


Introduction
Competitive interactions between exotic and native parasitoids are seldom studied in the context of biological control programmes before the introduction of an exotic natural enemy (Harvey et al. Reference Harvey, Poelman and Tanaka2013; Abram et al. Reference Abram, Franklin, Brodeur, Cory, McConkey, Wyckhuys and Heimpel2024). Most studies on environmental assessment for classical or neoclassical biological control have focused on direct nontarget attacks on organisms related to the pest species. This is partly due to logistical challenges associated with rearing native organisms with poorly known biology. Interspecific competition between parasitoids can be intrinsic and extrinsic (Godfray Reference Godfray1994; Harvey et al. Reference Harvey, Poelman and Tanaka2013). Intrinsic competition occurs when larvae of the two species develop inside the same individual host and either are forced to share resources (Poelman et al. Reference Poelman, Gols, Gumovsky, Cortesero, Dicke and Harvey2014) or one of the parasitoids kills the other. Extrinsic competition for resources occurs between adult parasitoids as they search and attack a host to lay their eggs (Ode et al. Reference Ode, Vyas and Harvey2022). Both types of competition are considered important factors in shaping community assemblages and may affect the efficacy of biological control of a target pest.
Biological control of Lygus (Hahn) species (Hemiptera: Miridae) with European parasitoid Peristenus digoneutis Loan (Hymenoptera: Braconidae) has been of interest to Canadians (Carignan et al. Reference Carignan, Stewart, Godin and Boivin2007) since Day (Reference Day1996) demonstrated its success in the northeastern United States of America. Goulet and Mason (Reference Goulet and Mason2006) reported the adventive migration of this parasitoid into Ontario and Quebec in the early 2000s, but after more than two decades, Lygus continue to cause damage to horticultural crops in those regions (Dumont et al. Reference Dumont, Mason, Lachance, Lucas, Cárcamo, Vankosky and Martel2024), bringing into question the parasitoid’s efficacy. Lygus bugs (L. keltoni, L. borealis, L. elisus, and L. lineolaris) are native pests of multiple crops throughout the temperate world. The insects feed on, for example, Brassicaceae and Fabaceae seed crops (Salt Reference Salt1945; Smith et al. Reference Smith, Rivard, Feng and Cárcamo2019; Aguiar-Cordero et al. Reference Aguiar-Cordero, Cárcamo and Prager2024; Cárcamo et al. Reference Cárcamo, Herle, Schwinghamer, Robinson, Reid and Gabert2024), horticultural crops (Rougoor Reference Rougoor2008), and greenhouse crops (Gillespie et al. Reference Gillespie, Foottit, Shipp, Quiring and Wang2003). Srivastava et al. (Reference Srivastava, Wist and Cárcamo2025) estimated field crop losses due to Lygus damage and pesticide costs on the Canadian prairies at approximately $40 million annually. Pesticide spraying, which is currently the standard control of Lygus bugs on crops (Cárcamo et al. Reference Cárcamo, Herle, Schwinghamer, Robinson, Reid and Gabert2024), adds to the economic costs and also has deleterious effects on the ecosystem (Schmidt-Jeffris Reference Schmidt-Jeffris2023) and human health (Valcke et al. Reference Valcke, Bourgault, Rochette, Normandine, Samuel and Belleville2017). Therefore, the potential economic, environmental, and health benefits from reduced pesticide spraying to control the pest that may accrue, if Lygus bug biological control were enhanced, could be significant and would outweigh the biological control’s potential negative impacts on biodiversity.
