


Emotion regulation is the process of influencing the experience, intensity and duration of an emotional experience, Reference Gross1 and is hypothesised to be underpinned by a complex interplay between frontal and limbic brain regions. Emotion regulation has been predominantly mapped to activation in regions of the prefrontal cortex (PFC), that is, the dorsolateral PFC (dlPFC) and ventrolateral PFC (vlPFC), the ventromedial PFC (vmPFC) including the orbitofrontal cortex (OFC), and adjacent structures such as the pre-supplementary motor area (preSMA) and the anterior cingulate cortex (ACC). Reference Etkin, Büchel and Gross2–Reference Ochsner and Gross6 Broadly, lateral prefrontal regions (dlPFC, vlPFC) are implicated in volitional control and the modulation of subcortical affective responses, Reference Braunstein, Gross and Ochsner3,Reference Zhang, Bo, Wager and Gross7 whereas the vmPFC is involved in emotion representation and stimulus valuation. Reference Hiser and Koenigs8 The preSMA and ACC are thought to support higher-order control processes, with the preSMA contributing to conflict monitoring, voluntary regulation and adapting behavioural responses to a changing context, and dorsal versus ventral ACC subregions differentially involved in expectancy violation and emotional conflict regulation, respectively. Reference Etkin, Büchel and Gross2,Reference Kohn, Eickhoff, Scheller, Laird, Fox and Habel9 Prefrontal regions are thought to exert top-down regulatory control of regions linked to emotional reactivity and processing, especially the amygdala. Reference Phillips, Ladouceur and Drevets4,Reference Ochsner and Gross6,Reference Underwood, Tolmeijer, Wibroe, Peters and Mason10 Emotion regulation encompasses a variety of strategies, which rely on overlapping yet partly distinct prefrontal substrates that work together to modulate emotional responses. Reference Braunstein, Gross and Ochsner3,Reference Zhang, Bo, Wager and Gross7
Patients suffering from social anxiety disorder show deficits in adaptive emotion regulation, which are thought to contribute directly to core symptoms such as heightened fear of negative evaluation, self-focused attention and avoidance of social situations. Reference Hofmann11,Reference Jazaieri, Morrison, Goldin and Gross12 Consequently, improving adaptive emotion regulation is a central goal of cognitive–behavioural therapy. Reference Smits, Julian, Rosenfield and Powers13 However, not all patients respond equally well, Reference Cuijpers, Miguel, Ciharova, Harrer, Basic and Cristea14 highlighting the need to enhance and personalise treatment approaches. Understanding the neurobiological mechanisms of emotion dysregulation in social anxiety disorder can help identify biomarkers of treatment response, explain individual differences and guide the development of targeted interventions. On the neurobiological level, models highlight disrupted fronto-amygdala connectivity in social anxiety disorder at rest and during emotional processing tasks. Reference Brühl, Delsignore, Komossa and Weidt15–Reference Heitmann, Feldker, Neumeister, Brinkmann, Schrammen and Zwitserlood20 During explicit emotion regulation (i.e. cognitive reappraisal), individuals with social anxiety disorder show reduced negative connectivity between the amygdala and dlPFC/vlPFC compared with healthy controls. Reference Goldin, Manber-Ball, Werner, Heimberg and Gross21 In contrast, during implicit emotion regulation (affective labelling), patients exhibit increased negative amygdala-vmPFC connectivity that correlates with symptom severity. Reference Young, Burklund, Torre, Saxbe, Lieberman and Craske22
Although functional connectivity studies provide valuable insights into an impaired emotion regulation network in social anxiety disorder, they have two crucial limitations. First, functional connectivity reflects correlative associations between brain regions and does not allow conclusions about the directionality. For instance, a negative correlation between prefrontal cortex and amygdala activity, often interpreted as top-down regulation, could equally reflect the prefrontal cortex dampening amygdala activity or, conversely, higher amygdala reactivity leading to lower prefrontal engagement. Functional connectivity alone cannot distinguish between these possibilities. Second, the complex process of emotion regulation involves a variety of subprocesses, which in turn, act in concert to execute emotion regulation successfully. Conventional functional connectivity analyses, particularly in task-based functional magnetic resonance imaging (fMRI), often focus on statistical associations between pairs of regions and may therefore provide only a limited perspective on broader network dynamics. In contrast, dynamic causal modelling (DCM) provides a data-driven, model-based approach that addresses both limitations: it estimates effective connectivity, capturing the direction of influence between regions, while simultaneously modelling how multiple regions interact dynamically within a network, potentially offering a more complete picture of the coordinated processes underlying emotion regulation. Reference Friston, Preller, Mathys, Cagnan, Heinzle and Razi23,Reference Seghier, Zeidman, Neufeld, Leff and Price24
Two studies have applied DCM to investigate effective connectivity during emotion regulation (cognitive reappraisal) in healthy controls. Steward et al confirmed the role of the vmPFC as the central node inhibiting amygdala activation during emotion regulation. Reference Steward, Davey, Jamieson, Stephanou, Soriano-Mas and Felmingham25 Furthermore, the amygdala showed an excitatory connection to the preSMA. Morawetz and colleagues highlighted the interplay within prefrontal regions during emotion regulation, with excitatory connections from the dlPFC to the inferior frontal gyrus (IFG) and inhibitory connections from the IFG to the dlPFC. Reference Morawetz, Bode, Baudewig, Kirilina and Heekeren26 They suggest an intricate feedback mechanism guiding the different components of successful emotion regulation (such as response selection). Together, these findings underscore the importance of both fronto-limbic connectivity and intra-prefrontal connectivity in successful emotion regulation. Disruptions in any of these circuits may contribute to the emotion regulation difficulties observed in social anxiety disorder.
