


Introduction
Rapeseed (Brassica napus), one of the world’s most crucial oil crops, produces straw that is increasingly recognized as a valuable agricultural resource due to its nutrient-rich composition (Stolarski et al. Reference Stolarski, Welenc and Krzyżaniak2024; Suchocki Reference Suchocki2024). However, despite its abundance, a significant portion of rapeseed straw is often underutilized or discarded, leading to resource waste and environmental pressure (Qiu et al. Reference Qiu, Cui and Gao2024; Wang et al. Reference Wang, Nabi and Sajid2024a). Rapeseed straw, a lignocellulosic biomass rich in cell wall components and protein, can be bio-converted via enzymatic hydrolysis or solid-state fermentation into nutritionally enhanced roughage. This strategy provides a viable pathway to mitigate forage shortages in livestock production (Zhang et al. Reference Zhang, Wang and Wang2025). This valorization strategy offers a practical pathway to alleviate the shortage of high-quality forage supply in livestock production (Chen et al. Reference Chen, Fan and Li2024; Kamal et al. Reference Kamal, Aldhalmi and Abd El-Hack2025; Shah et al. Reference Shah, Zhang and Shahid2025). Furthermore, the conversion of lignocellulosic agricultural residues like rapeseed straw into single-cell protein presents a promising strategy to alleviate the escalating global demand for sustainable protein in animal feed, thereby contributing to a more circular bioeconomy (Gao et al. Reference Gao, Khoo and Zhang2025; Sekoai et al. Reference Sekoai, Roets-Dlamini and O’Brien2024). Regarding structural properties, the epidermal cuticle of rapeseed straw possesses considerable resistance to mechanical breakdown due to its high degree of lignification. Regarding nutritional properties, anti-nutritional factors such as erucic acid and glucosinolates substantially reduce metabolizable energy by inhibiting voluntary intake and impairing nutrient digestion and absorption (Axentii and Codină Reference Axentii and Codină2024; Binczarski et al. Reference Binczarski, Zuberek and Fraczyk2025; Feng Reference Feng2007).
The physicochemical properties of rapeseed straw necessitate pretreatment and nutritional enhancement prior to practical application. The technical focus lies in employing physical or biological means to disrupt the compact architecture of lignin-carbohydrate complexes, thereby reducing their polymerization degree, while simultaneously eliminating or converting anti-nutritional constituents. Ensiling, enzymatic treatment, fermentation, and ammoniation improve rapeseed straw’s nutrition and feeding efficacy. These mainstream technologies demonstrate that rapeseed straw is a valuable resource for high-quality roughage.
As a pivotal forage processing method, ensiling offers distinct advantages in the preservation of feed nutrients. This technique effectively safeguards protein structural integrity while concurrently improving feed palatability, thereby promoting livestock feed consumption (Manganyi et al. Reference Manganyi, Tjelele and Mbatha2024). The ultimate quality of silage is largely dependent on the biological activity of lactic acid bacteria (LAB) during the fermentation process (Zhu et al. Reference Zhu, Zhao and Yan2025). The LAB effectively inhibit the growth and reproduction of spoilage microorganisms while concurrently driving the conversion of water-soluble carbohydrates (WSCs) into lactic acid (LA), thereby resulting in a decrease in system acidity (Wang et al. Reference Wang, Sun and Zhang2024b). The increased acidity within the silage system significantly inhibits the growth of various spoilage microorganisms, particularly the predominant species of the Enterobacteriaceae family, Clostridium, yeasts, and molds (Snyder et al. Reference Snyder, Martin and Wiedmann2024). Minimizing dry matter (DM) loss is a critical objective during silage storage. To this end, fully leveraging microbial resources to regulate the fermentation process while reducing energy consumption as much as possible to preserve nutrients is particularly important. The cell walls of straw and forage plants contain various polysaccharides, primarily consisting of cellulose, hemicellulose, and pectin (Undersander and Sharpe Reference Undersander and Sharpe2025). Cellulase (CE) has been successfully incorporated into forage silage and ruminant diets, showing notable efficacy in improving fiber utilization (Głazowska et al. Reference Głazowska, Gundersen and Heiske2025). This enzyme acts synergistically with rumen microbiota, promoting the breakdown of cellulose, the most abundant polysaccharide in plant cell walls, which enhances the rumen’s overall fiber-digesting efficiency. When applied to forage silage and grain feed, CE pretreatment increases their nutritional value by releasing more fermentable sugars and improving digestibility in livestock.
This study aimed to evaluate how the synergistic addition of Lactiplantibacillus plantarum and CE affects fermentation quality and microbial composition in rapeseed straw silage, hypothesizing that this combination optimizes the community structure by liberating fermentable sugars from structural carbohydrates.
Materials and methods
Substrate and silage preparation
Rapeseed straw was collected from the sampling plot of the Hulunbuir Grassland Ecosystem National Field Scientific Observation and Research Station, Chinese Academy of Agricultural Sciences. The study plot is geographically positioned at coordinates 49°23′13″ N, 120°02′47″ E, with an elevation varying between 627 and 635 m above sea level. The region has a mean annual temperature of −2.4°C and an average annual precipitation between 380 and 400 mm (Bo et al. Reference Bo, Dan and Ji2023). The plot features a plain topography with a sunny mid-slope aspect. The soil is classified as light loamy chernozem and dark chestnut soil developed on loess parent material.
Whole rapeseed plants were collected at physiological maturity (pod stage) on August 20, 2023. Following seed separation through mechanical threshing, the remaining straw was harvested using a commercial straw collector (Model HFD 50 × 80, Linyi Fuda Tools Co., Ltd., Beijing, China). The collected rapeseed straw was wilted for 3 hours under field conditions before being mechanically chopped into segments with an average length of 3 cm. Representative fresh samples (400 g) were rapidly frozen on-site and subsequently transported to the laboratory for proximate composition analysis. The determined DM (57.47%) of the rapeseed straw, along with other chemical compositions and microbial counts, is presented in Table 1. In this study, two external additives were introduced. The initial additive comprised L. plantarum strain MTD-1, sourced from Luke Biological Technology Co., Ltd. (located in Jiangsu, China), which exhibited a viable bacterial concentration of 1 × 1010 colony-forming units (CFU)/g. Prior to ensiling, the L. plantarum strain was cultured in de Man, Rogosa, and Sharpe (MRS) broth at 30°C to prepare the inoculant. The second was a commercial CE AC (derived from Acremonium cellulolyticus), supplied by Meiji Seika Pharma Co., Ltd. (Japan), possessing a total enzymatic activity of 50,000 U/g. The trial comprised four treatments: a control treatment receiving distilled water (CK), a treatment with L. plantarum alone (Lp treatment, applied at 1.0 × 10⁶ CFU/g of fresh weight), a CE only treatment (CE treatment, with an application rate of 50,000 U/g of fresh weight), and a synergy between microbial inoculants and exogenous enzymes (designated as combination of Lactiplantibacillus plantarum and cellulase [CELp] co-treatment). The dose of 1.0 × 10⁶ CFU/g was chosen as it is optimal for dominating the microbial community, rapidly reducing pH, and enhancing fermentation quality without causing over-acidification. The CE dosage of 50,000 U/g fresh weight was determined through preliminary experiments assessing enzymatic activity on rapeseed straw substrate (Ma et al. Reference Ma, Li and Zhao2025; Sifeeldein et al. Reference Sifeeldein, Xianjun and Zhihao2018; Wang et al. Reference Wang, Wang and Zhou2018). A completely randomized experimental design was implemented, with six biological replicates per treatment. All treatments received the same volume of distilled water. Approximately 250 g of chopped rapeseed straw were aseptically packed into polyethylene fermentation bags (32 × 26 cm). Following vacuum sealing, all samples were subjected to fermentation at a constant temperature of 25°C for 60 days. To ensure uniformity of fermentation conditions, all bags were maintained in identical positions throughout the ensiling period.