In western Canada, P. digoneutis has the potential to establish in the southern regions of the prairies where Lygus bugs have two generations (Haye et al. Reference Haye, Olfert, Weiss, Gariepy, Broadbent and Kuhlmann2013). Currently, native natural enemies do not provide sufficient suppression, and the only management option is through pesticides. Fernández et al. (Reference Fernández, Laird, Herle, Goulet and Cárcamo2018) documented the phenology of the native parasitoids in southern Alberta: Peristenus dayi Goulet is a spring species found mainly in Adelphocoris lineolatus Goeze (Hemiptera: Miridae), of European origin, but its native host is unknown; P. mellipes Cresson has a broad range in seasonal activity from late spring to mid-summer; and P. howardi Shaw occurs mostly in mid- to late summer. The first two species are transcontinental in distribution, whereas P. howardi’s distribution is limited to western North America (Goulet and Mason Reference Goulet and Mason2006). Exotic Peristenus digoneutis has two generations per year, and its phenology is expected to overlap with P. mellipes and P. howardi, similarly to how Peristenus mellipes and Leiophron lygivorous Loan (Hymenoptera: Braconidae) in eastern Canada attack the two generations of Lygus (Lachance et al. Reference Lachance, Broadbent and Sears2001; Day Reference Day2005; Goulet and Mason Reference Goulet and Mason2006; Carignan et al. Reference Carignan, Stewart, Godin and Boivin2007). Past surveys of Lygus nymphal parasitism in western Canada have revealed low overall levels in alfalfa and close to no (0) parasitism in canola or yellow mustard, Sinapis alba Linnaeus (both Brassicaceae) (Braun et al. Reference Braun, Erlandson, Baldwin, Soroka, Mason, Foottit and Hegedus2001; Gillespie et al. Reference Gillespie, Foottit, Shipp, Quiring and Wang2003; Fernández et al. Reference Fernández, Laird, Herle, Goulet and Cárcamo2018). In contrast to past surveys, Floate et al.’s (Reference Floate, Wilches-Correal, Ibarra-Galvis, Pawluk, Roy and Cárcamo2024) 2022–2024 survey showed that P. howardi has recently started to attack Lygus nymphs (L. elisus, L. keltoni, and L. borealis) in canola and other crops, with peak parasitism of some nymphal cohorts surpassing 80%. Given the more arid and colder climate of the prairies compared to conditions in eastern Canada and Europe and given also the phenological differences (Day Reference Day2005; Dumont et al. Reference Dumont, Mason, Lachance, Lucas, Cárcamo, Vankosky and Martel2024), one would expect that the native species would have a competitive advantage over P. digoneutis due to P. howardi’s adaptation to local climatic conditions and seasonal host phenology (Day Reference Day2005). These findings on the seasonal activity and increasing levels of parasitism of Lygus bugs by P. howardi reflect emerging trends not observed in earlier surveys (e.g., Fernández et al. Reference Fernández, Laird, Herle, Goulet and Cárcamo2018) and may be influenced by shifting climate conditions or by changes in host plant use.
To our knowledge, only two studies have assessed competitive interactions among parasitoids of Lygus bugs. Lachance et al. (Reference Lachance, Broadbent and Sears2001) conducted laboratory assays to determine the extent of intrinsic competition between combinations of two exotic species (P. digoneutis and P. relictus Loan = stygicus) and native species. The native species included Peristenus pallipes Curtis and P. pseudopallipes Loan of the first and second generation of Lygus (Zhang et al. (Reference Zhang, Ridenbaugh and Sharanowski2017) synonymised both of these species to P. mellipes), as well as Leiophron lygivorous Loan (Hymenoptera: Braconidae), from Ontario and Quebec. Due to logistical constraints in rearing the native species, only a few replications were completed for the Leiophron and exotic Peristenus combinations. Lachance et al. (Reference Lachance, Broadbent and Sears2001) concluded that P. digoneutis was a superior in-host competitor than P. relictus or L. lygivorous was. Lachance et al. (Reference Lachance, Broadbent and Sears2001) did not expect intense competition and major suppression of the native species because the native Peristenus spp. have an earlier seasonal activity and wider host range than P. digoneutis does. Luo et al. (Reference Luo, Zhang and Wu2018) conducted a series of laboratory assays to assess interspecific intrinsic and extrinsic competition between Peristenus spretus Chen and van Achterberg (endemic to China) and P. relictus (Eurasian), which are important parasitoids of Apolygus lucorum Meyer-Dür (Hemiptera: Miridae), a mirid pest of cotton (Malvaceae) in China. They concluded that, regardless of the order of exposure to the parasitoid or of the densities of adult wasps and A. lucorum nymphs, P. spretus was the dominant competitor. They attributed the results to P. spretus’s faster development (2.2 days shorter) compared to that of P. relictus. Field studies in the northeastern United States of America (Day Reference Day2005) and California (Pickett et al. Reference Pickett, Nieto, Bryer, Swezey, Stadtherr and Wisheropp2013) have demonstrated that the exotic Peristenus species become the dominant parasitoids of Lygus bugs but that they do not extirpate the native parasitoids because the latter persist in other hosts, including Closterotomus norvegicus Gmelin, Leptopterna dolobrata Linnaeus, and Adelphocoris lineolatus (all Hemiptera: Miridae).
