


Introduction
Invasive grasses are a global threat because they reduce native plant abundance and diversity, alter wildlife habitat, and transform ecosystem function (Pyšek et al. Reference Pyšek, Jarošík, Hulme, Pergl, Hejda, Schaffner and Vilà2012). One primary mechanism for these shifts is that invasive grasses can increase wildfire activity in dryland ecosystems that historically supported sparse native vegetation that was not fire-adapted (Brooks et al. Reference Brooks, D’Antonio, Richardson, Grace, Keeley, DiTomaso, Hobbs, Pellant and Pyke2004; D’Antonio and Vitousek Reference D’Antonio and Vitousek1992; Fusco et al. Reference Fusco, Finn, Balch, Nagy and Bradley2019). As invaded areas burn more frequently, native vegetation is damaged or killed and cannot regenerate as quickly as invasive grasses, thereby creating an invasive grass–fire cycle that is not easily broken (D’Antonio and Vitousek Reference D’Antonio and Vitousek1992). Evidence indicates that invasive grasses have become more widespread and abundant in recent decades (Smith et al. Reference Smith, Allred, Boyd, Davies, Jones, Kleinhesselink, Maestas, Morford and Naugle2022; Wilder et al. Reference Wilder, Jarnevich, Baldwin, Black, Franklin, Grissom, Hovanes, Olsson, Malusa, Kibria and Li2021), with much of this increase attributable to land-use intensification (Chambers and Wisdom Reference Chambers and Wisdom2009) and climate change (Abatzoglou and Kolden Reference Abatzoglou and Kolden2011). Understanding how invasive grasses change fuel characteristics and wildfire behavior and which management treatments can control their abundance is essential to reduce negative impacts to ecosystem health.
Multiple studies have sought to understand the influence of invasive grasses on fuel or fire characteristics and evaluated treatment effectiveness (reviewed in DiTomaso Reference DiTomaso2000; Laushman et al. Reference Laushman, Munson and Villarreal2020). However, these studies typically focus on a single species or treatment, occur within a limited spatial extent and over a short time window, narrowly define a wildfire or fuel metric, and do not fully consider how treatment characteristics modify their effectiveness. Synthesis across studies is needed to reveal the broad role of invasive grasses in the fire cycle and identify strategies for effectively controlling their abundance. Integrating results can allow comparison of patterns of wildfire and treatment effectiveness across ecoregions with different environmental contexts and invasive grass species to inform management strategies. Expanding knowledge of the invasive grass–fire cycle and treatment effectiveness can also reveal gaps in our understanding to help facilitate future research.
The Intermountain West has experienced rapid spread of multiple invasive grass species and associated increases in wildfire frequency in recent decades (Balch et al. Reference Balch, Bradley, D’Antonio and Gómez-Dans2013; Brooks Reference Brooks1999b; Crist et al. Reference Crist, Belger, Davies, Davis, Meldrum, Shinneman, Remington, Welty and Mayer2023; Hanselka Reference Hanselka1988; Mack Reference Mack1981). The distributions of these invasive grass species are largely controlled by precipitation seasonality and temperature, with annual cool-season (C3) grass species occupying ecoregions that primarily receive October to March precipitation, including cheatgrass (Bromus tectorum L.) and medusahead [Taeniatherum caput-medusae (L.) Nevski] in northern latitudes, and red brome (Bromus rubens L.) and Mediterranean grass [Schismus arabicus Nees and Schismus barbatus (Loefl. ex L.) Thell] in southern latitudes. Perennial warm-season (C4) grass species, including buffelgrass [Pennisetum ciliare (L.) Link] and Lehmann’s lovegrass (Eragrostis lehmanniana Nees), occur in ecoregions that receive April to September precipitation in southern latitudes (Wilder et al. Reference Wilder, Jarnevich, Baldwin, Black, Franklin, Grissom, Hovanes, Olsson, Malusa, Kibria and Li2021).
Periods of high precipitation can encourage the growth of invasive grasses that fill interspaces between sparsely distributed native vegetation with continuous fine fuels that can dry out and persist, creating potential for wildfire (Brooks et al. Reference Brooks, D’Antonio, Richardson, Grace, Keeley, DiTomaso, Hobbs, Pellant and Pyke2004; Brooks and Matchet Reference Brooks and Matchett2006). In a previous comparison of fire effects across ecoregions, many of the invasive grass species in the Intermountain West increased fire occurrence rates, size, and frequency, with up to a 3-fold increase in fire occurrence for areas invaded by S. barbatus and P. ciliare (Fusco et al. Reference Fusco, Finn, Balch, Nagy and Bradley2019). Knowledge of the feedback between fuel and fire, the metrics used to assess changes in the fire cycle, and the time required for fuel accretion can increase understanding of how invasive grasses alter fire regimes. Comparing how different invasive grass species in the Intermountain West change these fuel characteristics and fire behavior can inform risk evaluation of valued ecosystem and human assets.
Management treatments to reduce invasive grasses and mitigate wildfire risk are widely implemented across drylands and require large resource investments (Munson et al. Reference Munson, Yackulic, Bair, Copeland and Gunnell2020). The size, costs, and complexity of these treatments have been increasing over the last few decades (Copeland et al. Reference Copeland, Munson, Pilliod, Welty, Bradford and Butterfield2018), making it imperative to find strategies that are effective and long-lasting. Common methods to reduce invasive grasses include chemical treatments that consist of herbicide application, mechanical removal, applying weed-suppressive bacteria, prescribed fire, targeted livestock grazing, seeding of desired species, and soil manipulation (DiTomaso Reference DiTomaso2000). Treatment success can depend on the methods used, how and when they are applied, and the environmental context of targeted areas. For example, herbicides may target specific plant taxa and only be effective if they are applied at a critical stage of plant development (Hirsch et al. Reference Hirsch, Monaco, Call and Ransom2012). A combination of treatment types or repeat treatment applications may increase effectiveness because there is more than one mechanism to suppress invasive grasses or a reduction in the time elapsed since treatment, respectively. Comparing the influence of different treatment types for the same invasive grass species and evaluating whether the same treatment type works across different invasive grass species allow natural resource managers to understand the context in which management actions are effective.
We conducted a systematic review and meta-analysis of peer-reviewed scientific literature (1985 to 2023) on the aforementioned four cool-season annual and two warm-season perennial invasive grass species in drylands of the Intermountain West. Our objectives were to compare: (1) the influence of different invasive grass species on fuel and wildfire characteristics and (2) the effectiveness of different treatment types on reducing the amount of the focal invasive grass species.
Materials and Methods
Literature Search
We used Google Scholar to search the scientific literature with combinations of focal invasive grass species names (common and scientific, including all synonyms) and keywords such as “control”, “treatment”, “fire”, or “fire behavior” (Supplementary Table S1). Schismus spp. include S. arabicus, S. barbatus, or both, as most literature we reviewed combined the two species due to difficulty differentiating them in the field where they both occur. Peer-reviewed journal articles were selected for review if they: (1) were conducted in the Intermountain West or adjoining ecoregion; (2) had quantifiable data related to fuel, wildfire behavior, or management treatments to reduce one or more focal species; and (3) were conducted in a natural system or in controlled lab, greenhouse, or field environments meant to simulate this system (actively cropped or urban systems were excluded). We conducted Google searches for relevant management databases containing references to peer-reviewed literature to capture studies not found in Google Scholar searches. Eragrostis lehmanniana, P. ciliare, and Schismus spp. had a particularly limited body of qualifying peer-reviewed literature, so we narrowly expanded our search to include study areas outside the Intermountain West, and quantitative technical reports that fit all other criteria were included. We also identified primary research articles from review articles and included them in our review and analyses.
