


1. Introduction
The continental microvertebrate fauna from the latest Cretaceous of Europe remains poorly known. This is especially true for the mammals which are among the least known world-wide. The Late Cretaceous fossil record of European mammals is characterized by highly contrasting geographical distribution and faunal content. It shows a concentration of the sites in two well-separated areas with distinct mammalian composition: (1) Western Europe, corresponding to the palaeogeographic area of the Ibero-Armorican Island, where about ten Campano-Maastrichtian sites have yielded only therians (eight named species); (2) Eastern Europe, corresponding to the palaeogeographic area of the Hațeg Island, where some thirty sites from the latest Campanian and the Maastrichtian of Romania have yielded only multituberculates (four or five species in Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Vasile, Brusatte and Norell2022 and Solomon et al. Reference Solomon, Codrea, Venczel and Smith2022).
In Western Europe, three late Cretaceous mammal localities are known in Spain and Portugal (Antunes et al. Reference Antunes, Sigogneau-Russell and Russell1986; Pol et al. Reference Pol, Buscalioni, Carballeira, Francés, López Martinez and Marandat1992; Gheerbrant and Astibia, Reference Gheerbrant and Astibia1999, Reference Gheerbrant and Astibia2012). All the others are in France, and mostly in southern France (Provence, Hérault) and in the northern Pyrenees (Ledoux et al. Reference Ledoux, Hartenberger, Michaux, Sudre and Thaler1966; Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997; Tabuce et al. Reference Tabuce, Vianey-Liaud and Garcia2004; Martin et al. Reference Martin, Case, Jagt, Schulp and Mulder2005; Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). In the Pyrenean area, microvertebrate faunas with mammals have been reported by the first author of this paper from the Maastrichtian Auzas Marls Formation in the Petites Pyrénées (departments of Haute-Garonne and Ariège), with first discoveries in 1992 and 1994 (Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997), and with a recent find published in 2021 (Gheerbrant and Teodori Reference Gheerbrant and Teodori2021). Preliminary field work by Gheerbrant et al. (Reference Gheerbrant, Abrial and Cappetta1997) on the Cassagnau Hill, in Haute-Garonne, reported in particular on two local promising vertebrate localities named Tricouté. One site, Tricouté 1, yielded microvertebrates, and the other, Tricouté 2, yielded only macrofauna (dinosaurs). Later on, Laurent (Reference Laurent2003) reported a third Tricouté locality (Tricouté 3), with macroremains of crocodilians and dinosaurs. Further prospecting and excavation on the Cassagnau Hill of the Marnes d’Auzas fossiliferous levels was conducted in the early 2000s. Two microvertebrate sites of Tricouté, one new (Tricouté 4, see below), have been intensively sampled for this study. After a long and tedious period of processing and sorting of the sampled sediments, we herein report from these sites of a new microvertebrate assemblage including mammals. The mammals document an unexpected taxonomic assemblage of multituberculates and eutherians which was previously unknown in Europe prior to this report and the preliminary one by Marković et al. (Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025). They belong to new taxa which are here described and studied in detail.
The microvertebrate sites from the Auzas Marls Formation are of great interest because they are well-dated by a marine microfauna and a characean flora, and because they are among the latest known from the Mesozoic in Europe. They provide key fossil evidence for knowledge of the evolution at the K/Pg transition in Europe which remains the most poorly documented in Laurasia. They are close in age, i.e. late Maastrichtian, with other Pyrenean sites from the Tremp Formation (e.g. López-Martίnez et al. Reference López-Martίnez, Canudo, Ardèvol, Suberbiola, Orue-Etxebarria, Cuenca-Bescós, Ruiz-Omeñaca, Murelaga and Feist2001). Notably, the late Maastrichtian Tricouté sites from the Auzas Marls Formation and the early Danian site of Fontllonga-3 from the Tremp Formation (Lopez-Martinez and Pelaez-Campomanes, Reference López Martínez and Pelaez-Campomanes1999) have yielded the continental microvertebrate assemblages that most closely bracket the K/Pg boundary in Europe. In addition, these sites are close to each other on a regional scale (Pyrenees), providing unparalleled palaeontological data on the evolution during the K/Pg transition at a local scale in Europe.