Our objective in the present study was to assess interspecific competition between Peristenus digoneutis and native Peristenus spp. from western Canada in laboratory experiments. We hypothesised that native species (P. mellipes or P. howardi) are not outcompeted, as measured by progeny count, by the exotic species, particularly in simultaneous interactions between females in a laboratory setting.
Materials and methods
Rearing both species of parasitoids each year was not possible due to the scarcity of P. howardi during the early years of the study and the consistently high overwintering mortality of both in most years. Therefore, for practical reasons, the focus native species were P. mellipes during the first two years and P. howardi in the last year of the study after COVID pandemic restrictions were lifted for government employees.
2018 and 2019
Native Peristenus mellipes were reared in the laboratory at the Lethbridge Research and Development Centre (Lethbridge, Alberta, Canada) from fourth and fifth instars of Lygus spp. (mainly Lygus keltoni Schwartz, but also L. elisus Van Duzee and L. borealis Kelton) that had been collected from ball mustard, Neslia paniculata Linnaeus Desvaux (Brassicaceae), in an urban park (Peenaquim Park, Lethbridge, Alberta, Canada, 49° 42′ 59.0″ N, 112° 50′ 53.3″ W) on 9 June 2017. Groups of about 200 nymphs were reared in 5-L buckets equipped with a 2-mm mesh screen at the bottom. Peristenus larvae could then fall onto a jar with moist vermiculite to form a cocoon and pupate. This mass-rearing method was adopted from the one described by Haye (Reference Haye2004). The Peristenus mellipes used in 2019 came from laboratory assays performed in 2018.
Exotic Peristenus digoneutis were purchased as cocoons shipped in July of each year from the Alampi Beneficial Insect Rearing Laboratory, State of New Jersey, Department of Agriculture (Ewing, New Jersey, United States of America; (https://www.nj.gov/agriculture/divisions/pi/prog/beneficialinsect.html). Upon arrival, the cocoons were moved into a quarantine facility, where they were allowed to emerge in a cage and mate. Next, individual mated wasps were isolated in glass micropipettes and presented with a second- or third-instar Lygus nymph to confirm attacking females, which were then transferred to a separate cage before they were used in the competition assays.
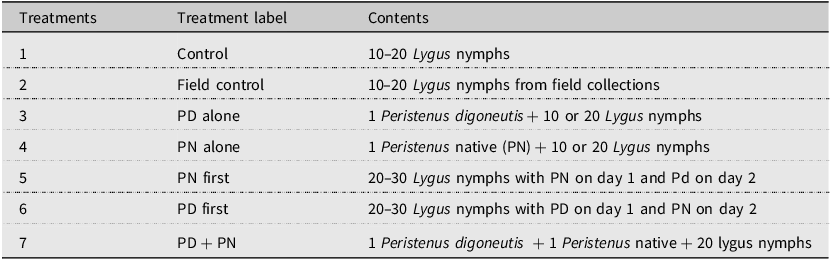
Assays were conducted in 0.75-L cylindrical containers with meshed screens on the lids for airflow (Luo et al. Reference Luo, Zhang and Wu2018). Generally, the Lygus nymphs used at the second- or third-instar stage were obtained from lab-reared colonies of Lygus keltoni or Lygus elisus, but occasionally field-collected nymphs (dominated by L. keltoni), potentially already parasitised, had to be used. From 20 to 30 nymphs were added to each container, depending on availability. During this period, two controls – one with nymphs from the laboratory and one with nymphs from field collections – without parasitoid wasps were included, and the control nymphs were allowed to reach the adult stage to confirm they were one of the two dominant pest species (L. keltoni or L. elisus). Competition assays included sequential tests, with nymphs exposed to the exotic or the native species first on day 1 and to the other species on day 2 (Table 1). The order of the parasitoids that the nymphs were exposed to was selected at random or based on availability. A third competition treatment included exposing Lygus nymphs to the exotic and native parasitoid simultaneously over 24 hours. Due to high mortality or low emergence of female parasitoids in the lab (similar to Lachance et al.’s Reference Lachance, Broadbent and Sears2001 study), a few individuals were used 2–3 times. Individual effects were included as a random factor in the GLIMMIX models, as discussed in the Data analysis section, below.
