Introduction
In avian species with biparental care, a pair’s reproductive success, defined as the number of nestlings fledged per breeding season, often hinges on investments made by both parents (Beissinger et al. Reference Beissinger, Wunderle, Meyers, Saether and Engen2008; Snyder et al. Reference Snyder, Wiley and Kepler1987; White et al. Reference White, Collazo, Dinsmore, Llerandi-Román and Devore2014; Wilson et al. Reference Wilson, Field and Wilson1995, Reference Wilson, Wilson and Field1997; Wright et al. Reference Wright, Toft, Enkerlin-Hoeflich, Gonzalez-Elizondo, Albornoz and Rodríguez-Ferraro2001). There is much variation among species regarding how males participate in offspring care and how much male participation is affected by paternity uncertainty (Cockburn Reference Cockburn2006; Ketterson and Nolan Reference Ketterson and Nolan1994; Møller Reference Møller2000; Møller and Birkhead Reference Møller and Birkhead1993). In some species, males take turns with the female for egg incubation (Cockburn Reference Cockburn2006), while in others, both may defend the nesting territory (Cockburn Reference Cockburn2006). There are other systems in which males and females perform different roles in rearing offspring. In many parrots, such as the Green-rumped Parrotlet Forpus passerinus, females spend close to 80% of the daylight period inside the nest during the incubation period while the males are outside foraging for food to provide the female and eventually the nestlings (Waltman and Beissinger Reference Waltman and Beissinger1992). Once nestlings hatch, the feeding rate depends on brood size, such that the more nestlings at the nest, the higher the delivery rates (Waltman and Beissinger Reference Waltman and Beissinger1992). In the Green-rumped Parrotlet, males preferentially feed older nestlings and often do not feed younger nestlings. In comparison, females feed younger nestlings more frequently, especially in larger broods, potentially because females spend more time inside the nest than males and are more responsive to the higher begging frequency of younger nestlings (Budden and Beissinger Reference Budden and Beissinger2009).
For avian species in which males provide food to nestlings and females, deficiencies in food provisioning can be detrimental to reproductive success (Bart and Tornes Reference Bart and Tornes1989; Waltman and Beissinger Reference Waltman and Beissinger1992). Studies of male removal in Northern House Wren Troglodytes aedon populations have shown that the absence of the male may lead to decreased offspring survival, and male parental care was especially critical during periods of poor resource availability (Bart and Tornes Reference Bart and Tornes1989). In species with biparental care and asynchronous hatching, such as most parrot species, the failure of one parent to provide food can significantly reduce reproductive success. For example, mortality among wild Green-rumped Parrotlet nestlings is highest within 12 days of hatching, with later hatched nestlings frequently found with empty crops (Stoleson and Beissinger Reference Stoleson and Beissinger1997). In these cases, mortality was primarily due to inequitable food distribution rather than a lack of food available to the parents (Stoleson and Beissinger Reference Stoleson and Beissinger1997; Vigo-Trauco et al. Reference Vigo-Trauco, Martínez-Sovero and Brightsmith2024). Characterising differences in food provisioning and other parental care strategies between captive and wild or reintroduced populations can provide helpful information to managers of threatened species maintained in captive breeding programmes.
Captivity may also affect parental care performance due to the absence of natural stimuli that regulate the initiation of behaviours crucial for reproductive success, such as foraging (Gilby et al. Reference Gilby, Mainwaring, Rollins and Griffith2011; Mason Reference Mason2010). One characteristic of captivity affecting food provisioning by breeding pairs is the continuous availability of food. In captive Zebra Finch Taeniopygia guttata, parents visited their nests more frequently and provided twice the amount of food compared with their free-living counterparts despite similar brood sizes (2–5 nestlings per nest; Gilby et al. Reference Gilby, Mainwaring, Rollins and Griffith2011). However, food distribution was less equitable among captive parents with larger broods, with greater variation observed in the number of regurgitations per nestling in larger broods (Gilby et al. Reference Gilby, Mainwaring, Rollins and Griffith2011). In captive Budgerigars Melopsittacus undulatus, females feed smaller nestlings independently of their age, whereas males tend to initiate feeding when nestlings beg more (Stamps et al. Reference Stamps, Clark, Arrowood and Kus1985). In another species of parrots in the wild, Crimson Rosella Platycercus elegans, males deliver higher feeding rates than females and tend to feed the first hatched nestlings more frequently. In contrast, the females distribute food more evenly (Krebs et al. Reference Krebs, Cunningham and Donnelly1999). However, it remains unclear whether food availability in captivity similarly influences food provisioning behaviour across other species, as responses to captivity vary widely.