Searches generated 810 unique scientific literature sources published between 1985 and 2023, of which 204 were selected for analysis based on our selection criteria. Selected sources were organized in a database according to the research objectives they addressed: (1) fuel and wildfire characteristics, (2) treatment effectiveness, or (3) both (Costanzo and Munson Reference Costanzo and Munson2025). For each study, we extracted background information including the study location, ecoregion (EPA Level III, https://www.epa.gov/eco-research/level-iii-and-iv-ecoregions-continental-united-states), and plant community and then summarized major findings relevant to research objectives. If a general location or unspecified location was provided, we approximated the center of the study area using the best information provided in the study area description.
Fuel and Fire Characteristics
Studies exploring the effects of invasive grasses on fuel and fire characteristics were organized into categories that corresponded to the two primary parts of the invasive grass–fire cycle: (1) “fuel,” focused on how invasive grasses influence fuel characteristics and how fuel regenerates after wildfire; and (2) “fire,” focused on how fuel created by invasive grasses influences wildfire behavior and other characteristics. Fires described in each study were characterized by type, including prescribed and managed burns, wildfires, models, and laboratory studies as long as they provided quantifiable data. Fires used for targeted control of invasive grasses were not included in this portion of the study. For the fuel studies, we extracted metrics that quantified the amount of fuel (e.g., biomass or grass cover in burned and unburned areas) from text or tables, or we visually approximated from figures with a separate data point for each monitoring point, study site, and most common metric measured (Costanzo and Munson Reference Costanzo and Munson2025).
Treatment Effectiveness
For studies that assessed treatments to reduce the amount of invasive grasses, we extracted treatment type, the number of times treated, time since first and most recent treatment, and whether plots had burned before treatments. We extracted raw values on the amount of invasive grasses in treated and untreated areas for each monitoring point, study site, and the metric measured (Costanzo and Munson Reference Costanzo and Munson2025).
Data Analysis
Effect sizes were calculated as the natural log of the ratio of invasive grass amount between burned and unburned or treated and control pairs to standardize our response variable across different monitoring metrics. Multiple effect-size data points could be contributed by a single study if multiple monitoring periods, distinct sites, treatment types, and/or treatment intensities were evaluated. We used linear regressions to evaluate how the effect size changed with time since fire for each invasive grass species except for T. caput-medusae, which did not have sufficient data. Prescribed or managed fires and wildfires were included in this analysis. We limited the analysis to <15 yr after fire, as data beyond this time were scarce for all species.
ANOVA was performed to compare effect sizes among treatment characteristics. To evaluate effectiveness by treatment type, we only used data from the application of a single treatment type, not a combination of treatments. Effectiveness was statistically compared across treatment type within each species, as well as among species within each treatment type. We also conducted ANOVA for each species to compare the effect sizes for different numbers of treatments (once, twice, or three or more times). All data met assumptions of normality. We used Tukey’s Honestly Significant Difference (HSD) tests to perform pair-wise comparisons on these different factors using the emmeans package (Lenth Reference Lenth2025). For all ANOVAs, there had to be a minimum of three estimates of effect size from separate studies within each category considered for comparison to be included. We used linear regressions to evaluate how time since treatment explained variation in the effect size. All analyses were conducted in R v. 4.5.0, and figures were generated using the ggplot2 package (Wickham Reference Wickham2016).
Results and Discussion
Study Characteristics
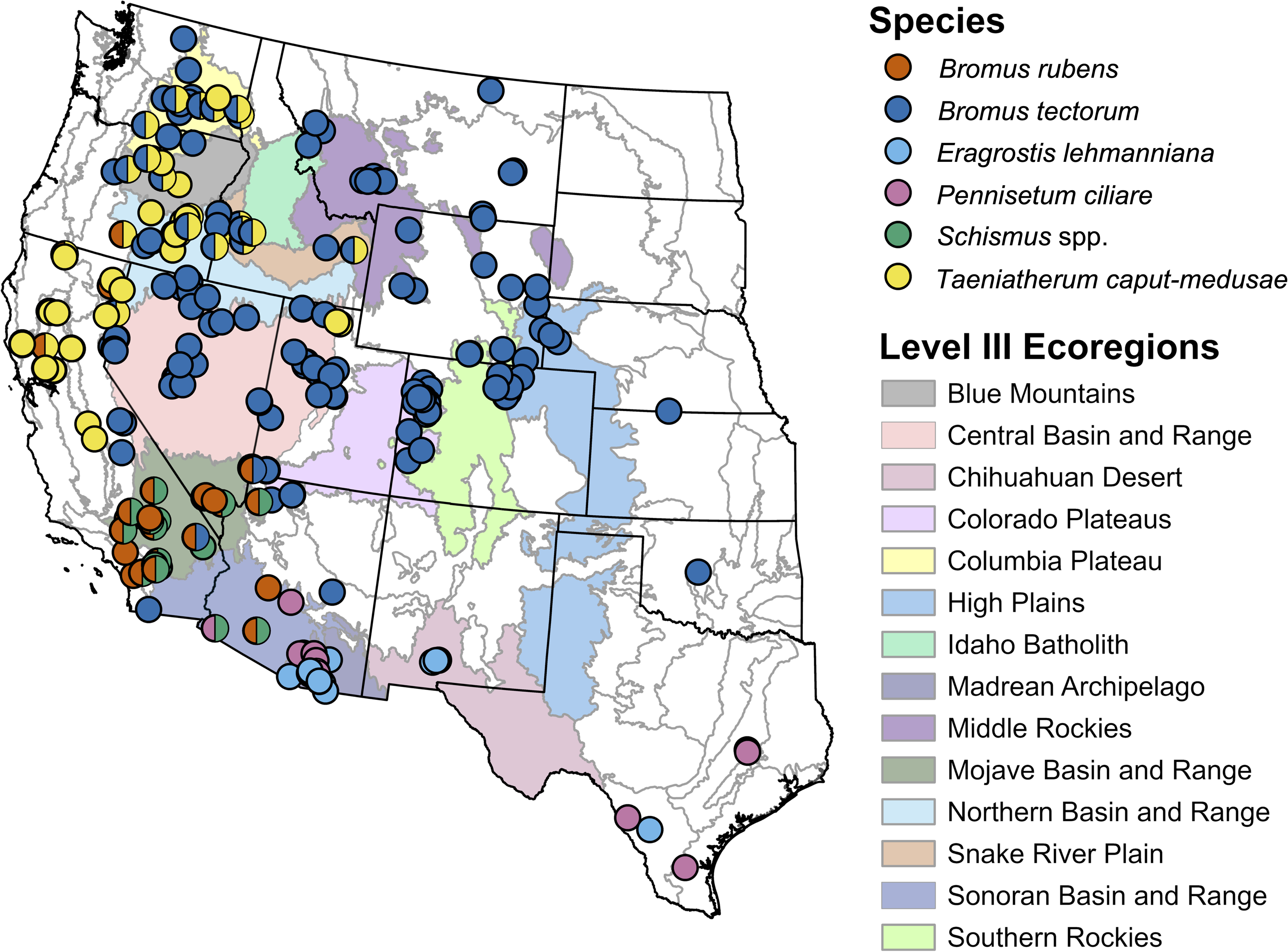
Studies on B. tectorum were the most broadly distributed, with a focus in the Great Basin (Central and Northern Basin and Ranges, Snake River Plain ecoregions), Columbia Plateau, Colorado Plateaus, and High Plains (Figure 1). Studies on T. caput-medusae geographically overlapped with B. tectorum to some degree but spanned farther west into portions of California. Bromus rubens and Schismus spp. studies occurred in the Mojave Basin and Range and isolated portions of the Sonoran Basin and Range, while P. ciliare studies largely occurred in the Sonoran Basin and Range but included some portions of Texas. Studies on E. lehmanniana were concentrated in the Madrean Archipelago and Chihuahuan Desert.
Location of studies on dominant invasive grasses in the Intermountain West, USA, and adjoining ecoregions by species and EPA Level III ecoregion. Other study locations with information relevant to the review that are not shown include Hawaii, USA (1), Mexico (4), South Africa (1), and Australia (2). Ecoregions indicated by color on the map and legend are those in which five or more studies were conducted.

