Introduction
The increasing human pressure on biodiversity has led researchers to call the present period the Anthropocene (Dirzo et al. Reference Dirzo, Young, Galetti, Ceballos, Isaac and Collen2014; Steffen Reference Steffen2021). Anthropogenic changes have already deforested more than one-third of neotropical forests, and about 55% of the remaining forests have low or medium integrity (Grantham et al. Reference Grantham, Duncan, Evans, Jones, Beyer and Schuster2020). Habitat degradation promotes the local or global extinction of species (Ceballos et al. Reference Ceballos, Ehrlich and Dirzo2017, Reference Ceballos, Ehrlich and Raven2020; Develey and Phalan Reference Develey and Phalan2021) and their ecological functions (Galetti et al. Reference Galetti, Guevara, Côrtes, Fadini, von Matter and Leite2013; Magioli et al. Reference Magioli, Ferraz, Chiarello, Galetti, Setz and Paglia2021), and consequently, negatively impacts various ecosystem services provided by natural areas and resources, such as pest control (Bianchi et al. Reference Bianchi, Booij and Tscharntke2006), water provision (Dodds et al. Reference Dodds, Perkin and Gerken2013), and carbon storage (Molotoks et al. Reference Molotoks, Stehfest, Doelman, Albanito, Fitton and Dawson2018), in addition to increasing exposure to infectious diseases (Barbier Reference Barbier2021; Wilkinson et al. Reference Wilkinson, Marshall, French and Hayman2018). However, extinctions do not occur randomly, and species with certain morphological and functional characteristics are more affected by habitat loss than others (Dirzo et al. Reference Dirzo, Young, Galetti, Ceballos, Isaac and Collen2014). Forest species with larger body sizes and frugivorous diets are more prone to disappearing in degraded areas (Bogoni et al. Reference Bogoni, Pires, Graipel, Peroni and Peres2018; Burivalova et al. Reference Burivalova, Lee, Giam, Sekercioglu, Wilcove and Koh2015; Coelho et al. Reference Coelho, Raniero, Silva and Hasui2016).
Species in the family Cracidae are large birds (~63% weighing over 1 kg; Dunning Reference Dunning2007) and fruit consumers (Wilman et al. Reference Wilman, Belmaker, Simpson, Rosa, Rivadeneira and Jetz2014), suffering from both habitat loss and hunting (Brooks Reference Brooks2006). Of the 56 species distributed across the Americas, 22 are included in some threat category on the global Red List (IUCN 2025), making it the most threatened bird family on the continent (Brooks Reference Brooks2006). The Black-fronted Piping-Guan Pipile jacutinga is a forest species endemic to a heavily exploited biome, the Atlantic Forest (Ribeiro et al. Reference Ribeiro, Metzger, Martensen, Ponzoni and Hirota2009; Rosa et al. Reference Rosa, Brancalion, Crouzeilles, Tambosi, Piffer and Lenti2021; Vale et al. Reference Vale, Tourinho, Lorini, Rajão and Figueiredo2018). It is nationally and globally threatened (‘Endangered’; Birdlife International 2018; MMA 2022), weighs about 1.25 kg (Dunning Reference Dunning2007), and has a strong relationship with the palm heart Euterpe edulis (Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997), which plays an important ecological, economic, and social role in the Atlantic Forest (Chagas et al. Reference Chagas, Salk, Vidal, Souza and Brancalion2021; Sales et al. Reference Sales, Kissling, Galetti, Naimi and Pires2021; Trevisan et al. Reference Trevisan, Fantini, Schmitt-Filho and Farley2015). For E. edulis and at least 51 other plant species (Bello et al. Reference Bello, Galetti, Montan, Pizo, Mariguela and Culot2017), the Black-fronted Piping-Guan is an important disperser of large seeds (Galetti et al. Reference Galetti, Guevara, Côrtes, Fadini, von Matter and Leite2013) and its disappearance, along with other large frugivores, can negatively impact seed dispersal in defaunated areas (Bovo et al. Reference Bovo, Ferraz, Magioli, Alexandrino, Hasui and Ribeiro2018; Galetti et al. Reference Galetti, Guevara, Côrtes, Fadini, von Matter and Leite2013).
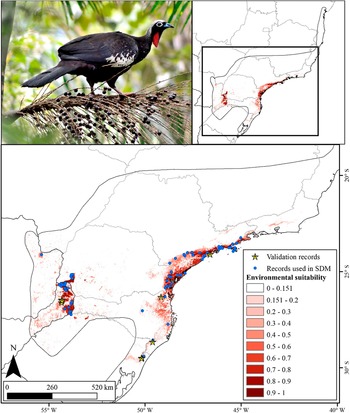
Originally, the Black-fronted Piping-Guan inhabited the entire Atlantic Forest south of Bahia state, up to Paraguay and Argentina (BirdLife International 2018) (Figure 1), but due to habitat loss and hunting (Silveira et al. Reference Silveira, Soares and Bianchi2008) it is currently restricted to the largest and isolated forest remnants in the states of São Paulo, Paraná, Santa Catarina, Rio Grande do Sul, Paraguay, and Argentina (Bernardo et al. Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011; Cockle and Bodrati Reference Cockle and Bodrati2011; Esquivel et al. Reference Esquivel, Zarza, Tiffer-Sotomayor, Díaz, Pérez and Velázquez2019). Natural populations of the species have become extinct in the states of Bahia (SEMA 2017), Espírito Santo (Chaves et al. Reference Chaves, Duca, Pinto, Rosa, Magnago, Filho, Fraga, Formigoni and Chaves2019), Rio de Janeiro (Bergallo et al. Reference Bergallo, Rocha, Alves and van2000), and Minas Gerais (Leal et al. Reference Leal, Marques and Silva2018), and even in the states and countries where it still occurs, there are several cases of local extinctions (Silveira et al. Reference Silveira, Soares and Bianchi2008), especially in semi-deciduous seasonal forests. In Paraguay, the species was recorded in at least 16 locations in the 1990s but currently survives only in the Mbaracayú Forest Nature Reserve (Esquivel et al. Reference Esquivel, Zarza, Tiffer-Sotomayor, Díaz, Pérez and Velázquez2019; Hugo Castillo 2021, personal communication). In Argentina, there are recent records only in the northern and central portions of Misiones (Cockle and Bodrati Reference Cockle and Bodrati2011; eBird 2025).
Historical and current distribution (following BirdLife International 2018), and records of the Black-fronted Piping-Guan (recent and historical). The historical records were extracted from Portal da Biodiversidade (ICMBio 2021).
