, total cover of gypsophiles (closely associated with gypsum-affiliated soil) did not differ among years. Moreover, the two gypsophile perennial forbs showed disparate patterns: E. argophylla was among the most stable of any species while A. leiosolenus was among the least stable. Further evaluating the importance of affiliation with gypsum compared with general traits (e.g., life span) may be useful in forecasts of change, as results internationally thus far are mixed for gypsum (Sánchez et al., Reference Sánchez, Alonso-Valiente, Albert and Escudero2017; Luzuriaga et al., Reference Luzuriaga, Ferrandis, Flores and Escudero2020) and for other special soils (e.g., Coppi et al., Reference Coppi, Lazzaro and Selvi2022; Ottaviani et al., Reference Ottaviani, Méndez-Castro, Conti, Zelený, Chytrý, Doležal, Jandová, Altman and Klimešová2022).
Conclusions and conservation implications
Metrics of community composition and structure exhibited high stability across three measurement years, varying widely in climatic conditions within an 18-year period that was generally dry. There was no consistent trend for total cover or turnover in species to vary with affiliation for gypsum-associated soil. As a group, perennial forbs, including two species of special conservation status (A. leiosolenus and A. californica), were the least temporally stable, together with the annual plant community contingent on favorable weather years. As the ability to persist in situ may be particularly important for unique communities and edaphic specialists on special soils, competition from invading, non-native plants is a potential threat to native species on special soils (Bishop et al., Reference Bishop, DeNittis and McGovern2023). All four of the non-native plants we detected during our study were annuals, and no directional trend in abundance of these species was detectable as they fluctuated with precipitation, similar to native annuals, and were absent in the dry last year of our study (2025). The shrub component of these gypsum-associated soils appears highly stable, exemplified by P. fremontii with minimal turnover and among the least post-drought dieback. Although the special-status species A. leiosolenus and A. californica may have persistence mechanisms such as a long-lived soil seed bank, their consistent decline aboveground across study years and the net sharp decline during the 18-year study suggest that further understanding the reliability of their persistence mechanisms and long-term spatiotemporal variation may help distinguish potential cyclical fluctuations from directional declines. Further determining potential relationships of the stability or turnover in other plant species surrounding the special-status species as a measure of habitat quality may also aid in conservation strategies of these species and their unique gypsum-associated communities.
Our results suggest that there may not be a binary classification of edaphic specialist species as either stable or dynamic in abundance on special soils during environmental change. Instead, our results suggest that there may be a range of responses within different subsets of plant communities, including specialist and more widespread species inhabiting special soils.
Open peer review
For open peer review materials, please visit http://doi.org/10.1017/dry.2026.10030.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/dry.2026.10030.
Data availability statement
The data that support the findings of this study are in Supplementary Table S2.
Acknowledgments
We thank Dianne Bangle, Sierra Jaeger, Adam Taylor, Kaden Ashdown, Carole-Lynna Benhamou, Ian Clark, Kevin Donnelly and Robert Pougnier for help with field data collection; the National Park Service (Lake Mead National Recreation Area) and Bureau of Land Management (Southern Nevada District) for permitting data collection; Alice Newton and Kent Turner (National Park Service) for supporting establishment of the study; and the Handling Editor and an anonymous reviewer for comments on the manuscript.
Author contribution
S.R.A. conceived the study and collected 2008 field data, and L.P.C. and S.R.A. coordinated the 2020 and 2025 data collection. S.R.A. performed statistical analyses and wrote the first draft of the manuscript. Both authors edited the manuscript.
Financial support
Data collection in 2008 was funded by the National Park Service (Lake Mead National Recreation Area) through a cooperative agreement with the University of Nevada, Las Vegas organized by Kent Turner and Alice Newton.
Competing interests
The authors declare none.






 Open access
Open access
Comments
20 November 2025
Dr. Osvaldo Sala and Dr. David Eldridge
Editors-in-Chief, Drylands
Dear Drs. Sala and Eldridge:
We are excited to submit a manuscript as a Research Article entitled “Stability and turnover in gypsum-associated plant communities during an 18-year period” for review to your journal.
Special soils with extreme properties form insular habitats that often support endemic species and unique communities that are priorities for conservation. Our study focused on the uncertainties for how these unique communities on special soils may change over multi-decade periods, including during droughts. In the Mojave Desert, USA, we examined change in plant communities on unique gypsum soils during an 18-year period spanning 2008-2025. Within the perennial plant community, we found evidence for both turnover and stability. With the exception of a species most strongly associated with gypsum, we found high turnover within perennial forbs, including two rare, conservation-priority species which declined sharply. In contrast, community structure, such as rank-abundance curves, remained little changed as transient species were replaced by other transients, preserving community structure. Moreover, while major mortality was reported in shrubs for non-gypsum soils, we found that the shrub component was highly stable during drought on these gypsum soils. We conclude the manuscript with habitat conservation applications from the findings.
This is an original manuscript, not under consideration for publication elsewhere or soon to be submitted elsewhere.
Sincerely,
Scott R. Abella, PhD
Associate Professor
Editor-in-Chief, Ecological Restoration
scott.abella@unlv.edu