Of the 204 selected articles, 68 provided information about invasive grass effects on fuel and fire characteristics, 125 provided information on treatment effectiveness, and 11 articles fit into both categories (Table 1). Bromus tectorum was the most frequently studied, with 57.3% of articles, followed by T. caput-medusae at 21.6%, B. rubens at 13.7%, P. ciliare at 10.8%, Schismus spp. at 8.3%, and E. lehmanniana being the least frequently studied at 7.8%.
Number of articles that report fuel and fire characteristics and management treatment effectiveness for dominant invasive grass species in the Intermountain West, USA.

a Columns/rows do not add up to totals due to articles that either fit into both categories or studied more than one grass species.
Fuel and Fire Characteristics
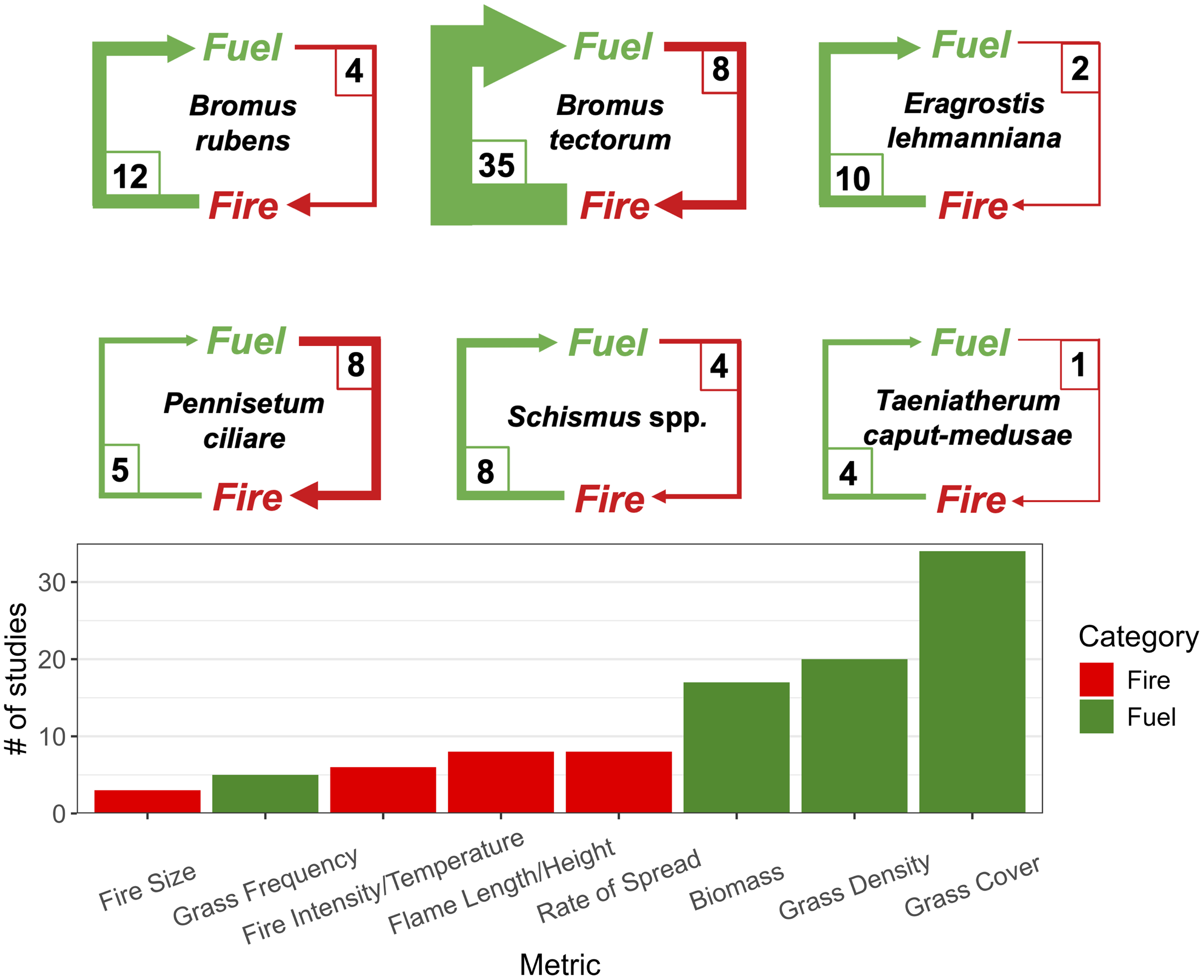
There were 61 articles that characterized the influence of invasive grasses on fuel characteristics and how fuels regenerate following wildfire, while 21 articles focused on invasive grass controls on fire behavior. Fuel articles were two to five times more abundant than fire articles for B. rubens, B. tectorum, E. lehmanniana, and Schismus spp., while there were two times as many fire articles for P. ciliare, and the fewest overall number of articles for T. caput-medusae, with only one of them focused on fire (Figure 2).
Number of studies that address the cycle on how dominant invasive grasses of the Intermountain West influence fuel characteristics and regeneration after wildfire (Fuel), and how fuel created by invasive grasses influences wildfire behavior and other characteristics (Fire) shown by species (top) and the four most common metrics studied in each study category (bottom).

The most common metric characterizing fuel was grass cover (34 studies), with density, biomass, and frequency used less frequently in 20, 17, and 5 studies respectively (Figure 2). These fuel characteristics were frequently tracked following a fire for as long as decades with an average monitoring period of 7 yr. Rate of spread was the most common fire metric (8 studies), with flame length/height, fire intensity/temperature, and fire size also reported. Most studies conducted field measurements of prescribed burns (34 articles) or wildfires (35 articles), but 4 articles performed laboratory experiments, and 16 articles generated models for invasion or burn risk based on historical or current fire data.
The greater number of studies on fuel compared to fire were partially due to the difficulty in accessing a fire and measuring its behavior in the field (Johnston and Chapman Reference Johnston and Chapman2014; Link et al. Reference Link, Hill and Bansal2019; Morris et al. Reference Morris, Morris and Surface2016) or in a burning device (Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007) and suggests opportunity for field-deployed sensors and remote sensing technologies to improve understanding of invasive grasses’ influence on fire behavior. The common use of canopy cover to characterize how invasive grasses influence fuel is due to its widespread inclusion in monitoring protocols, but this metric differs from fuel in that it does not consider the density, shape, arrangement, and total amount of biomass that could be consumed in a fire. Studies that combine grass canopy cover and density with biomass (Brooks Reference Brooks1999a; DiTomaso et al. Reference DiTomaso, Kyser, George, Doran and Laca2008; Jones et al. Reference Jones, Chambers, Board, Johnson and Blank2015; Prevéy et al. Reference Prevéy, Knochel and Seastedt2014) can improve predictions of fire probability and provide important allometries for estimating fire behavior, including flame length and rate of spread. Many estimates of cover were to assess post-fire recovery, and fewer studies assessed fuel before the occurrence of the fire, which could improve early warning of potential fire behavior before a fire occurs (Banks and Baker Reference Banks and Baker2011; Porensky et al. Reference Porensky, Perryman, Williamson, Madsen and Leger2018).
Effects on Fire Behavior
While fire behavior data were not substantive enough to perform meta-analysis, quantitative studies found that warm-season perennial grasses P. ciliare and E. lehmanniana were associated with higher fuel loads and higher-severity fire than in nearby uninvaded or restored areas (Kupfer and Miller Reference Kupfer and Miller2005; McDonald and McPherson Reference McDonald and McPherson2011b, Reference McDonald and McPherson2013; McKenna et al. Reference McKenna, Glenn, Erskine, Doley and Sturgess2017; Miller et al. Reference Miller, Friedel, Adam and Chewings2010; Schlesinger et al. Reference Schlesinger, White and Muldoon2013). Conversely, studies on cool-season annual grasses B. rubens and Schismus spp. associated these grasses with low-severity, quick-spreading fire behavior (Brooks Reference Brooks1999a; Fuentes-Ramirez et al. Reference Fuentes-Ramirez, Veldman, Holzapfel and Moloney2016; Moloney et al. Reference Moloney, Mudrak, Fuentes-Ramirez, Parag, Schat and Holzapfel2019).