2. Material and methods
2.a. Material
The new microvertebrate material studied here comes from the Tricouté 1 site reported by Gheerbrant et al. (Reference Gheerbrant, Abrial and Cappetta1997) and from the new site of Tricouté 4 here reported for the first time. The fossil material from Tricouté 4 was collected in a two-ton sample of sediment. For extraction of the Tricouté 4 sediment, the fossiliferous level was cleared and uncovered using a mechanical shovel over an area of approximately 12 m2 (Figure 2a). The sampled fossiliferous sediments, which consist of calcified fluvial bioclastic sandstone, were processed by acid attacks at the CR2P-MNHN laboratory for microvertebrate separation. This process yielded approximately 40 kg of microvertebrate concentrate, which was sorted under a binocular microscope with the help of several collaborators (see Acknowledgements). The sorting of the microvertebrate remains was completed in 2011.
2.b. Dental anatomy and measurements
For multituberculate mammals, the identification of the tooth position follows Kielan-Jaworowska et al. (Reference Kielan-Jaworowska, Cifelli and Luo2004) and the cusp formula is counted from the labial to the lingual side, with each number of cusps per row being separated by a colon. For the eutherian mammals, we use the classic tribosphenic molar terminology of Van Valen (Reference Van Valen1966).
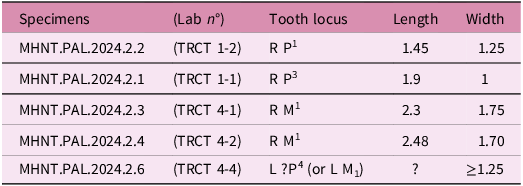
Linear measurements were taken on tooth specimens with a binocular microscope, to the accuracy of two decimal places, and on STL digital models using the software Meshlab (2021 version). An exception is the specimen MHNT.PAL.2024.2.5, for which the lengths were measured with Meshlab on the 3D digital model of the reconstructed tooth to obtain the best estimate of the size of the unfractured original specimen. Volumes of teeth (enamel, dentine and pulp cavity) were measured on 3D digital models using the software 3D Slicer 5.6.2 and Mimics Innovation Suite. Abbreviation of measurements: L, H, W: length, height and width.
2.c. CT scans
High-resolution computed microtomographies (μCT) of the material described here were performed at the AST-RX platform of the MNHN, Paris, using a GE Sensing and Inspection 3 Technologies phoenix|x-ray v|tome|x L240-180 CT scanner. Scan parameters for the different specimens are provided in Table 1. Data were reconstructed using datos|x reconstruction software Phoenix|x-ray. The segmentation, analysis and visualization of the image stacks were made by F. Goussard (CR2P-MNHN) using the software Mimics Innovation Suite (Materialise, Research Edition Release 21, https://www.materialise.com/en/medical/software/mimics).
CT scan parameters for the mammal material from the Tricouté microsites TRCT 1 and 4 (Maastrichtian of the Petites Pyrénées, France)

2.d. Cladistic analyses
We performed two cladistic analyses of the mammals studied here, one for the multituberculates and another one for the eutherians. For our parsimony analysis, we used the software TNT 1.6 (Goloboff et al. Reference Goloboff, Farris and Nixon2008). Traditional search (standard analysis) was made with 100 replications and swapping algorithm TBR. For the multituberculates, 15 000 trees were retained in memory for the analyses, whereas for the eutherians 10 000 trees were retained in memory for the analyses. The coding of the characters starts from 0.
Multituberculates. Our phylogenetic analysis of the multituberculates is based on the character matrix of Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022) which was used in their study of the kogaionids (Supplementary Text S1). This matrix is in turn derived from the earlier works of Csiki-Sava et al. (Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018), Wang et al. (Reference Wang, Meng and Wang2019) and Weaver et al. (Reference Weaver, Varricchio, Sargis, Chen, Freimuth and Wilson Mantilla2020). The matrix includes 130 characters (10 ordered) and 56 taxa. The coding of one character was changed with respect to Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022) (see text).
Eutherians. Our phylogenetic analysis of the eutherians is based on the character matrix of Gheerbrant and Teodori (Reference Gheerbrant and Teodori2021), previously used for the study of Azilestes ragei and modified from Archibald and Averianov (Reference Archibald and Averianov2012) and Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013) (Supplementary Text S2). This matrix is primarily devoted to the study of the relationships of the eutherian and placental mammals. It includes 408 characters (six ordered) and 77 taxa. In this matrix, 33 characters have been coded for cf. Azilestes yvettae n. sp., all concerning the upper molar morphology. The coding of a few characters was changed with respect to Gheerbrant and Teodori (Reference Gheerbrant and Teodori2021) and Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013). This concerns especially character 62 (state 62-1, instead 62-0, molar cups robust for Valentinella).