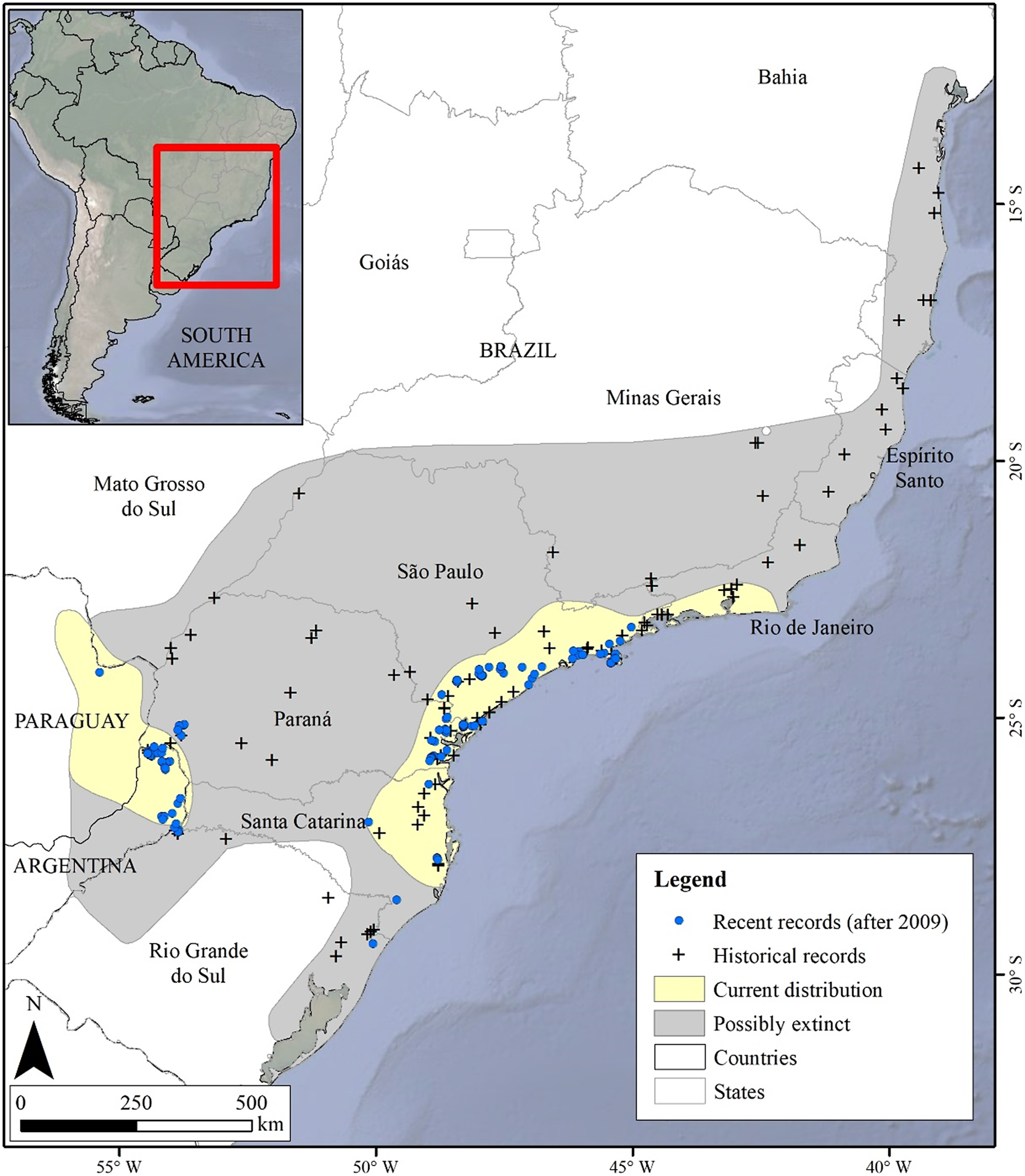
Figure 1. Long description
Inset at top left shows South America with a red box highlighting southeastern Brazil. The main map covers parts of Brazil, Paraguay, and Argentina, with state and country borders labeled. The legend at lower right defines symbols: blue dots for recent records (after 2009), black crosses for historical records, pale yellow for current distribution, and gray for possibly extinct areas. Recent records cluster in southern São Paulo, Paraná, Santa Catarina, and eastern Paraguay. Historical records are widely scattered, including Espírito Santo, Minas Gerais, and Rio Grande do Sul. The current distribution (yellow) forms a fragmented band from eastern Paraguay through southern Brazil, mainly along the Atlantic Forest. Gray shading marks areas where the species is possibly extinct, extending northward and inland from the current range. Scale bar at bottom left indicates distances up to 500 kilometers. Cardinal directions are marked with north at bottom left.
The impacts suffered by the species and the contraction of its distribution area have led to the evaluation of various strategies to assist in its conservation, such as translocation, specifically reintroduction (Phalan et al. Reference Phalan, Barbosa, Bernardo, Bosso, Chaves and Corrêa2020; Sick Reference Sick1997; Silveira et al. Reference Silveira, Soares and Bianchi2008), defined as “the intentional movement and release of an organism within its native range from which it has disappeared” (IUCN/SSC 2013). The Black-fronted Piping-Guan has been successfully bred in captivity over the past decades, and it is estimated that there are about 200 individuals in ex situ institutions in Brazil (Phalan et al. Reference Phalan, Barbosa, Bernardo, Bosso, Chaves and Corrêa2020). This fact, along with a large number of experiences with reintroductions of the species itself and other Cracidae (Angulo and Barrio Reference Angulo and Barrio2004; Bernardo et al. Reference Bernardo, Desbiez, Olmos and Collar2014; Pereira and Wajntal Reference Pereira and Wajntal1999; Souza L.D.C. et al. Reference Souza, Pereira, Azevedo, Bernardo, Develey and Ruiz-Miranda2020), can help increase the likelihood of success of future reintroductions (World Pheasant Association and IUCN/SSC Re-Introduction Specialist Group 2009). Besides benefiting the Black-fronted Piping-Guan itself, the translocation of a large frugivore can be important for restoring the complexity of seed dispersal in defaunated areas (Carvalho et al. Reference Carvalho, Lucas and Côrtes2021), being a first step in the recovery of ecological processes (Ghazoul and Chazdon Reference Ghazoul and Chazdon2017).
Here, we used two tools to identify the most suitable areas for the conservation and potential translocation of the Black-fronted Piping-Guan: Species Distribution Modelling (SDM) (Guisan et al. Reference Guisan, Thuiller and Zimmermann2017; Peterson et al. Reference Peterson, Soberón, Pearson, Anderson, Martínez-Meyer and Nakamura2011) and citizen science (Haklay et al. Reference Haklay, Dörler, Heigl, Manzoni, Hecker, Vohland, Vohland, Land-Zandstra, Ceccaroni, Lemmens, Perelló and Ponti2021). Based on presence records (points) and environmental variables, SDM produces a map of environmental suitability, indicating where the areas with the highest probabilities of species occurrence are located (Peterson et al. Reference Peterson, Soberón, Pearson, Anderson, Martínez-Meyer and Nakamura2011). The application of SDM to assist in species conservation has aided threatened species and can be the first step in planning the translocation of the species considering its entire original range (Anoop et al. Reference Anoop, Babu, Nagarajan and Sen2020; Elia et al. Reference Elia, Haig, Johnson, Marcot and Young2015; Martínez-Meyer et al. Reference Martínez-Meyer, Peterson, Servín and Kiff2006). However, presence data with high spatial accuracy are scarce for many species (Lees et al. Reference Lees, Rosenberg, Ruiz-Gutierrez, Marsden, Schulenberg and Rodewald2020), and suitable data from citizen science platforms can be very useful for building reliable models (i.e. selecting only data that meet certain criteria; Steen et al. Reference Steen, Elphick and Tingley2019). Thus, we aimed in this study to (1) update the potential distribution of the Black-fronted Piping-Guan, (2) validate the SDM by using new records, and (3) identify potentially suitable areas for translocation.
Methods
Study area
The Atlantic Forest is a tropical rainforest biome extending for 1.4 million km² along the Brazilian coast, reaching Paraguay and Argentina in its southern portion. In Brazil and Paraguay, the biome was severely replaced by anthropogenic land use and cover, with only 12–28% and 9% of its original vegetation remaining, respectively (Ponte et al. Reference Ponte, Mack, Wohlfart, Rodas, Fleckenstein and Oppelt2017; Rezende et al. Reference Rezende, Scarano, Assad, Joly, Metzger and Strassburg2018; Ribeiro et al. Reference Ribeiro, Metzger, Martensen, Ponzoni and Hirota2009). In Argentina, forest cover is high compared with neighbouring countries (60%), but the three countries have faced an historic loss of mature forest in the last decades (Projeto MapBiomas Mata Atlântica Trinacional 2021; Rosa et al. Reference Rosa, Brancalion, Crouzeilles, Tambosi, Piffer and Lenti2021). The Atlantic Forest houses 891 bird species, of which 223 (25%) are endemic (Vale et al. Reference Vale, Tourinho, Lorini, Rajão and Figueiredo2018), and 10% of the taxa are threatened (MMA 2022). Due to its rich biodiversity, the biome is considered a hotspot (Mittermeier and Rylands Reference Mittermeier, Rylands, Dellasal and Goldstein2018), but habitat loss and other anthropogenic impacts have already caused extinction and functional loss (Develey and Phalan Reference Develey and Phalan2021; Pereira et al. Reference Pereira, Dantas, Silveira, Roda, Albano and Sonntag2014). The area used for modelling the Black-fronted Piping-Guan was the geographical range proposed by the International Union for Conservation of Nature (IUCN), i.e. 1,062,683 km² (BirdLife International 2018) (Figure 1), and was selected by including all the recent presence records and all the areas with potential for conservation and translocations.