Annuals such as B. rubens, B. tectorum, Schismus spp., and T. caput-medusae introduced fire risk to historically sparse shrublands where they typically invade by increasing fuel load and continuity across shrub interspaces and expand the fire season by drying out earlier than native herbaceous vegetation (Brooks Reference Brooks1999b; D’Antonio and Vitousek Reference D’Antonio and Vitousek1992). Because many native shrubs and other life-forms in these arid communities have not experienced frequent wildfire, they are vulnerable to damage and mortality from fire (Brooks et al. Reference Brooks, D’Antonio, Richardson, Grace, Keeley, DiTomaso, Hobbs, Pellant and Pyke2004; D’Antonio and Vitousek Reference D’Antonio and Vitousek1992). Perennials P. ciliare and E. lehmanniana invaded multiple plant community types, though E. lehmanniana is more common in grasslands and may increase fire frequency and severity, with neutral to negative effects to native grasses (Banks and Baker Reference Banks and Baker2011; Ellsworth et al. Reference Ellsworth, Wrobleski, Kauffman and Reis2016; Roundy et al. Reference Roundy, Chambers, Pyke, Miller, Tausch, Schupp, Rau and Gruell2018; Thacker et al. Reference Thacker, Ralphs and Monaco2009). Quantitative fire behavior information was most abundant for B. tectorum, as studies took advantage of abundant long-term cover and fire data, especially across the Great Basin and Mojave Desert, to associate high B. tectorum cover with fire risk (Balch et al. Reference Balch, Bradley, D’Antonio and Gómez-Dans2013; Bradley et al. Reference Bradley, Curtis, Fusco, Abatzoglou, Balch, Dadashi and Tuanmu2018; Knapp Reference Knapp1998; Link et al. Reference Link, Keeler, Hill and Hagen2006; Williamson et al. Reference Williamson, Fleishman, Mac Nally, Chambers, Bradley, Dobkin, Board, Fogarty, Horning, Leu and Wohlfeil Zillig2020). Extending these approaches across ecoregions and to other dominant invasive grass species (e.g., T. caput-medusae) could help to fill knowledge gaps and inform risk assessments for newer invasions.
Invasive Grass Fuel Regeneration through Time
We found that the difference in the amount of invasive grasses between burned (managed fires and wildfires) and unburned areas significantly changed through time for B. tectorum (P < 0.05) and E. lehmanniana (P < 0.1), but not for other species (Figure 3; Supplementary Table S2). The amount of B. tectorum was initially lower in burned areas, then became equal with unburned areas in the first 2 yr, followed by increases through time such that the invasive grass was 22 times higher in burned compared with unburned areas after 10 yr. In contrast, the amount of E. lehmanniana was initially higher in burned areas, but decreased through time such that burned areas reached the same effect size as unburned areas after 3 yr. This discrepancy is likely tied to the similarity in structure, life history, and fire history of the invasive grass compared with the native vegetation. Eragrostis lehmanniana is a perennial grass that can increase immediately following fire through its extensive root system but then competes with other perennial grasses that are similar in structure and fire history over longer periods of time. Studies found that fire increases germinability and seedbank density of E. lehmanniana (Ruyle et al. Reference Ruyle, Roundy and Cox1988; Snyman and van Wyk Reference Snyman and van Wyk2005; Sumrall et al. Reference Sumrall, Roundy, Cox and Winkel1991) but does not increase establishment of adult plants in the long term (Ayala-A et al. Reference Ayala-A, Ortega-S, Fulbright, Rasmussen, Drawe, Synatzske and Litt2012; Bock and Bock Reference Bock and Bock1992; McDonald and McPherson Reference McDonald and McPherson2011a; McGlone Reference McGlone2013; McGlone and Huenneke Reference McGlone and Huenneke2004). Evidence of similar patterns was also observed for P. ciliare (Cox et al. Reference Cox, Ibarra-F and Martin-R1990; Fensham et al. Reference Fensham, Donald and Dwyer2013; Snyman and van Wyk Reference Snyman and van Wyk2005).
Effect size (ln[amount in burned/unburned]) of invasive grass species related to time since fire, colored by warm-season perennial (red) and cool-season annual (blue) grasses. Taeniatherum caput-medusae was omitted due to insufficient data. Statistical significance: *P < 0.1; **P < 0.05.

Conversely, studies largely concluded that fire resulted in higher establishment of B. tectorum and the other cool-season annual grasses over time (Banks and Baker Reference Banks and Baker2011; Berleman et al. Reference Berleman, Suding, Fry, Bartolome and Stephens2016; Bishop et al. Reference Bishop, Gill, McMillan and St Clair2020; Brooks Reference Brooks2002, Reference Brooks2011; Brown and Minnich Reference Brown and Minnich1986; Callison et al. Reference Callison, Brotherson and Bowns1985; Davy and Dykier Reference Davy and Dykier2017; Getz and Baker Reference Getz and Baker2008; Gill et al. Reference Gill, O’Connor, Rhodes, Bishop, Laughlin and St Clair2018; Horn and St Clair Reference Horn and St Clair2017; Jurand and Abella Reference Jurand and Abella2013; Kulpa et al. Reference Kulpa, Leger, Espeland and Goergen2012; McGlone et al. Reference McGlone, Springer and Covington2009; Rodhouse et al. Reference Rodhouse, Irvine and Bowersock2020; Shinneman and Baker Reference Shinneman and Baker2009; Underwood et al. Reference Underwood, Klinger and Brooks2019; West and Hassan Reference West and Hassan1985), even though observations of seed death and reduced germinability were common immediately following fire (Abella et al. Reference Abella, Engel, Lund and Spencer2009; Berleman et al. Reference Berleman, Suding, Fry, Bartolome and Stephens2016; Bishop et al. Reference Bishop, Gill, McMillan and St Clair2020; Brooks Reference Brooks2002; Davy and Dykier Reference Davy and Dykier2017; Gill et al. Reference Gill, O’Connor, Rhodes, Bishop, Laughlin and St Clair2018). Of the annual grasses, only Schismus spp. had high establishment immediately following fire (Bishop et al. Reference Bishop, Gill, McMillan and St Clair2020; Brooks Reference Brooks2002; Gill et al. Reference Gill, O’Connor, Rhodes, Bishop, Laughlin and St Clair2018; Steers and Allen Reference Steers and Allen2012), which may be attributable to its fire-resilient seed (Esque et al. Reference Esque, Young and Tracy2010).
Recovery and invasion mechanisms following initial seed mortality were explored most frequently for B. tectorum in field and greenhouse studies, where researchers documented advantages in water and nutrient acquisition that allowed B. tectorum to respond more quickly than native plants and outcompete native perennial bunchgrass seedlings (Gundale et al. Reference Gundale, Sutherland and DeLuca2008; Hassan and West Reference Hassan and West1986; Melgoza et al. Reference Melgoza, Nowak and Tausch1990). Many of the native plants where annual grasses occur in the Intermountain West are shrubs that access deep soil moisture and do not compete as much with shallow-rooted annuals. However, many of these shrubs are not historically fire-adapted and can be lost from frequent, invasive grass–carried wildfires (Crist et al. Reference Crist, Belger, Davies, Davis, Meldrum, Shinneman, Remington, Welty and Mayer2023; Fuentes-Ramirez et al. Reference Fuentes-Ramirez, Veldman, Holzapfel and Moloney2016), further enabling shifts to invasive grass–dominated communities (D’Antonio and Vitousek Reference D’Antonio and Vitousek1992; Mack Reference Mack1981).