Abbreviations: RI= retention index; CI= consistency index.
2.e. Institutional abbreviations
The material sample (mostly mammal teeth) studied here is recorded and catalogued in the collection of the Musée d’Histoire Naturelle de Toulouse (MHNT, France), under the numbers with prefix MHNT.PAL. (see Supplementary Table S1). Numbers with the acronym TRCT refer to an informal laboratory numbering of the specimens that was made during the sorting of sediments after their concentration by acid attack processing.
CR2P: Centre de recherche en Paléontologie, Paris; MNHN: Museum National d’Histoire Naturelle (France).
3. Geographical, geological and stratigraphic contexts of the Tricouté microvertebrate sites (Figures 1 and 2)
Geological map of the Petites Pyrénées (Haute-Garonne, France) with geographical (red rectangle and star) and stratigraphic location of the microvertebrate sites of Tricouté TRCT 1 and 4 (modified from Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997). The Tricouté sites belong to the middle part of the Auzas Marls Formation which is dated to the late Maastrichtian.

View of the microsite Tricouté 4 from the Maastrichtian of the Petites Pyrénées discovered in 2001 (photo of the 2002 excavation) (a) and stratigraphic section (b) in the silico-clastic series (sandstones and marls rich in intraclasts and bioclasts) from the fluviatile channel which outcrops at the site and which belongs to the middle part of the Auzas Marls Formation. In this locality, about two tons of sediment were sampled in the fossiliferous level indicated by the symbol Ŧ and were processed in laboratory by acid attack for microvertebrate remains research. Caption of the lithological facies seen in the Tricouté 4 section (B): 1: Thin sandstones, ferruginous in places; 2: Sandstones rich in intraclasts and cross-bedded in places; 3: Cross-bedded sandstones; 4: Fine sandstones; 5: Marls; 6: Limestones; 7: Microconglomerate; 8: Microvertebrate bioclasts. Ŧ: fossiliferous level rich in microremains of vertebrates.

The microvertebrates reported and studied herein were found in the site of Tricouté 1 (TRCT 1; see Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997), and in the new site of Tricouté 4 (TRCT 4), both of which are located on the Cassagnau Hill, close to the city of Marignac-Laspeyres (Department of Haute-Garonne, France). These fossil localities belong to the Southern flank of the Aurignac Anticline (Figure 1) and the structural area of the Petites Pyrénées and the Sub-Pyrenean Zone. The geological and stratigraphic context of the late Cretaceous deposits in the Sub-Pyrenean Zone, especially in the Aurignac Anticline, was described in detail by Babinot et al. (Reference Babinot, Freytet, Amiot, Bilotte, de Lapparent de Broin, Colombo Piñol, Durand, Feist, Floquet, Gayet, Lange-Badré, Masriera, Massieux, Medus, Tambareau, Ullastre and Villatte1983), Bilotte (Reference Bilotte1990, Reference Bilotte1994), Bilotte and Andreu (Reference Bilotte and Andreu2006), Bilotte et al. (Reference Bilotte, Tambareau and Villatte1983), Laurent et al. (Reference Laurent, Bilotte and Le Loeuff2002), Lepicard (Reference Lepicard1985), and Lepicard et al. (Reference Lepicard, Bilotte, Massieux, Tambareau and Villatte1985).
The microvertebrate-bearing levels of the Tricouté sites belong to the Auzas Marls Formation, which in turn belongs to a local Late Cretaceous paralic sedimentary series dated as Campano-Maastrichtian. In the Petites Pyrénées, the Maastrichtian sedimentary series includes indeed successively, from the base to the top, the Saint Martory Marls Formation, the Nankin Limestones Formation and the Auzas Marls Formation (Lepicard et al. Reference Lepicard, Bilotte, Massieux, Tambareau and Villatte1985; Bilotte and Andreu, Reference Bilotte and Andreu2006). This series overlies the Campanian formations of the Labarre Sandstones and the Lower Red Marls, both of which occur in the Mas d’Azil area (Plantaurel massif). At the top of the Cassagnau Hill, the Auzas Marls beds are overlain by the Cassagnau evaporites and the Tucauou lacustrine limestones which belong to the lowermost part of the Danian Aurignac Limestone Formation.
