


Cattle and fungi
Anaerobic zoosporic fungi inhabit the rumen of cattle where they engage in a symbiotic relationship with their host. First characterized by Orpin (Reference Orpin1975), these fungi have likely inhabited, and evolved in, the gastrointestinal (GI) tracts of large herbivores for millions of years (Theodorou et al., Reference Theodorou, Mennim, Davies, Zhu, Trinci and Brookman1996). In the rumen, fungi play a pivotal role in the degradation of recalcitrant fibrous feeds; as such, they are typically more abundant in animals fed low quality, high fibre diets (Hartinger and Zebeli, Reference Hartinger and Zebeli2021). Fungi are estimated to make up 8 – 20% of microbial biomass (Orpin, Reference Orpin and Cooke1981; Rezaeian et al., Reference Rezaeian, Beakes and Parker2004) and provide the host ruminant with volatile fatty acid (VFA), amino acids and vitamins (Hartinger and Zebeli, Reference Hartinger and Zebeli2021). Although fungi have long been intertwined with sustaining cattle life, only recently has the utility of exogenous fungi, their components and their fermentation products been realized for enhancing animal health and production in the dairy sector. Fungi can contain and produce a multitude of beneficial products including enzymes, amino acids, citric acid, polyphenols, mannan, adenosine and sterols (Chuang et al., Reference Chuang, Hsieh and Lee2020). These compounds can have powerful antioxidant, anti-inflammatory, hydrolytic and pro- and prebiotic effects in cattle, leading to enhanced immune function, rumen fermentation, growth, GI tract development, fertility and lactational performance (Beev et al., Reference Beev, Todorova and Tchobanova2007; Varada and Kumar, Reference Varada and Kumar2024).
Approximately 150,000 species of fungi have been classified to date (Bhunjun et al., Reference Bhunjun, Niskanen, Suwannarach, Wannathes, Chen, McKenzie, Maharachchikumbura, Buyck, Zhao and Fan2022), most of these species are probably not beneficial in the diets of cattle and some are pathogenic and/or produce mycotoxins, which can have negative ramifications for cattle health and productivity (Gallo et al., Reference Gallo, Giuberti, Frisvad, Bertuzzi and Nielsen2015). Species that have been used in, or to produce, feed additives for cattle include anaerobic fungi (AF): Piromyces spp., Orpinomyces spp., Anaeromyces spp., Neocallimastix spp. and obligately aerobic fungi: Trichoderma spp., Aspergillus niger, Aspergillus oryzae and yeast: Saccharomyces cerevisiae (Król et al., Reference Król, Słupczyńska, Wilk, Asghar and Cwynar2023; Varada and Kumar, Reference Varada and Kumar2024). The aim of this review is to consolidate literature investigating the utility of fungal feed additives in cattle, examine their benefits and limitations and highlight areas of research which require further exploration.
Classification, production and testing of fungal feed additives
Fungal feed additives can be broadly split into mixed fermentation products, direct-fed microbials (DFM) or probiotics and purified fermentation products or cell components. Mixed fermentation products, sometimes referred to as cultures, can contain some or all of: viable or non-viable cells, cultivation media and products of fermentation, including metabolites and enzymes (Krishna, Reference Krishna2005). The terms DFM and probiotic are often used interchangeably to describe additives with a live component; however, they have slightly different definitions. Direct-fed microbials are defined as additives that only contain live or naturally occurring microbes (FDA, 1995), whereas probiotics are defined as live microorganisms, that when administered in adequate amounts, confer a health benefit upon the host (FAO/WHO, 2001), so may contain metabolites, enzymes and live organisms. Purified fermentation products are specific compounds or molecules that have been produced by fungal fermentation; and then, extracted from the fermentation media. In the context of this review, the most relevant fermentation products are enzymes, the profile of which is dependent on fungal species, fungal strain, genetic modification, cultivation method and cultivation substrate (Krishna, Reference Krishna2005). The use of fungal cell components in cattle nutrition is predominantly focused on yeast. The intracellular soluble component of yeast cells is a valuable source of nucleotides, vitamins and lipids, and the insoluble cell walls are rich in mannan-oligosaccharides (MOS) and β-Glucans (Shurson, Reference Shurson2018).
The production of all fungal feed additives requires fermentation. Commercial techniques are solid-state fermentation (SSF) or submerged fermentation (SmF); both involve cultivating fungi on a substrate in the presence (SmF) or absence (SSF) of free-flowing liquid. Solid-state fermentation tends to have higher yields, cost less and have a lower risk of contamination than SmF (Jonathan et al., Reference Jonathan, Tania, Tanjaya and Katherine2021). However, when the entire culture media is harvested from SSF to produce mixed fermentation products, there can be lack of homogeneity throughout the product, caused by difficulties agitating a solid fermentation bed (Krishna, Reference Krishna2005). This can be problematic when the recommended feeding doses of mixed fermentation products are low, potentially leading to heterogenous distribution of the active components and lack of efficacy in the final feedstuff. Conversely, SmF typically occurs in stirred-tank reactors, which makes achieving physical and chemical homogeneity considerably easier (Jonathan et al., Reference Jonathan, Tania, Tanjaya and Katherine2021). The purpose of fungal fermentation can be solely to produce feed additives for livestock, or additives can be obtained from by-products of other fermentative processes, such as brewing (Shurson, Reference Shurson2018). To obtain specific fungal enzymes, metabolites or cell components, purification steps are required. For enzymes, this involves filtration, solvent extraction, precipitation and chromatography (Jonathan et al., Reference Jonathan, Tania, Tanjaya and Katherine2021). Cell components can be obtained using ultrafiltration, subcellular fractionation, enzymatic digestion, Concanavalin-A chromatography and sodium dodecyl sulphate extraction (Utama et al., Reference Utama, Oktaviani, Balia and Rialita2023). Most fungal products are dried and stored anaerobically for stability. The technique for drying products with an active component is particularly important as moist heat can disrupt fungal cells; commonly used methods for drying DFM and probiotics include rotolouver, fluid-bed and tunnel drying (Shurson, Reference Shurson2018).