The Puerto Rican Parrot Amazona vittata is a species in which variation in parental care may relate to reproductive success, which can impact conservation management strategies. Efforts to save the Puerto Rican Parrot have been on-going since the late 1960s when it was first listed as “Critically Endangered”, a status that it still holds over 60 years later (IUCN 2022). The captive breeding programme for Puerto Rican Parrots began in 1972, but achieving consistent reproductive success has proven challenging for both wild and captive populations. To investigate factors contributing to low reproductive success, a study was conducted from 1987 to 1990 to assess the nesting attentiveness of wild Puerto Rican Parrots in the El Yunque wild population (Wilson et al. Reference Wilson, Field and Wilson1995, Reference Wilson, Wilson and Field1997). In this study, it was observed that female nest attendance decreased as the nestlings grew older, while male nest attendance increased once the chicks hatched, although males did enter the nest sporadically during early chick development. The study also revealed that female attentiveness to the nest was crucial for the success or failure of the nest, with longer breaks and less time spent inside the nest predicting greater nest failure during incubation and early nestling-rearing periods (e.g. up to day 20 after the first nestling hatched; Wilson et al. Reference Wilson, Field and Wilson1995, Reference Wilson, Wilson and Field1997). Since that study, a second population has been established at Rio Abajo Commonwealth Forest, first as a captive population on 30 April 1993 when the first group of 10 Puerto Rican Parrots arrived, hereafter called the Rio Abajo Captive population, and then as a reintroduced population on 19 November 2006, hereafter called the Rio Abajo Wild population.
Notably reproductive success differs between the two populations, with a mean annual reproductive success from 2015 to 2018 of 1.02 fledglings per pair in the Rio Abajo Captive population (SD = 0.133, range = 0.298) and 1.29 fledglings per pair in the Rio Abajo Wild population (SD = 0.179, range = 0.379; unpublished data Puerto Rican Parrot Recovery Program). Since the mid-2000s, the Puerto Rican Parrot Recovery Program has used cameras to monitor nest activity, allowing researchers to access previously unseen behaviours. However, it is still unknown how each member’s contribution during feeding visits affects the species’ reproductive success and how captive conditions and parental sex influence parental care behaviours. A deeper understanding of parental care patterns could enhance efforts to increase the number of released individuals to augment reintroduced and wild populations.
In this study, we examined reproduction in the Rio Abajo Captive and Rio Abajo Wild populations of the Puerto Rican Parrot to understand how captive conditions and sex relate to parental care behaviours, how parental care behaviours impact reproductive success, and how they might differ in captive vs wild populations. We monitored the nest activity of wild and captive populations using video recordings of daytime activity. We measured parental food allocation (number of nestlings fed, total bouts in a visit, and mean bout duration) and time spent inside the nest cavity. We hypothesised that parental care strategies would impact the number of nestlings that fledged from a nest. We expected a positive relationship between parental care efforts and the number of fledglings produced. We also hypothesised that males and females differ in their parental care performance. We expected females to exhibit more extended visits and more feeding bouts per visit than males. Since captivity is documented to affect physiology and behaviours (Cabezas et al. Reference Cabezas, Carrete, Tella, Marchant and Bortolotti2013; Mason Reference Mason2010; Ramos-Güivas et al. Reference Ramos-Güivas, Jawor and Wright2021), we hypothesised that there is a difference between captive and wild parental care. We expected to see more feeding bouts, longer duration of bouts, and more extended visits in the wild population.
Methods
Study site and species
We collected video recordings from the Rio Abajo Captive population (RA Captive) and Rio Abajo Wild population (RA Wild) at the Rio Abajo Commonwealth Forest in Puerto Rico (Rio Abajo; 18.2°N, 66.4°W; 366 m a.s.l.). The captive population was established in 1993 with 10 birds from a captive population held at El Yunque National Forest. The wild population was established in 2006 through annual releases of captive-bred individuals. By the time this study was conducted, the population comprised a mix of both previously released individuals and wild-born birds. The captive population houses around 220 individuals, with yearly fluctuations due to births, deaths, translocations, and releases. Each year, the captive population has at least 40–60 nesting pairs. The size of the wild population at the Rio Abajo Commonwealth Forest ranged from 74 to 134 individuals during the study period, from 2015 to 2018, with a general trend of population growth.
The Puerto Rican Parrot exhibits a monogamous mating system with biparental care. They use secondary cavities in trees for nesting. Artificial cavities are provided for captive and wild populations as part of the recovery programme and are equipped with cameras to observe nest activities. Only nests in artificial cavities were examined in our study. During incubation, which lasts approximately 26 days, the female is solely responsible for tending to the eggs. Clutches can range from one to four eggs but typically consist of three eggs laid between January and February (Snyder et al. Reference Snyder, Wiley and Kepler1987; Puerto Rican Parrot Recovery Program unpublished data). Hatching within broods is asynchronous, and fledging occurs between 55 and 75 days post-hatching. The timing of fledging varies among nestlings and across breeding pairs, reflecting individual and pair-specific differences in development and parental care (unpublished data, Puerto Rican Parrot Recovery Program). The male plays a crucial role in providing food to the female during incubation and the early nestling period, while the female is still brooding the nestlings, and the females use some of the food provided by the male to feed the nestlings. The male also provides food directly to the nestlings immediately after hatching. The female joins the male on extended foraging excursions once the nestlings are able to thermoregulate. After the foraging excursions begin, both males and females enter the nest and feed the nestlings (Snyder et al. Reference Snyder, Wiley and Kepler1987).
Daily management of the captive and wild populations included scheduled feeding and restricted human activity to minimise disturbance, particularly near breeding areas. For the captive population, nest checks were conducted primarily on Mondays and Fridays, involving direct inspections and occasional manipulation of eggs or chicks. Nests of the wild population were monitored using video when available to reduce human disturbance. Nest checks occurred throughout the week, based on nest conditions and parental behaviour, particularly after 09h00, to align with natural visitation patterns. These practices aimed to support breeding success while minimising disruptions to nesting pairs.