Across species, many studies concluded that fuel regeneration and fire risk are most closely tied to short-term precipitation patterns rather than fire characteristics directly (Brooks Reference Brooks2011; Fensham et al. Reference Fensham, Donald and Dwyer2013; Geiger and McPherson Reference Geiger and McPherson2005; Jarnevich et al. Reference Jarnevich, Young, Cullinane Thomas, Grissom, Backer and Frid2020; Knapp Reference Knapp1998; McDonald and McPherson Reference McDonald and McPherson2011a; Moloney et al. Reference Moloney, Mudrak, Fuentes-Ramirez, Parag, Schat and Holzapfel2019; Peeler and Smithwick Reference Peeler and Smithwick2018; Taylor et al. Reference Taylor, Brummer, Rew, Lavin and Maxwell2014). Short-term climate projections could aid land managers in determining when and how to allocate resources and manage risk in invaded areas following fires (Smith et al. Reference Smith, Allred, Boyd, Davies, Kleinhesselink, Morford and Naugle2023).
Treatment Characteristics
Chemical treatment was the most common treatment type among all species, occurring in 58% of treatment studies and consisting of herbicide application (Table 2). Across all chemical treatments, 28 different herbicides were used, with the most common being imazapic (46 studies) and glyphosate (31). Aminopyralid (8), picloram (6), tebuthiuron (6), fluazifop (5), indaziflam (5), and rimsulfuron (5) were other commonly used herbicides (Supplementary Table S3). Imazapic was extensively employed to control both B. tectorum and T. caput-medusae, as it specifically targets annual grasses by inhibiting enzymes in new rather than mature plant tissue and can be applied during a time when co-occurring native vegetation is dormant (Mangold et al. Reference Mangold, Parkinson, Duncan, Rice, Davis and Menalled2013; Shaner and O’Connor Reference Shaner and O’Connnor1991; Terry et al. Reference Terry, Madsen, Gill, Anderson and St Clair2021a). Indaziflam is used similarly in a preemergence stage, but has longer seedbank persistence and has only recently become more widely used (Sebastian et al. Reference Sebastian, Nissen, Sebastian and Beck2017; Terry et al. Reference Terry, Madsen, Gill, Anderson and St Clair2021a) These herbicides differ functionally from postemergence contact herbicides, which can potentially have larger impacts on non-target plant species if applied nonselectively. Fluazifop was the herbicide most commonly used for B. rubens and Schismus spp. This herbicide does not affect broadleaved plants and can remain active in the soil (Allen et al. Reference Allen, Cox, Tennant, Kee and Deutschman2005; Cox and Allen Reference Cox and Allen2008; Steers and Allen Reference Steers and Allen2010). The nonselective herbicide glyphosate was the herbicide most commonly used for E. lehmanniana and P. ciliare (Biedenbender and Roundy Reference Biedenbender and Roundy1996; Buerdsell and Lehnhoff Reference Buerdsell and Lehnhoff2023; Crimmins and McPherson Reference Crimmins and McPherson2008; Dixon et al. Reference Dixon, Dixon and Barrett2002; McDonald and McPherson Reference McDonald and McPherson2013; Rowe et al. Reference Rowe, Sprague, Fastiggi and Staker2023; Sumrall et al. Reference Sumrall, Roundy, Cox and Winkel1991).
Description and number of articles studying different treatment types intended to reduce dominant invasive grass species in the Intermountain West, USA.

Seeding was the second most common treatment type (29% of studies) and was primarily intended to revegetate native species to compete with invasive species. These treatments were common for B. rubens, B. tectorum, and T. caput-medusae, but were used in two or fewer studies for all other species. Only about a third of these studies evaluated seeding as the only treatment, as most applied seed following chemical or fire treatments to suppress reinvasion. Most studies used native seed, but a small number evaluated using non-native species such as forage kochia [Bassia prostrata (L.) A.J. Scott] or crested wheatgrass [Agropyron cristatum (L.) Gaertn.] to better compete with the target invasive grass (Aryal and Islam Reference Aryal and Islam2018; Clements et al. Reference Clements, Harmon and Blank2022; Davies and Johnson Reference Davies and Johnson2017; Davies and Sheley Reference Davies and Sheley2011; Davies et al. Reference Davies, Boyd, Johnson, Nafus and Madsen2015b; Porensky et al. Reference Porensky, Perryman, Williamson, Madsen and Leger2018; Rose et al. Reference Rose, Hild, Whitson, Koch and Van Tassell2001; Stonecipher et al. Reference Stonecipher, Thacker, Welch, Ralphs and Monaco2019; Thacker et al. Reference Thacker, Ralphs and Monaco2009; Urza et al. Reference Urza, Weisberg, Chambers, Board and Flake2019). When compared, seeding non-native species outperformed native species at suppressing target invasive grasses in some cases, with studies finding highest success using A. cristatum to control B. tectorum and T. caput-medusae in the Great Basin (Clements et al. Reference Clements, Harmon and Blank2022; Davies et al. Reference Davies, Boyd, Johnson, Nafus and Madsen2015b; Thacker et al. Reference Thacker, Ralphs and Monaco2009). In studies testing different seeding rates, higher rates of seeding increased both seeding success and invasive grass suppression (Aryal and Islam Reference Aryal and Islam2018; Porensky et al. Reference Porensky, Perryman, Williamson, Madsen and Leger2018, Reference Porensky, Baughman, Williamson, Perryman, Madsen and Leger2021; Rose et al. Reference Rose, Hild, Whitson, Koch and Van Tassell2001; Schantz et al. Reference Schantz, Sheley and Hardegree2019). Seeding rates varied widely based on the seeded species and invasion severity.
Targeted prescribed fire was applied in 24% of studies and was most often applied repeatedly in an attempt to deplete the seedbank of the invasive grass and prevent reinvasion. Prescribed fire was used for all invasive grass species except B. rubens and Schismus spp. Fire showed limited to no effects on decreasing invasive grasses when applied alone, especially when monitored beyond 2 yr (Ball Reference Ball2014; Germino and Lazarus Reference Germino and Lazarus2020; Sheley et al. Reference Sheley, Bingham and Davies2012a). Some studies observed an increase in invasive grass when fire was applied alone (Davy and Dykier Reference Davy and Dykier2017; McDonald and McPherson Reference McDonald and McPherson2011a; Pyke et al. Reference Pyke, Shaff, Chambers, Schupp, Newingham, Gray and Ellsworth2022; Roundy et al. Reference Roundy, Chambers, Pyke, Miller, Tausch, Schupp, Rau and Gruell2018). However, many studies observed that burning before herbicide application increased treatment effectiveness (Calo et al. Reference Calo, Brause and Jones2012; Davies Reference Davies2010; Davies and Sheley Reference Davies and Sheley2011; Kessler et al. Reference Kessler, Nissen, Meiman and Beck2015; Schantz et al. Reference Schantz, Sheley and Hardegree2019; Seedorf et al. Reference Seedorf, Clark and Nissen2022; Sheley et al. Reference Sheley, Carpinelli and Morghan2007, Reference Sheley, Bingham and Davies2012a) and that implementing pre-fire grazing or post-fire mechanical and/or seeding efforts effectively suppressed reinvasion following fire (Berleman et al. Reference Berleman, Suding, Fry, Bartolome and Stephens2016; Daehler and Goergen Reference Daehler and Goergen2005; Davies et al. Reference Davies, Nafus and Johnson2013, Reference Davies, Boyd, Bates and Hulet2015a, Reference Davies, Bates, Boyd and Svejcar2016a, Reference Davies, Bates, Boyd, O’Connor and Copeland2021a, Reference Davies, Bates, Perryman and Arispe2021b; Davies and Boyd Reference Davies and Boyd2018; Diamond et al. Reference Diamond, Call and Devoe2012; Jones et al. Reference Jones, Chambers, Board, Johnson and Blank2015; McDonald and McPherson Reference McDonald and McPherson2011a).