Black-fronted Piping-Guan records
We used only presence records from 2009 to 2021, gathered from the literature, researchers, and citizen science platforms, excluding records made before 2008 (hereafter referred to as ‘historic records’) since they cannot represent the environments where the species occurs nowadays due to local extinctions. We conducted a literature search by using the species’ scientific names ‘Aburria jacutinga’ and ‘Pipile jacutinga’ in Web of Science, Google Scholar, and, for a comprehensive and efficient search, the Brazilian Library of Theses and Dissertations, the only local online database of this sort in July 2021. We also personally contacted researchers who used camera traps to study mammals to share with us opportunistic records of the species. We gathered presence records from two online platforms to fill gaps in the original occurrence area. In February 2019, we sent individual requests to 624 WikiAves (wikiaves.com.br) users who had contributed photographs or audio recordings of the Black-fronted Piping-Guan and used eBird (ebird.org) to search for records where the users share the exact location obtained with a GPS device (confirmed by personal communication). We gathered 207 recent records of Black-fronted Piping-Guan, from which 143 (69%) were from citizen scientists.
To avoid spatial dependence between records we applied a rarefaction using 7 km distance, defined from the maximum individual movement recorded in fieldwork focused on the species (Alecsandra Tassoni 2021, personal communication). We used the tool Spatially Rarefy Occurrence Data for SDMs, from SDMtoolbox v2.4 package (Brown et al. Reference Brown, Bennett and French2017) in ArcGIS 10.3 (ESRI 2014), selecting 60 points for modelling.
After building the model, we used the same method to search for records from 2021 to 2024 in areas with no records in our former database. The data set of 10 records gathered were used to validate our model by extracting their suitability value.
Environmental data
We used topographic (Danielson and Gesch Reference Danielson and Gesch2011), bioclimatic (Fick and Hijmans Reference Fick and Hijmans2017), and land cover variables (Hansen et al. Reference Hansen, Potapov, Moore, Hancher, Turubanova and Tyukavina2013; Souza C.M. et al. Reference Souza, Shimbo, Rosa, Parente, Alencar and Rudorff2020) (see Supplementary material S1). The land cover variable available from MapBiomas is categorical, consisting of various land cover classes. Therefore, we selected only the pixels classified as ‘Forest Formation’ (class 3) and used the Focal Statistics tool within ArcGIS 10.3 (ESRI 2014) with a moving square window of 33 × 33 pixels to generate a ‘percentage of forest cover’ variable. Forest formation classification in MapBiomas is defined as “Forest Formation includes natural forest (excludes Forest Plantation) areas of more than 0.5 ha with trees with a minimum height of 5 m and tree canopy cover that varies for each type of original forest formation (60 to 80%)” (Projeto MapBiomas Mata Atlântica Trinacional 2021). For two small sections within the modelled area but not mapped by MapBiomas, we used the forest vegetation percentage map (Hansen et al. Reference Hansen, Potapov, Moore, Hancher, Turubanova and Tyukavina2013) to complement the forest cover percentage layer.
To avoid including correlated variables in the model, we conducted a Pearson correlation analysis and excluded variables with values greater than 0.7 or less than -0.7 (Dormann et al. Reference Dormann, Elith, Bacher, Buchmann, Carl and Carré2013), resulting in eight variables available for model construction (Table 1). All variables were resampled to a resolution of 1 km and clipped according to the species distribution (BirdLife International 2018).
Variables used in the Black-fronted Pipin-Guan distribution model
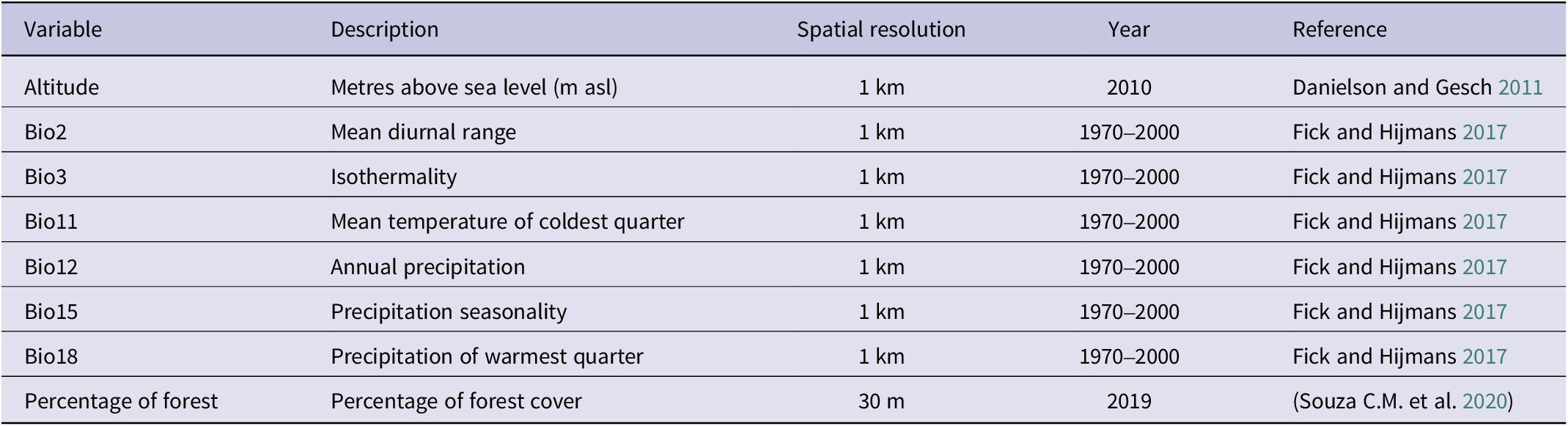
Table 1. Long description
The table contains eight columns labeled Variable, Description, Spatial resolution, Year, and Reference. From top to bottom, the variables are Altitude, Bio2, Bio3, Bio11, Bio12, Bio15, Bio18, and Percentage of forest. Altitude is described as metres above sea level with a spatial resolution of 1 km, year 2010, and reference Danielson and Gesch 2011. Bio2 is mean diurnal range, 1 km, 1970 to 2000, Fick and Hijmans 2017. Bio3 is isothermality, 1 km, 1970 to 2000, Fick and Hijmans 2017. Bio11 is mean temperature of coldest quarter, 1 km, 1970 to 2000, Fick and Hijmans 2017. Bio12 is annual precipitation, 1 km, 1970 to 2000, Fick and Hijmans 2017. Bio15 is precipitation seasonality, 1 km, 1970 to 2000, Fick and Hijmans 2017. Bio18 is precipitation of warmest quarter, 1 km, 1970 to 2000, Fick and Hijmans 2017. Percentage of forest is percentage of forest cover, 30 m, 2019, Souza C M et al. 2020.