Video data collection
Video recordings were collected during the 2017–2018 breeding seasons of the RA Captive population and the 2015–2018 breeding seasons of the RA Wild population. We initially recorded 28 pairs, but due to equipment and nesting failure, we only obtained parental care data from 15 pairs (see Supplementary material). After our validation procedure (see below), we ended up with seven pairs from the RA Captive population and seven from the RA Wild population.
We used remote 24-hour video recording equipment to evaluate parental care behaviours during daylight in both populations. For RA Captive, we used a 16-channel DRV (1 Hikvision AcuSense iDS-7216HQH1-M2/S Turbo HD). The cost for the DVR was US$350.00. For RA Wild, we used a portable DVR (6 KJB Security Products DVR506 720p) and 16 channels DRV (1 Hikvision AcuSense iDS-7216HQH1-M2/S Turbo, 1 Hikvision 16 channel 720p DVR with 2TB HDD). The cost of the portable DVR was US$489.99. We used multiple models of cameras (SWPRO-1080FLD-US 2.1MP, Speco Technologies HT71TG 2MP, Swan 1080 ANLG.HD Dome Cam.3.6 mm) that were placed at least 30–45 cm in height from the bottom of the nest. The cost of the camera equipment ranged between US$79.99 and US$100.00 per unit, depending on the brand and model. The area captured in the video image showed a view from the top of the nesting material where the female and nestlings were located, and the bottom of the wire mesh that served as a ladder for the birds to enter and exit the nest. The video files were downloaded from the DVR into external hard drives (Seagate Expansion 4TB, Seagate Backup Plus 5TB, Seagate Expansion 5TB, and Seagate Backup Plus 6TB). The cost of each hard drive ranged between US$120.00 and US$150.00.
To measure parental food provisioning, we began video-scoring daylight periods after the first nestling hatched and continued for two weeks. Managers found that the first week is critical for nestling survival since survival after the first week is associated with a high probability of fledging (Puerto Rican Parrot Recovery Program unpublished data). The scored daylight period ran from 05h30 to 09h30 in the morning and from 15h30 to 19h30 in the afternoon. We selected these periods with the intention of capturing at least one visit from the male. Additionally, the programme staff observed that males from both captive and wild populations at Rio Abajo visit the nest throughout the day, especially early in the morning and late in the afternoon. During male visits, females typically leave the nest. This pattern allowed us to measure the time spent inside the nest for both sexes and the parental care behaviours for each sex. With the food provided to the females, the females provide food to the nestlings throughout the day and night. Captive populations were fed at 06h30, and the wild birds were provided with supplemental feeding at 07h00. Staff activities were then reduced to a minimum until 09h00, at which point facility maintenance and other activities began. For both populations, we collected video images for at least four days over two weeks, with 84% of the total observations taken on Tuesdays and Thursdays, 15% on Mondays and Wednesdays, and 1.4% on Fridays and Saturdays. This schedule was followed to minimise the effects of nest checks and other disruptive maintenance activities on the captive population, which occurred on Mondays and Fridays and could cause parents to leave their nests. For sex identification, we followed each individual once they entered the nest and collected their data until they exited the nest. A total of 480 hours of video recording were scored, with 32 total hours per pair.
Parental care behaviour scoring
The data from the video recordings were collected by trained volunteer scorers and one master scorer (BR-G). The trained volunteers were undergraduate students, professional biologists, and other professionals recruited through social media advertisements and word of mouth. The volunteer scorers were each provided with a written data collection protocol and a live video training session that included examples of measurement of each variable. After the video training session, each scorer was assigned some or all observation days for a single pair, depending on their availability. Their progress was monitored weekly by email, text messaging or video conferences, where questions and difficulties were discussed. Of the 47 initial volunteers, only 20 submitted their work, and only 17 completed it. Assignments for scorers who did not complete their work were reassigned to other volunteers or the master scorer.
Parental care behaviours were collected during a parental visit, defined as the first time a parent appears in camera view. The visit ended when the parent climbed the wire mesh ladder and was not seen on the camera for at least five seconds. We measured the following behaviours: the number of nestlings fed, the number of feeding bouts, the duration of each feeding bout, and the time each parent spent inside the nest (hereafter duration inside). The number of nestlings fed in a visit was the total number of nestlings a parent fed at least once. We defined a feeding bout as each event in which a parent feeds a nestling by locking its beak and regurgitating food while making quick head-bobbing movements. The feeding bout ends when both birds unlock their beaks and separate. We calculated the duration of each feeding bout. Then, for each adult visit, we calculated the mean duration of feeding bouts in our analysis. To measure the duration inside the nest, we started scoring when the adult bird appeared in the camera image, even while still hanging on to the wire mesh that served as a ladder to enter and exit the nest. The time inside ended when the individual bird was no longer visible in the image for at least five seconds.
Data validation
After data collection, we conducted a validation test for observer reliability, variable reliability, and pair source reliability. A total of 17 volunteer scorers and 15 breeding pairs were evaluated. For the validation, the master scorer scored at least five randomly selected visits for each scorer and all scored pairs. We conducted a Spearman’s rank correlation between the master scorer and each volunteer scorer. We considered a non-significant correlation between the volunteer scorer and the master scorer to represent a failed agreement. When a scorer’s or pair’s data failed to agree on two of the four factors, data for that pair were then re-collected by the master scorer unless the video quality was judged too poor for accurate data scoring (one pair; see below). We removed a variable from further analysis if it failed to agree with more than 50% of all scorers’ data.