However, no studies to date have applied DCM to investigate emotion regulation in social anxiety disorder; existing work has focused on emotion processing. During emotional face processing, patients with social anxiety disorder show reduced inhibitory vmPFC–amygdala connectivity, suggesting disrupted automatic top-down regulation. Reference Minkova, Sladky, Kranz, Woletz, Geissberger and Kraus27,Reference Sladky, Höflich, Küblböck, Kraus, Baldinger and Moser28 Altered connectivity also involves the dlPFC, with controls showing stronger vmPFC inhibition and amygdala excitation of the dlPFC compared with patients. Reference Sladky, Höflich, Küblböck, Kraus, Baldinger and Moser28 Notably, because emotional faces are disorder-relevant threat cues in social anxiety disorder, their passive viewing may already engage regulatory processes.
The present study aimed to address this gap by using DCM to examine disturbances in the effective connectivity of emotion regulation networks in individuals with social anxiety disorder. Specifically, we sought to (a) replicate previously reported alterations in effective connectivity during emotion processing in social anxiety disorder, and (b) investigate how these networks are modulated during controlled emotion regulation. To link network-level dysfunctions to clinically relevant outcomes amenable to psychotherapy, we further examined (3) whether differences in effective connectivity predict diagnostic group (patients social anxiety disorder versus healthy controls), and relate to individual symptom severity and self-reported emotion regulation abilities. Based on prior DCM studies in healthy individuals and emotion processing research in social anxiety disorder, we hypothesised that patients would show reduced inhibitory influence from prefrontal regions, particularly the vmPFC, on the amygdala, alongside altered intra-prefrontal connectivity during emotion regulation. We further expected that these neural alterations would differentiate patients from healthy controls, and be associated with greater social anxiety symptoms and less adaptive emotion regulation strategies.
Method
Sample
This project was part of the larger ‘Transdiagnostic Investigation of Emotional Processing in Depression and Anxiety’ study. Project data were collected between October 2018 and July 2022 at the University of Münster, Germany. Neuroimaging data for 102 participants (social anxiety disorder n = 61, healthy controls n = 41) were eligible for this study (for sample characteristics, see Table 1). For a comprehensive flowchart including the total of participants assessed within the larger study as well as a detailed description of inclusion criteria, see the project preregistration on OSF (https://osf.io/s6jqc). Participants were diagnosed with social anxiety disorder using the Structured Clinical Interview for DSM-IV. Reference Wittchen29 Subsequent assessment confirmed that all included individuals also met diagnostic criteria for social anxiety disorder according to the DSM-5. 30 An additional diagnosis of acute or remitted major depressive disorder was permitted within the social anxiety disorder group; however, individuals with any other current or lifetime psychiatric disorders were excluded, except in cases of mild to moderate alcohol and/or substance misuse. The use of psychotropic medication was allowed for patients with social anxiety disorder, with the exception of benzodiazepines. For the healthy control group, any current or lifetime history of mental disorders was an exclusion criterion. All procedures contributing to this work comply with the ethical standards of the Helsinki Declaration of 1975, as revised in 2013, and were approved by the local ethics commission (‘Ethik Kommission der Ärztekammer Westfalen-Lippe und der Westfälischen Wilhelms-Universität’, 2017-579-f-S). Written informed consent was obtained from all participants.
Comparison of demographic characteristics (patients with social anxiety disorder versus healthy controls) for whole sample

LSAS, Liebowitz Social Anxiety Scale; BDI-II, Beck Depression Inventory II; CERQ, Cognitive Emotion Regulation Questionnaire.
Social anxiety diagnoses were established via the Structured Clinical Interview for DSM-IV and confirmed according to DSM-5 criteria. N = 102. Information given as mean (s.d.) unless otherwise indicated.
a. Corrected for inhomogeneous variances.
b. All 18 participants reporting psychotropic medication were taking antidepressants. Three took multiple medications (n = 2 an additional antidepressant, n = 1 a low-potency typical antipsychotic).
*p < 0.05, *** p < 0.001.