Treatments for laboratory competition assays between the exotic Peristenus digoneutis (PD) and native (PN) Peristenus mellipes or P. howardi from 2018, 2019, and 2023

Table 1. Long description
The table presents treatments for laboratory competition assays between the exotic Peristenus digoneutis (PD) and native (PN) Peristenus mellipes or P. howardi from 2018, 2019, and 2023. It consists of seven rows and three columns. The columns are labelled Treatments, Treatment label, and Contents. The rows detail various treatments involving Lygus nymphs and parasitoid wasps. Treatment 1 is a control with 10-20 Lygus nymphs. Treatment 2 is a field control with 10–20 Lygus nymphs from field collections. Treatment 3 involves 1 Peristenus digoneutis plus 10 or 20 Lygus nymphs. Treatment 4 involves 1 Peristenus native (PN) plus 10 or 20 Lygus nymphs. Treatment 5 involves 20–30 Lygus nymphs with PN on day 1 and PD on day 2. Treatment 6 involves 20–30 Lygus nymphs with PD on day 1 and PN on day 2. Treatment 7 involves 1 Peristenus digoneutis plus 1 Peristenus native plus 20 Lygus nymphs.
The rearing of the parasitoids from the assays followed the methods outlined by Whistlecraft et al. (Reference Whistlecraft, Haye, Kuhlmann, Muth, Murillo and Mason2010). In a 0.75-L plastic tub, Lygus nymphs were fed romaine lettuce twice a week until they died or reached the adult stage. Rearing took place at room temperature (22 °C), under a 16:8–hour light:darkness photoperiod and approximately 40% relative humidity. Parasitoid larvae were allowed to mature in the rearing containers for the remainder of the summer under the same conditions. In September and October, the vermiculite in each Petri dish was searched, and the number of cocoons was counted using a stereoscope at 10× magnification. The cocoons were then cold-acclimated at 12 °C for 2–4 weeks before overwintering at 3 °C for 6–7 months. In June or July of the following year, the cocoons were transferred to room temperature to allow adult wasps to emerge for identification. Due to COVID-19 laboratory access restrictions in 2020, material from the 2019 assays could not be reared to adults after overwintering.
Peristenus digoneutis came from Lygus lineolaris Palisot de Beauvois nymphs collected in southern Quebec in forage fields during the summer of 2022 and shipped to Lethbridge, where they were reared and overwintered, as described in the 2018 and 2019 section, above. The native Peristenus species used in the 2023 competition assays was P. howardi. The majority of these parasitoids were collected as adults (aspirated from sweep nets) from various forage and field crops in southern Alberta in July 2023, and a few were reared from Lygus nymphs collected during the previous year from similar fields.
The assays for 2023 were completed in similar containers and using the same methods as those described for the years 2018 and 2019, above but did not include the two sequential competition assays (Table 1). Instead of rearing the Lygus nymphs until Peristenus larvae emerged, the nymphs were dissected during the fifth-instar stage, about 7–10 days after the adult wasp was removed, as Floate et al. (Reference Floate, Wilches-Correal, Ibarra-Galvis, Pawluk, Roy and Cárcamo2024) validated in a survey of Peristenus parasitism. Any Peristenus larvae from dissections were stored in 95% ethanol and processed using polymerase chain reaction screening to determine species.
Molecular polymerase chain reaction determination
In 2023, molecular Peristenus determination (P. digoneutis or native) was performed, following the methods that Tilmon et al. (Reference Tilmon, Danforth, Day and Hoffmann2000) developed and that Floate et al. (Reference Floate, Wilches-Correal, Ibarra-Galvis, Pawluk, Roy and Cárcamo2024) validated to document parasitism by western Peristenus species. This method uses a two-step molecular approach: (1) cytochrome c oxidase 1 (CO1) gene amplification and (2) restriction digest of the polymerase chain reaction product. Extraction of DNA and amplification followed Tilmon et al. (Reference Tilmon, Danforth, Day and Hoffmann2000) and Simon et al.’s (Reference Simon, Frati, Beckenbach, Crespi, Liu and Flook1994) methods. After the polymerase chain reaction cycle was completed, Alu 1 enzyme and fluorescent dye were added, according to the recommended protocol for polymerase chain reaction products. The samples were then sorted as either P. digoneutis or native Peristenus, based on the banding pattern shown under ultraviolet illumination. We sequenced the samples to evaluate whether the method successfully differentiates the Peristenus species under evaluation in this study. For sequencing, the extracted DNA was amplified with the universal primers, HCO2198 (5′-TAA ACT TCA GGG TGA CCA AAA AAT CA-3′) and LCO1490 (5′-GGT CAA CAA ATC ATA AAG ATA TTG G-3′; Folmer et al. Reference Folmer, Black, Hoeh, Lutz and Vrijenhoek1994), and using general methods described in Hebert et al. (Reference Hebert, Cywinska, Ball and deWaard2003). The amplicons were visualised by electrophoresis in 1% agarose gels and purified using the QIAGEN QIAquick Gel Extraction Kit (Qiagen Inc., Valencia, California, United States of America) following the manufacturer’s protocols. The purified product was sent for sequencing to the University of Calgary (University Core DNA Services, Calgary, Alberta, Canada). We compared the sequences with those available in GenBank (https://www.ncbi.nlm.nih.gov/genbank/).