Chemical and microbial characteristics of substrates before ensiling

Table 1 Long description
The table presents the chemical and microbial characteristics of rapeseed straw before ensiling, focusing on dry matter, protein content, fiber, carbohydrates, and microbial counts. Rapeseed straw contains 57.47% dry matter and 4.73% crude protein, with acid detergent fiber at 28.40% and neutral detergent fiber at 43.83%. Water-soluble carbohydrates are measured at 4.24%. Microbial analysis shows lactic acid bacteria at 4.49 log10 CFU per gram of fresh weight, aerobic bacteria at 5.60, yeasts at 6.02, coliform bacteria at 7.49, and mold at 7.69. The standard error of the mean is provided for each measurement, indicating variability in the data. The high presence of mold and coliform bacteria suggests potential challenges in microbial management during ensiling.
DM, dry matter; FM, fresh materials; CFU, colony-forming units.
Analytical procedures for evaluating silage quality
After a 60-day fermentation period, both fresh rapeseed straw and the resulting silage samples were dried in a forced-air oven at 65°C for 48 hours to determine DM content (Xue et al. Reference Xue, Liu and Wang2019). For the purpose of subsequent chemical analysis, all specimens were pulverized by a grinder fitted with a 1 mm sieve (Model FM100, manufactured by Teste Instrument Co., China). Crude protein (CP) concentration was determined following the standard Kjeldahl procedure (Davidson et al. Reference Davidson, Mathieson and Boyne1970). The neutral detergent fiber (NDF) and acid detergent fiber (ADF) analyses were performed following the fiber analytical procedure established by Van Soest et al. (Reference Van Soest, Robertson and Lewis1991). The organic acid (LA), acetic acid (AA), propionic acid (PA), and butyric acid (BA) concentrations in the sample filtrates were determined by high-performance liquid chromatography equipped with an ultraviolet detector (detection wavelength: 210 nm). The chromatographic conditions were set as follows: 3 mmol/L perchloric acid solution served as the mobile phase, the flow rate was maintained at 1.0 mL/min, and the column temperature was controlled at 50°C (You et al. Reference You, Du and Ge2021). The concentration of WSC content was determined according to the anthrone colorimetric method (Thomas Reference Thomas1977; Yu et al. Reference Yu, Wang and Cao2020).
Microbial counts in fresh materials (FM) were determined by plate counting (CFU/g FM). LAB and coliforms were enumerated on MRS agar and violet red bile agar at 30°C for 48 hours. Molds and yeasts were counted on potato dextrose agar and nutrient agar at 30°C for 24 hours (You et al. Reference You, Du and Ge2021).
Bacterial community analysis
Ten grams of silage samples were mixed with 90 mL of sterile water and subjected to extraction by shaking at 160 rpm for 2 hours at 4°C to obtain microbial samples (Xiao et al. Reference Xiao, Sun and He2025). The resulting suspension underwent filtration via double-layered sterile gauze followed by multiple rinses with sterile water, thereby concentrating the remaining microbial biomass. Subsequent centrifugation was performed at 10,000 × g for 15 minutes at 4°C, and the resulting bacterial pellet was stored in an ultra-low temperature freezer at −80°C (Moore et al. Reference Moore, Arnscheidt and Krüger1999).
Total microbial genomic DNA was isolated by adhering precisely to the protocol provided with a commercially available DNA extraction kit (TIANamp Bacteria DNA Kit, model DP30202; Tiangen Biotech Co., Ltd., Beijing, China). Subsequently, the V3–V4 hypervariable region of the bacterial 16S rRNA gene was amplified using a universal primer pair (forward sequence: 5′-ACTCCTACGGGAGGCAGCA-3′; reverse sequence: 5′-GGACTACHVGGGTWTCTAAT-3′) and a standardized polymerase chain reaction system and protocol (Ni et al. Reference Ni, Wang and Zhu2017). Bioinformatic analysis of sequencing data, including diversity indices and microbial community composition, was conducted on the Omicsmart cloud platform (https://www.genedenovo.com/; accessed on 30 May 2024). The functional profiles of the bacterial community and the relative abundance of key enzymes (e.g., L-lactate dehydrogenase, xylanases, and β-glucosidase) were predicted using PICRUSt2 based on the 16S rRNA gene sequencing data and the Kyoto Encyclopedia of Genes and Genomes (KEGG) database.
Statistical evaluation
All data are presented as the mean of six replicates. A one-way ANOVA was conducted in SPSS (version 27.0) to analyze nutritional and fermentation parameters, with Duncan’s multiple range test applied for post hoc comparisons where significant differences (p < 0.05) were detected (Tallarida and Murray Reference Tallarida and Murray1987). The principal coordinate analysis (PCoA) plots were generated with the ggplot2 package, incorporating 95% confidence ellipses to clarify the treatments’ separation patterns (Valero Mora Reference Valero Mora2010). Microbial raw data were downloaded from the Dior platform, and subsequent analysis of the Venn diagram was drawn with the online analysis platform (https://www.omicshare.com/tools/home/report/reportpcoa2.html). Upset and Mantel test figures were analyzed using the online analysis website (https://chiplot.online/upset_plot.html) (Diniz-Filho et al. Reference Diniz-Filho, Soares and Lima2013; Li et al. Reference Li, Ge and Wang2021b). The bar chart and functional prediction chart of the relative abundance percentage of bacterial genus were drawn using Origin2024. The linear discriminant analysis (LDA) figure was drawn with the online analysis website, setting the threshold to >4 (https://www.bioincloud.tech/task-meta) (Chang et al. Reference Chang, He and Dang2022; Du et al. Reference Du, You and Jiang2022). The author annotated microbial community functions using the KEGG database and performed functional enzyme annotation for bacteria by employing the EC enzyme database.