Experimental paradigm
Magnetic resonance imaging (MRI) comprised a structural T1-weighted scan, diffusion tensor imaging (DTI) and fMRI, including a face-matching task, an emotion regulation task and resting state. Only the emotion regulation task was relevant for the present project. Participants completed an event-related emotion regulation task adapted from established paradigms. Reference Kuo, Fitzpatrick, Metcalfe and McMain31 A schematic overview of the task is shown in Fig. 1. All procedural details as well as statistical analysis of behavioural rating data during scanning are provided in the Supplementary Material, Part A1 available at https://doi.org/10.1192/bjp.2026.10628.
Schematic illustration of the event-related emotion regulation paradigm with the conditions ‘observe’ and ‘observe and decrease emotions’. Images are selected from the Karolinska Directed Emotional Faces database. Reference Lundqvist, Flykt and Öhman32 Image IDs: AF01ANS (upper row), BM34NES (lower row). ISI, interstimulus interval.

Image acquisition and preprocessing
MRI data were acquired on a 3 T Siemens PRISMAfit scanner. Standard preprocessing (realignment, slice-time correction, co-registration, segmentation, normalisation to Montreal Neurological Institute space, and smoothing) was performed in SPM12 (for Windows; Wellcome Department of Cognitive Neurology, University College London, UK; www.fil.ion.ucl.ac.uk/spm). Motion criteria and all acquisition parameters are described in the Supplementary Material, Part A3.
Statistical analysis
The statistical analysis plan, including hypotheses, was preregistered at OSF on 28 February 2023 (https://doi.org/10.17605/OSF.IO/CBM6Z). All changes made since preregistration are summarised in a deviation protocol also available at OSF (https://osf.io/duym6/files/ct568).
DCM
For clarity, only the main steps of the DCM analysis are described here; full technical details, preprocessing parameters and further explanations of DCM as a method are provided in the Supplementary Material, Part A4 and Supplementary Tables 1 and 2. DCM is a hypothesis- and model-driven approach used to infer directed (i.e. causal) connectivity between brain regions that best explains observed neural responses. Reference Friston, Harrison and Penny33 It distinguishes between intrinsic connectivity (baseline coupling between regions) and modulatory effects (changes induced by experimental conditions), with connection strengths expressed in hertz (Hz): positive values indicate excitatory influences, and negative values inhibitory ones. Reference Friston, Preller, Mathys, Cagnan, Heinzle and Razi23,Reference Stephan, Penny, Moran, den Ouden, Daunizeau and Friston34
Node selection and time-series extraction
To define network nodes, first- and second-level general linear models were performed to identify regions consistently engaged during observation and regulation of negative faces. Time-series were extracted from five regions of interest (amygdala, vmPFC, dlPFC, vlPFC, preSMA) following established guidelines. Reference Zeidman, Jafarian, Corbin, Seghier, Razi and Price35 For each participant, the first eigenvariate of voxels around individual maxima was calculated and adjusted for task-related effects.
Model specification and group analysis
The DCM model included bidirectional intrinsic connections between all nodes, driving inputs to the amygdala, and modulatory effects of the task conditions (‘observe’ and ‘regulate’). Individual models were estimated and group differences were examined using parametric empirical Bayes, Reference Friston, Litvak, Oswal, Razi, Stephan and Van Wijk36 including regressors for mean connectivity, group (social anxiety disorder versus healthy controls) and gender. Bayesian model reduction and averaging identified parameters contributing most to model evidence. Effects are reported as posterior probabilities, representing the likelihood that a connection exists in the population given the data; a posterior probability > 0.99 was considered very strong evidence. Reference Kass and Raftery37
Leave-one-out cross-validation
To evaluate predictive utility, leave-one-out cross-validation (LOOCV) tested whether connectivity differences could predict diagnostic group (patients with social anxiety disorder versus healthy controls), symptom severity (Liebowitz Social Anxiety Scale (LSAS)) Reference Stangier and Heidenreich38 and emotion regulation difficulties (Cognitive Emotion Regulation Questionnaire (CERQ) – dysfunctional subscale). Reference Loch and Hiller39 For each participant, the group-level parametric empirical Bayes model was re-estimated excluding their data, and the resulting model was used to predict their individual outcomes. Predicted and observed values were then correlated to assess predictive accuracy. Significant correlations indicate that group-level connectivity differences have meaningful utility for predicting individual characteristics above chance. Reference Zeidman, Jafarian, Seghier, Litvak, Cagnan and Price40
Results
Behavioural ratings during fMRI paradigm
For full descriptive statistics and strategy frequencies, see Supplementary Tables 3–4 and Supplementary Fig. 2. Ratings from nine participants were missing because of technical issues (healthy controls n = 7; patients with social anxiety disorder n = 2). During the observe condition, two separate 2 (valence) × 2 (group) analyses of variance were conducted, examining angry versus neutral faces and fearful versus neutral faces, respectively. Both analyses showed strong main effects of valence (angry: F(1,84) = 51.0, p < 0.001; fearful: F(1,84) = 34.53, p < 0.001), with negative faces eliciting more negative affect than neutral faces. In both analyses, the valence × group interaction was significant (angry: F(1,84) = 4.39, p = 0.039; fearful: F(1,84) = 6.35, p = 0.014), indicating a larger increase in negative affect from neutral to negative faces in the social anxiety disorder group than in healthy controls. Across groups, emotion regulation led to more positive ratings for negative faces (angry: F(1,84) = 7.08, p = 0.009; fearful: F(1,84) = 8.96, p = 0.004). Condition × group interactions were not significant (angry: F(1,84) = 0.23, p = 0.636; fearful: F(1,84) = 0.12, p = 0.735), indicating that participants with social anxiety disorder continued to rate their affect as more negative than healthy controls even after regulation. Groups did not differ in self-reported regulation success (angry: t (84) = −0.14, p = 0.444; fearful: t(83) = −0.91, p = 0.183). Post-scan reports indicated that both groups used a mix of adaptive and maladaptive regulation strategies, although the social anxiety disorder group descriptively endorsed maladaptive strategies more often (Supplementary Table 4).