Size comparisons
Ten females of P. howardi and P. digoneutis were selected from laboratory-emerged specimens to compare the length of a number of morphological traits (head breadth, front and hind tibia lengths, mesoscutum breadth, and metasomal tergite 1 length) that may indicate overall adult size. The traits were selected from a list used in Zhang et al.’s (Reference Zhang, Ridenbaugh and Sharanowski2017) taxonomic revision of the genus. Measurements were taken with a Leica microscope (Ivesta 3 integrated camera; Leica, Wetzlar, Germany) at various resolutions, depending on the trait.
Data analysis
The main response variable of interest was the count of the progeny of native and exotic Peristenus that resulted from the various competition assays. Assessing the fitness effects of competition was not practical due to rearing challenges and was beyond the scope of the study. The level of field parasitism of Lygus nymphs by native Peristenus wasps was used to adjust the number of native parasitoids that emerged from competition assays that required field-collected nymphs. This was done to ensure that the results would not be biased in favour of the native species for those assays. Counts of native or exotic Peristenus progeny from the various competition treatments were compared using the GLIMMIX procedure in SAS, version 9.4 (SAS Institute, Cary, North Carolina, United States of America), with a Poisson response distribution and a log-link function. Individual wasps were given a unique label, and the potential effect of using them more than once on the outcome of competition was tested by adding this variable as a random factor in the GLIMMIX model. We used hierarchical (i.e., mixed) models that included random effects that were associated with the structures of grouping in the experimental design and sampling: block was the spatial or temporal experimental replication; exotic_individual_id (to assess repeat use of exotic individuals) and native_individual_id (for repeated use of native individuals) were subjects of repeated measures; and number of_nymphs or source (lab or field) was included to model stratification and repeated measures, where appropriate. By including random effects, we modelled the nonindependence of observations in groups. Each model was designed to analyse the structure and scale of its data. A simpler GLIMMIX model with a gamma distribution for two-group comparison using an F-test was used to analyse the body-size differences between the two wasp species. A chi-squared test was used to analyse the proportion of native P. howardi progeny that emerged in the simultaneous (extrinsic) competition test with females of P. digoneutis wasps. This analysis was done with the prop.test function in base R, version 4.4.1 (R Project; available at https://cran.r-project.org/bin/windows/base/old/4.4.1/).
Results
The analysis for each year revealed no significant effects of any of the treatments on the counts of native Peristenus progeny in the various assays. In 2018 (GLIMMIX procedure with Poisson distribution, F 3,122 = 2.12, P = 0.10) and in 2019 (F 3,75 = 1.34, P = 0.27), numerically more progeny were found in the assays that included only the native species than in those that included sequential or simultaneous exposure to the exotic female wasp (Fig. 1). The number of progeny was low and the variability high in all the assays, and no significant statistical differences were observed. More replicates might determine whether the differences are significant. In 2023, the counts of native P. howardi (F 1,14 = 0.34, P = 0.57) larvae in assays with or without competition did not differ significantly (Fig. 1). In the “simultaneous” treatment, in which the two species were allowed to compete for Lygus nymphal hosts, the number of native wasps was higher than those of the exotic species (99 versus 70). The proportion of native wasps from this treatment was significantly higher than 0.5 (χ2 = 42.5, df = 1, P = 1.327e–5).
Mean counts and data range for native Peristenus mellipes (2018, 2019; n = 9–76) or P. howardi (2023; n = 15–17) progeny from the laboratory competition assays. The SAS PROC GLIMMIX analysis with DIST = POISSON revealed no statistical differences, P > 0.05. Abbreviations are defined as follows: pmonly, P. mellipes only; seqpd, sequential assay with P. digoneutis first; seqpm, sequential assay with P. mellipes first; sim, simultaneous test; phonly, P. howardi only.