Mechanical treatment, which consisted of invasive grass removal by machine or by hand was used in 21% of treatment studies and was the most common treatment for B. rubens and Schismus spp. (6 and 4 studies, respectively). These techniques often required multiple applications to effectively suppress invasive grass (Thomson et al. Reference Thomson, Meyer and Whitcomb2021; Valliere et al. Reference Valliere, Balch, Bell, Contreras and Hilbig2019) and were labor intensive (Rowe et al. Reference Rowe, Sprague, Fastiggi and Staker2023). Alone, mechanical treatments were relatively ineffective (Marushia et al. Reference Marushia, Cadotte and Holt2010; Schact and Stubbendieck Reference Schacht and Stubbendieck1985; Steers and Allen Reference Steers and Allen2010; Swanson et al. Reference Swanson, Swanson, Murphy, McAdoo and Schultz2016; Wolden and Stromberg Reference Wolden and Stromberg1997), but when applied with chemical and/or fire treatments they often increased treatment effectiveness by reducing the existing canopy and increasing seedbank exposure for fires or herbicides like imazapic (Daehler and Goergen Reference Daehler and Goergen2005; Jones et al. Reference Jones, Chambers, Board, Johnson and Blank2015; Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Ott et al. Reference Ott, McArthur and Roundy2003; Thomson et al. Reference Thomson, Meyer and Whitcomb2021).
Livestock grazing, or clipping meant to simulate grazing, was applied in 15% of treatment studies. Grazing was used in only one study apiece for B. rubens, E. lehmanniana, and P. ciliare, and not at all for Schismus spp.; and was almost exclusively used in areas of the Great Basin, Northern California and on the Columbia Plateau for B. tectorum, T. caput-medusae, and other invasive plant species in those regions. Grazing had limited effects when applied alone (Diamond et al. Reference Diamond, Call and Devoe2012; Grupenhoff and Molinari Reference Grupenhoff and Molinari2021; Porensky et al. Reference Porensky, Baughman, Williamson, Perryman, Madsen and Leger2021) but showed effectiveness when applied pre- or post-fire (Davies et al. Reference Davies, Bates, Boyd and Svejcar2016a, Reference Davies, Bates, Boyd, O’Connor and Copeland2021a, Reference Davies, Bates, Perryman and Arispe2021b; McDonald and McPherson Reference McDonald and McPherson2011a) and increased herbicide effectiveness in some cases (Lehnhoff et al. Reference Lehnhoff, Rew, Mangold, Seipel and Ragen2019; Whitson and Koch Reference Whitson and Koch1998). When evaluating grazing as a strategy, studies stressed that the seasonality, rainfall amount, and stocking rate were important factors to ensure that invasive grasses were palatable to livestock and that effects on native species were mediated (Condon and Pyke Reference Condon and Pyke2018; Davies et al. Reference Davies, Nafus, Boyd, Hulet and Bates2016b; Davy et al. Reference Davy, Roche, Robertson, Nay and Tate2015; DiTomaso et al. Reference DiTomaso, Kyser, George, Doran and Laca2008; Hempy-Mayer and Pyke Reference Hempy-Mayer and Pyke2008; James et al. Reference James, Gornish, DiTomaso, Davy, Doran, Becchetti, Lile, Brownsey and Laca2015; Porensky et al. Reference Porensky, Perryman, Williamson, Madsen and Leger2018, Reference Porensky, Baughman, Williamson, Perryman, Madsen and Leger2021; Rhodes et al. Reference Rhodes, Plowes and Gilbert2023).
Soil treatments, including physical manipulation of the soil surface (e.g., tilling, mounding) and amendments such as sucrose application intended to tie up soil nutrients, were used in 13% of studies. Soil treatments were used in one study each for B. rubens and P. ciliare and none at all for E. lehmanniana or Schismus spp. Studies that manipulated the soil surface via tilling or creating microtopography found that surface manipulations increased native seeding success (Jernigan et al. Reference Jernigan, McClaran, Biedenbender and Fehmi2016; Johnston Reference Johnston2019; Schantz et al. Reference Schantz, Sheley and Hardegree2019; Whitson and Koch Reference Whitson and Koch1998) and increased herbicide effectiveness in some cases (Johnston Reference Johnston2015; Schantz et al. Reference Schantz, Sheley and Hardegree2019). A carbon source added to tie up nitrogen was usually applied alone and had mixed effects: activated carbon tended to decrease B. tectorum up to 6 yr following application (Kulmatiski Reference Kulmatiski2011; Kulmatiski and Beard Reference Kulmatiski and Beard2006), while sucrose addition was largely ineffective (Mata-Gonzalez et al. Reference Mata-Gonzalez, Hunter, Coldren, McLendon and Paschke2008; Mazzola et al. Reference Mazzola, Chambers, Blank, Pyke, Schupp, Allcock, Doescher and Nowak2011).
Biological treatments, which typically consisted of applying different strains of the annual grass–targeting bacterium Pseudomonas fluorescens, occurred in 6% of studies and were only used on B. tectorum and T. caput-medusae. These treatments were largely ineffective in the field for both species (Germino and Lazarus Reference Germino and Lazarus2020; Pyke et al. Reference Pyke, Shaff, Gregg and Conley2020; Reinhart et al. Reference Reinhart, Carlson, Feris, Germino, Jandreau, Lazarus, Mangold, Pellatz, Ramsey, Rinella and Valliant2020; Tekiela Reference Tekiela2020), and studies that did report success only observed decreases in invasive grasses within the first 3 yr following treatment before populations rebounded (Germino and Lazarus Reference Germino and Lazarus2020; Kennedy Reference Kennedy2018; Lazarus and Germino Reference Lazarus and Germino2021; Lazarus et al. Reference Lazarus, Germino, Brabec, Peterson, Walker and Moser2020).
Effectiveness among Treatment Types
Chemical treatments were significantly more effective than mechanical or biological treatments for B. tectorum, more effective than fire treatments for P. ciliare, and more effective than mechanical treatments for Schismus spp. (Figure 4; Supplementary Table S2). While not significant for all species, mean effect sizes for chemical treatments were lower than for other treatment types for all six invasive grass species. This particular analysis does not capture the effects of treatment combinations, but these findings are consistent with our review in the “Treatment Characteristics” section, which found that biological, mechanical, and fire treatments were rarely effective alone and required combination with other treatments to effectively control invasive grasses.
Effect size (ln[amount in treated/control]) by species and treatment type. Only single treatment type, not combinations of treatment type, was included. Points and lines within box plots indicate mean and median values, respectively. Different letters indicate significant differences (P < 0.05) among species for a single treatment type. An asterisk (*) under a bracket indicates a significant difference (P < 0.05) among treatments within a single species. All species × treatment combinations had effect sizes significantly less than zero, except for Bio and Fire treatments for Bromus tectorum, Chem treatment for Eragrostis lehmanniana, and Bio treatment for Taeniatherum caput-medusae. Category abbreviations: Bio, Biological; Chem, chemical; Fire, prescribed fire; Graze, livestock grazing; Mech, mechanical; Seed, seeding; Soil, soil.

Studies on chemical treatments demonstrated opportunities to increase effectiveness by comparing herbicide types (Monaco et al. Reference Monaco, Osmond and Dewey2005; Shive et al. Reference Shive, Kuenzi, Sieg and Fulé2013; Young Reference Young2004), rates (Harvey et al. Reference Harvey, Simanonok, Rew, Prather and Mangold2020; Hirsch-Schantz et al. Reference Hirsch-Schantz, Monaco, Call and Sheley2014; Roldan-Nicolau et al. Reference Roldan-Nicolau, Terrazas, Montesinos, Pi and Tinoco-Ojanguren2020), and timing or method of application (Burnett and Mealor Reference Burnett and Mealor2015; Hirsch-Schantz et al. Reference Hirsch-Schantz, Monaco, Call and Sheley2014; Kyser et al. Reference Kyser, Peterson, Davy and DiTomaso2012; Sebastian et al. Reference Sebastian, Nissen and Rodrigues2016; Sheley et al. Reference Sheley, Bingham and Davies2012a), but also acknowledged the trade-off of having a negative effect on native and seeded species. Preemergence herbicides such as imazapic or indaziflam are, in general, very effective at controlling invasive grass and can potentially control invasives without significant effects on native and seeded species, as they are applied while perennial herbaceous species are dormant (Sebastian et al. Reference Sebastian, Nissen, Sebastian and Beck2017; Seedorf et al. Reference Seedorf, Clark and Nissen2022). However, studies assessing their effects on non-target species had mixed results, with some studies finding negative impacts on native and seeded grass and forb species, especially shortly following herbicide application (Baker et al. Reference Baker, Garner and Lyon2009; Brisbin et al. Reference Brisbin, Thode, Brooks and Weber2013; Elseroad and Rudd Reference Elseroad and Rudd2011; Mangold et al. Reference Mangold, Orloff, Parkinson and Halstvedt2015; Sheley et al. Reference Sheley, Carpinelli and Morghan2007; Terry et al. Reference Terry, Madsen, Gill, Anderson and St Clair2021a).