Results and discussion
This study systematically analyzed the differential effects of L. plantarum addition, CE treatment, and their combined application on rapeseed straw silage, with a particular focus on elucidating the synergistic influence between the two. By integrating the experimental results with existing literature data, the research offers novel inferred metabolic profiles of explanations and robust experimental evidence for improving the quality of lignocellulosic silage through the optimization of microbial–enzyme co-processing technologies.
Effects of additives on the quality of rapeseed straw silage
As a feed ingredient, rapeseed straw exhibits inherently low CP content and high structural fiber levels, necessitating effective improvement to meet the nutritional requirements of ruminants. The effectiveness of ensiling fermentation was largely influenced by the initial population of epiphytic LAB on the raw material, with this bacterial density typically being a key determinant of the final silage quality. In practice, the natural load of epiphytic LAB on untreated rapeseed straw often falls below the minimum threshold required for high-quality ensiling (<105 CFU/g), whereas undesirable microorganisms, such as spoilage bacteria, frequently exceed recommended limits (>10⁴–10⁸ CFU/g). Such an unfavorable initial microbial profile significantly increases the risk of fermentation failure or compromised silage quality (You et al. Reference You, Du and Ge2021). Therefore, the addition of exogenous LAB inoculants represents an effective strategy for enhancing the quality of rapeseed straw silage. As a bioprocessing technology, ensiling fermentation can modify the structure of cellulose and lignin in the raw material, thereby optimizing its overall nutritional value (Liu et al. Reference Liu, Wang and Wang2024). Table 2 reveals that the core indicators for evaluating silage fermentation quality consist of the system’s pH and organic acid profile. For instance, L. plantarum predominantly transforms WSCs into short-chain organic acids through its metabolic activity. Concurrently, acid hydrolysis and enzymatic digestion can effectively degrade the digestible components of plant cell walls, which not only reduce fiber content but also promote an overall increase in acidity (Letizia Reference Letizia2024). In this experiment, compared with the CK treatment, the Lp treatment and CELp co-treatment exhibited a significant decrease in pH by 10.5% and 12%, respectively, whereas their LA content increased significantly by 38% and 33%, respectively (p < 0.05). The CELp co-treatment exhibited the highest LA content (47.68 g/kg DM). Compared to the CK treatment, the AA content (27.47 g/kg DM) decreased significantly in the CE, Lp, and CELp co-treatment, while the Lp co-treatment had significantly higher AA content (19.23 g/kg DM) than the CELp and CE treatments (p < 0.05). These findings are consistent with previous reports that L. plantarum acidifies the silage environment by enhancing homolactic fermentation. However, we did not detect BA and PA in these silages.
Fermentation quality and chemical composition in rapeseed straw silage

Table 2 Long description
The table compares fermentation quality and chemical composition of rapeseed straw silage under different treatments: control, Lactiplantibacillus plantarum, cellulase, and a combination of cellulase and Lactiplantibacillus plantarum. Key findings include the combination treatment having the lowest pH and highest lactic acid content, indicating better fermentation quality. The control treatment has the highest pH and lowest dry matter content, suggesting less effective fermentation. Propionic and butyric acids were not detected in any treatments. In terms of chemical composition, the combination treatment also shows higher dry matter content compared to control. Significant differences are noted in acid detergent fiber and neutral detergent fiber content across treatments, with the combination treatment generally showing lower fiber content, indicating improved digestibility.
DM, dry matter; CP, crude protein; ADF, acid detergent fiber; NDF, neutral detergent fiber; propionic acid and butyric acid were not detected in all wheat straw silages. Control, control treatment; CE, rape straw inoculated with cellulase treatment; Lp, rape straw was treated by inoculating Lactiplantibacillus plantarum; CELp, rapeseed straw was inoculated with cellulase and Lactiplantibacillus plantarum for treatment. SEM, standard error of mean. a, b, and c significant differences at p < 0.05 level. ND, not detected.
Effects of additives on fiber content in rapeseed straw
Numerous studies have validated that the combined use of microbial and enzymatic approaches markedly decreases the concentrations of NDF and ADF within the feedstock (Cherdthong et al. Reference Cherdthong, Suntara and Khota2020). Compared to the CK treatment, the CELp co-treatment exhibited significant reductions in ADF (17.23% of DM), NDF (49.57% of DM), and hemicellulose content (32.34% of DM; p < 0.05), while both the CE and CELp co-treatments showed a significant increase in DM content (p < 0.05). These improvements are likely attributed to the microbial–enzyme-mediated degradation of structural carbohydrates during fermentation. The breakdown of cellulose and hemicellulose polymers releases soluble low-molecular-weight compounds, which can serve as metabolic substrates for microbial growth while enhancing the bioavailability of nutrients for animal digestion. Notably, the CE treatment exhibited higher residual hemicellulose content compared to other treatments. This may be attributed to insufficient degradation of cellobiose, a product of cellulolytic hydrolysis. In this study, Xanthomonas became the dominant genus in the CE treatment, but its secreted β-glucosidase activity was relatively low (Fig. 3D), leading to the accumulation of cellobiose in the system. Although Xanthomonas could improve silage quality by producing antibacterial AA and reducing fiber crystallinity, its metabolic profile favored the accumulation of soluble oligosaccharides, resulting in only partial depolymerization of fibers. This metabolic characteristic ultimately explains the relatively limited cellulose degradation observed in the CE treatment.
Effects of additives on bacterial community structure in rapeseed straw
The present investigation was centered on elucidating the synergistic interactions between bacterial and enzymatic components within the silage processing system. The results demonstrated that this co-treatment significantly improved both the fermentation quality and the in vitro digestibility of nutrients. To elucidate the underlying microbiological basis, we systematically analyzed the diversity and composition of the bacterial community in rapeseed straw silage using 16S rRNA gene amplicon sequencing (Table 3). Sequencing of all samples yielded a total of 3,463,385 raw reads. After stringent quality control filtering, 3,460,406 high-quality valid reads were retained, with the number of sequences per sample ranging from 58,896 to 131,649. The sequencing coverage of all samples exceeded 0.99, indicating that the sequencing depth was sufficient to comprehensively capture the bacterial taxa present in the samples.
Diversity indices of rapeseed straw inoculated without or with cellulose or L. plantarum

Table 3 Long description
The table compares diversity indices of rapeseed straw treated with different inoculants, including L. plantarum and cellulase. The number of sequences and valid sequences is significantly higher in treatments with L. plantarum, cellulase, or both, compared to the control. Observed OTU numbers are highest in the control and L. plantarum treatments, while the lowest is in the combined treatment. Goods' coverage is consistently high across all treatments. The Chao1 index is highest in the control and lowest in the combined treatment, indicating a decrease in estimated species richness. The Shannon index, which measures diversity, is highest in the control and lowest in the L. plantarum treatment, suggesting variations in species evenness. All differences are statistically significant, with p-values less than 0.001.