Statistical approach
We first used Spearman’s rank correlation analyses to examine relations between parental care behaviours at the individual level and used the mean values for each individual over the two weeks.
To assess which factor affects parental care behaviour, we used the log+1 transformation to achieve normality and then used a mixed-model analysis with repeated measures for each individual. We fitted a separate mixed-model analysis for each parental care behaviour. The fixed effects for each analysis were the population (captive and wild), parental sex, mean nestling age at the time of the visit, number of nestlings in the nest at the time of the visit, pair duration (number of breeding seasons together), and a random effect of pair ID to control for repeated measures. We also included an interaction of sex and population in the mixed-model analysis.
We conducted a generalised linear model with Poisson distribution to analyse the relationship between the number of fledglings produced by each pair and parental care behaviours (total bouts, bout mean duration, and duration inside). Given the nature of the dependent variable (number of fledglings), a Poisson distribution was appropriate. We assessed overdispersion by comparing residual deviance to the degrees of freedom. For these models, we used each pair’s mean across the study period for each behaviour. We first conducted a generalised linear model with both populations in the same analysis. Then, we conducted a separate population analysis to reduce the possible effects of the population (captivity and wild). We also did a Wilcoxon test to compare fledgling production between the populations using the fledgling production of the studied pairs.
We examined the potential confounding effects of inbreeding and pair duration on the reproductive success metric of fledgling production. Inbreeding coefficient information came from the studbook database maintained using PopLink 2.52 software (Faust et al. Reference Faust, Bergstrom, Thompson and Bier2019). A generalised linear model with Poisson distribution was employed to assess associations with fledgling production.
We used JMP® Pro17 (SAS Institute Inc., Cary, NC, 2024) for analyses and R Studio team (2023).
Results
Validation
Our validation results showed that one pair failed the validation. Two scorers evaluated the pair RAW Chalet 2018, and both validations failed (see Supplementary material). The video quality of the recordings for this pair was poor, making scoring for both the volunteers and the main scorer difficult, and thus, we decided to remove this pair. Of the parental care factors measured, we found a low-scoring agreement for one variable, i.e. number of nestlings fed, from multiple scorers, where only 6 of 17 (35.3%) scorers passed the validation (see Supplementary material). We found that individual identification of each nestling in the nest (nestling ID) was difficult, especially in the earlier stages. For this reason, we removed the nestling ID variable from further analysis. After the validation results, the master scorer completed the data for three pairs and collected all the data from one pair.
Parental care behaviour correlations
After the validation, we retained three parental care behaviours used for our analysis: total bouts, mean bout duration, and duration inside the nest. Our results from Spearman’s rank correlations between the measured parental care behaviours showed that all three were correlated. The strongest correlation was between the mean bout duration and the total bouts (n = 28, r = 0.797, P <0.05; Figure 1a). The second strongest correlation was between duration inside and total bouts (n = 28, r = 0.686, P <0.001; Figure 1b). The correlation between the duration inside and the mean bout duration was the weakest (n = 28, r = 0.651, P < 0.05; Figure 1c). Although all three variables showed statistically significant positive correlations, we retained all three factors for further analysis, as each captures a different aspect of parental behaviour.
Spearman’s rank correlation of parental care behaviours, where each data point represents the mean value for an individual over the two-week study period. The open circle represents females and the closed circle represents males. (a) Correlation between total bouts in a visit and mean bout duration. (b) Correlation of total bouts in a visit and duration inside the nest. (c) Correlation of mean bout duration and duration inside. All correlations are significant at P <0.05.

Factors that influence parental care behaviours
In our linear mixed-model analysis, we first found that for total bouts, the random effect of pair ID accounts for about 8% of the variation (Table 1). Population, parental sex, and mean nestling age were shown to affect total bouts (Table 1). No other fixed effects significantly affected total bouts (Table 1). Both females and males from the captive population tended to have lower total bouts than their wild counterparts (Figure 2a).
Results of generalised linear-mixed models predicting parental care behaviours. Estimates of fixed and random effects associated with parental care behaviours. The values presented are results from log+1 transformed data. Total bouts n = 630, mean bout duration n = 630, duration inside n = 514

* Indicates results significant at alpha = 0.05.
Raw data from the mixed-model results for parental care behaviours and their significant effects. The analysis was performed with the log+1 transformed values for the dependent variable, but untransformed values are presented here for ease of interpretation. The dots represent the mean values for each sex and the SE for each mean. (a) The relationship of total bouts to population and sex, in which population and sex both showed a significant effect. (b) The relationship of mean bout duration to population and sex, in which population and sex both showed a significant effect. (c) The relationship of duration inside the nest to population and sex, in which only sex shows a significant effect.

Our results for mean bout duration showed that the random effect of pair ID accounted for approximately 6% of the variation (Table 1). Of the fixed effects, population and parental sex were shown to affect mean bout duration (Figure 2b). Females from the wild population tended to have a higher mean bout duration than captive females. Also, males from the wild population showed higher variation in this behaviour than captive males (Figure 2b).
For duration inside the nest, pair ID accounted for approximately 7% of the variation (Table 1). Among the fixed effects, only parental sex had a significant effect (Table 1). Males tended to spend less time inside the nest than females in both populations (Figure 2c).