DCM results
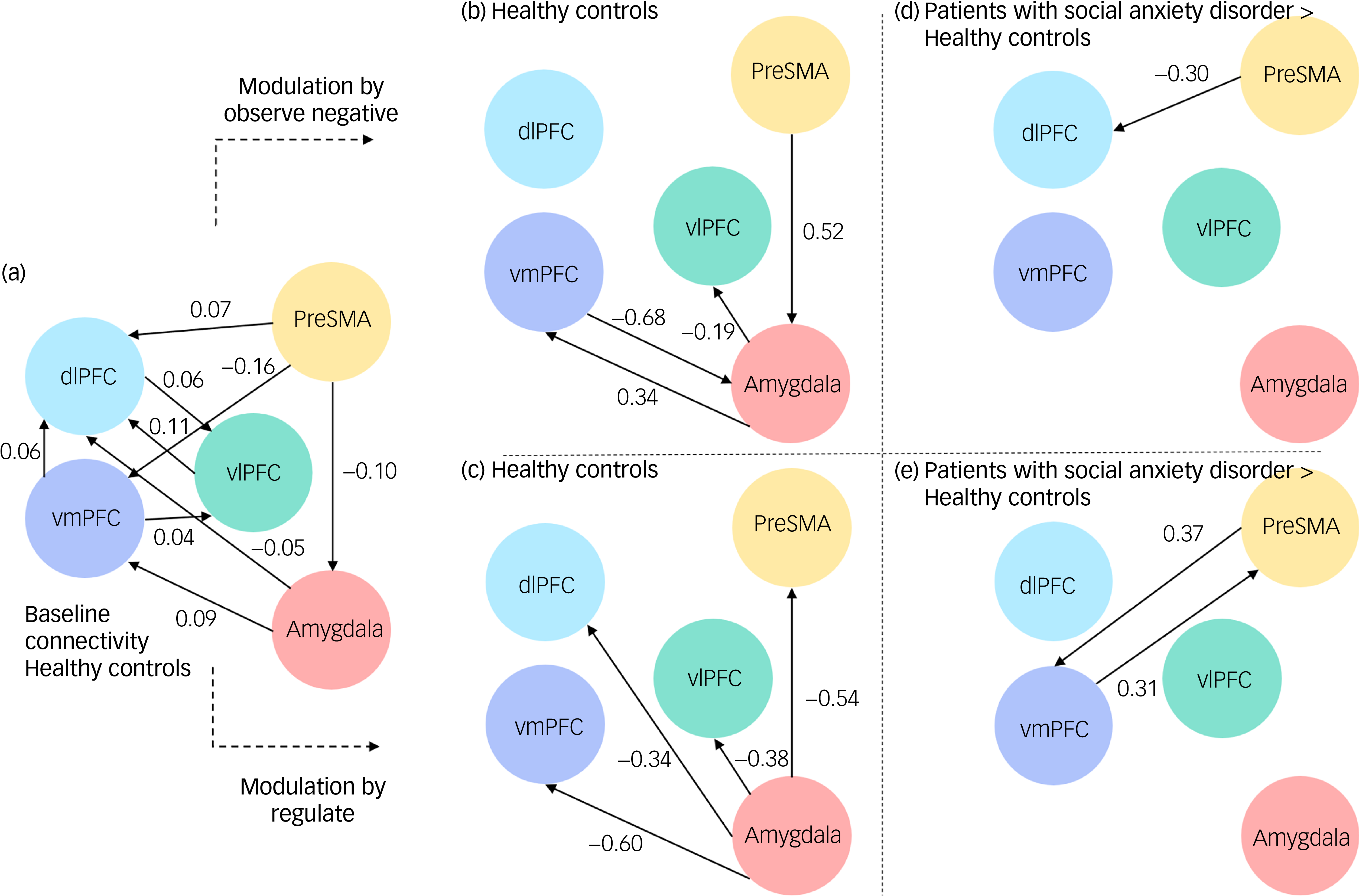
The expected values and posterior probability for all connections are reported in Supplementary Tables 5.1–5.3. Since input was mean-centred in this study, the baseline connectivity reflects the mean connectivity across both conditions (emotion processing and regulation). Accordingly, the reported condition-specific modulations (i.e. increases or decreases in connection strength) reflect changes relative to this averaged baseline, rather than overall connectivity strength. To aid interpretation, we first describe the effective connectivity patterns observed in healthy controls during the ‘observe’ and ‘regulate’ conditions, which serve as a reference model of typical network functioning. Group differences are then reported relative to this healthy control reference, that is, values indicate how connectivity in social anxiety disorder differs from healthy controls under the same condition (see also Fig. 1).