FM, fresh materials; OTU, operation classification unit; No., number; Control, control treatment; CE, rapeseed straw inoculated with cellulase treatment; Lp, rapeseed straw inoculated with L. plantarum treatment; CELp, rapeseed straw inoculated with cellulase and L. plantarum treatment. a, b, c significant differences at p < 0.05 level.
The Chao1 index reflects species richness within the microbial community, while the Shannon index incorporates information on both richness and evenness. Analysis revealed that all additive treatments (Lp, CE, and CELp) exhibited significantly lower Chao1 and Shannon indices compared to the CK treatment (p < 0.05). This decline in diversity primarily stems from the inhibitory effect of bacteriocins produced by LAB on other bacterial taxa, leading to a simplification of the species composition within the community (Soltani et al. Reference Soltani, Biron and Said2022). Upset plots and PCoA were employed to examine the compositional variations in bacterial communities across silage samples subjected to distinct experimental treatments. The Upset plot results indicated that the FM, CK treatment, Lp treatment, CE treatment, and CELp co-treatment contained 498, 121, 178, 245, and 47 unique operational taxonomic units (OTUs), respectively, with 121 OTUs shared across all treatments (Fig. 1A). PCoA based on Bray–Curtis distances revealed clear separation among samples from different treatments in the coordinate space, indicating significant treatment effects, along with high aggregation of samples within each treatments, demonstrating good reproducibility (Fig. 1B). Beta-diversity analysis revealed statistically significant differences (p < 0.05) in bacterial communities among treatment, confirming that silage intervention treatments serve as crucial determinants in reshaping the microbial architecture of rapeseed straw silage.
Effects of different additives on the microbial community structure of rapeseed straw. Panel (A) presents Venn and Upset plots, highlighting the common and unique operational taxonomic units (OTUs) between fresh material and silage. Panel (B) displays the principal coordinate analysis (PCoA) based on Bray–Curtis distances, revealing compositional differences among treatments. The relative abundances of microbial taxa are shown at the phylum level (C) and bacterial genus level (D), demonstrating shifts in community structure. Finally, panel (E) employs the LEfSe tool to identify statistically significant microbial biomarkers in mixed silage.

Figure 1 Long description
Panel A shows Venn and Upset plots illustrating common and unique Operational Taxonomic Units (OTUs) among treatments: FM, Control, Lp, CE and CELp. The Venn diagram highlights shared and unique OTUs, while the Upset plot quantifies these differences. Panel B presents a Principal Coordinate Analysis (PCoA) based on Bray-Curtis distances, showing compositional differences among treatments with clear separation and high reproducibility. Panel C displays relative abundances of microbial taxa at the phylum level across treatments, with different colors representing various phyla such as Firmicutes and Proteobacteria. Panel D shows relative abundances at the bacterial genus level, highlighting shifts in community structure with genera like Lactobacillus and Acetobacter. Panel E uses the LEfSe tool to identify statistically significant microbial biomarkers in mixed silage, with LDA scores indicating the impact of different treatments on microbial composition. Each panel provides insights into the microbial community structure and the effects of various additives on rapeseed straw silage.
Taxonomic profiling of the bacterial communities in rapeseed straw silage was conducted at the phylum and genus levels. As illustrated in Fig. 1C, Cyanobacteria and Proteobacteria were identified as the predominant phyla within the untreated FM. After the fermentation process, the relative abundance of Firmicutes increased significantly, becoming the predominant phylum in all silage samples. It is noteworthy that LAB are taxonomically classified under the phylum Firmicutes, which includes several fermentation-related genera such as Lactobacillus, Enterococcus, and Lactococcus. In this experiment, all exogenous additive treatments (Lp, CE, and CELp) exhibited significantly higher abundance of Firmicutes compared to the untreated CK treatment.
This dynamic shift in microbial community structure reflects the typical successional pattern observed during ensiling fermentation. As the fermentation environment transitions from aerobic to anaerobic, the dominant phylum of the microbial community typically shifts from Proteobacteria to Firmicutes. This successional process is primarily driven by two factors: first, the selective pressure exerted by the anaerobic and acidic environment on specific microbial taxa; second, the unique metabolic traits of Firmicutes, such as their ability to produce acids under anaerobic conditions and to secrete various enzymes with specific functions (Wang et al. Reference Wang, Zhang and Liu2023).
(Fig. 1D) illustrates the genus-level community composition of the FM and various silage treatments after 60 days of fermentation. Analysis revealed that Pseudomonas was the dominant genus in the FM. These aerobic microorganisms proliferate rapidly during the initial aerobic phase of raw material storage, consuming substantial amounts of oxygen and soluble sugars, which can lead to increased temperature and accelerated spoilage. Following complete ensiling fermentation, Lactobacillus became the predominant genus across all silage samples. Compared with the CK treatment, all intervention treatments (Lp, CE, and CELp) demonstrated that the increased relative abundance of Lactobacillus in the Lp treatment displayed the most substantial colonization by the Lactobacillus genus. This succession phenomenon stems from the metabolic differences among bacterial genera. Lactobacillus efficiently utilizes soluble sugars in the substrate, rapidly generating and accumulating LA through the homolactic fermentation pathway, which induces a swift decline in system pH and effectively suppresses the growth competition from other microorganisms (Zhao et al. Reference Zhao, Sun and Chang2024). The relative abundance of Lactobacillus in the CELp co-treatment was higher than that in the CE treatment. Meanwhile, a higher abundance of Xanthomonas was observed in both additive treatments, and LDA effect size (LEfSe) analysis indicated a significant enrichment of this bacterium in the CELp co-treatment (Fig. 1E). Previous studies have reported a significant negative correlation between the abundance of Xanthomonas and ADF content in barley silage systems, suggesting that this bacterium may acquire nutrients for growth by decomposing the cellulose and hemicellulose components of plant cell walls (Sun et al. Reference Sun, Na and Li2021). Particularly noteworthy is the significant proliferation of Lactobacillus observed in the CELp co-treatment. This is likely attributable to the synergistic effect between the bacterium and enzyme, which promoted further cellulose degradation and consequently supplied a more abundant pool of fermentable sugar substrates for beneficial microbial communities represented by Lactobacillus.