Species Distribution Modelling
We used the maxent algorithm within the Maxent software version 3.4.4 (Phillips and Dudík Reference Phillips and Dudík2008; Phillips et al. Reference Phillips, Anderson and Schapire2006, Reference Phillips, Anderson, Dudík, Schapire and Blair2017a, Reference Phillips, Dudik and Schapireb) due to its strong performance (Gobeyn et al. Reference Gobeyn, Mouton, Cord, Kaim, Volk and Goethals2019; Qiao et al. Reference Qiao, Soberón and Peterson2015) and positive results obtained in applications aimed at species conservation (Bovo et al. Reference Bovo, Ferraz, Ribeiro, Lins, Barbosa and Previdente2021; Ferraz et al. Reference Ferraz, Oliveira, Attias and Desbiez2021; Morato et al. Reference Morato, Ferraz, Paula and Campos2014). The model was generated from 10 repetitions, using the bootstrap resampling method, with 10,000 pseudoabsence points, 30% of the data for testing, random seed, convergence threshold of 10^-5, and a maximum of 500 iterations. The model was evaluated based on omission errors (false negative or Type I error) and overprediction (false positive or Type II error). We also used two measures based on sensitivity (the model’s ability to correctly predict presences) and specificity (the model’s ability to correctly predict pseudoabsences): area under the ROC curve (AUC) and True Skill Statistics (TSS). AUC measures the area under the receiver operating characteristic (ROC) curve, constructed from false positive and true positive values (Fielding and Bell Reference Fielding and Bell1997). It ranges from 0 to 1, indicating a good model for values between 0.7 and 0.9, and an excellent model above 0.9 (Peterson et al. Reference Peterson, Soberón, Pearson, Anderson, Martínez-Meyer and Nakamura2011). TSS is calculated by the formula: sensitivity + specificity - 1 (Allouche et al. Reference Allouche, Tsoar and Kadmon2006). Its values range from -1 to +1, being acceptable above 0.4 and ‘almost perfect’ above 0.8 (Landis and Koch Reference Landis and Koch1977).
We cut the final average model using the ‘Maximum test sensitivity plus specificity Cloglog threshold’ (= 0.151) to transform it into a binary model (suitable or unsuitable areas). Pixels with values above this threshold are considered suitable, while pixels with lower values are considered unsuitable. We chose this threshold because it is minimally affected by the number of presence records (Liu et al. Reference Liu, Newell and White2016), performs well when only presence data are used (Liu et al. Reference Liu, White and Newell2013), and has satisfactory omission and overprediction error rates.
To calculate the minimum area needed to sustain a long-term population of the species, we used the average density of the species from Bernardo et al. (Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011): 1.51 individuals/km². We excluded values from coastal islands (Ilha do Cardoso and Ilhabela) as they represent much higher densities than other continental areas. There are no studies exploring the minimum viable population for Black-fronted Piping-Guan or other species within the genus. Therefore, we relied on available values for the Red-billed Curassow Crax blumembachii, a very similar species with available information, i.e. belongs to the same family, frugivorous, and endemic to the Atlantic Forest. We used the value of 138 individuals, representing a minimum viable population in a more conservative scenario (Bonfim et al. Reference Bonfim, Cordeiro, Peres, Canale and Bernardo2019). By multiplying the density values by the minimum viable population, we found an area of 208 km², which we considered the minimum area capable of sustaining a Black-fronted Piping-Guan population. Thus, we identified suitable areas larger than this threshold as environmentally adequate for the reintroduction. We then examined how these related to Strictly Protected Areas, defined here as those in IUCN Categories I–III (Dudley Reference Dudley2008), to quantify areas with adequate protection for the species.
Results
The potential distribution model for the Black-fronted Piping-Guan showed excellent statistical performance (AUC = 0.973 ± 0.006 and TSS = 0.876 ± 0.071; Figure 2), with high sensitivity (94%) and high specificity (93%). Forest cover percentage (70.9%) was the most important variable in explaining the probability of species presence, with an increase in forest cover leading to a higher probability of species presence (S2 and S3). The area potentially suitable for the species was 73,802 km² (6.9% of the original distribution; Figure 2), mainly concentrated in two large blocks: the region near the coast from Rio de Janeiro to Rio Grande do Sul (highlighted in Figure 3), and the region including Foz do Iguaçu, Argentina, and Paraguay (highlighted in Figure 4). All areas with recent records used in the modelling process was indicated as suitable.
Black-fronted Piping-Guan distribution model cut by the Maximum test sensitivity plus specificity Cloglog threshold (0.151). The suitable areas are shown in different shades of red.
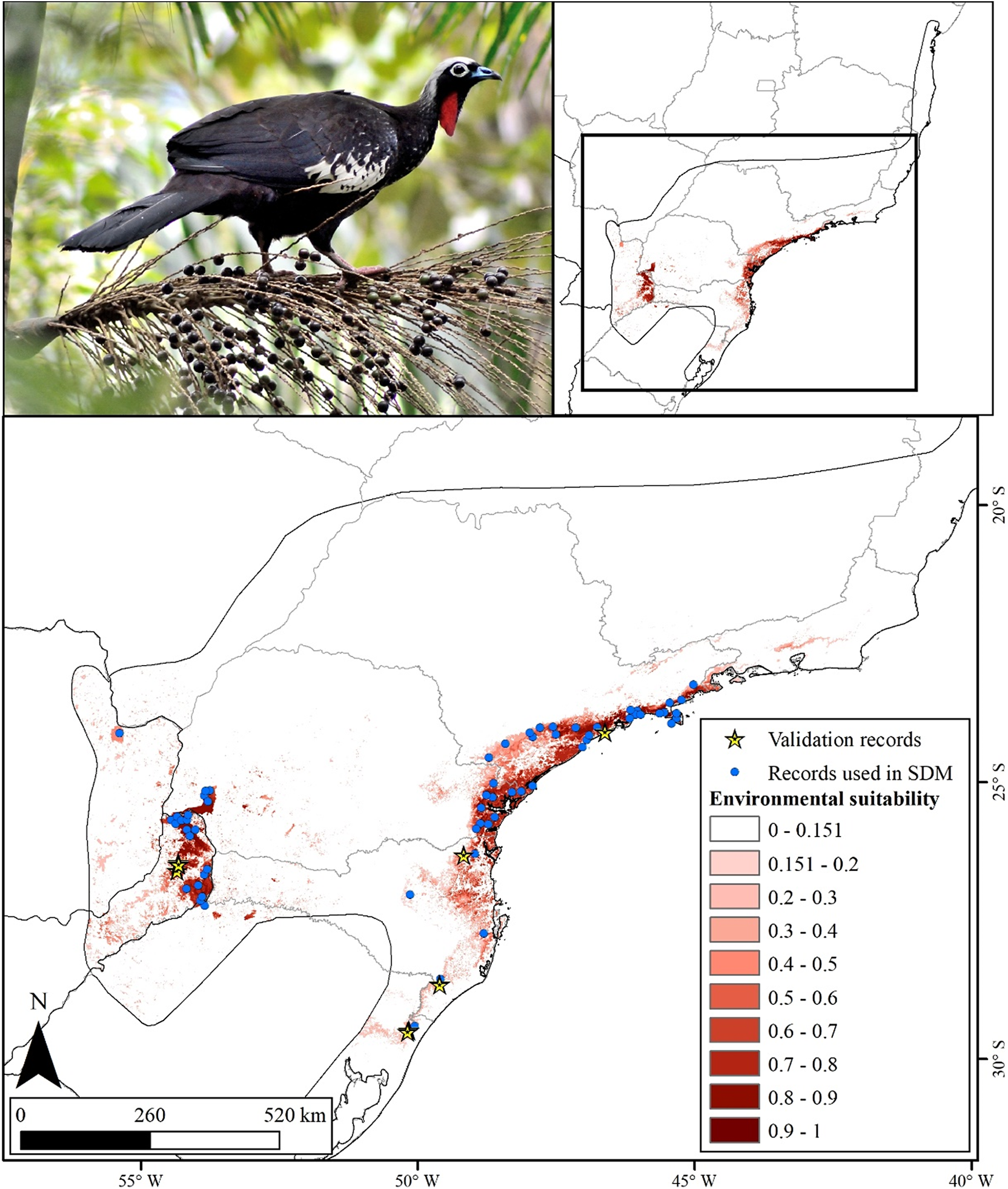
Figure 2. Long description
The upper left panel displays a color photograph of a Black-fronted Piping-Guan perched on a palm branch. The upper right panel contains a simplified outline map of eastern South America with a black rectangle highlighting the study region. The main lower panel is a detailed map of southeastern Brazil, oriented with north at the bottom left. Environmental suitability is represented by a gradient from white to dark red, with white indicating low suitability and dark red indicating high suitability. Suitability classes are labeled in the legend as 0 to 0.151, 0.151 to 0.2, 0.2 to 0.3, 0.3 to 0.4, 0.4 to 0.5, 0.5 to 0.6, 0.6 to 0.7, 0.7 to 0.8, 0.8 to 0.9, and 0.9 to 1. The highest suitability areas (darkest red) are concentrated in the southeast, especially along the Atlantic Forest corridor. Blue dots mark records used in species distribution modeling, and yellow stars indicate validation records. The scale bar at the bottom left shows distances of 0, 260, and 520 kilometers. Latitude and longitude lines are labeled along the map edges.