In vitro and in vivo techniques are used to test the effects of fungal feed additives. When the primary mode of action (MOA) of a product is enzymatic, its activity will usually be assessed by quantifying the rate of accumulation of the product of the reaction that the enzyme catalyses when combined with a standard substrate. However, the conditions under which these assays are performed are often not representative of the conditions found in the digestive tract of cattle, where pH, temperature and inhibitory co-factors can all affect enzyme efficacy (Meale et al., Reference Meale, Beauchemin, Hristov, Chaves and McAllister2014). The in vitro gas production (GP) technique overcomes some of these challenges, where rumen fermentation kinetics are modelled by measuring GP, when substrates are incubated with microbial inoculum derived from rumen digesta or fluid (Theodorou et al., Reference Theodorou, Williams, Dhanoa, McAllan and France1994). The GP technique is a high throughput method that can be used to test the effects of feed additives on digestibility and gas composition, providing a better insight into the effects that enzymatic products could have in vivo, compared to gas chromatography, high-performance liquid chromatography or dinitrosalicylic acid assays. In vitro GP does not mimic rumen motility or mastication, and the microbial diversity present in inoculum may not be typical of the populations in the rumen, especially if rumen fluid that excludes protozoa and most fungi is used, limiting the conclusions that should be drawn from GP experimentation (Yáñez-Ruiz et al., Reference Yáñez-Ruiz, Bannink, Dijkstra, Kebreab, Morgavi, O'Kiely, Reynolds, Schwarm, Shingfield and Yu2016). When the MOA of fungal products is pre- or probiotic, digestive tract simulations are equally valuable. These assays focus on screening product viability and actions under conditions found in the stomach and small intestine; for cattle, these assays are less developed than rumen simulations and present a valuable area of further refinement. Intestinal conditions to measure pH and bile stability can be mimicked using incubations of potassium dihydrogen phosphate, bovine bile, calcium chloride and pancreatin (Vinyard et al., Reference Vinyard, Sarmikasoglou, Bennett, Arce-Cordero, Aines, Estes and Faciola2021). Haemolytic and immunomodulation activities can be measured using cell and co-cultures to examine the effects of additives on immune cells, epithelial structure and integrity and bacteria (Papadimitriou et al., Reference Papadimitriou, Zoumpopoulou, Foligné, Alexandraki, Kazou, Pot and Tsakalidou2015). When fungal additives contain viable cells, activity is usually expressed as colony- (CFU) or thallus-forming units (TFU), per unit weight of product (Król et al., Reference Król, Słupczyńska, Wilk, Asghar and Cwynar2023). CFU is a measure of the number of cells able to reproduce on a media to form colonies rather than a measure of cells viability per se. Interpretation can be confounded by the fact that colonies can be formed from one cell or from multiple cells that have aggregated, and the incidence of aggregation is increased following heat treatment, for example, pelleting feed (Kwolek-Mirek and Zadrag-Tecza, Reference Kwolek-Mirek and Zadrag-Tecza2014).
In vivo experimentation is the gold standard for testing fungal feed additives in cattle. Using animals in research is expensive and time consuming and requires ethical consideration. Hence, pre-screening products using in vitro techniques to determine potential utility and optimal inclusion rates is important economically and ethically before proceeding to animal studies. In vivo experiments can measure the effects of treatments on primary response variables such as growth, health and lactation, with these variables likely to influence the industry uptake of additives. However, characterizing the biological mechanisms that lead to beneficial health and production responses is critical for product development and refinement. When improved rumen fermentation is the desired outcome from supplementation, ruminally cannulated cattle can be used to investigate the effects of additives on feed and feed component degradation using in sacco techniques (Ørskov et al., Reference Ørskov, Hovell and Mould1980). Rumen access provides the cleanest and most accurate means to determine effects on rumen microbial community composition and diversity, which is useful for products with digestibility and health MOAs. Measuring the effects of products with purported health benefits can be done non-invasively using indicators such as faecal score, behaviour, and biological markers in milk (Džermeikaitė et al., Reference Džermeikaitė, Bačėninaitė and Antanaitis2023). However, haematological, histological and blood biochemical variables can provide considerably more information on the effects of treatment on immunity and digestive tract health and development. Studies that can measure the effects of treatments at cellular, tissue and whole-body system levels undoubtedly provide the most comprehensive insight into the efficacy of fungal feed additives.
Anaerobic fungi
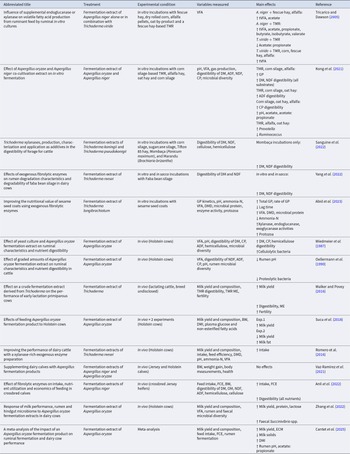
AF play a pivotal role in degrading lignocellulosic compounds in the rumen and are efficient fibre degrading microbes (Hess et al., Reference Hess, Paul, Puniya, Van der Giezen, Shaw, Edwards and Fliegerová2020). This physiological group of fungi has flagellated zoospores which reach and colonise deep inside fibrous material, making regions which are usually unavailable to fibrolytic bacteria, accessible through the physical splitting of plant tissue (Bhagat et al., Reference Bhagat, Kumar, Kumari and Bharti2023; Du et al., Reference Du, Zhou, Li, Zhang, Wang, Yao, Chen, Liu and Cao2024). This penetrative capability is compounded by the remarkable suite of enzymes that AF encode for, including polysaccharidases, glycosidases, esterases and pectinolytic lyases, which are employed to rapidly hydrolyse fibre components entering the rumen (Bhagat et al., Reference Bhagat, Kumar, Kumari and Bharti2023). Given that cell wall digestibility in the rumen is limited to 60 % under ideal conditions (Van Soest, Reference Van Soest1994), supplementing cattle with AF that have efficient cell wall degrading capabilities could be a useful strategy to enhance feed efficiency. However, it is AF's vulnerability to oxygen that limits the accessibility of supplemental AF as feed additives on farm. A summary of the research examining anaerobic fungal additives, discussed in this review, is shown in Table 1.
Summary of treatments, variables, and main effects, in studies examining the use of anaerobic fungal additives in dairy cattle

Abbreviations: NDF = neutral detergent fibre; DM = dry matter; DMD = dry matter disappearance; GP = gas productions; VFA = volatile fatty acids; N = nitrogen; ADF = acid detergent fibre; TMR = total mixed ration; tVFA = total volatile fatty acids; DMI = dry matter intake; OM = organic matter; CP = crude protein; CF = crude fibre; BW = body weight; TCA = trichloroacetic acid; ADG = average daily gain; TDN = total digestible nutrients.