Interestingly, the LEfSe analysis identified Xanthomonas as a significantly enriched genus in the CELp co-treatment, displaying a strong negative correlation with ADF and NDF contents (Yu et al. Reference Yu, Yan and Wang2024). Literature suggests that Xanthomonas species possess a sophisticated array of carbohydrate-active enzymes, particularly CEs and hemicellulases, facilitated by TonB-dependent receptors for efficient polysaccharide scavenging (Blanvillain et al. Reference Blanvillain, Meyer and Boulanger2007). Furthermore, acting as a primary degrader, Xanthomonas dismantles the recalcitrant lignocellulosic matrix into fermentable oligosaccharides, which subsequently serve as essential substrates to support the rapid proliferation and LA production of Lactobacillus (Chukwuma et al. Reference Chukwuma, Rafatullah and Tajarudin2021). Regarding biological safety, the rapid establishment of anaerobic conditions and the decline in pH below 4.5 effectively inhibit the expression of virulence factors and the survival of aerobic Xanthomonas strains, thereby neutralizing their potential pathogenic risk (Dey and Raghuwanshi Reference Dey and Raghuwanshi2024; Shah et al. Reference Shah, Guo and Adnan2026).
Functional prediction of environmental adaptation traits was performed through comparative analysis with the KEGG database
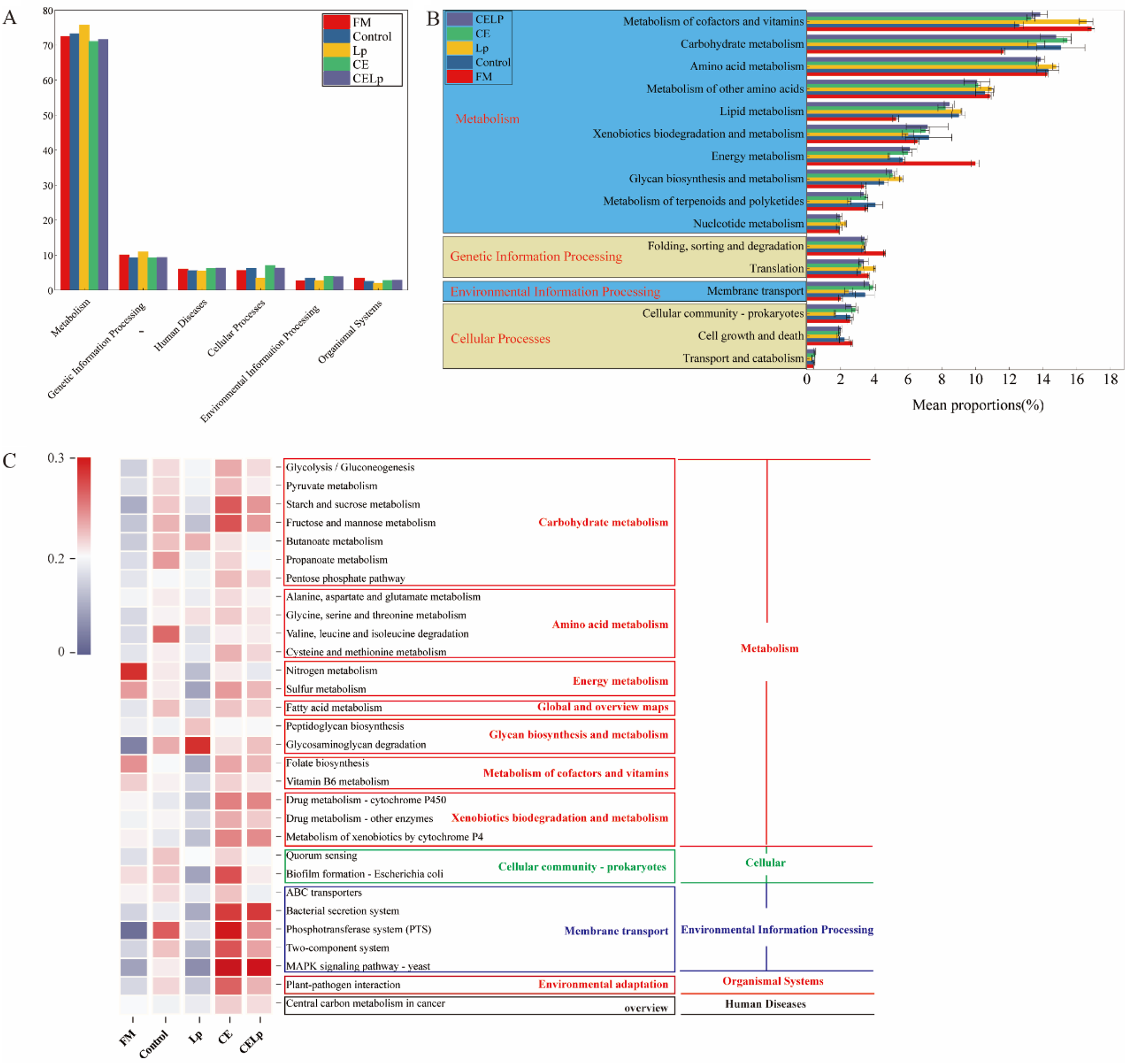
Deciphering the functional traits and metabolic potential of the silage microbiome offers critical insights into the fermentation process. In this study, based on the KEGG database, the author systematically analyzed the predicted functions of bacterial communities across levels 1–3 using the PICRUSt tool (Fig. 2). At the first level of pathway classification, functions related to “Metabolism” were predominant, confirming that during ensiling fermentation, bacteria actively utilize various fermentable substrates for metabolic transformations, thereby forming a diverse network of metabolic pathways. Overall, ensiling fermentation is essentially a complex biochemical process driven by microorganisms, which achieve the decomposition of substrates and remodeling of metabolites through their unique metabolic networks. This study confirms that the exogenous addition of L. plantarum and CE can effectively enhance the carbohydrate metabolism function of the silage system, a result consistent with the earlier findings. The underlying reaction involves L. plantarum rapidly lowering the system pH through the production of organic acids, thereby inhibiting the growth of harmful microorganisms; simultaneously, CE directly acts on the structural polysaccharides of plant cell walls, promoting their depolymerization. This synergistic interaction between the bacterium and enzyme supplies more abundant available substrates to functional fermenting microorganisms, thereby activating and strengthening the relevant metabolic pathways (Li et al. Reference Li, Guo and Zhao2021a; Si et al. Reference Si, Wang and Liu2023). During the ensiling fermentation process, several pathways associated with energy metabolism, including nitrogen metabolism, sulfur metabolism, and the metabolism of cofactors and vitamins, were suppressed to varying degrees. This observation is consistent with the previous report (Wang et al. Reference Wang, Li and Zhao2022). The occurrence of the aforementioned phenomenon may be related to the high cellulose and lignin content in ensiled materials such as alfalfa, which are chemically similar to rapeseed straw. The complex fibrous structure of these materials may impose certain limitations on bacterial metabolic activity. At the same time, the results of this study diverge significantly from previous reports that demonstrated a significant enrichment of energy metabolism pathways during corn ensiling (Xu et al. Reference Xu, Wang and Rinne2021), whereas the current research indicates that the exceptionally high-fiber content of rapeseed straw constitutes a critical limiting factor. Its complex physical structure obstructs the activation of core metabolic pathways, such as energy and cofactor metabolism, thereby diminishing overall microbial metabolic activity and potentially exerting a negative impact on silage fermentation quality.