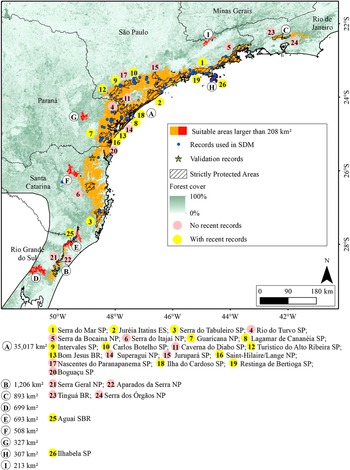
Macro region of Serra do Mar with indications of the suitable areas for the Black-fronted Piping-Guan and the Protected Areas. The letters are ordinated from the larger to smaller area and the use of different colours for the areas are just to provide a better visualisation. SP: State Park. ES: Ecological Station. NP: National Park. BR: Biological Reserve. SBR: State Biological Reserve.
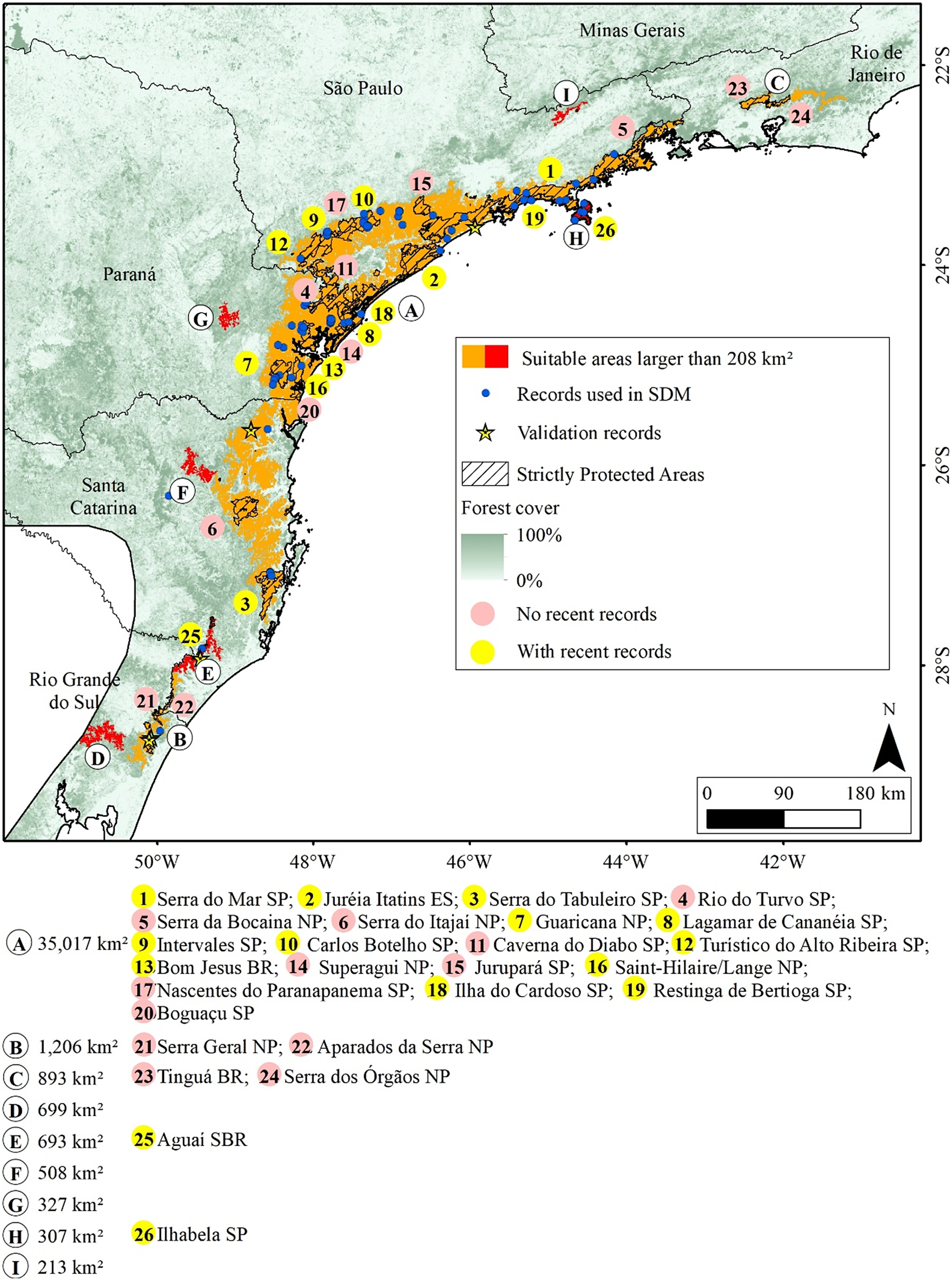
Figure 3. Long description
This is a color-coded regional map of southeastern Brazil, anchored at São Paulo in the northwest and extending southeast along the Serra do Mar mountain range through Paraná, Santa Catarina, and ending in Rio Grande do Sul. Red polygons indicate suitable areas larger than 208 square kilometers for the Black-fronted Piping-Guan, concentrated along the Serra do Mar corridor. Blue dots mark records used in species distribution modeling, with yellow stars for validation records. Hatched overlays denote strictly protected areas. Forest cover is shaded from dark green (100 percent) to white (0 percent). Pink circles indicate locations with no recent records, while yellow circles show areas with recent records. Twenty-six protected areas are labeled with numbers inside colored circles, ordered by area size and detailed in the legend. Major parks and reserves include Serra do Mar S P, Juréia Itatins E S, Serra do Tabuleiro S P, and others, with their respective area sizes and types (S P for State Park, E S for Ecological Station, N P for National Park, B R for Biological Reserve, S B R for State Biological Reserve) listed below the map. The legend clarifies all symbols and color codes. A north arrow and scale bar are present in the lower right.
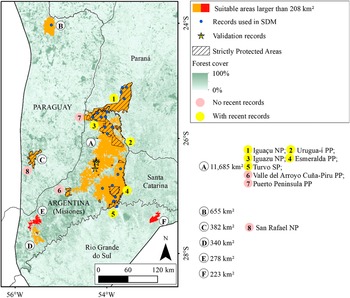
Macro region of Misiones with indications of suitable areas for the Black-fronted Piping-Guan and the Protected Areas. The letters are ordinated from the larger to smaller area and the use of different colours for the areas are just to provide a better visualisation. SP: State Park. PP: Provincial Park. NP: National Park.