The deposits of the Auzas Marls Formation which outcrop in the Petites Pyrénées are about 100 metres thick, and their sedimentological facies gradually evolve upward from marine coastal and paralic to continental (limnic). They yielded a diverse fossil assemblage including marine, freshwater and terrestrial species of algae, invertebrates and vertebrates. The Auzas Marls Formation comprises the last sedimentary deposits of Cretaceous age from the Sub-Pyrenean Zone. Its late Maastrichtian age is well constrained based on its microflora and microfauna contents, with the occurrence of several marker species of charophytes (Clavatoraceae such as Septorella brachycera and Septorella ultima), foraminifera (Laffiteina mengaudi, Hellenocyclina beotica), ostracods (Paracandona krsticae), rudists and nannoflora (Massieux et al. Reference Massieux, Tambareau and Villatte1979; Bilotte, Reference Bilotte1985, Reference Bilotte, Graciansky De, Hardenbol, Jacquin and Vail1998; Lepicard, Reference Lepicard1985; Lepicard et al. Reference Lepicard, Bilotte, Massieux, Tambareau and Villatte1985; Tambareau et al. Reference Tambareau, Hottinger, Rodriguez Lazaro, Villatte, Babinot, Colin, Garcia Zarraga, Rocchia and Guerrero1997; Laurent et al. Reference Laurent, Bilotte and Le Loeuff2002). The Maastrichtian age is also supported by large benthic foraminifers occurring in the underlying Nankin Limestone Formation, such as Orbitoides apiculata, Lepidorbitoides socialis, and Siderolites calcitrapoides (see Lepicard, Reference Lepicard1985).
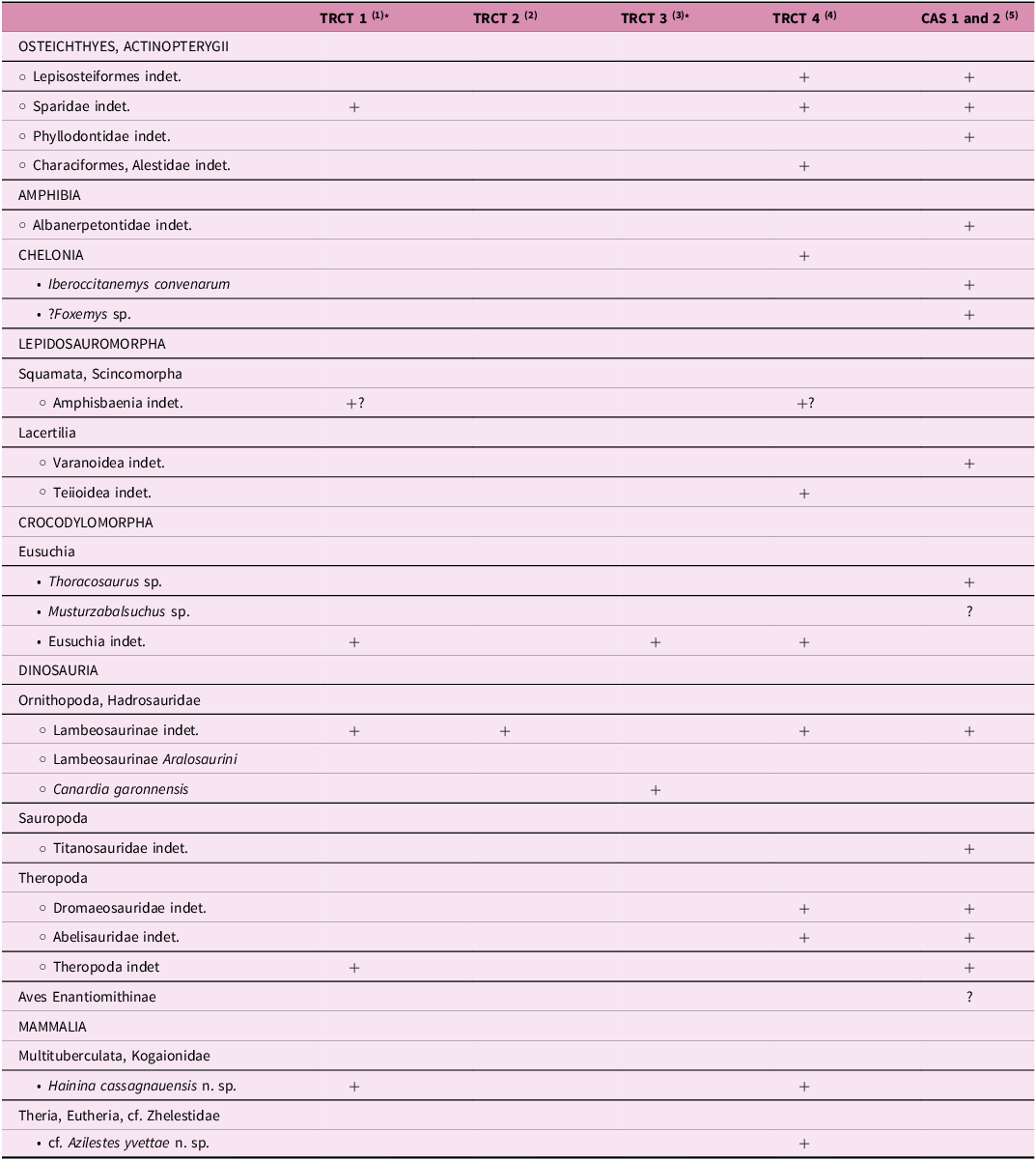
4. Vertebrates from the Auzas Marls formation
The Auzas Marls Formation has yielded one of the richest vertebrate assemblages known from the latest Cretaceous of southern France (Laurent et al. Reference Laurent, Bilotte and Le Loeuff2002). It comprises both macrovertebrates and microvertebrates found at several sites (see Table 2) in the Petites Pyrénées, in particular on the Cassagnau Hill. They include selachians (Palaeogaleus sp., Rhinobatos sp., and especially the Maastrichtian species Rhombodus binkhorsti), osteichthyans (Lepisosteiformes, Phyllodontidae, Sparidae), amphibians (Albanerpetontidae), turtles (the non-marine Bothremydidae Iberoccitanemys convenarum and ? Foxemys), squamates (?Scincomorpha, ?Amphisbaenia, and Varanoidea), early