PICRUSt analysis of dynamic changes in bacterial functional profiles under different treatments (n = 3). (A) Level 1 metabolic pathways. (B) KEGG ortholog functional predictions at level 2, showing the relative abundance of the top 16 significantly differential metabolic functions. (C) KEGG functional predictions at level 3, displaying the relative abundance of the top 30 metabolic functions. FM, fresh materials; Control, untreated sample; Lp, rapeseed straw inoculated with Lactiplantibacillus plantarum; CE, rapeseed straw inoculated with cellulase; CELp, rapeseed straw co-inoculated with Lactiplantibacillus plantarum and cellulase.

Figure 2 Long description
The image contains three distinct graphs labeled A, B and C, each representing different levels of bacterial functional profiles under various treatments (Figure 2). A bar graph labeled A shows Level 1 metabolic pathways with categories such as metabolism, genetic information processing, environmental information processing and cellular processes. The x-axis is labeled with these categories and the y-axis shows the count. Different treatments are represented by color-coded bars: FM, Control, Lp, CE and CELp. Graph B is a horizontal bar graph showing KEGG ortholog functional predictions at Level 2, with the x-axis labeled 'Mean proportion (percent)' and categories such as metabolism of cofactors and vitamins, lipid metabolism and cellular processes. Each category has multiple bars representing different treatments. Graph C is a heatmap showing KEGG functional predictions at Level 3, displaying the relative abundance of the top 30 metabolic functions. The heatmap includes categories like carbohydrate metabolism, amino acid metabolism and cellular community prokaryotes, with color intensity indicating relative abundance. Treatments FM, Control, Lp, CE and CELp are shown on the x-axis.
Prediction of enzymatic functions in microbial communities based on the KEGG database
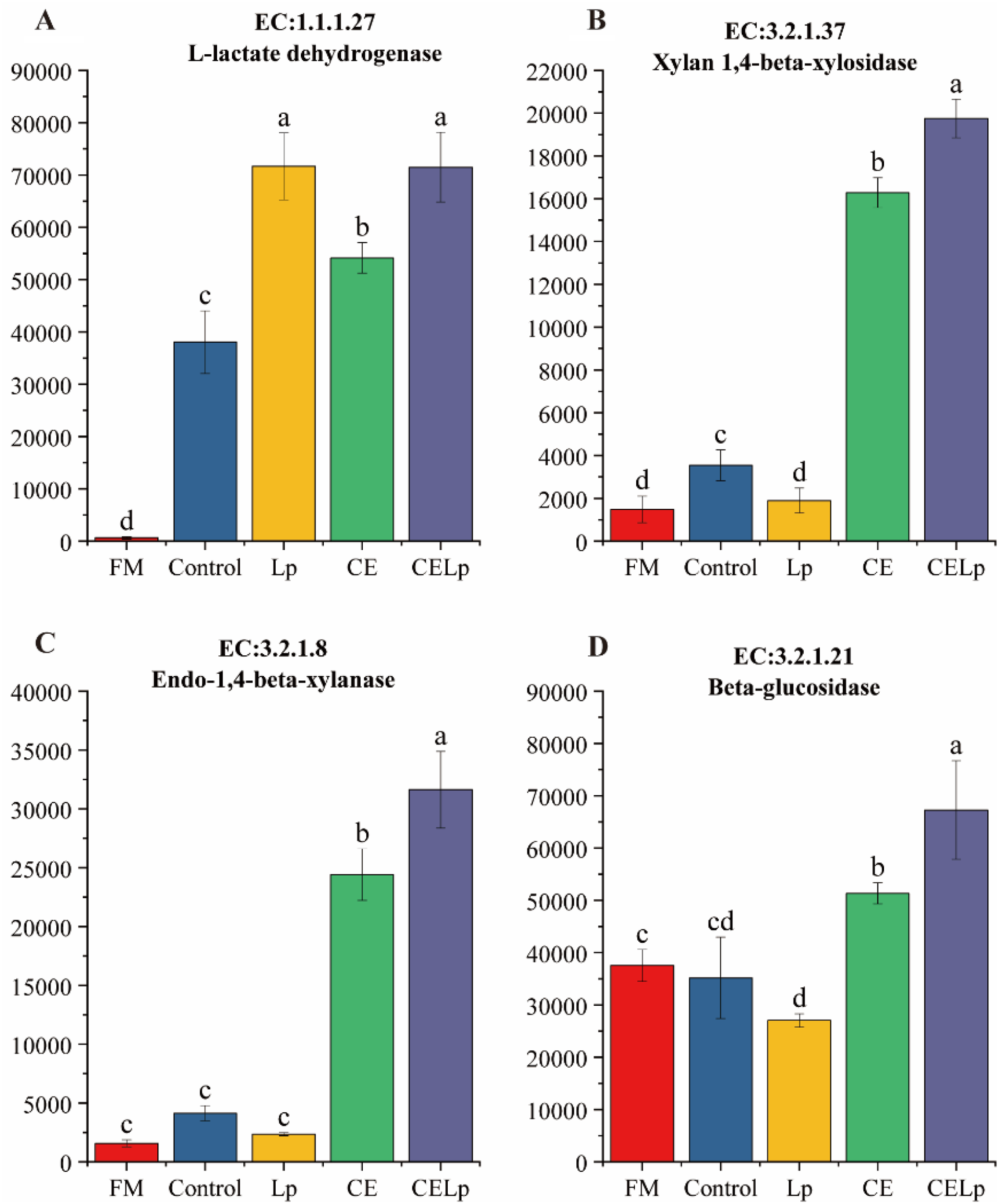
L-lactate dehydrogenase (EC: 1.1.1.27) is a key enzyme in the glycolysis and gluconeogenesis pathways (Fig. 3A). Its core biochemical function is to catalyze the reduction of pyruvate to L-lactate while oxidizing the coenzyme NADH to NAD⁺. In this study, the expression levels of L-lactate dehydrogenase were significantly higher in all additive treatments (Lp, CE, and CELp) compared to the CK treatment. Notably, the enzyme expression in the CELp co-treatment was significantly higher than that in the CE treatment. This result demonstrated that compared to single enzyme treatment, the synergistic action of bacteria and enzymes can more efficiently drive the conversion of pyruvate to L-lactate, thereby accelerating system acidification.
Changes in key enzymes of the cellulolytic bacterial community during silage fermentation of rapeseed straw with additives. (A) L-lactate dehydrogenase. (B) Xylan 1,4-beta-xylosidase. (C) Endo-1,4-beta-xylanase. (D) Beta-glucosidase. The error bars in the bar chart represent standard deviation (SD). There are significant differences (p < 0.05) in the average values between different letters.