Figure 1. Long description
The dot plot presents the mean counts and data range for native Peristenus mellipes and P. howardi progeny from laboratory competition assays conducted in 2018, 2019, and 2023. The x-axis represents different treatments: pmonly, seqpd, seqpm, sim, and phonly. The y-axis shows the least-squares means of native Peristenus ranging from 0 to 15. Each dot represents individual data points, with error bars indicating the range of data. The treatments are grouped by year: 2018, 2019, and 2023. The plot shows variations in progeny counts across different treatments and years, with some treatments exhibiting higher variability and outliers. All values are approximated.
In 2018, competition treatment effects were statistically significant for P. digoneutis (F 3,83 = 6.57, P = 0.0005). The exotic P. digoneutis produced fewer progeny under treatment conditions where it competed simultaneously with adult native wasps than in the assays without competition (P = 0.0042) and under treatment conditions where a native wasp had attacked a nymph the day before the exotic wasp attacked the same nymph, thereby resulting in within-host larval competition (P = 0.0406; Fig. 2). In 2019, the counts were similar across all treatments (F 3,72 = 0.17, P = 0.9178; Fig. 2). In 2023, when P. howardi was the native adult competitor in the simultaneous competition treatments, no statistical difference in the counts of P. digoneutis progeny occurred compared to the P. digoneutis progeny counts in the treatment without competition (F 3,14 = 1.56, P = 0.23; Fig. 2).
Mean counts and data range for the exotic Peristenus digoneutis progeny from the laboratory competition treatments from 2018 (n = 24–76), 2019 (n = 9–36), and 2023 (n = 15–17). SAS PROC GLIMMIX with DIST = POISSON revealed statistical differences only in 2018 (F 3,83 = 6.57, P = 0.0005). Abbreviations are defined as follows: pdonly, P. digoneutis only; seqpd, sequential assay with P. digoneutis first; seqpm, sequential assay with P. mellipes first; sim, simultaneous test.

Figure 2. Long description
The box-and-whisker plot displays the least-square means of exotic Peristenus progeny across various treatments for the years 2018, 2019, and 2023. The x-axis represents different treatments: pdonly, seqpd, seqpm, and sim. The y-axis indicates the least-square means of exotic Peristenus, ranging from 0 to 15. Each treatment category contains three vertical box plots corresponding to the years 2018, 2019, and 2023. The boxes show the interquartile range (Q1 to Q3), with the median (Q2) marked by a line inside each box. Whiskers extend to the minimum and maximum values, and outliers are represented by individual points. In 2018, statistical differences are noted, with some treatments showing higher variability and outliers. The treatments include pmonly, seqpd, seqpm, and sim, each with distinct data ranges and median values. The plot highlights the variability and trends in the exotic Peristenus progeny counts across different treatments and years.
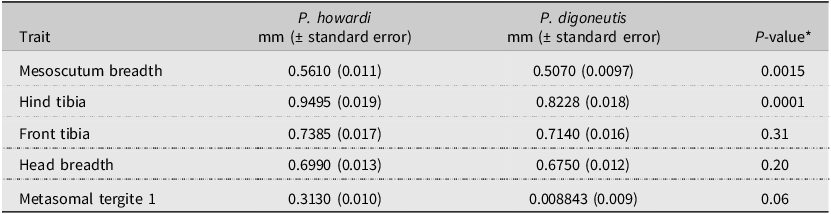
All five body measurements (Table 2) suggested that P. howardi females are slightly larger than P. digoneutis females, although not all measurements had statistically significant differences. The breadth of the head and the length of the front tibia were slightly shorter for P. digoneutis than for P. howardi but not statistically so (P > 0.20 for both traits). On the other hand, the breadth of the mesoscutum and the length of hind tibia were significantly greater in P. howardi than in P. digoneutis (P < 0.0015; Table 2). The metasomal tergite 1 was also smaller for P. digoneutis than for P. howardi, but the statistical differences were marginally significant (P = 0.06).
Means and standard errors for five body measurements comparing Peristenus digoneutis and P. howardi from laboratory emerged females

Table 2. Long description
The table presents a comparison of five body measurements between Peristenus digoneutis and P. howardi females, including the mesoscutum breadth, hind tibia length, front tibia length, head breadth, and metasomal tergite 1 length. The table has five rows and four columns. The columns are labelled Trait, P. howardi in millimetres with standard error, P. digoneutis in millimetres with standard error, and P-value. P. howardi females are generally larger than P. digoneutis females. The mesoscutum breadth and hind tibia length show significant differences, with P. howardi being larger. The head breadth and front tibia length are slightly shorter for P. digoneutis but not significantly so. The metasomal tergite 1 is smaller for P. digoneutis, with marginal statistical significance.