branching eusuchian crocodilians (Thoracosaurus sp. and ?Musturzabalsuchus sp.), non-avian dinosaurs (Theropoda, Titanosauridae, and Hadrosauridae such as Canardia garonnensis), pterosaurs (Azhdarchidae), birds (?Enantiornithines) and undetermined mammals (Buffetaut et al. Reference Buffetaut, Laurent, Loeuff and Bilotte1997 a, b; Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997; Laurent et al. Reference Laurent, Buffetaut and le Loeuff2000; Laurent et al. Reference Laurent, Bilotte and Le Loeuff2002; Prieto-Márquez et al. Reference Prieto-Márquez, Dalla Vecchia, Gaete and Galobart2013).
Updated faunal list of the vertebrates from the Auzas Marls Formation recovered at the Cassagnau Hill (Petites Pyrénées, France). TRCT: Tricouté; CAS: Cassagnau; *microvertebrate sites. (1) Gheerbrant et al. (Reference Gheerbrant, Abrial and Cappetta1997); (2) Laurent (Reference Laurent2003); (3) Laurent (Reference Laurent2003), Prieto-Márquez et al. ( Reference Prieto-Márquez, Dalla Vecchia, Gaete and Galobart2013); (4) this work; (5) Laurent et al. (Reference Laurent, Bilotte and Le Loeuff2002), Laurent (Reference Laurent2003), Brochu (Reference Brochu2004), Pérez-García et al. (Reference Pérez-García, Ortega and Murelaga2012)
4.a. The Tricouté microvertebrate sites
The first discovery of microvertebrates in the Auzas Marls Formation was reported by Gheerbrant et al. (Reference Gheerbrant, Abrial and Cappetta1997) at several sites at the Cassagnau Hill in the Petites Pyrénées, in particular at the Tricouté 1 site (TRCT 1). Preliminary prospecting and excavations conducted in TRCT 1 in 1992 and 1994 sampled approximately 200 kg of sediment, yielding a few teeth of undetermined species and genera of osteichthyans, crocodilians, squamates, and non-avian dinosaurs (Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997). New field research conducted by one of us (EG) in 2001 and 2002 included, in particular, new sampling at the site TRCT 1. It also led to the discovery of the new microvertebrate site Tricouté 4 (TRCT 4). This site corresponds to a lens of fossiliferous sandstones of a fluviatile channel whose lithological and sedimentological facies are identical to those of TRCT 1. TRCT 4 was excavated and intensively sampled in the search for microvertebrates (see Section 2. Material and Methods, and Figure 2a).