Paul et al. (Reference Paul, Deb, Punia, Singh and Kumar2010) isolated and cultured 10 Piromyces spp. from Nilgai (Boselaphus tragocamelus) and wild cattle (no breed specified) and quantified their effect on in vitro rumen fermentation of wheat straw, when inoculated with rumen fluid from buffalo. All five isolates obtained from wild cattle increased the digestibility of wheat straw and increased fungal biomass in incubations. However, none of the Piromyces isolates obtained from Nilgai increased the true digestibility of wheat straw, and only one isolate increased fungal biomass. This highlights substrate specificity differences between fungi of the same genus, isolated from cattle and Nilgai, with those cultured from cattle more useful at hydrolysing the fibre present in wheat straw. Dietary differences between Nilgai and cattle likely explain this, with Nilgai favouring forbs and shrubs and cattle preferentially consuming forages (Sheffield, Reference Sheffield1983). The species of Piromyces present in the rumens of cattle should be effective at hydrolysing the fibres their host preferentially consumes, including the wheat straw fibres investigated by Paul et al. (Reference Paul, Deb, Punia, Singh and Kumar2010). Alternatively, limited effects from Nilgai-isolated Piromyces could indicate that they were not equipped to survive in the microbial community present in the buffalo rumen fluid.
Sirohi et al. (Reference Sirohi, Choudhury, Dagar, Puniya and Singh2013) isolated Orpinomyces spp. and Anaeromyces spp. from rumen fluid collected from Karan × Friesian cattle and quantified the effect of incubating their cultures with NDF from wheat straw on in vitro fermentation. Acetate production was increased by the addition of all fungal cultures as was the digestibility of NDF and total microbial biomass production. Incubations containing Orpinomyces spp. yielded increased NDF digestibility compared with incubations containing Anaeromyces spp., suggesting the former is more useful for enhancing fibre degradation than the latter. In accordance with this, when Du et al. (Reference Du, Zhou, Li, Zhang, Wang, Yao, Chen, Liu and Cao2024) introduced Oprinomyces sp. YF3, cultured from the rumens of Qinghai yaks, into rumen fluid from dairy goats, an increase in acetate, acetate: propionate ratio and total volatile fatty acid (tVFA) production from the in vitro fermentation of wheat straw was measured. This was likely associated with the enhancement of exoglucanase and acetyl esterase activities and an increase in Ruminococcus and Pyramidobacter abundance from Oprinomyces addition, highlighting a synergistic relationship between AF and fibre degrading bacteria. Although it is commercially impractical to harvest AF from one ruminant and transfer to another, this study does indicate the highly efficient fibre degrading capabilities of Oprinomyces sp. YF3 and its ability to propagate favourable microbial dynamics in the rumen. Kumar et al. (Reference Kumar, Seghal, Mir, Jha and Ganie2012) inoculated rumen fluid with broth containing lab-cultured Orpinomyces joyonii zoospores to quantity the effect of AF addition on the in vitro fermentation kinetics of wheat straw, paddy straw, and a TMR consisting of both straws and 40 % concentrate. AF addition increased the digestibility of DM, ADF and NDF in all substrates, and AF incubations had greater esterase activities than controls. Esterases are key enzymes involved in the hydrolysis of ester linkages between lignin and hemicellulose; thus, the increase in their activities from AF supplementation likely caused the increased digestibility of substrates. The capacity of AF to enhance the digestibility of fibre has been presented by all in vitro studies discussed. However, maintaining anaerobic conditions to maintain AF viability in a lab is less challenging than on farm. In vitro experimentation is undoubtedly valuable, but for supplemental AF to have industry utility, applications must be translatable to on-farm practicality and more in vivo studies are required to validate effects. Although AF cannot grow in the presence of oxygen, some species can tolerate aerobic environments for several hours (Trinci et al., Reference Trinci, Lowe, Milne and Theodorou1988). Thus, in conditions where feed or animals can be regularly inoculated with fresh isolates that will be ingested into a favourable environment for fungal survival, their effects on performance and digestibility could be harnessed. However, most farmers would lack the time and infrastructure to culture AF and regularly inoculate cattle or feeds.
When Saxena et al. (Reference Saxena, Sehgal, Puniya and Singh2010) dosed buffalo with 250 ml/day broth containing Piromyces sp. WNG-12 or Orpinomyces sp. C-14 isolates, beneficial lactation responses were measured. Increases in milk yield and milk solids were realized from treatment; although, the authors declared significance at P < 0.1 and did not state specific P-values; so, effects might be tendencies and not meet the more robust significance level of P < 0.05. Rumen fermentation variables from treatments were significantly (P < 0.05) enhanced, with increased rumen VFA, total N, trichloroacetic acid (TCA) precipitable N, decreased ammonia N and zoospore count in rumen fluid. In lactating buffalo, Sehgal et al. (Reference Sehgal, Mir, Jha, Kumar and Sharma2012) measured enhanced rumen fermentation performance from inoculating Orpinomyces joyonii zoospores at 10.7 × 106 CFU/ml/kg feed into a wheat straw, paddy straw, concentrate and green fodder TMR, daily. Over an experimental period of 105 d, using five buffalo per treatment, the digestibility of DM, OM, NDF, ADF and CP was increased from treatment, as was that of tVFA, total N, ammonia N and zoospore count in rumen fluid. Increased energy availability from enhanced nutrient digestibilities would suggest that O. joyonii supplementation could improve production responses. However, feed intake and lactational performance of buffalo were unaffected, indicating that digestibility of feeds was not increased sufficiently to cause a detectable production response or perhaps that the study was statistically underpowered. The literature examining AF supplementation in lactating cattle is sparse, but further exploration of AF effects on lactational biology is worthwhile and could improve the performance of cattle fed high levels of recalcitrant fibre. Notably, cattle on low quality fibre diets are often situated in lower income systems, where the use of any fungal additive could be limited by socio-economic factors (Balehegn et al., Reference Balehegn, Duncan, Tolera, Ayantunde, Issa, Karimou, Zampaligré, André, Gnanda and Varijakshapanicker2020).