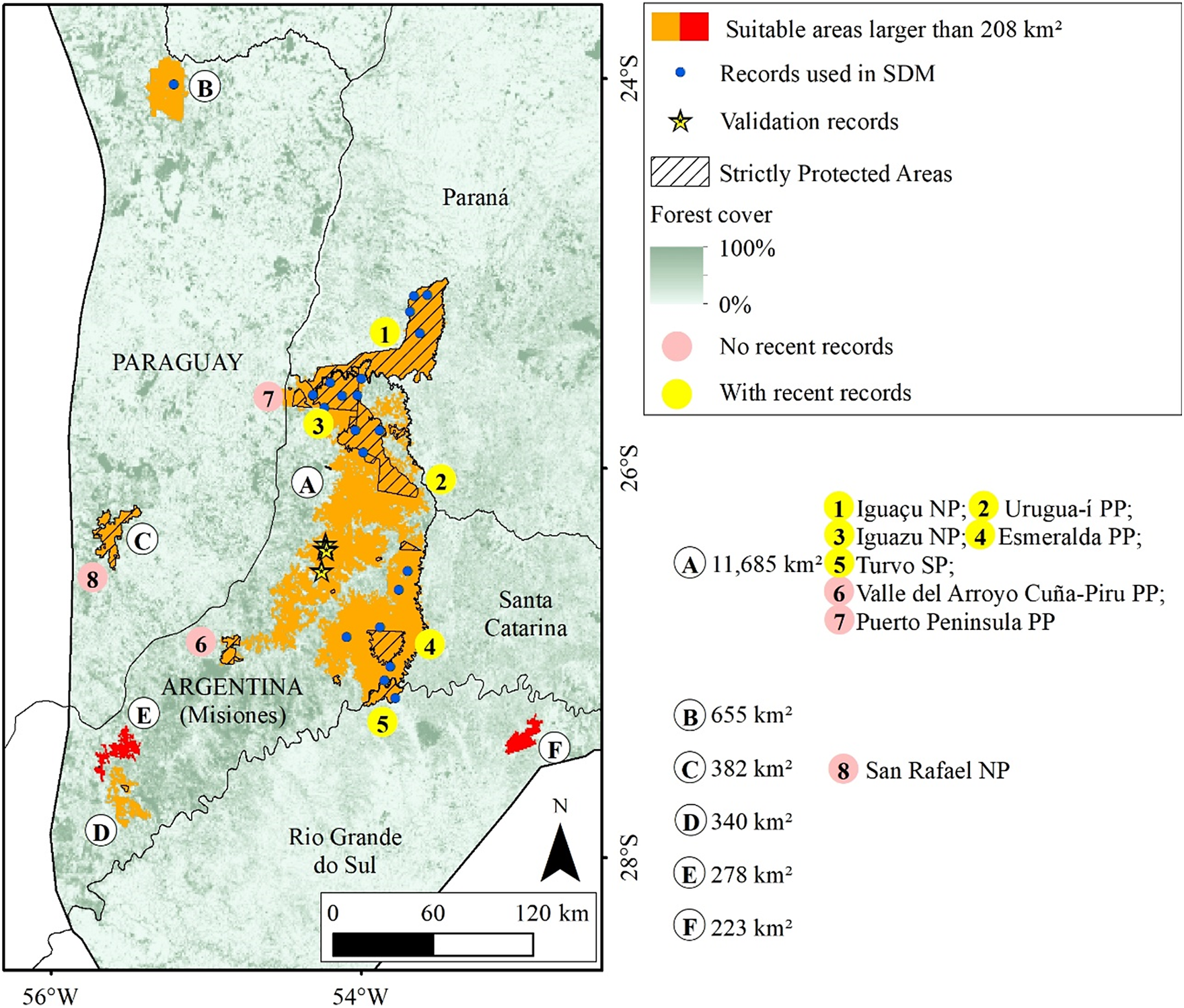
Figure 4. Long description
Starting at the center, area A (11,685 km super 2) covers central Misiones, marked in orange for high suitability, with hatched overlays indicating strictly protected areas. Blue dots within A show S D M records, and yellow stars indicate validation records. Yellow circles with numbers 1 to 5 highlight Iguaçu N P, Urugua-i P P, Iguazu N P, Esmeralda P P, and Turvo S P, all with recent records. Pink circles 6 and 7 mark Valle del Arroyo Cuña-Piru P P and Puerto Peninsula P P, with no recent records. To the northwest, area B (655 km super 2) is also suitable. Southwest lies area C (382 km super 2), and further south are D (340 km super 2), E (278 km super 2), and F (223 km super 2), each shown in red. Area 8, San Rafael N P, is in the west, marked with a pink circle. Forest cover is shaded from dark green (100 percent) to white (0 percent). The legend clarifies all symbols and color codes. The map includes a north arrow and a scale bar (0 to 120 km).
All the records collected after the model was built (after 2021) fell almost entirely within suitable areas. Only one record was placed on an unsuitable area, but it was just 50 m away from a suitable pixel, likely due to a resolution issue (Figure 2).
We identified 15 areas larger than 208 km², totalling 53,435 km² (Figures 3 and 4), of which 17,838 km² (33.4%) are inside Strictly Protected Areas (I, II, and III IUCN categories). In 9 of these 15 areas, there are no recent records (Figure 3, areas C, D, F, G, and I; Figure 4, areas C, D, E, and F), although they are all close to occurrences of the species (<200 km). Noteworthy among the suitable areas without recent records are the mountainous region in Rio de Janeiro (Figure 3, area C), the southern portion of the province of Misiones (Figure 4, areas D and E), San Rafael National Park (Figure 4, area C), and the Serra da Mantiqueira (Figure 3, area I).
Discussion
The model showed excellent performance and was able to correctly predict the areas where the Black-fronted Piping-Guan still occurs, indicating a massive contraction in the species distribution range compared with its historical distribution. The suitable areas with recent records are mainly within Strictly Protected Areas, but there are also important areas for the species that are not protected. Our validation proves that the model presented is updated, being able to predict all new recent records. Additionally, there are suitable areas without recent records that could be targeted for reintroduction programmes.
The suitable areas for the Black-fronted Piping-Guan are mainly concentrated in two macro regions, which we will call Serra do Mar (areas in Figure 3) and Misiones (areas in Figure 4). The Serra do Mar region includes areas near the coast from São Paulo to Rio Grande do Sul, with a small suitable area in the mountainous region of Rio de Janeiro. The Serra do Mar and Paranapiacaba ranges, in the states of São Paulo and Paraná, possibly house the largest population of the species (Bernardo et al. Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011; Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997; Silveira et al. Reference Silveira, Soares and Bianchi2008; Torre et al. Reference Torre, Bastiani, Fratoni and Freitas2020), with several Strictly Protected Areas (Figure 3) with recent records, such as Serra do Mar State Park, Ilha do Cardoso State Park, Ilhabela State Park, Intervales State Park, Lagamar de Cananéia State Park, and Guaricana National Park.
The Misiones macro region includes the Argentine province of Misiones and nearby areas of Paraná, Santa Catarina, Rio Grande do Sul, and Paraguay, being the Atlantic Forest region farthest from the coast with the highest forest cover. The species occurs in several Strictly Protected Areas in this region, such as Iguaçu/Iguazu National Parks, Urugua-í Provincial Park, and Turvo State Park. However, despite having a high percentage of forest cover (61% in the province of Misiones; Projeto MapBiomas Mata Atlântica Trinacional 2021), this is the Atlantic Forest area that has suffered the greatest forest loss in recent years. From 2000 to 2019, about 11,865 km² of forest vegetation were replaced by other land uses in the Atlantic Forest of Paraguay and Argentina, which is equivalent to 10.5% of the biome’s extent in these countries (Projeto MapBiomas Mata Atlântica Trinacional 2021).