* SAS PROC GLIMMIX with DIST = GAMMA; replication = 10 females for each species.
Discussion
Currently, biological control science operates in an environment of risk aversion. This environment motivates scientists to assess the risk of direct impacts of parasitoids on nontarget native organisms and the potential displacement of native fauna through indirect interactions such as competition (Abram et al. Reference Abram, Franklin, Brodeur, Cory, McConkey, Wyckhuys and Heimpel2024). Performing these studies and, especially, assessing competition outcomes can be challenging due to constraints on raising native species with poorly known life histories and often obscure taxonomies. For example, Lachance et al. (Reference Lachance, Broadbent and Sears2001) attempted to assess within-host larval competition for the two native species, Peristenus pallipes and P. pseudopallipes (both synonymised as P. mellipes by Zhang et al. Reference Zhang, Ridenbaugh and Sharanowski2017), and for Leiophron lygivorus, from Ontario with the exotic P. digoneutis and P. relictus from Europe. However, those authors were unable to obtain sufficient Peristenus individuals that successfully survived the long overwintering period to complete the assays and score the outcome of competition. Based on the assays with Leiophron lygivorus, Lachance et al. (Reference Lachance, Broadbent and Sears2001) concluded that the exotic species, particularly P. digoneutis, were superior in-host competitors. In the present study, nearly a decade of effort was required to successfully overwinter the native Peristenus, to source and synchronise emergence of the exotic species, and to generate sufficient replicates for assessing competitive interactions under simplified laboratory conditions. This effort is consistent with past rearing challenges with Peristenus species, as reported by Lachance et al. (Reference Lachance, Broadbent and Sears2001) and Haye (Reference Haye2004). Based on these experiences, the expectation of assessing indirect impacts on potential competitors as part of environmental assessments of insect biological control – classical or neoclassical – is often too costly and impractical.
Our results suggest that the native Peristenus mellipes and P. howardi are not suppressed by competition with P. digoneutis in laboratory assays. In 2018, slightly lower numbers of P. mellipes progeny were produced in the presence of competitor larvae or of an adult female than without either, but the results were not statistically significant. Higher replication would be necessary to rule out potential competition effects. Lachance et al. (Reference Lachance, Broadbent and Sears2001) tested competition between P. mellipes (= P. pseudopallipes or P. pallipes) and only the more polyphagous P. relictus, not against P. digoneutis: in all of their tests, albeit with low sample sizes, the exotic species was found to be a superior competitor. The idea that P. mellipes in eastern Canada might be vulnerable to some displacement within Lygus hosts is supported by Goulet and Mason (Reference Goulet and Mason2006), who observed significant dominance by P. digoneutis in the community of parasitoids in some sites in Quebec and Ontario. Interestingly, contrary to Goulet and Mason’s (Reference Goulet and Mason2006) field observations, in our lab assays in 2018, we observed a significant reduction in the number of P. digoneutis progeny in treatments where the competing native species P. mellipes was present: the P. mellipes population on the prairies may be a superior competitor than the species’ population in eastern Canada. In fact, before Zhang et al.’s (Reference Zhang, Ridenbaugh and Sharanowski2017) revision, Goulet and Mason (Reference Goulet and Mason2006) considered the western populations of P. mellipes a distinct species complex: P. carcamoi and P. otaniae Goulet.
Surveys of Lygus lineolaris nymphal parasitism in Ontario (Dumont et al. Reference Dumont, Mason, Lachance, Lucas, Cárcamo, Vankosky and Martel2024) and in Quebec (Labrie and Cárcamo, unpublished data 2023, 2024) also suggest that P. digoneutis has become the dominant species developing in Lygus nymphs, potentially at the expense of native Peristenus but not of Leiophron. Direct evidence for displacement remains lacking because recent partial survey data do not provide evidence that the native Peristenus spp. have been suppressed in their native habitats. The species are expected to survive in their native plant hosts because their host ranges are wider than those of P. digoneutis (Day Reference Day2005). This expectation is supported by data from areas where exotic Peristenus have been established for several years: in the northeastern United States of America, Day (Reference Day2005) showed that an increased dominance of P. digoneutis suppressed populations of native Peristenus in Lygus, but native Peristenus remained common parasitoids in other mirids, including Leptopterna dolobrata and Adelphocoris lineoleatus. In central California, United States of America, P. relictus has become established and seems to suppress Lygus, along with the native parasitoid (Peristenus nr. pallipes), but again the native species occurs at high frequency in another mirid, Closterotomus norvegicus Gmelin (Pickett et al. Reference Pickett, Nieto, Bryer, Swezey, Stadtherr and Wisheropp2013). This is of interest because P. relictus is considered a more generalist parasitoid than P. digoneutis is because it attacks several genera within the Miridae (Haye Reference Haye2004; Haye et al. Reference Haye, Kuhlmann, Goulet and Mason2006). Yet, in California, the native Peristenus nr. pallipes is as prevalent as P. relictus in C. norvegicus nymphs (Pickett et al. Reference Pickett, Nieto, Bryer, Swezey, Stadtherr and Wisheropp2013). An extensive survey of Lygus bugs and other mirid nymphs in multiple crops and in noncultivated plants throughout the season in eastern Canada would likely show similar patterns of coexistence of native and exotic Peristenus in plant bugs other than Lygus.