Several studies have examined the effect of AF in young cattle. Because fungi are some of the first microbes to inhabit the rumen (Li et al., Reference Li, Shi and Na2023), exogenous AF could propagate rumen microbial colonization and enhance digestive function. However, none of the studies identified by this review examined AF supplementation in pre-ruminant calves, highlighting an area for future research to examine if supplemental AF could enhance rumen development through improved solid feed degradation. Kumar et al. (Reference Kumar, Sehgal, Puniya and Kumari2015) found beneficial effects on growth and digestibility in young buffalo aged 5–7 months, when incorporating Neocallimastix sp. GR-1 zoospores into wheat straw-based diets. Lab cultured zoospores were inoculated into straw blocks at 106 zoospores/kg feed over a 172-day experimental period. Animals fed fungal treated straw had improved growth rates (16%) and feed efficiencies (28.7%) compared to controls. Enhanced animal performance was likely caused by the increase in OM, DM, NDF, ADF, CP, crude fibre (CF) and cellulose digestibilities of wheat straw from zoospore treatment. Dey et al. (Reference Dey, Sehgal, Puniya and Singh2004) measured the effect of supplemental Orpinomyces C-14 in twelve 10-month-old Tharparkar × Holstein Friesian cattle on a 50:50 wheat straw: concentrate diet. Animals were orally drenched once per week with 160 ml of fungal culture containing 106 CFU/ml. Similar to the observations of Kumar et al. (Reference Kumar, Sehgal, Puniya and Kumari2015), Dey et al. (Reference Dey, Sehgal, Puniya and Singh2004) reported that calves drenched with AF had greater weight gain (15.7%) than control calves and increased digestibilities of DM, CP, N free extracts, CF, NDF and ADF. Rumen pH and ammonia N were lower in fungal treated cattle, and total N and TCA precipitable N were increased. Physical disruption of plant tissues by Orpinomyces hyphae likely caused an increase in cellulose surface area available for degradation, leading to the increase in ADG observed in this study. Notably, weekly drenching was sufficient to stimulate this effect, suggesting that supplementation altered the dynamics of the rumen microbiome for longer than the life cycle of the Orpinomyces C-14 administered, which would typically be 24 – 36 h (Lowe et al., Reference Lowe, Griffith, Milne, Theodorou and Trinci1987). The rumen microbiome is normally extremely resistant to the introduction of exogenous microbes (Weimer, Reference Weimer2015), but perhaps because AF are already present in the rumen, it may be easier to alter their proportions compared with the introduction of microbes which do not normally reside there. Investigations into how long AF population levels remain functionally elevated post supplementation would inform the intervals that could be employed in dosing strategies; the utility of exogenous AF in cattle would be considerably greater if dosing did not need to occur daily to realize beneficial effects. Paul et al. (Reference Paul, Deb, Punia, Das, Singh, Ashar and Kumar2011) found that dosing buffalo calves (weight 148 ± 12 kg, age undeclared) every 8 d with Neocallimastix sp. CF-17 zoospores and rhizomycelia was optimal to enhance digestibility and growth rates. The first experiment in this study examined two dosing techniques over a 4-week experimental period, oral dosing of 200 ml liquid AF culture containing 106 TFU/ml and the same dose of AF encapsulated in an oxygen resistant biopolymer (composition undeclared). Only calves administered with encapsulated Neocallimastix had greater ADG than control calves. Both liquid and encapsulated treatments led to improved OM digestibility, but only encapsulated AF caused an improvement in NDF digestibility and only liquid AF treatment calves had increased DM and ADF digestibilities. Notably, both groups of treatment calves consumed increased TDN. When different dosing frequencies of encapsulated AF were investigated in a further experiment, there was no difference between growth rates in calves dosed every 4 days and calves dosed every 8 d, with both groups having increased growth (20 %) compared to controls. This indicates that a dosing frequency of at least 8 d can improve calf performance and encapsulation enhanced calf growth compared with oral dosing of liquid culture. The biopolymer used by Paul et al. (Reference Paul, Deb, Punia, Das, Singh, Ashar and Kumar2011) only afforded fungi protection from oxygen for 12 h, which would still make on-farm storage challenging. Research to develop a more resistant polymer to protect AF from oxygen would increase the accessibility of AF to farmers.
Limited research examining the effects of supplemental AF in cattle has realized consistent responses in improved nutrient digestibility, which are often paired with improvements in animal performance. The fibre-degrading capabilities of AF make them an appealing tool for ruminant producers and nutritionists, only hindered by challenges in handling and storage which arise from the anaerobic conditions required for AF viability. An area of research which could help overcome these challenges is the resting stage of AF's life cycle. This stage is poorly understood but is believed to allow fungi to survive for prolonged periods in aerobic conditions and has been proposed as the reason why AF can be cultured from faeces that has been exposed to oxygen for long periods of time (Hess et al., Reference Hess, Paul, Puniya, Van der Giezen, Shaw, Edwards and Fliegerová2020). Comprehension and characterization of the resting stage of AF could significantly expedite the development of AF supplements for cattle and, as such, requires investigation.
Obligately aerobic fungi
The rumen is considered an anaerobic environment. However, small amounts of oxygen enter the rumen during feeding and regurgitation, and this is likely why obligate aerobes including Aspergillus and Trichoderma spp. can be isolated from rumen fluid (Almeida et al., Reference Almeida, Duarte, Abrão, Freitas, Geraseev and Rosa2012). Although the relative abundance of obligately aerobic fungi is small compared to AF and other microbes in the rumen, a diversity of species can persist (Wang et al., Reference Wang, Huang, Chen, Song, Peng, He, Huang, Huang, Wang and Lan2024). This is demonstrated by Almeida et al. (Reference Almeida, Duarte, Abrão, Freitas, Geraseev and Rosa2012), who isolated 49 species from the genera: Aspergillus, Gliocladium, Paecilomyces, Rhizophus and Scedosporium, from the rumen fluid of dairy cattle fed tropical forages. Outside of the rumen, obligate aerobes are predominantly used for the fermentation products that they can produce. These filamentous fungi are often characterized as ‘enzyme factories’ and have been used in human food fermentation for many years (Abdullahi et al., Reference Abdullahi, Atiku and Umar2021). In cattle nutrition, the main focus of obligately aerobic fungi is in biotransformation to produce enzyme rich feed materials, where fungi can be cultivated on low-value substrates to produce high-value active compounds that can act as digestibility enhancers (Thomas et al., Reference Thomas, Larroche and Pandey2013). A summary of the research examining obligately aerobic fungal additives, discussed in this review, is shown in Table 2.
Summary of treatments, variables and main effects, in studies examining the use of obligately aerobic fungal additives in dairy cattle

Abbreviations: DM = dry matter; NDF = neutral detergent fibre; DMD = dry matter disappearance; GP = gas productions; VFA = volatile fatty acids; N = nitrogen; ME = metabolizable energy; TMR = total mixed ration; tVFA = total volatile fatty acids; ADF = acid detergent fibre; DMI = dry matter intake; OM = organic matter; CP = crude protein; CF = crude fibre; BW = body weight; FCE = feed conversion efficiency; ECM = energy-corrected milk.