Several studies explored strategies to minimize herbicide impact to native species, and even promote their recovery, in invaded sites (Beck et al. Reference Beck, Sebastian and Chapman1995; Davies et al. Reference Davies, Boyd, Johnson, Nafus and Madsen2015b; Horn and St Clair Reference Horn and St Clair2017; Shive et al. Reference Shive, Kuenzi, Sieg and Fulé2013; Tjelmeland et al. Reference Tjelmeland, Fulbright and Lloyd-Reilley2008). For example, some studies found that lower rates of imazapic were suitably effective at decreasing invasive grasses while preventing excessive damage to native and seeded species (Burnett and Mealor Reference Burnett and Mealor2015; Kyser et al. Reference Kyser, DiTomaso, Doran, Orloff, Wilson, Lancaster, Lile and Porath2007; Rinella et al. Reference Rinella, Bellows, Davy, Forero, Hatler and James2021) and that seeding native species a year following herbicide application prevented detrimental effects (Davies et al. Reference Davies, Madsen, Nafus, Boyd and Johnson2014; Munson et al. Reference Munson, Long, Decker, Johnson, Walsh and Miller2015). Seeding techniques such as furrows, seeding below the active herbicide zone, and activated carbon seed coatings also mediated preemergence herbicide effects on seeded species (Terry et al. Reference Terry, Madsen, Gill, Anderson and St Clair2021b).
Mechanical, grazing, fire, and physical soil treatments were often resource intensive or created more disturbance that enhanced invasive grass growth (Rhodes et al. Reference Rhodes, Plowes and Gilbert2023; Rowe et al. Reference Rowe, Sprague, Fastiggi and Staker2023); and biological and seeding treatments were rarely effective alone (Germino and Lazarus Reference Germino and Lazarus2020; Owen et al. Reference Owen, Sieg and Gehring2011; Pyke et al. Reference Pyke, Shaff, Gregg and Conley2020; Reinhart et al. Reference Reinhart, Carlson, Feris, Germino, Jandreau, Lazarus, Mangold, Pellatz, Ramsey, Rinella and Valliant2020; Sheley et al. Reference Sheley, Vasquez, Chamberlain and Smith2012b; Tekiela Reference Tekiela2020; Uselman et al. Reference Uselman, Snyder, Leger and Duke2014). Variation in treatment combinations within species included as few as 4 unique combinations of treatment types (Schismus spp.) to as many as 26 (B. tectorum) (Supplementary Figure S1). Overall, combining treatment types was generally more effective than any one treatment strategy (Brisbin et al. Reference Brisbin, Thode, Brooks and Weber2013; Calo et al. Reference Calo, Brause and Jones2012; Daehler and Goergen Reference Daehler and Goergen2005; Davies Reference Davies2010; Davies and Boyd Reference Davies and Boyd2018; Davies and Sheley Reference Davies and Sheley2011; Davies et al. Reference Davies, Bates, Boyd, O’Connor and Copeland2021a; Diamond et al. Reference Diamond, Call and Devoe2012; Hirsch-Schantz et al. Reference Hirsch-Schantz, Monaco, Call and Sheley2014; Johnston Reference Johnston2015; Jones et al. Reference Jones, Chambers, Board, Johnson and Blank2015; Kessler et al. Reference Kessler, Nissen, Meiman and Beck2015; Lehnhoff et al. Reference Lehnhoff, Rew, Mangold, Seipel and Ragen2019; Porensky et al. Reference Porensky, Perryman, Williamson, Madsen and Leger2018; Schantz et al. Reference Schantz, Sheley and Hardegree2019; Seedorf et al. Reference Seedorf, Clark and Nissen2022; Sheley et al. Reference Sheley, Bingham and Davies2012a; Terry et al. Reference Terry, Madsen, Gill, Anderson and St Clair2021b; Whitson and Koch Reference Whitson and Koch1998; Wolden and Stromberg Reference Wolden and Stromberg1997).
Effectiveness among Species
Chemical treatment was significantly more effective, on average, at reducing T. caput-medusae compared with B. tectorum (Figure 4; Supplementary Table S2). This may have been due to environmental context, as T. caput-medusae tends to have a later phenology and to be more spatially constrained than B. tectorum (Bradley Reference Bradley2013), although there was a large amount of variation in treatment effects on both species overall and the median effects were similar. Conversely, prescribed fire treatments were significantly more effective for decreasing T. caput-medusae compared with B. tectorum and E. lehmanniana (Figure 4). Prescribed fire alone had a neutral effect on B. tectorum, or the invasive grass recovered to untreated levels or higher within the first few years following treatment (Davies Reference Davies2010; Davies and Dean Reference Davies and Dean2019; Jones et al. Reference Jones, Chambers, Board, Johnson and Blank2015; Pyke et al. Reference Pyke, Shaff, Lindgren, Schupp, Doescher, Chambers, Burnham and Huso2014; Roundy et al. Reference Roundy, Chambers, Pyke, Miller, Tausch, Schupp, Rau and Gruell2018; Schacht and Stubbendieck Reference Schacht and Stubbendieck1985; Vermeire et al. Reference Vermeire, Rinella and Strong2021). On the other hand, spring or summer burns very effectively controlled T. caput-medusae in warm-winter climates, likely driving the significant difference from B. tectorum in this analysis, but were less effective in cold-winter climates (Kyser et al. Reference Kyser, Doran, McDougald, Orloff, Vargas, Wilson and DiTomaso2008). While other studies observed some control in the first few years following fire treatments, they noted eventual increases in T. caput-medusae similar to that of B. tectorum over time (Davies and Dean Reference Davies and Dean2019; Davy and Dykier Reference Davy and Dykier2017). Studies that combined fire as a treatment alongside supplementary treatments such as seeding, grazing, or herbicide were typically more effective than using fire alone (Davies and Boyd Reference Davies and Boyd2018; Diamond et al. Reference Diamond, Call and Devoe2012).