The most important variable for the model was the percentage of forest cover, which was expected, since the species is found only in large forest remnants (Bernardo et al. Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011; Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997). There are few records of the species in anthropised areas near large forest blocks, such as in altered forests and even Pinus sp. plantations (Bernardo et al. Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011; Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997), making it difficult to determine if the absence of the species in more fragmented landscapes with less forest cover is strictly due to the species’ environmental requirements or if hunting is responsible for its disappearance from fragmented areas to where people have access. Possibly, a combination of both factors restricts the species to large forest fragments, where hunting pressure, though present, has not yet been sufficient to extinguish the population. The Black-fronted Piping-Guan may be similar to two other related species considered restricted to primary forests, Great Curassow Crax rubra and Great Tinamou Tinamus major , which were recorded occurring without differences in occupancy probability in primary and secondary forests where hunting was eliminated (Whitworth et al. Reference Whitworth, Beirne, Flatt, Huarcaya, Cruz Diaz and Forsyth2018).
The original vegetation of the Atlantic Forest is currently restricted to a few large forest blocks and many small and disconnected remaining fragments (Projeto MapBiomas 2021; Ribeiro et al. Reference Ribeiro, Metzger, Martensen, Ponzoni and Hirota2009). In the Brazilian Atlantic Forest, the percentage of forest cover has remained stable in recent years (Projeto MapBiomas 2021). However, this stability in quantity hides the reduction of mature forests, replaced in numbers by passive and active restoration, as most land use and cover maps consider forests of different ages in the same category (Rosa et al. Reference Rosa, Brancalion, Crouzeilles, Tambosi, Piffer and Lenti2021). Native forests take many decades to mature and recover levels of biomass and species richness close to the original (Martin et al. Reference Martin, Newton and Bullock2013; Rozendaal et al. Reference Rozendaal, Bongers, Aide, Alvarez-Dávila, Ascarrunz and Balvanera2019). For large forest frugivores like the Black-fronted Piping-Guan, such restoration plantings will be usable in a few decades (Chazdon et al. Reference Chazdon, Peres, Dent, Sheil, Lugo and Lamb2009; Powell et al. Reference Powell, Stouffer and Johnson2013), while the reduction in the availability of mature forests indicates that habitat loss continues to advance and threaten the species.
Argentina and Paraguay also show a recent increase in forest restorations, but even so, the amount of forest, regardless of age, is decreasing. In Argentina, despite still having more than 60% of the native forest vegetation standing, about 10% of the forest cover that existed in 2000 has been lost (1,834 km²), and the species has already been extirpated in several locations, especially in the north of Corrientes and south of Misiones, and some areas in the centre and west of Misiones (Cockle and Bodrati Reference Cockle and Bodrati2011). In Paraguay, the situation is more critical. More than a quarter (27%, 9,247 km²) of the forest vegetation existing in 2000 has been converted to other land uses (Projeto MapBiomas Mata Atlântica Trinacional 2021), which helps to explain the currently confirmed presence of the species in only one location. In this interior portion of the Atlantic Forest, the Argentine forests are the main refuges for the species, as the large neighbouring forest areas in Paraguay, and especially in Brazil, are scarce and isolated (Cockle and Bodrati Reference Cockle and Bodrati2011).
Deforestation, besides directly causing habitat loss, also promotes the increase of human pressure in the remaining areas, such as poaching (Brodie et al. Reference Brodie, Giordano, Zipkin, Bernard, Mohd-Azlan and Ambu2015; Cockle and Bodrati Reference Cockle and Bodrati2011; Laurance and Balmford Reference Laurance and Balmford2013). Thus, even standing forests are more accessible to hunters and palm tree collectors, for example. Due to their size, members of the Cracidae family, especially guans and curassows, have always been considered an important food resource by indigenous peoples and local communities (Brooks Reference Brooks and Brooks1999; Johnson Reference Johnson1993). Also, there are many reports of hunts involving large numbers of individuals killed in a short time (Sick Reference Sick1997). Even in pristine areas like the Amazon, the constant removal of individuals by hunting impacts the population (Barrio Reference Barrio2011; Johnson Reference Johnson1993). In fragmented areas like the Atlantic Forest, the situation is even worse, due to human pressure throughout its territory, even in conserved and protected areas (Bodrati and Cockle Reference Bodrati and Cockle2006; Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997). In forest fragments, where movement is limited and there is no flow of individuals from a source, local extinction due to hunting is often the unwanted result for a species like the Black-fronted Piping-Guan (Robinson Reference Robinson, Schelhas and Greenberg1996). Additionally, the species is further disadvantaged by its ‘docile’ habit, staying perched on the same branch and feeding even in the presence of people (Carolina Minõ 2025, personal communication). Hunting-related factors may be more decisive for the presence or absence of a species than only habitat fragmentation, as found for the Red-billed Curassow in southern Bahia (Rios et al. Reference Rios, McGowan, Collar, Benchimol, Canale and Olmos2020).
Potential areas for translocations
All blocks identified as highly suitable for the species but without recent records are within its original distribution and are close to areas with recent records. Some have historical records, suggesting that the species once occurred in these areas but probably suffered local extinctions.
The mountainous region in Rio de Janeiro (Figure 3, area C), the largest of these areas, includes parts of the Serra dos Órgãos National Park and the Tinguá Biological Reserve, where the species was reported to be locally extinct a few decades ago (Mallet-Rodrigues et al. Reference Mallet-Rodrigues, Parrini and Pacheco2007) and has already been identified as a priority for reintroductions (Silveira et al. Reference Silveira, Soares and Bianchi2008). In the same region, two reintroductions of the species were carried out in the Guapiaçu Ecological Reserve. The first was conducted in 2007 and 2008, along with the Red-billed Curassow (Bernardo et al. Reference Bernardo, Rubim, Bueno, Begotti, Meirelles and Donatti2011, Reference Bernardo, Desbiez, Olmos and Collar2014), when at least 40 individuals of Black-fronted Piping-Guan were released, but individuals of both species were not recorded after 2014 (Bernardo Reference Bernardo2010; Bernardo and Locke Reference Bernardo and Locke2014). The second release, involving three individuals, was carried out in 2018 by the Jacutinga Project; however, two were predated and the third was recaptured to prevent further predation (A. Tassoni 2021, personal communication). Although it presents a large area suitable for the species, it is important to note that hunter activity still exists in the protected areas of this region (Constantino Reference Constantino2018), which could hinder new reintroduction attempts.
Four suitable areas near Misiones are isolated forest blocks, without connectivity to the forest block where the species still exists. One of them, San Rafael National Park in Paraguay (Figure 3, area C), had records until 1998 (Esquivel et al. Reference Esquivel, Peris, Fraga, Clay, Bodrati and del Castillo2007), but now is possibly locally extinct (Hugo Castillo 2021, personal communication). The other areas (Figure 3, areas D, E, and F) do not have Strictly Protected Areas. The smallest of the nine is located in Serra da Mantiqueira, on the border between São Paulo, Minas Gerais, and Rio de Janeiro, where the Jacutinga Project reintroduced 56 individuals from 2016 to 2024 (Souza L.D.C. et al. Reference Souza, Pereira, Azevedo, Bernardo, Develey and Ruiz-Miranda2020). The species was possibly extinct in the region, despite a record in 2001 (Portes and Monteiro Reference Portes and Monteiro2004). Other releases are planned and should be carried out in the coming years.
The remaining three areas are forest fragments in Paraná, Santa Catarina, and Rio Grande do Sul (Figure 3, areas G, F, and D, respectively). Although they are not Strictly Protected Areas, they are close to suitable areas with recent records and connected by forest vegetation. The absence of historical records in these areas may be due to low sampling effort in the surroundings, and due to the presence of the Black-fronted Piping-Guan in nearby areas, it may indicate a false absence of the species.
Most of the records used in this study are within Strictly Protected Areas, which represent the last refuges for the species. However, although considered Strictly Protected Areas, the presence of hunters and palm tree collectors within these places has been known for decades (Bodrati and Cockle Reference Bodrati and Cockle2006; Constantino Reference Constantino2018; Galetti and Chivers Reference Galetti and Chivers1995; Galetti et al. Reference Galetti, Martuscelli, Olmos and Aleixo1997). The lack of effective enforcement allows many of these protected areas not to fulfil their protection role, mainly due to the reduced number of enforcement agents (Galetti and Chivers Reference Galetti and Chivers1995; Munhoes Reference Munhoes2020). Park rangers themselves point out other difficulties, such as lack of training and equipment (Munhoes Reference Munhoes2020), which makes the effectiveness of Strictly Protected Areas for the protection of highly hunted species quite limited.