Trichoderma spp. are soil-borne fungi with global distribution; they synthesize a range of fibrolytic enzymes and antimicrobial compounds (Król et al., Reference Król, Słupczyńska, Wilk, Asghar and Cwynar2023). Sanguine et al. (Reference Sanguine, Cavalheiro, Garcia, Dos Santos, Gandra, E Buschinelli, da Paz, Fonseca and Leite2022) quantified the effects of xylanases produced by the SSF of Trichoderma koningii and Trichoderma pseudokonigii on the in vitro rumen digestibility of a range of forages. The authors only declare results from incubations using Mombaça (Panicum maximum), where xylanases from both Trichoderma spp. increased DM and NDF digestibility, suggesting that there was possibly no effect on the other feeds examined. The xylanases were found to be stable at a pH range of 3–9.5 and were thermostable for 1 h at 45–50 °C. The pH stability of exogenous enzymes (EE) can limit their efficacy in the rumen (Beauchemin et al., Reference Beauchemin, Colombatto and Morgavi2004). However, the range of stability in these Trichoderma EE is far greater than pH fluctuations that would occur in a healthy rumen, suggesting that stability would not limit their use in vivo, but limited effects on the digestibility of feeds other than Mombaça, might. When incubating 0.2 g DM sesame seed coats with Trichoderma longibrachiatum extract containing xylanase, endoglucanase and exoglucanase, Abid et al. (Reference Abid, Jabri, Yaich, Malek, Rekhis and Kamoun2023) measured improvements in in vitro rumen fermentation variables. A dose of 4 µl/g DM substrate decreased the lag time for the start of fermentation and increased the rate of GP, DM disappearance (DMD), VFA production, hydrolysis of NDF and rumen fibrolytic enzyme activity, whereas lower doses of 1 and 2 µl/g DM had no effect on fermentation variables. This highlights that although Trichoderma enzymes can modulate the fermentation of low-quality feedstuffs in vitro, this is dose-dependent. As with pH stability, inappropriate dose rates of EE are frequently cited as potential explanations for when EE are ineffective in ruminants (Beauchemin et al., Reference Beauchemin, Colombatto and Morgavi2004; Meale et al., Reference Meale, Beauchemin, Hristov, Chaves and McAllister2014).
Yang et al. (Reference Yang, Guevara-Oquendo, Refat and Yu2022) measured the effects of seven doses of Trichoderma reesei-derived xylanase and cellulase on in vitro and in sacco rumen fermentation of Faba bean silage (0, 0.25, 0.5, 0.75, 1, 1.25, and 1.5 ml/kg DM silage). 0.5 and 7. 5 g DM Faba bean silage were used in in vitro and in sacco incubations, respectively. Exogenous enzymes improved DMD and NDF degradation kinetics both in vitro, using DaisyII incubators, and in sacco, following the protocol of Ørskov et al. (Reference Ørskov, Hovell and Mould1980). The soluble and potentially degradable NDF fraction of Faba bean silage was increased with increasing dose of EE; whilst, the undegradable fraction was linearly decreased. However, DMD was cubically improved, with the intermediate EE dose of 1 ml/kg DM silage having the highest digestibility of 56.54%. This shows that higher doses of EE do not always exert greater effects on digestibility, further emphasizing the importance of characterizing optimal doses of EE. Notably, authors measured a robust correlation between in vitro and in sacco DMD (r = 0.98, P < 0.01) and NDF degradability (r = 0.84, P < 0.01); this demonstrates the utility of in vitro rumen simulations for screening the effects of enzymatic products on digestibility variables.
Romero et al. (Reference Romero, Macias, Ma, Martins, Staples, Beauchemin and Adesogan2016) did not realize consistently beneficial lactation responses when supplementing two fermentation extracts of T. reesei containing xylanase and cellulase + xylanase to 66 lactating Holstein cattle. Extracts were sprayed onto a dairy TMR and the xylanase treatment increased DM, OM and CP intakes. During the 10-week experiment, the treatments had limited effects on lactation. There was no improvement in milk solids or feed efficiency from either treatment; although, during weeks 3, 6 and 9, cattle treated with xylanase had higher milk yield than controls. This transient response limits the evidence that the Trichoderma extract investigated could consistently improve milk yields on farm; although, the authors suggest that the tendency (P > 0.05) of treatments to improve milk yield and solids is sufficient to infer enhanced lactation. A tendency of Trichoderma fermentation extract to improve lactational performance was also measured by Walker and Povey (Reference Walker and Povey2016). Fifty primiparous cows were recruited on a 12-week study to quantify the effect of pre-treating a TMR made up of grass silage, corn silage and concentrate with Trichoderma extract at 750 ml/T DM. During the first 9 weeks of the study, milk yield tended to be higher in cows consuming the treatment TMR; however, in the last 3 weeks of the study, milk production was significantly increased. This perhaps suggests that the effects of Trichoderma extract supplementation take several weeks to translate into production responses. Notably, the pregnancy rate of cows consuming Trichoderma extract was also 20% higher than control cattle, indicating potential benefits to fertility from supplementation. This effect requires further investigation and may be associated with the production of antimicrobials by Trichoderma (Zin and Badaluddin, Reference Zin and Badaluddin2020), potentially reducing the risk of conception limiting conditions, such as metritis and endometritis.