Time since Treatment
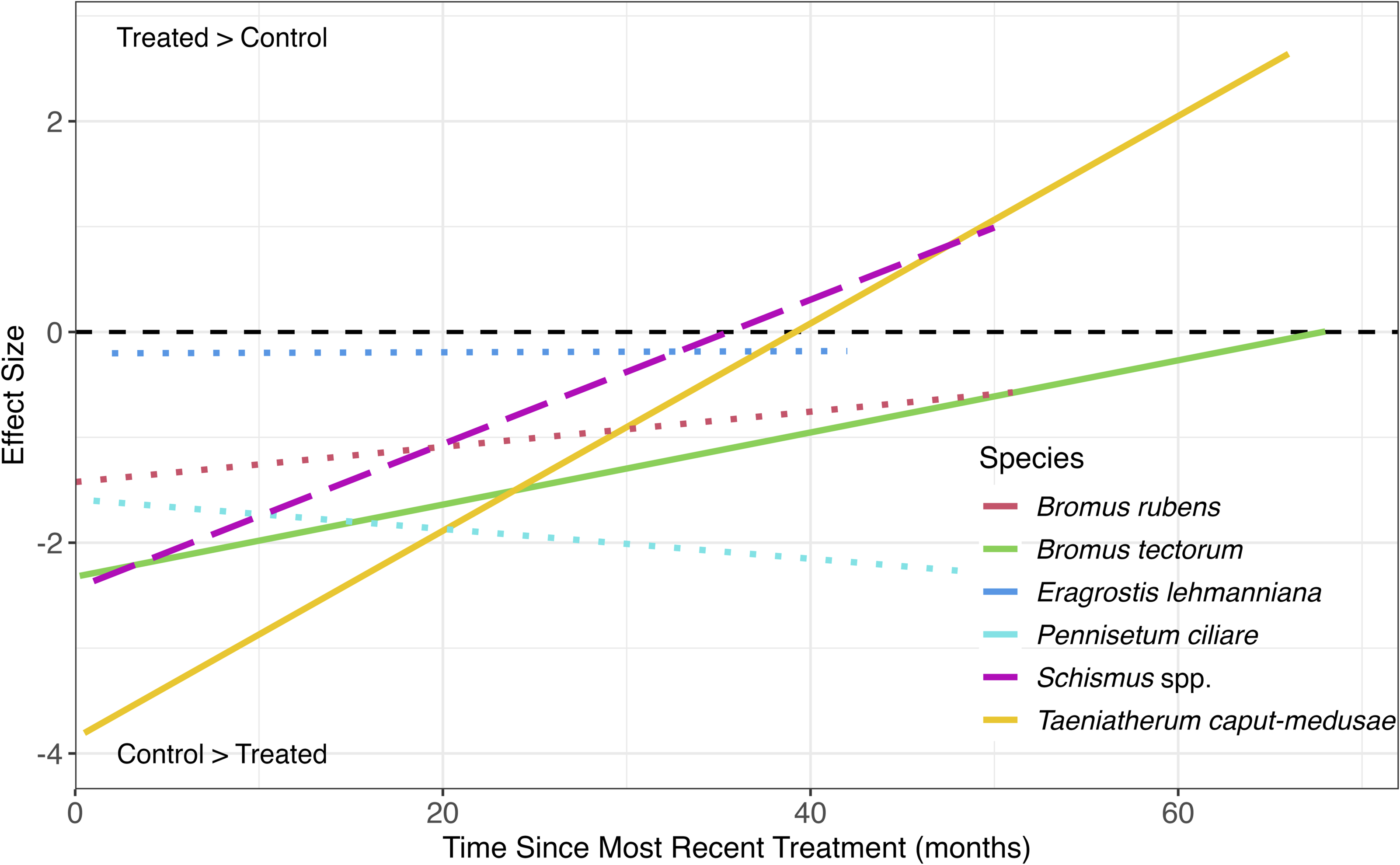
Treatments intended to reduce T. caput-medusae were most effective compared with other invasive grasses immediately after treatment. This effect diminished through time such that invasive grasses in treated areas matched control areas within 40 mo (Figure 5). The effect size immediately after treatment for B. tectorum was less than that of T. caput-medusae, but the treatment effectiveness decreased less rapidly to reach control levels within 60 mo. There was a trend for decreasing treatment effectiveness over time for Schismus spp. (P < 0.1), while treatment effectiveness of the other invasive grass species did not change through time (Supplementary Table S2). Differences in the duration of treatment effectiveness among species suggest variation in the regeneration and persistence traits of individual invasive grass species, including the relatively fast regrowth of annual compared with perennial species. The size and severity of the invasions and underlying environmental conditions could also affect treatment longevity of different treatment types and could be considered in future treatment review efforts. Only about one-third of the 136 studies used in this portion of the review evaluated treatment effectiveness beyond 2 yr, and only 6 studies evaluated treatment effectiveness beyond 5 yr (Davies and Dean Reference Davies and Dean2019; Davies et al. Reference Davies, Bates, Boyd and Svejcar2016a; Kulmatiski Reference Kulmatiski2011; Pyke et al. Reference Pyke, Shaff, Chambers, Schupp, Newingham, Gray and Ellsworth2022; Stonecipher et al. Reference Stonecipher, Thacker, Welch, Ralphs and Monaco2019; Urza et al. Reference Urza, Weisberg, Chambers, Board and Flake2019). Longer and more frequent monitoring than occurred in previous studies can improve evaluations of treatment effectiveness, including using remote sensing (Bradley et al. Reference Bradley, Curtis, Fusco, Abatzoglou, Balch, Dadashi and Tuanmu2018; Dickson et al. Reference Dickson, Sisk, Sesnie and Bradley2015; James et al. Reference James, Gornish, DiTomaso, Davy, Doran, Becchetti, Lile, Brownsey and Laca2015; Mealor et al. Reference Mealor, Cox and Booth2012).
Treatment effect size (ln[amount in treated/control]) related to time since most recent treatment by invasive grass species indicated by linear regression lines. Statistical significance at P < 0.05 indicated by solid lines; trend at P < 0.1 indicated by dashed regression line; no statistical significance indicated by dotted lines. Black dashed line indicates effect size of zero, with values above the line indicating an increase in invasive grass following treatment and vice versa.

Repeat Treatments
Studies addressed diminishing treatment effectiveness by exploring whether repeat treatments caused a greater reduction of invasive grasses. Applying the same treatment three or more times was significantly more effective than a single-treatment application for P. ciliare, and treatment effectiveness significantly increased with treatment frequency for B. tectorum (Figure 6; Supplementary Table S2). Chemical, mechanical, grazing, and fire were the most commonly repeated treatments across all species. These results suggest the added benefit of investing more to increase the number of treatments and that a multiyear management strategy is more likely to lead to sustained reduction of invasive grass species. While herbicide application could be effective at reducing invasive grasses, there was often potential for reinvasion if the remaining grasses were not retreated (Baker et al. Reference Baker, Garner and Lyon2009; Farrell and Gornish Reference Farrell and Gornish2019; Vermeire et al. Reference Vermeire, Rinella and Strong2021) and the persistent seedbank was not targeted (Rowe et al. Reference Rowe, Sprague, Fastiggi and Staker2023; Sebastian et al. Reference Sebastian, Nissen, Sebastian and Beck2017). Similarly, treatments such as seeding native species or mowing could require multiple treatments to exert enough pressure to successfully suppress invasive grasses (Davies et al. Reference Davies, Boyd, Johnson, Nafus and Madsen2015b; Valliere et al. Reference Valliere, Balch, Bell, Contreras and Hilbig2019). Whether combining treatments to overcome barriers associated with a single treatment could reduce the need for treatment repetitions is worth further investigation.
Effect size (ln[amount in treated/control]) of applying the same treatment type 1, 2, or 3 + times for Bromus rubens, Bromus tectorum, Pennisetum ciliare, and Taeniatherum caput-medusae. Points and lines within box plots indicate mean and median values, respectively. Different letters indicate statistically significant (P < 0.05) differences among treatment times within each species.

Conclusions
Dominant invasive grass species in the Intermountain West increase the cover, density, and frequency of fuel and magnify the fire area, rate of spread, and flame length that collectively amplify wildfire risk. Changes in fuel and fire characteristics depend on the species, with warm-season perennial species causing high-severity fire and cool-season annuals generating fast-moving but low-severity fire. Chemical treatments were the most common and effective treatment type among all species, although other treatment types could be equally or more effective under certain management and environmental conditions. Despite their effectiveness, longevity of treatments was 3 to 5 yr, and in many cases less, and sustained reduction was dependent on repeat treatments for two of the six dominant invasive grass species. Our findings, based on comparisons across invasive grass species, ecoregions, and treatment types, provide unique insight on a wide range of fuel and fire metrics and the conditions under which treatments can be effective.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.1017/inp.2025.10037
Acknowledgments
We would like to thank Katie Laushman and Alex Croydon for their assistance in literature selection and data collection for this project. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Funding statement
This work was supported by the U.S. Geological Survey Restoration Assessment and Monitoring Program for the Southwest in the Ecosystems Mission Area.
Competing interests
The authors declare no conflicts of interest.








 Open access
Open access