Laboratory assays are highly simplified artificial constructs that likely exaggerate real-life ecological tri-trophic interactions among plant hosts, herbivores, and parasitoids and may not predict field outcomes accurately. That Peristenus species such as P. pseudopallipes (= mellipes) from the northeast United States of America is highly attracted to volatiles of Erigeron flowers is well established (Shahjahan Reference Shahjahan1974; Halloran et al. Reference Halloran, Mauck, Fleisher and Tumlinson2013). Likewise, working in cotton fields in China, Xia et al. (Reference Xia, Zhang, Williams III and Lu2023) demonstrated that Peristenus spretus Chen and van Achterberg (Hymenoptera: Braconidae) preferred to attack Apolygus lecorum (Hemiptera: Miridae) in trap strips of buckwheat, Fagopyrum esculentum Moench (Polygonaceae), than in cotton, Gossypium hirsutum (Malvaceae). That native Peristenus species in western Canada and the exotic P. digoneutis also exhibit differential preferences to plant hosts of Lygus bugs is therefore also a reasonable expectation. Such partitioning of the Lygus bug resources by plant hosts is expected to reduce competitive interactions among exotic biocontrol agents and native parasitoids and to have additive mortality effects to reduce pest populations.
Based on the laboratory assays in the present study, we conclude that the exotic P. digoneutis would not outcompete the two native Peristenus species that are expected to have some overlap in seasonal activity if the exotic was relocated and became established in western Canada. In fact, P. howardi produced more progeny than P. digoneutis did when females competed for Lygus nymphs. This result is consistent with some of our measurements that showed that P. howardi is larger than P. digoneutis. Our findings contrast with statements by Zhang et al. (Reference Zhang, Ridenbaugh and Sharanowski2017), who highlighted P. howardi as the most vulnerable to become “extinct” if P. digoneutis were to be relocated to the Canadian prairies. Furthermore, P. howardi occurs outside the Prairies Ecozone in the interior of British Columbia, Canada (Acheampong and Cárcamo, unpublished data), and in the Pacific Northwest of the United States of America, where it has a flexible life history: some populations are univoltine, and others are bivoltine (Day et al. Reference Day, Baird and Shaw1999). At some sites, P. howardi adults result from a combination of nondiapausing larvae and from larvae produced the previous year (Day Reference Day2005), demonstrating a life-history plasticity that should make the species less vulnerable to competition. Differences in synchronisation with their nymphal host instars have been documented to vary for Peristenus species in the eastern United States of America and likely also occur in western Canada. Finally, even if native Peristenus species were supressed by the exotic Peristenus digoneutis on the prairies, the potential direct benefits that may accrue from biological control and the economic and environmental benefits that would result from reduced pesticide spraying are expected to outweigh this risk (Srivastava et al. Reference Srivastava, Wist and Cárcamo2025).
Acknowledgements
This work was supported by grants awarded to H.C. by the Western Grains Research Foundation and the Alberta Canola Producers Commission (ACPC-133-AR20; AFC2020F093R). The authors gratefully acknowledge field and laboratory support from S. Daniels, R. Brandt, P. Eudes, A. Efetha, J. Lawal, R. Oliver, S. Berri, B. Taylor, M. Kanashiro, A. Pawluk, K. Beres, A. Strain, and B. Bennett. The foundational scientific research on Peristenus biology over two decades that allowed completion of this project would not be possible without the internal human and physical resources provided by Agriculture and Agri-Food Canada.
Competing interests
The authors declare that they have no competing interests in any form in relation to the above submission to The Canadian Entomologist.



 Open access
Open access