Fermentation products of A. niger and A. oryzae have also realized benefits for rumen fermentation kinetics and cattle performance. Tricarico and Dawson (Reference Tricarico and Dawson2005) measured the effect of an A. niger extract with xylanase activities of 135 xylanase units (XU)/ml on VFA production from fescue hay, dry rolled corn, alfalfa pellets, oat by-product and a fescue hay-based TMR, in a batch culture rumen simulation using rumen fluid from a cannulated heifer. The A. niger extract increased tVFA production from the TMR, fescue hay, and alfalfa pellets, as well as increased molar concentrations of acetate, propionate, butyrate, isobutyrate, and valerate from TMR and acetate from fescue hay and alfalfa incubations. Notably, when the A. niger extract was combined with a Trichoderma viride extract containing endoglucanase activity of 5 carboxymethyl cellulase (CMCU)/ml, the acetate to propionate ratio in the TMR incubation was decreased and tVFA production from the TMR, dry rolled corn, fescue hay, and alfalfa pellets was higher than when either extract was incubated on its own. This suggests an additive effect between fermentation products of Aspergillus and Trichoderma, where the combination of metabolites and enzymes in both products has increased tVFA production from substrate fermentation. Kong et al. (Reference Kong, Lu, Liu, Zhang, Jiang, Wang, Wang and Li2021) treated a corn silage-based TMR, alfalfa hay, oat hay and corn silage with a fermentation extract from the co-cultivation of A. oryzae and A. niger (AOAN) to measure the effects of treatment on in vitro rumen fermentation. The AOAN decreased pH in all incubations and increased tVFA production from all substrates apart from alfalfa hay. Acetate and the acetate to propionate ratio were increased in oat hay and alfalfa hay incubations. As in the study of Tricarico and Dawson (Reference Tricarico and Dawson2005), the effects of this AOAN on VFA production were substrate specific. Kong et al. (Reference Kong, Lu, Liu, Zhang, Jiang, Wang, Wang and Li2021) observed improvements in DM, ADF and NDF digestibilities in all feed from AOAN treatment, except for ADF in alfalfa hay. This highlights the ability of AOAN to enhance fibre digestibility across substrates. Microbiome analysis revealed that AOAN also influenced α and β diversity; the prevalence of Prevotella was increased and Ruminococcus decreased in TMR, alfalfa and oat hay incubations; however, microbial diversity was unaffected in corn silage incubations. This indicates that fungal extracts could alter rumen fermentation through influencing microbial composition in the rumen or solely enhancing the digestibility of feeds with bioactive compounds present in the extract. Further investigation of these mechanisms would provide insight into how to maximise the digestibility-enhancing capacity of AOAN.
Zhang et al. (Reference Zhang, Jin, Jiang, Xie and Mao2022) supplemented 10 multiparous Holstein cows with 5 g/day A. oryzae extract (Nutri-Ferm; Nutri-AD International, Belgium) for 5 weeks. Cows fed A. oryzae extract had increased milk yield (1.5 kg/day) compared to controls and higher yields and concentrations of milk protein and lactose; whilst, milk fat production was unaffected. Rumen VFA production and proportions were unaffected as was rumen microbial diversity. Interestingly, the composition of the faecal microbiome was altered by treatment, with A. oryzae treated cows having an increased abundance of unclassified Succinivibrio spp. This genus is involved in the production of succinate and may be associated with increased caecal propionate production (Iqbal et al., Reference Iqbal, Zhang, Yang, Zou, Li, Liang, Wei and Lin2018). Zhang et al. (Reference Zhang, Jin, Jiang, Xie and Mao2022) measured an increase in faecal propionate percentage and decreased acetate: propionate ratio; as well as increased faecal pH in treatment cows, which should promote caecal epithelial health and enhance hindgut VFA absorption. The hindgut is an important site for VFA production and absorption, yet is often overlooked in cattle nutrition (Sanz-Fernandez et al., Reference Sanz-Fernandez, Daniel, Seymour, Kvidera, Bester, Doelman and Martín-Tereso2020). These results suggest that further investigation into the effects of A. oryzae in the cattle hindgut could be warranted.
When Oellermann et al. (Reference Oellermann, Arambel, Kent and Walters1990) supplemented five non-lactating Holstein cows with five doses of A. oryzae extract (0, 1, 2, 4, 6 g/day) in a randomized block design study, supplementation had limited effects on rumen fermentation. A dose of 1 g/day decreased rumen pH, however, none of the doses influenced VFA production or the digestibilities of NDF, ADF and CP. Doses of 2, 4 and 6 g/day decreased the number of proteolytic bacteria in the rumen but did not increase the proliferation of AF, cellulolytic bacteria, amylolytic bacteria, or protozoa. Arambel et al. (Reference Arambel, Wiedmeier and Walters1987) suggests that A. oryzae can increase rumen proteolysis. So, perhaps, the population of proteolytic bacteria was suppressed by the endogenous proteolytic activity of A. oryzae in the experiment of Oellermann et al. (Reference Oellermann, Arambel, Kent and Walters1990). However, this was not paired with increased CP digestibility so is not functionally advantageous. Conversely, when Wiedmeier et al. (Reference Wiedmeier, Arambel and Walters1987) supplemented four non-lactating Holstein cows with 2.63 g/day A. oryzae extract (Amaferm®; BioZyme Inc, St Joseph, MO, USA), in a 4 × 4 Latin square design experiment, the digestibility of CP was increased, as was DM and hemicellulose digestibilities. Supplementation with A. oryzae extract increased the absolute and proportional amount of cellulolytic bacteria in rumen fluid, suggesting that A. oryzae increased structural carbohydrate hydrolysis and the proliferation of other fibrolytic microbes. This highlights the different effects that similar fungal products can have on the rumen microbiome; with Oellermann et al. (Reference Oellermann, Arambel, Kent and Walters1990) observing an increase in proteolysis paired with a decrease in proteolytic microbes, and Wiedmeier et al. (Reference Wiedmeier, Arambel and Walters1987) observing an increase in fibre degradation paired with an increase in fibrolytic microbes. Sucu et al. (Reference Sucu, Moore, VanBaale, Jensen, Sanz-Fernandez and Baumgard2018) found that feeding primiparous and multiparous Holstein cows (n = 33) 15 g/day of the same A. oryzae extract as Wiedmeier et al. (Reference Wiedmeier, Arambel and Walters1987) in a TMR consisting of alfalfa hay and steam-flaked corn, milk yield was increased by 3.1 kg/day compared to controls. A. oryzae treated cows achieved peak yield two weeks earlier than control cows, with average milk yields 4.5 kg/day higher in weeks three and four postpartum and no difference in volumetric concentrations of milk solids between groups. In this study, treatment administration began 21-day pre-calving, suggesting potential benefits from A. oryzae extract supplementation in the pre-fresh period that could lead to increased milk yields in the transition period. However, in a second experiment using 445 multiparous Holstein cows, A. oryzae extract supplementation (15 g/day) decreased milk yield by 0.8 kg/day. The DM intake of treatment cows was approximately 1 kg less per day than controls, which could indicate improved feed conversion efficiency (FCE); however, statistical analysis was not performed on these data. Aspergillus oryzae supplemented cows did have increased milk fat content, but ECM was unaffected. The forage to concentrate ratio (F:C) in TMRs offered to cattle may have led to different responses in these experiments. In the first study, F:C was 59:41 and in the second it was 44:56. Although both rations were corn and alfalfa based, the increased fibre content of the TMR in Experiment 1 may have had improved digestibility due to the fibrolytic activity of A. oryzae, leading to increased energy availability and consequently increased milk yield. These two experiments demonstrate that even when the same dose of the same A. oryzae extract is administered to cows on similar diets, very different responses from treatment can occur. This hinders the use of fungal fermentation products in cattle, as inconsistent outcomes do not encourage industry uptake. The effects of the A. oryzae extract, Amaferm®, in dairy cattle have been assessed using meta-analysis. The results of the analysis of 26 studies by Cantet et al. (Reference Cantet, Palladino, Ocasio, Bargo and Ipharraguerre2025), found decreases in milk fat and protein, and increases in milk yield, ECM, DMI, rumen pH and acetate: propionate in the rumen. This demonstrates the power of meta-analysis for consolidating and analysing the effects of treatments where individual studies present inconsistencies. These analyses can increase the industry acceptance of such products, by providing strong evidence of the effects likely to be observed on farm.