Reproductive success and parental care behaviours
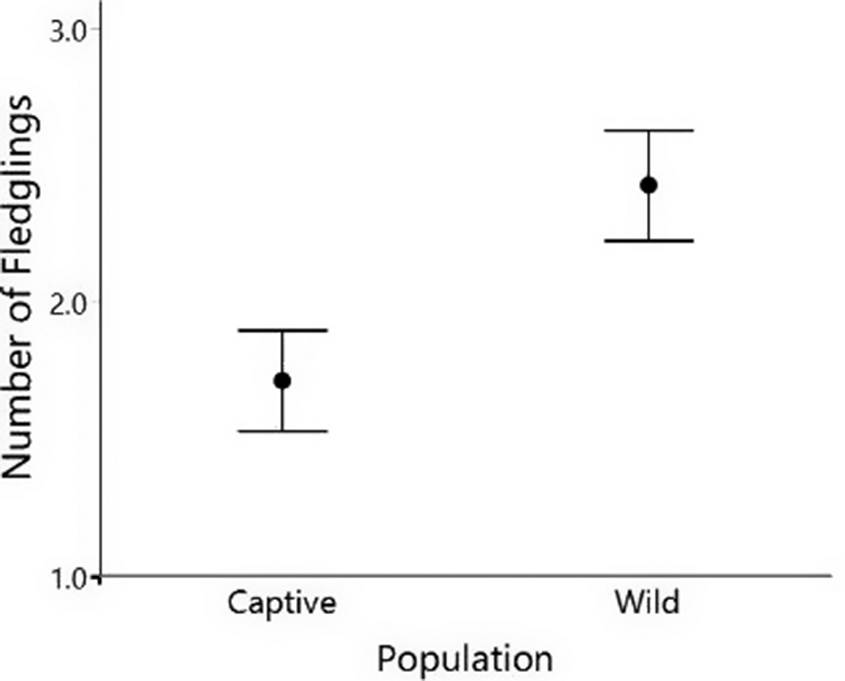
The 14 pairs in our study produced a mean of 2.07 fledglings with an SD of 0.616. The total number of fledglings produced by the wild population was higher than in the captive population for the pairs under the study (P = 0.04; RA Captive mean = 1.71, SD = 0.488; RA Wild mean = 2.42, SD = 0.534; Figure 3).
Comparison of fledgling production between the populations. Each dot represents the mean values for each population, and the range is the SE. Mean production was calculated using the fledglings produced by the 14 pairs in the study.

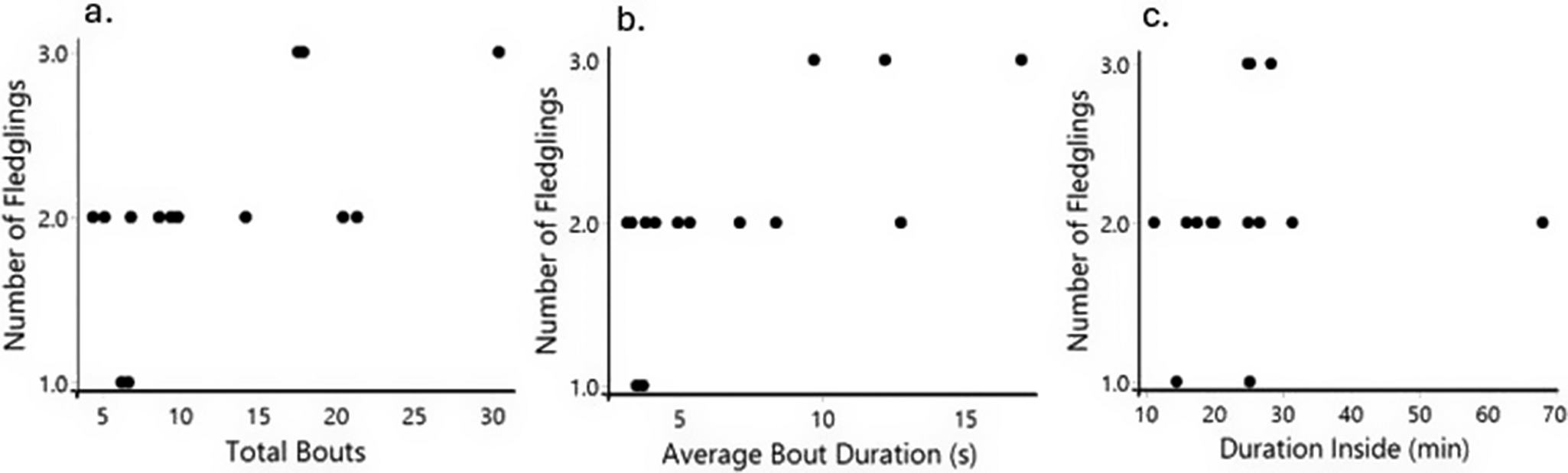
In our generalised linear model with Poisson distribution and incorporating both populations, we found that the mean bout duration (n = 14, SE = 0.017, P = 0.03) was significantly related to the number of fledglings produced (Figure 4). The number of fledglings increased with longer duration of feeding bouts during the first two weeks of nestling rearing (Figure 4). On the other hand, total bouts (n = 14, SE = 0.009, P = 0.14; Figure 4) and duration inside the nest (n = 14, SE = 0.005, P = 0.51; Figure 4) did not predict the number of fledglings produced by pairs.