Effective connectivity in healthy controls
Figure 1 illustrates the effective connectivity between nodes within healthy controls regarding baseline connectivity (a) and its modulation by the two conditions (b, c). During the processing of negative emotional faces, healthy controls demonstrated enhanced positive modulation from the amygdala to the vmPFC (expected value: 0.34 Hz, posterior probability: 1), and negative modulation from the amygdala to the vlPFC (expected value: −0.19 Hz, posterior probability: 1). Furthermore, they showed negative modulation from the vmPFC back to the amygdala (expected value: −0.68 Hz, posterior probability: 1), and positive modulation from the preSMA to the amygdala (expected value: 0.52 Hz, posterior probability: 1). During emotion regulation, healthy controls showed greater negative modulation from the amygdala to all of the prefrontal regions (dlPFC: expected value: −0.34 Hz, posterior probability: 1; vlPFC: expected value: −0.38 Hz, posterior probability: 1; vmPFC: expected value: −0.60 Hz, posterior probability: 1) and the preSMA (expected value: −0.54 Hz, posterior probability: 1).
Differences in effective connectivity between patients with social anxiety disorder and healthy controls
Figure 2 illustrates the difference between healthy controls and social anxiety disorder in their modulation of connectivity by the two conditions (d, e). During the observation of negative faces, patients with social anxiety disorder demonstrated enhanced negative connectivity from the preSMA to the dlPFC compared with healthy controls (expected value: −0.30 Hz, posterior probability: 0.99). During emotion regulation, patients with social anxiety disorder showed enhanced positive connectivity form the preSMA to the vmPFC (expected value: 0.37 Hz, posterior probability: 0.99), and vice versa (expected value: 0.31 Hz, posterior probability: 0.99), compared with healthy controls.
Baseline connectivity in (a) healthy controls and its modulation by (b) condition ‘observe negative’ and by (c) condition ‘regulate’. Group differences between participants with social anxiety disorder in their modulation by (d) condition ‘observe negative’ and by (e) condition ‘regulate’. dlPFC, dorsolateral prefrontal cortex; PreSMA, pre-supplementary motor area; vlPFC, ventrolateral prefrontal cortex; vmPFC, ventromedial prefrontal cortex.

There was no evidence for connectivity differences between healthy controls and patients with social anxiety disorder regarding connections between the amygdala and prefrontal regions in either condition.
LOOCV
For the observed differences in effective connectivity between healthy controls and patients with social anxiety disorder (i.e. regarding the connections ‘preSMA→dlPFC’, ‘preSMA→vmPFC’, ‘vmPFC→preSMA’), LOOCV was performed to determine whether these parameters could significantly predict diagnostic group, symptom severity and emotion regulation difficulties, respectively. Regarding classification of diagnostic group, the overall effect of connectivity differences resulted in a significant out-of-sample correlation of r = 0.24 (p = 0.009) between the predicted and observed diagnostic group (see Supplementary Table 6 and Supplementary Fig. 3). For symptom severity (LSAS scores), a marginal significant out-of-sample correlation (r = 0.17, p = 0.048) was found only for the difference in effective connectivity during the ‘observe’ condition (regarding the connection ‘preSMA→dlPFC’, Supplementary Table 7 and Supplementary Fig. 4). However, this effect was accompanied by wide confidence intervals, suggesting limited robustness and should therefore be interpreted with caution. No significant out-of-sample correlation was observed for emotion regulation difficulties (CERQ – dysfunctional subscale, Supplementary Table 8 and Supplementary Fig. 5).
Discussion
This study investigated differences in effective connectivity during emotion processing and regulation between controls and patients with social anxiety disorder, using DCM. In healthy controls, during the observation of negative facial expressions, the best-performing DCM model suggested ‘bottom-up’ communication from the amygdala to prefrontal regions (i.e. the vmPFC and vlPFC), and ‘top-down’ from prefrontal regions (i.e. vmPFC and the preSMA) to the amygdala. During emotion regulation, we unexpectedly observed negative modulation from the amygdala to all included prefrontal regions. Contrary to our hypothesis, patients with social anxiety disorder did not differ in their amygdala–frontal connectivity from controls, but only regarding their effective connectivity within prefrontal regions, with a negative modulation from the preSMA to the dlPFC during emotion processing, and bidirectional positive modulations between the preSMA and the vmPFC during emotion regulation. Using LOOCV, the overall pattern of connectivity differences significantly predicted diagnostic group, indicating that these alterations carried discriminative value for distinguishing patients from controls. A marginal association also emerged for symptom severity regarding the predictive strength of the preSMA→dlPFC connection during the observation of negative faces. No significant predictive relationship was found for emotion regulation difficulties.