There is limited literature examining the use of Aspergillus products in young cattle. Anil et al. (Reference Anil, Chatterjee, Singh, Yadav and Mohammad2022) measured the effect of two Aspergillus niger fermentation extracts with xylanase and cellulase activities of 8000 and 16,000 U/kg DM and 12,000 and 24,000 U/kg DM, respectively, in 7-month-old crossbred Jersey heifers. Heifers were fed a TMR consisting of concentrates, chaffed rice paddy straw and oat fodder, and both treatments increased TMR digestibility, feed intake and FCE. Conversely, when Vaz-Ramírez et al. (Reference Vaz-Ramírez, Curbelo-Rodríguez and Ortiz-Colón2021) supplemented pre-weaned Jersey and Holstein calves 2 g/day of A. niger and A. oryzae fermentation product, no effects on growth or health status were measured. It is possible that aerobic fungi and their fermentation products could enhance the performance of calves through increased fibre digestion and rumen development. However, there is sparse evidence to validate or refute this hypothesis and the topic requires further investigation.
Yeasts
The use of yeast additives in cattle has been rigorously researched and reviewed (Beev et al., Reference Beev, Todorova and Tchobanova2007; Shurson, Reference Shurson2018; Cangiano et al., Reference Cangiano, Yohe, Steele and Renaud2020; Baker et al., Reference Baker, Kraft, Karnezos and Greenwood2022), and the volume of work is such that comprehensively evaluating all the literature examining yeast supplementation in dairy cattle is beyond the scope of this paper; as such we will discuss the principles of adding yeast to cattle diets, and associated advantages and challenges. Saccharomyces cerevisiae supplements can be purely live cells, cultures containing live or inactive cells and fermentation metabolites, nutritional by-products from brewing, or cell components (Shurson, Reference Shurson2018). Viable yeast additives are sometimes beneficial in the rumens of mature cattle and calves, where the probiotic effects of S. cerevisiae can stimulate the growth of cellulolytic and lactate-utilizing bacteria (Baker et al., Reference Baker, Kraft, Karnezos and Greenwood2022). Propagation of these microbial communities often leads to enhanced fibre digestion, increased butyrate production, a lower acetate to propionate ratio, and increased pH stability in the rumen; which can cause beneficial responses in DMI, FCE, rumen development, lactation, growth, reduced output of CH4 and animal health (Beev et al., Reference Beev, Todorova and Tchobanova2007; Baker et al., Reference Baker, Kraft, Karnezos and Greenwood2022). Saccharomyces cerevisiae is a facultative anaerobe, so can de-oxygenate the rumen, producing a more favourable environment for anaerobic microbe growth and fibre degradation (Baker et al., Reference Baker, Kraft, Karnezos and Greenwood2022). This is particularly important in calves, where transitioning from liquid to solid feed relies on microbial and histological development in the rumen. Yeast supplementation can lead to improvements in feed intake and growth in calves. However, of 17 studies reviewed by Cangiano et al. (Reference Cangiano, Yohe, Steele and Renaud2020) examining the effects of yeast culture or live yeast in calves, only five studies realized an increase in weight gain (Lesmeister et al., Reference Lesmeister, Heinrichs and Gabler2004; Galvão et al., Reference Galvão, Santos, Coscioni, Villaseñor, Sischo and Berge2005; Zhou et al., Reference Zhou, Zhen, Yan, Li and Wgh2009; Brewer et al., Reference Brewer, Anderson, Yoon, Scott and Carlson2014; Villot et al., Reference Villot, Ma, Renaud, Ghaffari, Gibson, Skidmore, Chevaux, Guan and Steele2019), and no studies measured an effect on FCE. The effects of yeast supplementation on GI tract development were much more consistent: of six studies that measured the effect of treatment on gut, haematological or microbiome variables, five found a positive effect (Kaldmäe et al., Reference Kaldmäe, Suurmets, Järveots, Suuroja and Kärt2008; Hučko et al., Reference Hučko, Bampidis, Kodeš, Christodoulou, Mudřik, Poláková and Plachý2009; Brewer et al., Reference Brewer, Anderson, Yoon, Scott and Carlson2014; Xiao et al., Reference Xiao, Alugongo, Chung, Dong, Li, Yoon, Wu and Cao2016; Harris et al., Reference Harris, Liang, Sharon, Sellers, Yoon, Scott, Carroll and Ballou2017), but only Brewer et al. (Reference Brewer, Anderson, Yoon, Scott and Carlson2014) also realized an increase in growth. This indicates that S. cerevisiae does not always improve primary response variables in calves but reliably promotes health and gut development.
Similarly to filamentous fungal additives, the effects of yeast cultures can be probiotic, prebiotic and/or nutritional. Cultures contain live or inactive cells, culture media, and compounds produced during fermentation, including organic acids, peptides, enzymes, esters and alcohols. The profile of compounds produced is dependent on S. cerevisiae strain, the composition of the culture media and the conditions under which fermentation takes place (Shurson, Reference Shurson2018). Saccharomyces cerevisiae is rich in amino acids, lipids, minerals and vitamins that can be utilized directly by cattle, or by rumen microbes to produce VFAs and microbial protein (Baker et al., Reference Baker, Kraft, Karnezos and Greenwood2022). The bioavailability of nutrients in these products can be highly variable and can be reduced by the formation of Maillard products from high drying temperatures (Shurson, Reference Shurson2018). Similarly to products containing only active yeast, yeast cultures can enhance the abundance and activities of beneficial microbes in the rumen, leading to improved FCE, productivity and animal health. However, because the composition of different yeast cultures can be varied depending on culture conditions, yeast strain and viability, there can be differences in animal responses to supplementation (Beev et al., Reference Beev, Todorova and Tchobanova2007).