Relationship between parental care behaviours and fledglings produced. A total of 14 pairs is represented, where each dot represents the mean for the first two weeks of each pair. (a) Total bouts did not have a statistically significant relationship with the number of fledglings produced. (b) Mean bout duration shows a statistically significant relationship with the number of fledglings. (c) Duration inside showed no statistically significant effect on fledglings produced.
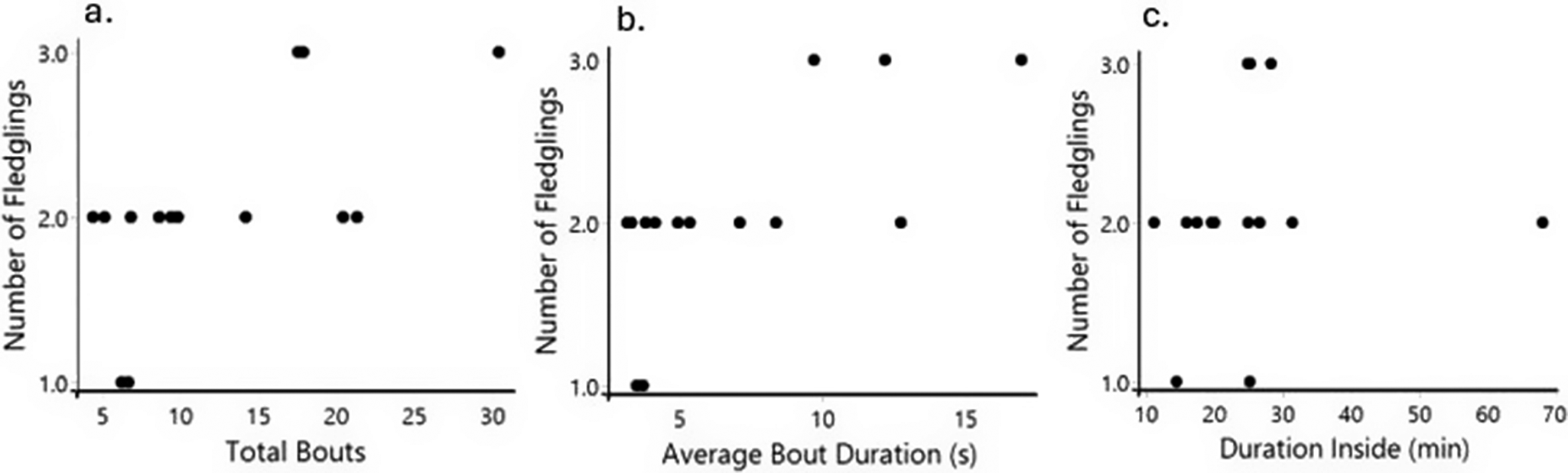
When we ran the analysis separated by population, with the caveat of smaller sample size, we found that none of the behaviours related to fledgling production in the captive population (n = 7, total bout, SE = 0.053, P = 0.93; bout mean duration, SE = 0.103, P = 0.59; duration inside, SE = 0.037, P = 0.92). However, for the wild population, bout mean duration (n = 7, SE = 0.015, P = 0.03) did relate to the number of fledglings but not the total bouts (n = 7, SE = 0.010, P = 0.37) and duration inside (n = 7, SE = 0.005, P = 0.34).
Analysis showed that reproductive success, as measured by the number of fledglings, was not contingent on parental inbreeding (n = 8, SE = 4.252, P = 0.81). For the inbreeding coefficient, due to individuals with incomplete pedigree records, inbreeding values could only be determined for eight breeding pairs (n =8). The mean for the pairs for which we had inbreeding values was 0.128, with a range of 0.078.
Discussion
Parental care behaviours can determine the success or failure of a breeding attempt in species with parental care. Here, we examined parental care behaviours and the number of nestlings that fledge in wild and captive populations of the Puerto Rican Parrot, a Critically Endangered species with biparental care. As we predicted, our results show that reproductive success is related to the parents’ mean bout duration when feeding the nestlings during a visit, showing a positive relationship with the number of fledglings produced by the pair. We also predicted associations between parental sex and population related to parental care behaviours. We found that parental sex and population (wild vs captive) affect parental care behaviours. In addition, we found that the nestling’s mean age at the visit is related to the total bouts in a visit. While we acknowledge that the number of nests examined was small, these results do demonstrate that during an early stage of nestling development, food provisioning strategies by the parents of a threatened parrot play an essential role in reproductive success and that the parent’s sex, captive conditions, and age of nestlings affect parental care behaviours.
Parental care strategies and fledgling production
We found that the mean feeding bout duration was related to the number of fledglings produced, such that longer feeding bouts were associated with a greater number of fledglings produced per breeding pair. A nest characterised by extended intervals of parent foraging excursions and insufficient food provisioning may lead to higher expenditure of energy by chicks when begging for food, potentially impacting their development (Royle et al. Reference Royle, Hartley and Parker2006) or survival (Stoleson and Beissinger Reference Stoleson and Beissinger1997). Starvation is a common cause of nestling mortality in species of birds with asynchronous hatching (Slagsvold and Wiebe Reference Slagsvold and Wiebe2007; Stoleson and Beissinger Reference Stoleson and Beissinger1997). Our study suggests that parents with longer feeding bouts per visit can increase the probability of providing enough food to all nestlings, potentially increasing their survival. Having more bouts during a visit was not related to the production of fledglings. More feeding bouts do not guarantee that the nestlings will be well-fed since feeding attempts can be interrupted by other individuals or a lack of coordination between parent and nestling.