Limitations of our approach and important questions
We limited our approach to seeking environmentally suitable areas for the Black-fronted Piping-Guan without including spatial information on other factors that should be considered in conservation planning and reintroduction projects, such as hunting pressure, food availability, and micro-habitat quality. Although proxies for hunting pressure are available, including this layer in modelling could introduce spatial and temporal uncertainties (Barry and Elith Reference Barry and Elith2006). Spatial uncertainties are related to the resolution of available information, as these layers are generated from interpolations of data such as distance to urban centres and human density (Barry and Elith Reference Barry and Elith2006; Bogoni et al. Reference Bogoni, Peres and Ferraz2020), and even indicating areas with higher or lower hunting pressure on a broader scale, they may not be as effective for defining areas with lower hunting pressure for reintroducing a game species. Temporally, local actions can cause impacts that increase or decrease hunting pressure (Bragagnolo et al. Reference Bragagnolo, Gama, Vieira, Campos-Silva, Bernard and Malhado2019; Braga-Pereira et al. Reference Braga-Pereira, Bogoni and Alves2020). Concerning this issue, which is crucial in planning reintroductions, analyses focused on ‘candidate areas’ can be much more productive, even if they involve greater temporal and financial effort, as described by Gama et al. (Reference Gama, Malhado, Bragagnolo, Correia and Ladle2016) for the Alagoas Curassow Mitu mitu.
This more refined analysis, conducted locally, can also be useful for identifying the more specific conditions the species needs (Armstrong and Seddon Reference Armstrong and Seddon2008). The forests included in the forest cover percentage variable have different phytophysiognomies and levels of conservation (IBGE 2012; Rosa et al. Reference Rosa, Brancalion, Crouzeilles, Tambosi, Piffer and Lenti2021). Identifying structural and floristic traits favourable to the Black-fronted Piping-Guan is essential to increase the chances of success. Silveira et al. (Reference Silveira, Soares and Bianchi2008) mentioned the dependence on “conserved forests, with an abundance of Lauraceae and Myrtaceae”, traits that can be identified locally. To evaluate potential sites for reintroducing the White-winged Guan Penelope albipennis in Peru, researchers locally assessed plant diversity, water resources, and forest structure (Angulo and Barrio Reference Angulo and Barrio2004). As it is a species with a smaller distribution, it was possible to evaluate the areas in a few days. For the Black-fronted Piping-Guan, the SDM is useful for restricting available areas, complemented by local evaluation. Besides the mentioned environmental traits, it is important to consider factors such as the genetic and health status of the individuals to be reintroduced (Ferreira-Junior et al. Reference Ferreira-Junior, Dutra, Martins, Valkiūnas and Braga2018; Oliveira-Jr et al. Reference Oliveira-, Costa, Silveira and Francisco2016; World Pheasant Association and IUCN/SSC Re-Introduction Specialist Group 2009).
Another important tool is the Population Viability Analysis (PVA), which uses information about the species’ ecology and available resources to estimate survival probabilities. The PVA can be employed to determine the number of individuals to be released and for how long after the release (World Pheasant Association and IUCN/SSC Re-Introduction Specialist Group 2009). This approach has already been used for other Cracidae, such as the Red-billed Curassow (Bernardo and Locke Reference Bernardo and Locke2014; Bonfim et al. Reference Bonfim, Cordeiro, Peres, Canale and Bernardo2019), Salvin’s Curassow Mitu salvini (Johnson Reference Johnson1993), and the Black-fronted Piping-Guan itself, to aid in planning new releases (Alecsandra Tassoni 2021, personal communication). During an exercise considering only the reintroduced population in Serra da Mantiqueira, PVA results showed that more releases would be needed for the population to have a low probability of long-term extinction (Alecsandra Tassoni 2021, personal communication). Thus, the initial idea of releasing individuals in other areas was modified, increasing the population’s survival probability.
Conclusions
The potential distribution area of the Black-fronted Piping-Guan represents only 6.9% of the species’ original range. The most important variable for explaining the species’ presence probability was the percentage of forest cover. For reintroduction areas, we identified 15 areas larger than 208 km², nine of which do not have recent records and can be considered candidates for future reintroductions. We suggested a deeper analysis to include the hunting absence and the presence of necessary microclimatic conditions for the species to complement the initial area selection made by the SDM.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/S0959270926100501.
Acknowledgements
We are grateful to the Forest Science Department of the Luiz de Queiroz College of Agriculture, University of São Paulo (Brazil), the Wildlife Ecology, Management, and Conservation Lab (LEMaC), and the Graduate Program in Forest Resources (PPGRF). We thank Alecsandra Tassoni, SAVE Brasil, Hugo del Castillo, and Martjan Lammertink for shared information about the species, and all the birdwatchers and researchers who shared their data for this paper: Ademir Carosia, Ademir Costa, Ale Bianco, Alexandre Bianco, Alexandre Souza, André Guaraldo, André Menini, Andreas Oberhuber, Antonio Santos, Antonio Vilela, Arlei Antunes, Augusto M. Santos, Ben Phalan, Bianca Matinata and Legado das Águas, Bruno Lima, Camila Marques, Cárin Duarte, Carlos Brocardo, Carlos E. Nascimento, Carlos Gussoni, Carlos Moura, Carlos Neimar Kuhn, Carolina Minño, Cesar Medolago, Clécio Gomides, Dáfini Bruno, Daiane Carreira, Daniel Lamberg, Daniela Zaffignani, Danilo Junior, Diego Kondratzky, Diego Oscar, Douglas Bete, Eduardo Bergo, Fábio A.F. Jacomassa, Fábio Benedetti, Fabio Ming Hallais, Fabio Schunck, Fernando Jacobs, Fernando Straube, Francisco T. Tatit, Frederico Swarofsky, Gabriel Leite, Glauco Tonello, Guilherme Willrich, Guillermo Gil, Marcelo Cavicchia, Gustavo Muniz, Hector Bottai, Ian Thompson, Isede Calvani, Ivo Kindel, João J Soares, João Rosa, João Salvador, Jonas C. Kilpp, Jorge L. Wolff, José Dionísio Bertuzzo, Jose Mauro C. Monteiro, Josiane Bernardes, Kassius Santos, Katlin Fernandes, Leandro Corrêa, Lenice Amaral, Leonardo Casadei, Leticia Munhoes, Lindemberg Caranha, Luciano Breves, Lucilene Jacoboski, Luís Roberto Silva, Manuel Encabo, Marcelo Camacho, Marco Cruz, Marcos Faria, Marcos Henrique Achado, Mário Nunes, Michelli Gomes, Natalia Rongetta, Nicelio Silva, Norton Santos, Patricio Ramirez Llorens, Paulo Amstalden, Paulo Silveira, Pedro Develey, Pedro Sessegolo, Rafael Ritter, Rene Santos, Ricardo Barbosa, Roberta Montanheiro Paolino, Roberto Dall Agnol, Roberto Fusco Costa, Roberval Londrina, Rodrigo Conte, Rodrigo Y Castro, Romulo Silva, Ronald Santiago, Ronaldo Cardoso, Rubia W. Speck Loes, Sidnei Leal, Sidney J. Monteiro, Tania Gomes, Tiago Machado-de-Souza, Ulisses Nemetz, Victor Leandro-Silva, Vincent Kurt Lo and Vitor Mazzochi Aguiar. This work was partially financed by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001. We also thank the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the productivity fellowship (KMPMBF; #303940/2021-2) and long-term fellowship (AAAB; #350533/2023-7).