Figure 3 Long description
The image contains four bar charts labeled A, B, C and D, each representing enzyme activity levels across different treatments. Chart A shows L-lactate dehydrogenase activity with treatments FM, Control, Lp, CE and CELp. The highest activity is observed in CELp, followed by Lp, CE, Control and FM. Chart B displays Xylan 1,4-beta-xylosidase activity, with CELp showing the highest activity, followed by CE, Lp, Control and FM. Chart C illustrates Endo-1,4-beta-xylanase activity, where CELp has the highest activity, followed by CE, Lp, Control and FM. Chart D shows Beta-glucosidase activity, with CELp having the highest activity, followed by CE, Control, Lp and FM. Error bars represent standard deviation and significant differences are indicated by different letters above the bars.
Xylan 1,4-beta-xylosidase functions as a pivotal hydrolytic catalyst within the metabolic pathways of amino sugars and nucleotide sugars. Its primary function is to hydrolyze 1,4-beta-xylan, releasing xylose monomers (Fig. 3B). Xylose is a core component of the hemicellulose backbone in plant cell walls, making its release efficiency a crucial indicator for assessing cell wall degradation. The experimental results showed that the xylan 1,4-beta-xylosidase activity in the combined bacterium–enzyme treatments was significantly higher than that in either the bacterium- or enzyme-alone treatments. This indicates that the synergistic interaction between microorganisms and enzymes can more effectively disrupt plant cell wall structure, promote the release of intracellular nutrients, and thereby create a more favorable microenvironment for the ensiling fermentation process.
Endo-1,4-beta-xylanase is a key enzyme that catalyzes the hydrolysis of internal β-1,4-xylosidic bonds within xylan molecules (Fig. 3C). Experimental data showed that the expression level of this enzyme in the combined bacterium–enzyme treatments was significantly higher than in either the bacterium- or enzyme-alone treatments. This reveals a synergistic and promotive relationship between specific functional bacterial communities and xylanase activity within the silage system. This synergistic effect enhances the overall hydrolysis efficiency of xylosidic bonds by significantly upregulating the expression of endo-1,4-beta-xylanase.
β-Glucosidase exhibits bifunctional activity in the cellulolytic process: initially catalyzing the hydrolysis of cellodextrins to cellobiose, followed by the cleavage of cellobiose into D-glucose (Fig. 3D). This final step is essential for the full depolymerization of cellulose into bioavailable monosaccharides for microbial assimilation. This study demonstrated that the β-glucosidase expression level in the combined bacterium–enzyme treatments was significantly higher than in either the bacterium- or enzyme-alone treatments. This indicates that the synergistic interaction between CE and L. plantarum significantly enhances cellulose degradation efficiency, outperforming the supplementation of the bacterial inoculant alone. Such synergy is expected to more efficiently promote the overall fermentation process while supplying richer glucose as fermentative substrates for beneficial microorganisms such as LAB.
Analysis of silage quality, fiber characteristics, and their correlations with bacterial and enzymatic profiles
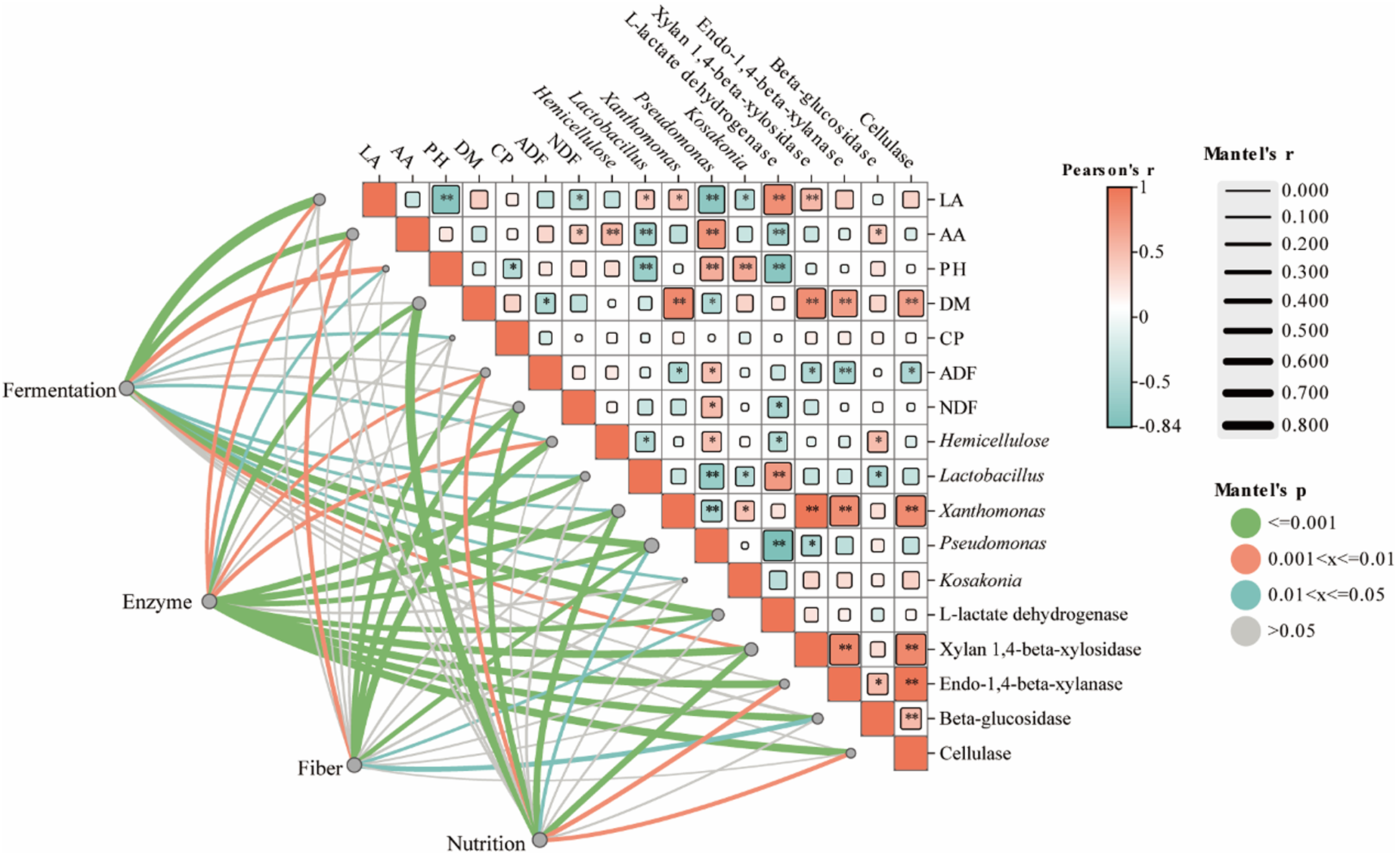
To systematically elucidate the relationships among fiber components, key enzyme activities, microbial community structure, and silage quality parameters, this study performed a correlation network analysis using the Mantel test (Fig. 4). The results indicated that the relative abundance of Lactobacillus showed a significant positive correlation with LA concentration, while exhibiting significant negative correlations with system pH and AA content. As a typical homofermentative LAB, Lactobacillus directly utilizes water-soluble sugars from the raw material to produce LA via the glycolytic pathway. The rapid accumulation of LA is the primary driver for the swift decline in pH within the silage system (Zhu et al. Reference Zhu, Zhao and Yan2025).