In our study, it was not possible to identify each nestling individually. The camera’s distance to the chicks’ location was 30–45 cm above the bottom of the nest. For this reason, we had to use wide-angle cameras, which can cause image size disproportion. Additionally, because the image requires infrared light, it lacks coloration, making it challenging to identify colour marks. Finally, marking requires nest intervention, and in a captive population this can create a disturbance that can trigger a stress response that could potentially affect nearby nests. Thus, it was not possible to assess whether parents showed feeding preferences for specific chicks as has been found in other parrots (Stamps et al. Reference Stamps, Clark, Arrowood and Kus1985). However, we do know the age of each nestling at the nest, and our results show that the mean nestling’s age at the time of the visit was related to the total feeding bouts in a visit, with females having longer bouts as the nestlings got older. In captive Budgerigars, female parents prefer to feed younger individuals even without begging (Stamps et al. Reference Stamps, Clark, Arrowood and Kus1985). One thing to consider for future studies is to measure the amount of food provided. In overfed nestlings, the digesta retention time may decline, making digestion of food less efficient and compromising the nestling’s health (Karasov and Wright Reference Karasov, Wright, Wright and Leonard2002). Monitoring the weight of nestlings should help prevent nestling loss, specifically for Critically Endangered species programmes.
Although sibling competition is generally thought to be low in parrots, food competition between older nestlings could interrupt feeding bouts of other nestlings, reducing feeding bout duration as increased age results in greater strength and manoeuvrability (Krebs et al. Reference Krebs, Cunningham and Donnelly1999). Although we did not measure interruptions of feeding bouts by nestmates, this behaviour was observed on multiple occasions in multiple pairs and could be a form of nestling competition. However, in parrots, it has been hypothesised that parents allocate food using different strategies to reduce competition between nestlings (Krebs et al. Reference Krebs, Cunningham and Donnelly1999). Although we attempted to identify the fed nestlings based on size, our validation results suggested that this scoring was unreliable across observers, so we did not use these data in analyses. Using machine learning algorithms to identify individual chicks and catalogue parental feeding could mitigate this difficulty.
Effects of captivity and sex
We examined how captivity affects parental care behaviours and whether these behaviours have sex-related differences. Our results show that population affects the total number and mean duration of the bouts. Captive parents made fewer and shorter feeding bouts than their wild counterparts, a pattern that contrasts with that found in Zebra Finches, where the captive individuals fed more food to nestlings than the wild individuals (Gilby et al. Reference Gilby, Mainwaring, Rollins and Griffith2011). In addition, parental sex significantly affected the duration of nest visits. In our study species, and most species of parrots, the female incubates, and the male provides food to the female during the incubation period, and to both nestlings and females during early nestling development (Forshaw and Knight Reference Forshaw and Knight2010; Snyder et al. Reference Snyder, Wiley and Kepler1987; Spoon et al. Reference Spoon, Millam and Owings2006; Waltman and Beissinger Reference Waltman and Beissinger1992). When we measured parental behaviours during the first two weeks after hatching, males of both populations tended to spend less time inside the nest than females. We note that our analyses of differences between wild and captive populations were based on particularly small samples and should, therefore, be treated with caution.
Captivity is known to modify the physiology and behaviour of individuals (Mason Reference Mason2010; Ramos-Güivas et al. Reference Ramos-Güivas, Jawor and Wright2021). Nesting behaviour is highly hormone-dependent (Angelier et al. Reference Angelier, Holberton and Marra2009, Reference Angelier, Wingfield, Tartu and Chastel2016), and disruptions in normal levels of related hormones due to disturbances in the captive environment can affect the expression of typical nesting behaviours. The proximity of other breeding pairs and anthropogenic disturbances can affect the time individuals spend inside the nest, consequently affecting nestlings’ feeding events. In this captive breeding programme, the breeding cages are approximately 1.5–3 m from each other, whereas in the wild, the closest nests are no less than 30 m apart. Parrots make loud vocalisations, and the proximity of breeding pairs can trigger continuous territorial displays such that birds attending the nest may emerge more frequently to observe and respond to these displays. Disrupted nest attendance by the female may lead to lower reproductive success, as previously seen in wild populations of this species (Wilson et al. Reference Wilson, Wilson and Field1997). The difference in nest proximity between wild and captive populations may contribute to the shorter feeding bouts observed in captivity and explain why wild pairs produce more fledglings than their captive counterparts.
We did find a tendency for wild males to exhibit more variation in total feeding bouts and mean feeding bout duration than their captive counterparts (Figure 2a and b). When females are constantly in the nest, it is possible that when the males enter on some visits, the nestlings are satiated, and thus males engage in fewer and shorter feeding bouts. However, the opposite may be true on other occasions, where males are the first to feed, and nestlings are hungrier. Foraging behaviour for wild individuals is unpredictable, and environmental effects such as distance from food sources can dictate how long and often males take to return with food to the nest (Grissot et al. Reference Grissot, Araya-Salas, Jakubas, Kidawa, Boehnke and Błachowiak-Samołyk2019) and the quantity of food they can provide. For the observation periods we measured, both sexes in captivity tended to have fewer total feeding bouts and shorter feeding bouts than their wild counterparts. Wild individuals experience substantial environmental heterogeneity, potentially influencing behavioural patterns shown by the different pairs, especially when the data were taken in different years.