As previous studies have shown, emotion processing and regulation relies on the intricate interplay of different prefrontal and limbic areas Reference Braunstein, Gross and Ochsner3,Reference Zhang, Bo, Wager and Gross7 : The vmPFC is widely recognised as a key hub within the brain’s emotion-processing network: It integrates input from the amygdala to evaluate the emotional significance of stimuli to generate adaptive regulatory responses (for review, see Hiser and Koenigs Reference Hiser and Koenigs8 ). In turn, the vmPFC has been shown to exert inhibitory influence on the amygdala, especially during automatic emotion regulation when demands for effortful emotional control are low. Reference Quirk and Beer41 In contrast to the vmPFC, the vlPFC and the dlPFC seem less directly involved in initial emotion processing per se, but rather engaged in higher-order cognitive control and effortful or explicit emotion regulation processes (such as reappraisal and attentional control, Reference Braunstein, Gross and Ochsner3 ). In light of this, the positive modulation from the amygdala to the vmPFC found during observation in healthy controls in our study might reflect the input of emotionally salient information from subcortical to cortical areas, enabling the vmPFC to assess the relevance of the emotional faces. In turn, the negative modulation from the vmPFC back to the amygdala can be interpreted as a feedback mechanism aimed at dampening amygdala output, since an emotional reactivity to the faces seemed unnecessary. Although participants were instructed to merely ‘observe’ the emotional face during one of the conditions, it can be assumed that the brain likely still engaged in some form of automatic emotion regulation as part of a functional handling of emotional information. The reciprocal interaction fits with models of implicit emotion regulation, where the vmPFC continuously monitors and adjusts emotional responses without requiring conscious effort or explicit instructions. Reference Braunstein, Gross and Ochsner3,Reference Quirk and Beer41
Notably, we did not observe modulatory connections from the vlPFC to the amygdala that would indicate effortful top-down regulatory control. Instead, we found negative modulatory connections from the amygdala to the vlPFC. Interestingly, the same pattern of negative amygdala–vlPFC modulation emerged during the emotion regulation condition, and extended to all included prefrontal regions. This is at odds with classical models of top-down emotion regulation, which emphasise prefrontal control over limbic regions (for meta-analysis, see Berboth and Morawetz Reference Berboth and Morawetz42 ). Negative connectivity from the amygdala to the prefrontal regions has often been interpreted as ‘emotional hijacking’, where strong bottom-up emotional signals suppress cognitive control regions to prioritise rapid emotional responses. Reference Bishop43 However, because our emotional faces were mildly negative and rated relatively neutral by healthy controls, this explanation seems unlikely. Unlike highly aversive stimuli used in other studies, our social anxiety disorder-relevant faces likely had limited impact on controls, reducing the need for additional regulation. The observed inverse modulation may instead reflect an adaptive mechanism, in which the amygdala fine-tunes prefrontal engagement to prevent excessive control and promote flexible regulation. Previous studies suggest that the amygdala can modulate prefrontal activity depending on context, supporting coordinated network dynamics rather than strictly top-down regulation. Reference Pessoa44 This view aligns with more recent perspectives highlighting substantial overlap between emotion-generating and emotion-regulating frontal areas. Reference Zhang, Bo, Wager and Gross7
Contrary to much of the existing literature on social anxiety disorder, Reference Bas-Hoogendam and Westenberg16 we did not observe altered prefrontal–amygdala connectivity in patients with social anxiety disorder compared with controls during either observation or instructed regulation. Several explanations may account for this finding. One possibility relates to methodological aspects. Although facial expressions are among the most frequently used stimuli in social anxiety disorder research, Reference Gentili, Cristea, Angstadt, Klumpp, Tozzi and Phan45 authors have argued that they lack ecological validity. Reference Heitmann, Feldker, Neumeister, Brinkmann, Schrammen and Zwitserlood20 Nevertheless, in our study these stimuli were rated as more negative by patients with social anxiety disorder compared with controls, and successfully elicited amygdala activation, suggesting that they were emotionally engaging. Another methodological factor concerns task design: unlike many previous studies that instructed participants to apply a specific strategy such as reappraisal, our paradigm allowed participants to choose their emotion regulation strategy. This approach enhances ecological validity by capturing more naturalistic regulatory behaviour, but it may also introduce greater inter-individual variability (e.g. in the specific prefrontal subregions engaged by different strategies), and thus reduce sensitivity to detect consistent amygdala–prefrontal coupling effects. Consistent with this, participants in both groups reported using a wide variety of adaptive and maladaptive strategies, which may have contributed to variability in prefrontal recruitment and the absence of robust group differences in amygdala–prefrontal connectivity.