Correlation analysis of nutritional components, fermentation parameters, key enzymes, fiber parameters, and bacteria in rapeseed straw silage. Red indicates positive correlations, and green indicates negative correlations. Asterisks denote statistical significance: *p < 0.05 and **p < 0.01. The square size represents the magnitude of correlation coefficients, while color intensity reflects the strength of the correlation.

Figure 4 Long description
The image presents a correlation analysis involving nutritional components, fermentation parameters, key enzymes, fiber parameters and bacteria in rapeseed straw silage (Figure 4). The top section features a matrix with squares representing correlation coefficients between various elements such as LA, AA, DM, CP, ADF, NDF and different bacteria like Lactobacillus and Acetobacter. Positive correlations are indicated in red, while negative correlations are shown in green. Asterisks denote statistical significance, with indicating less than 0.05 and * indicating less than 0.01. The square size reflects the magnitude of correlation coefficients and color intensity indicates the strength of the correlation. Below the matrix, lines connect different categories: Fermentation, Enzyme, Fiber and Nutrition, illustrating the relationships between these components. The lines are colored to represent the correlation strength, with red for positive and green for negative correlations.
ADF primarily consists of cellulose and lignin that are difficult for animals to digest and utilize, making it a key negative indicator for evaluating feed nutritional value. Correlation analysis revealed that ADF content showed significant negative correlations with CE activity, endo-1,4-beta-xylanase activity, xylan 1,4-beta-xylosidase activity, and the abundance of Xanthomonas. Furthermore, both the NDF and hemicellulose content exhibited significant negative correlations with L-lactate dehydrogenase activity, while hemicellulose content was also negatively correlated with the abundance of Lactobacillus. Although only CE was exogenously supplemented, its initial disruption of the fiber structure created accessible sites for attack by endogenous enzymes, such as Endo-1,4-beta-xylanase and Xylan 1,4-beta-xylosidase, which are either inherently present in the forage or secreted by microorganisms like Xanthomonas. These enzymes acted synergistically to collectively degrade the cellulose and hemicellulose that constitute the plant cell wall. The enhanced fermentation efficiency was observed in the CELp co-treatment and driven by a bidirectional bacteria–enzyme synergistic effect rather than a simple additive effect (Du et al. Reference Du, Xu and Jiang2025). First, the exogenous CE initiates the depolymerization of the recalcitrant plant cell wall, breaking down structural carbohydrates (such as ADF and NDF) into fermentable monosaccharides and oligosaccharides (Li et al. Reference Li, Yuan and Dong2018). This release of accessible carbon sources alleviates the substrate limitation typically encountered in high-fiber rapeseed straw, thereby directly promoting the rapid colonization and exponential proliferation of bacteria such as Lactobacillus. Second, the modulation of microbial diversity subsequently upregulates the endogenous enzymatic profile of the silage system. As evidenced by our functional prediction data (Fig. 3), the combined CELp co-treatment significantly enriched endogenous hydrolytic enzymes – specifically xylan 1,4-beta-xylosidase, endo-1,4-beta-xylanase, and beta-glucosidase – reaching expression levels significantly higher than those in either the CE or Lp individual treatments. This indicates that the CELp treatment induces the secretion of more endogenous hydrolytic enzymes to further dismantle the lignocellulosic matrix. Ultimately, this dual reaction ensures continuous fiber degradation and sustained homolactic fermentation, resulting in superior silage quality.
In conclusion, this study demonstrates that the combined application of L. plantarum and CE generates synergistic effects that significantly enhance the fermentation quality and nutritional value of rapeseed straw silage. The co-treatment promoted efficient LA production, accelerated acidification, and effectively degraded structural fibers (ADF, NDF, and hemicellulose), thereby improving substrate availability for microbial metabolism. Microbial community analysis revealed substantial enrichment of Lactobacillus and key functional enzymes, including L-lactate dehydrogenase, xylanases, and β-glucosidase, in the CELp co-treatment, which collectively enhanced carbohydrate metabolism and fiber decomposition. Correlation analyses confirmed robust interrelationships among critical silage parameters, microbial taxa, and enzymatic activities, underscoring the central role of microbial–enzyme synergism in optimizing the ensiling process. These findings establish inferred metabolic profiles as a foundation for developing tailored microbial–enzyme co-treatment strategies to upgrade lignocellulosic biomass into high-quality ruminant feed.
Conclusion
This study demonstrates that the CELp co-treatment significantly enhances the fermentation quality of rapeseed straw silage. Results from 16S rRNA sequencing revealed a marked enrichment of functional microorganisms, such as Lactobacillus and Xanthomonas. Furthermore, functional predictions indicated a significant upregulation of metabolic pathways related to homofermentative LA fermentation and fiber degradation, which was further supported by the predicted enhancement of key enzymes involved in fiber breakdown. However, as this study is primarily based on 16S rRNA sequencing, the observed improvements in functions and enzyme activities remain at the level of bioinformatic prediction. Thus, these findings provide preliminary scientific evidence and insights into the manner in which bacterial–enzymatic synergy improves silage quality.
Data availability statement
Sequence data that support the findings of this study have been deposited in the NCBI’s Sequence Read Archive under Bio-Project accession number PRJNA1297899.
Acknowledgements
This research was supported by the Research and Development of Collaborative Innovation Technology for the Whole-Chain Microbial Fermentation of Rapeseed Straw and Its Integrated Application Demonstration (2025KJHZ0041), the Research and Application Demonstration on Precision Nutrition Formulation of Straw-Based Total Mixed Rations and Green and Low-Carbon Livestock Production Technology (2025KJHZ0046), and the Inner Mongolia Autonomous Region Agricultural and Animal Science Technology Transfer and Transformation Fund (2024TG09) and Doctoral Fellowship (2023BSJJ01).
Author contributions
Y.X.: Writing – original draft, methodology, formal analysis, and visualization. Lichao He: Writing – review and editing, validation, and data curation. Z.L.: Methodology, software, and investigation. L.S.: Conceptualization, resources, and supervision. T.W.: Formal analysis, visualization, and writing – review and editing. P.S., W.M., and M.L.: Investigation, data curation, and validation. S.D.: Conceptualization, supervision, and funding acquisition.
Conflict(s) of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.

 Open access
Open access