A meta-analysis performed by Ondarza et al. (Reference Ondarza, Sniffen, Dussert, Chevaux, Sullivan and Walker2010) demonstrated that live yeast supplementation is associated with increased milk production in cattle, but that this was not associated with increased DMI, suggesting that improved lactation could be caused by enhanced rumen function and FCE rather than feed intake. As well as scavenging oxygen and improving fibre degradation, S. cerevisiae can prevent lactate accumulation in the rumen by outcompeting lactate producing microbes for fermentable sugars (Ding et al., Reference Ding, Chang, Zhao, Zhou, Ren and Meng2014) and stimulating the growth and activities of lactate metabolizing bacteria (Fonty and Chaucheyras-Durand, Reference Fonty and Chaucheyras-Durand2006). This leads to increased pH and pH stability in the rumen, limiting the risk of acidosis which is highly prevalent in cattle fed large quantities of starch; although some studies have found that live S. cerevisiae does not affect rumen pH (Ambriz-Vilchis et al., Reference Ambriz-Vilchis, Jessop, Fawcett, Webster, Shaw, Walker and Macrae2017). As with studies investigating aerobic and anaerobic fungal feed additives, variation from heterogenous yeast product composition can be compounded by dietary, environmental, and animal differences in experiments. Given the abundance of research examining yeast in cattle, the strength of evidence for the efficacy of specific cultures can sometimes be quantified through meta-analysis. This highlights how consolidation of existing research on individual fungal additives can provide greater insights into treatment effects without the need for further in vitro or in vivo experimentation.
Yeast cell walls contain MOS and β-glucans that can have beneficial effects on immunity, gut health, and productivity (Broadway et al., Reference Broadway, Carroll and Burdick Sanchez2015). Cell wall contents can be extracted from fragmented yeast cells or be a component of hydrolysed yeast cultures. MOS and β-Glucans are mycotoxin binders, can regulate the activation of the hypothalamic–pituitary–adrenal axis, modulate the immune system by interacting with immune cells, and enhance the phagocytosis of pathogens by leucocytes (Pedro et al., Reference Pedro, Lima, Fróis-Martins, Leal, Ramos, Martins, Cabrita, Fonseca, Maia and Vilanova2021). As biological response modifiers, MOS and β-Glucans can affect the functionality of macrophages and neutrophils which can lead to enhanced production of pro-inflammatory cytokines when animals are challenged by stress (Broadway et al., Reference Broadway, Carroll and Burdick Sanchez2015). Dairy cattle encounter stress throughout their lives, including birth, weaning, heat stress, social stress, disease and calving. Improved humoral and innate immunity from MOS and β-Glucan supplementation can increase the capacity of cattle to tolerate and recover from environmental and disease stressors (Ma et al., Reference Ma, Shah, Shao, Wang, Zou and Kang2020). In calves, yeast cell walls can enhance GI tract histology, increasing the length and width of papillae in the rumen due to increased butyrate production from Butyrivibrio spp., for which yeast and yeast cell walls provide a more favourable environment for growth (Ma et al., Reference Ma, Shah, Shao, Wang, Zou and Kang2020). Yeast culture supplementation has also led to increased villus height to crypt depth ratio in the duodenum, jejunum and ileum in neonatal calves (Xiao et al., Reference Xiao, Alugongo, Chung, Dong, Li, Yoon, Wu and Cao2016). The actions of yeast cultures and cell walls that lead to improved small intestine histology are not well defined, but are likely due to favourable manipulation of the gut microbiome, engulfing pathogens and mycotoxins and inhibiting their ability to bind to the epithelium, activating cytokine responses in mucosa, and improving epithelial barrier function through increased expression of tight junction proteins (Xiao et al., Reference Xiao, Alugongo, Chung, Dong, Li, Yoon, Wu and Cao2016; Cangiano et al., Reference Cangiano, Yohe, Steele and Renaud2020; Ma et al., Reference Ma, Shah, Shao, Wang, Zou and Kang2020). Considerably less research has been performed to investigate pure cell wall additives compared to yeast cultures and active yeast. When yeast cultures are hydrolysed to disrupt cells, they should be able to elicit the same benefits as cell wall products but have the advantage of containing other cell components and fermentation products that can also cause beneficial responses. Further research is required to define optimal inclusion rates and dietary interactions that can hinder the efficacy of yeast products, but as highlighted by the large volume of research that has already been performed, yeast supplementation typically improves cow and calf health, which is often paired with improvements in productivity.
Conclusion
This review highlights the potential of fungal feed additives to improve the productivity of dairy cattle through enhanced feed digestibility, microbial growth, gut health and development and immunity. AF have elite fibre degrading capabilities but the utility of these additives in dairy cattle is limited by oxygen vulnerability; research to overcome this via encapsulation or further investigation into the resting stage of the AF lifecycle could increase the accessibility of these additives to industry. Aerobic fungal products can be rich in enzymes, bioactive metabolites and nutrients, which can enhance rumen fermentation variables, productivity and favourably manipulate the rumen microbiome in cattle. However, inconsistent responses to treatments limit their uptake in industry and further research is required to define optimum inclusion rates, substrate specificity and interactions with endogenous microbes and co-cultivation products. Notably, most of the published literature examining AF and aerobic fungal additives comes from in vitro experimentation; a greater number of in vivo studies are required to define additive interactions in animal models, and meta-analyses of current literature could provide further insights into product efficacy. Conversely, yeast and yeast components have been researched comprehensively in vitro, in vivo and in meta-analyses. Investigation into potential synergies between yeasts and other fungal additives could increase the already broad range of benefits that can occur from yeast supplementation in cattle. Across all fungal additive types, existing research demonstrates that achieving consistent responses from treatments is often challenging, with inconsistent responses limiting industry uptake. Defining fungal additive interactions at dietary, cellular, and microbiome levels will increase the utility of these products on farm by informing optimal conditions for efficacy.
Funding statement
This study was supported by the University of Glasgow and Alltech as part of a PhD studentship.
Competing interests
No conflicts of interest.
Ethics approval
This study was purely desk based so did not require ethical approval.
Declaration of Generative AI and AI-assisted technologies in the writing process
The authors did not use any artificial intelligence assisted technologies in the writing process.


 Open access
Open access