At the same time, our results indeed showed aberrant effective connectivity in patients with social anxiety disorder relative to controls, not in fronto–amygdala pathways, but within prefrontal regions themselves. These findings raise the possibility that alterations within intra-prefrontal circuitry, rather than disrupted amygdala downregulation per se, may represent a characteristic neural feature of social anxiety disorder. Supporting this interpretation, the identified differences in intra-prefrontal connections significantly predicted diagnostic group (patients with social anxiety disorder versus healthy controls), indicating that these connectivity differences carry meaningful and generalisable information at the individual level. This interpretation resonates with recent criticisms about the limited reproducibility of fronto–amygdala abnormalities in social anxiety disorder, supported by a recent meta-analysis Reference Mizzi, Pedersen, Rossell, Rendell, Terrett and Heinrichs46 reporting no significant group effects in their meta-analytic seed-to-whole-brain functional connectivity analysis (including the amygdala as a seed region). Importantly, much of the existing evidence for fronto–amygdala dysfunction stems from non-directed functional (i.e. not effective) connectivity, which may overlook the more nuanced, directional dynamics revealed by effective connectivity approaches such as DCM.
Following this line of reasoning, our findings may point to altered intra-prefrontal dynamics in social anxiety disorder, specifically involving the preSMA. The preSMA is traditionally associated with higher-order motor control and response inhibition, but its role extends to affective and cognitive domains. Reference Kohn, Eickhoff, Scheller, Laird, Fox and Habel9 In the context of emotion, the preSMA has been shown to be involved in conflict monitoring, volitional control over emotional responses and preparing adaptive behavioural responses to salient stimuli. In our study, during observation of emotional stimuli in patients with social anxiety disorder, the preSMA exerted a stronger negative influence on the dlPFC compared with healthy controls, which could indicate reduced recruitment of dlPFC-based cognitive control. One tentative interpretation is that, even in the absence of explicit regulatory demands, patients with social anxiety disorder may engage the preSMA in ways that interfere with more adaptive forms of cognitive control, possibly reflecting premature or inefficient attempts to manage emotional responses. During instructed emotion regulation, patients with social anxiety disorder showed enhanced reciprocal connectivity between the preSMA and vmPFC. This increased bidirectional coupling suggests that the preSMA becomes more strongly integrated into vmPFC-based evaluative and regulatory processes when active regulation is required. Although the precise functional implications remain to be determined, this pattern could indicate heightened readiness to monitor or adjust behaviour in response to emotional salience. Reference Etkin, Büchel and Gross2 Heightened prefrontal engagement in social anxiety during emotion regulation has been reported regularly, and is often interpreted as a compensatory mechanism to help maintain performance despite underlying emotional challenges. Reference Brühl, Delsignore, Komossa and Weidt15 Supporting this notion, our meta-analysis Reference Schrammen, Roesmann, Rosenbaum, Redlich, Harenbrock and Dannlowski47 showed that psychotherapy in social anxiety disorder (and other anxiety disorders) was associated with reduced prefrontal activation (including the supplementary motor area) suggesting that such compensatory mechanisms may normalise following psychotherapy. Notably, our meta-analysis did not identify corresponding reductions in limbic regions such as the amygdala following treatment. Although any clinical implications must be drawn with caution, this aligns with contemporary approaches in treating anxiety, that do not aim at a direct reduction of fear, but emphasise strengthening flexible regulation capacities and the ability to act adaptively in the presence of heightened emotional arousal. Reference Craske48
Importantly, although these results shift attention away from fronto-amygdala dysfunction, interpretations remain tentative because of the methodological limitations and specific challenges of DCM. Since this study is the first to investigate emotion regulation in social anxiety disorder using DCM, we based our selection of regions of interest on the few prior DCM studies examining emotion processing in social anxiety disorder and emotion regulation in healthy controls (see OSF preregistration). Other brain regions not included in the model, such as the insula or anterior cingulate cortex, Reference Brühl, Delsignore, Komossa and Weidt15 may also contribute. Thus, future research should systematically investigate additional regions implicated in emotion regulation and its subprocesses in social anxiety disorder to provide comprehensive understanding of the underlying neural mechanisms. Further, although the observed differences in effective connectivity were found to predict diagnostic group, the same parameters did not reliably predict symptom severity (LSAS) or emotion regulation difficulties (CERQ – dysfunctional subscale). This suggests that these connectivity alterations may represent a broader categorical neural signature of social anxiety disorder rather than a continuous correlate of clinical severity and emotion regulation ability. The absence of continuous associations should be interpreted cautiously, as it may reflect limited statistical power rather than a true lack of relationship. Also of note is that the ecological validity of facial expressions has been doubted. Reference Heitmann, Feldker, Neumeister, Brinkmann, Schrammen and Zwitserlood20 Nevertheless, in our study, these stimuli were rated as more negative by patients with social anxiety disorder compared with controls and successfully elicited amygdala activation, suggesting that they were emotionally engaging.