Previous studies have found a relationship between pair duration and better parental coordination (Brooke Reference Brooke1978; Cooke et al. Reference Cooke, Bousfield and Sadura1981; Fowler Reference Fowler1993; Ollason and Dunnet Reference Ollason and Dunnet1986). This relationship was not statistically significant for the parental care behaviours measured in our study. Monogamous mated pairs, when a nesting attempt fails, could divorce and, if given the opportunity, choose a new mate. However, this option is not always available in captive conditions where the manager controls pairings. Non-compatible pairs can maintain bonds for extended periods while exhibiting poor reproductive performance, potentially linked to deficient parental care behaviours stemming from incompatibility.
Inbreeding
Our analysis revealed no significant association between reproductive success, as measured by the number of fledglings produced and pair-level inbreeding coefficients. While inbreeding depression is a well-documented phenomenon affecting various fitness components in birds (Hemmings et al. Reference Hemmings, Slate and Birkhead2012), our study did not detect a significant impact of inbreeding coefficients on fledgling production. This discrepancy could be attributed to the limited sample size (n = 8) and the relatively narrow range of inbreeding coefficients observed in our data set. These findings underscore the importance of considering species-specific and contextual factors when assessing the effects of inbreeding on reproductive success.
Conservation implications
The Puerto Rican Parrot has been a model for avian species conservation for over 50 years since its decline was first recognised in 1955 (Rodriguez-Vidal Reference Rodriguez-Vidal1959). The on-going decline of many parrot species (Berkunsky et al. Reference Berkunsky, Quillfeldt, Brightsmith, Abbudd, Aguilare and Alemán-Zelayaf2017; Snyder et al. Reference Snyder, McGowan, Gilardi and Grajal2000; Vergara-Tabares et al. Reference Vergara-Tabares, Cordier, Landi, Olah and Nori2020; Wright et al. Reference Wright, Lewis, Lezama-López, Smith-Vidaurre and Dahlin2018) means that captive reproductive programmes may become the norm in efforts to avoid extinction (Bowkett Reference Bowkett2009; Leus Reference Leus2011; Seal et al. Reference Seal, Foose, Ellis, Olney, Mace and Feistner1994). One of the main challenges for the Puerto Rican Parrot Program has been the production of fledglings, particularly in captivity. We studied parental care behaviours, explored their relationship to reproductive success, and found that mean feeding bout duration positively relates to reproduction. These parental behaviours can be affected by captive conditions, parental sex, and mean nestling’s age at the time of the parents’ visit. The parental care behaviours we studied are simple to measure and can be applied to any species with biparental care. Continuous video monitoring, potentially coupled with artificial intelligence-assisted scoring of parental provisioning performance, holds promise for improving captive productivity by allowing earlier intervention in nests with poor parental care and retroactively identifying sources of reproductive failure, while reducing the stress caused by anthropogenic manipulation. Future studies could monitor the level of competition among nestlings to assess the role it may play in the condition and survival of nestlings, and evaluate the effect of housing conditions and staff activities on parental care behaviours.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0959270925100208.
Acknowledgements
This research was supported by the Puerto Rico Department of Natural and Environmental Resources, the U.S. Fish and Wildlife Service (USFWS), and the World Parrot Trust. It was conducted with the permission of the Institutional Animal Care and Use Committee of New Mexico State University (IACUC protocol 2014-030 and 2021-014). We thank Miguel A. García-Bermudez, Ricardo Valentín de la Rosa, Tanya Martínez-Ramirez, Jong Piel Banchs Plaza, Roseanne Medina-Miranda, Alberto Álvarez, and Tomás Medina-Cortés from the PR DNER for their cooperation during the study. We thank Marisel López, Jafet Vélez, Iris Rodriguez-Carmona, Gabriel Benitez-Soto, Limary Ramírez-Esquilín, and Arelys Johnson-Camacho from the USFWS for their cooperation during the study. We thank members of the Wright Lab at the New Mexico State University for sharing their knowledge and helping improve the study with their suggestions during the development of the manuscript. We thank all the volunteers for helping to develop the methodology of data collection and for collecting the data: Kelly Fraguada, Pablo A. Sosa-Negron, Natasha L. Perez-Nieto, Mariana Travieso-Diffoot, Gabriela Fonseca-Cordero, Adriana Pérez-Vega, Adriana Zaruk-Sierra, Facundo Quintanela, Alejandro Rivera-Madera, Samantha Overcashier, Kiara Holloway, Kennysha Ramirez, Saelly M. Martinez-Rivera, Jheismar A. Santos-Rodriguez, Legna Alfonzo-Rosario, Rey Febo, Karla Vilches, Ashley Pacheco, Zulymar Ramos, Giselle Garcia-Negron, Gloriana Velazquez, Arleen Rodriguez-Serrano, Deyaneira M. Rodriguez-Muñoz, Luis Y. Colón-Dávila, Francisco Castro-Rivera, Gabriela Montalvo-Rodriguez, Wanda Gonzalez, and Ricardo Maldonado-Acosta.