Our paradigm allowed participants to freely choose their emotion regulation strategies, increasing ecological validity by capturing more naturalistic regulatory behaviour. However, this flexibility likely introduced substantial inter-individual variability. Consistent with this interpretation, participants in both groups reported using a wide variety of adaptive and maladaptive strategies, potentially contributing to heterogeneous prefrontal recruitment and the absence of robust group differences in amygdala–prefrontal connectivity. Although unlikely, habituation and/or carryover effects from the preceding face-matching task cannot be completely ruled out. Also, although formal sample size recommendations for DCM are lacking, studies suggest that 20–40 participants per group are generally sufficient for robust group-level results. Reference Goulden, Elliott, Suckling, Williams, Deakin and McKie49,Reference Ma, Braun, Steinberg, Bjork, Martin and Keen50 Cross-validation typically requires larger effect sizes and, although classification of categorical outcomes is generally more robust, it is particularly sensitive to sample size when predicting continuous outcomes. Reference Archer, Snell, Ensor, Hudda, Collins and Riley51,Reference Varoquaux52 Further, the present analyses did not explicitly account for comorbid major depressive disorder, which limits conclusions about diagnostic specificity. Importantly, the number of participants with social anxiety disorder without any history of depression was relatively small, precluding meaningful subgroup or covariate analyses. Since the high comorbidity rate reflects the typical clinical presentation of social anxiety disorder, as observed in other samples, the observed changes in connectivity can be generalised, even if they may not be specific to social anxiety disorder. Finally, a small minority of participants with social anxiety disorder were taking antidepressant medication. Importantly, all participants fulfilled diagnostic criteria for social anxiety disorder despite treatment. Nonetheless, potential effects of medication on effective connectivity cannot be fully ruled out and should be considered when interpreting the findings.
In conclusion, this study provides novel insights into the effective connectivity dynamics underlying emotion processing and regulation in healthy controls and patients with social anxiety disorder. Generally, in healthy controls, emotion processing and emotion regulation modulated fronto-amygdala circuits, whereas differences between healthy controls and patients with social anxiety disorder in effective connectivity were observed exclusively within prefrontal regions. These findings suggest that alterations in intra-prefrontal circuitry, rather than deficits in amygdala–prefrontal coupling, may characterise aberrant emotion regulation in social anxiety disorder. Although this finding was unexpected it adds to a growing body of evidence, challenging current prominent models of fronto-limbic inhibition in emotion regulation. Overall, these findings contribute to a more nuanced understanding of the altered neural dynamics underlying emotion regulation in social anxiety and may inform targeted therapeutic approaches.
Supplementary material
The supplementary material is available online at https://doi.org/10.1192/bjp.2026.10628
Data availability
The study protocol, the pre-registered statistical analysis plan and the deviation protocol can be found via the OSF project: https://osf.io/s6jqc. Data, analytic code and research material will be made available upon reasonable request from the corresponding author, E.J.L.
Acknowledgement
During the preparation of this work the authors used DeepL and ChatGPT in order to improve language and readability. After using this tool/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Author contributions
E.J.L., J. Böhnlein, S.M., U.D., L.S. contributed to the conception or design of the study. E.S., H.M., J. Böhnlein, S.M., J. Bauer, J.G., D.G., L.S., K.F., J.R., N.O., R.R., U.D., E.J.L. contributed to the acquisition. E.S., A.J.J., B.J.H., E.J.L., D.G. contributed to the analysis. E.S., A.J.J., B.J.H., E.J.L., T.M.S., R.A.V., A.M. contributed to the interpretation of data for the work. E.S., E.J.L., A.J.J., B.J.H. participated in drafting the work. All authors revised the final version critically for important intellectual content. All authors approved the final version to be published. All authors agreed to be accountable for all aspects of the work.
Funding
This work was funded by the Innovative Medizinische Forschung of the medical faculty of Münster (to E.J.L., grant numbers LE121703 and LE121904) and in part by the consortium grant Trajectories of Affective Disorders from the German Research Foundation (DFG) SFB/TRR 393 (project grant number 521379614). U.D. was funded by the DFG (grant numbers FOR2107 DA1151/5-1, DA1151/5-2, DA1151/9-1, DA1151/10-1, DA1151/11-1, project grant number 521379614) and the Interdisciplinary Center for Clinical Research of the Medical Faculty of Münster (grant Dan3/022/22). J.R. was supported by the LOEWE programme of the Hessian Ministry of Science and Arts (grant number LOEWE1/16/519/03/09.001(0009)/98).
Declaration of interest
J.R. received speaker’s honoraria from Janssen, Hexal, Neuraxpharm and Novartis. All remaining authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Transparency declaration
The authors affirm that the manuscript is an honest, accurate, and transparent account of the study being reported; no important aspects of the study have been omitted, and any discrepancies from the study as planned (and, if relevant, registered) have been explained (see also deviation protocol: https://osf.io/ct568).





 Open access
Open access
eLetters
No eLetters have been published for this article.