The two nearby microvertebrate sites TRCT 1 and TRCT 4, about 100 metres apart, correspond to two very localized outcrops of a microvertebrate-bearing cross-bedded sandstone lens. They belong to the detrital deposits of fluviatile channels that are gullying versicolor clays of palaeosols. These channel deposits are made of sandstones (with carbonate cement) and alternating clayey marls, silty or sandy marls, and calcareous marls rich in intraclasts, lithoclasts and bioclasts, the latter mostly consisting of bone and teeth debris (Figure 2b). The maximum visible thickness of the detrital channel levels in the TRCT 4 site is of about 0.5 m (Figure 2). They are intercalated in a paralic series of marls and limestones that forms most of the Auzas Marls Formation thickness, and they belong to the middle part of the Auzas Marls Formation. More precisely, the microvertebrate levels of TRCT 1 and TRCT 4 most likely belong to the sedimentary facies ‘PC4’ described by Lepicard (Reference Lepicard1985), which corresponds to the ‘distant paralic to continental domain’ and is located in the Cassagnau section at the top of the ‘Lower Ensemble’ of the Auzas Marls Formation (see Lepicard, Reference Lepicard1985: figures 40 and 46).
4.b. The Tricouté microvertebrate assemblage (Table 2)
The microvertebrate material recovered in the Tricouté sites includes dental and bone remains of bony fishes, amphibians, squamates, crocodilians, dinosaurs (incl. juvenile individuals) and mammals. They are associated with macroremains of turtles, crocodyliforms and dinosaurs (Table 2). Vertebrates are also associated with rare plant and invertebrate microfossils including charophytes oogons and benthic foraminifer tests.
Among the new assemblage recovered from TRCT 4, the actinopterygians are represented by small button-shaped teeth of a durophagous sparid and by sharp conical teeth of a carnivorous species of an undetermined taxa. Isolated rhombic-shaped scales suggest the presence of Lepisosteiformes. One small tooth (specimen with lab numbering TRCT 4-13) from TRCT 4 represents a characiform fish of the family Alestidae (Figure 3a–c). It is an incomplete lower tooth bearing a crown inflated on the lingual side and flat on the labial side. It preserves three sharp labio-lingually compressed cusps with a sharpening facet on the labial side (flat side).
Non-mammalian microvertebrate fauna from the Maastrichtian of the Tricouté 4 microsite (TRCT 4, Petites Pyrénées, France). (a–c) TRCT 4-13, Osteichthyes, Characiformes, Alestidae indet., lower tooth (from dentary) in occlusal, lingual and labial views (s.e.m. photographs). (d–f) TRCT 4-14, Squamata, Teiiodea, ?Barbatteiidae indet., right lower jaw in lingual (d), labial (e) and occlusal (f) views. (g–h), TRCT 4-15, Squamata, ?Amphisbaenia, isolated vertebra in ventral (g) and dorsal (h) views.

The herpetofauna of TRCT4 is quite diverse and is known from numerous microfossils (Table 2). Unfortunately, most of these microfossils are fragments of larger teeth or bones. This is particularly true for archosaurs, which are mainly represented by numerous fragments of the teeth of indeterminate eusuchian crocodiles, hadrosaurs, abelisaurids and, much more rarely, dromaeosaurids. Fossil remains of the microherpetofauna are scarcer. A new discovery is a vertebra that supports the presence of amphisbaenians (Figure 3g–h). Another one is that of an incomplete squamate dentary, specimen TRCT 4-14 (Figure 3d–f). Its posterior part and the symphysis are broken away, and only eight tooth positions are preserved in the specimen. The dentary TRCT 4-14 has a low profile and tapers anteriorly. It is slightly curved both lingually and dorsally. When preserved, the labial surface of the dentary is smooth and three large mental foramina arranged in a single row are preserved (Figure 3e). The Meckelian canal runs the full length of the preserved bone and is widely open lingually. The teeth are pleurodont in their mode of attachment. The only preserved tooth is cylindrical but worn, making any definitive assignment complicated. All these characters suggest close relationships, within the Laterata, with either the Teiioida or the Polyglyphanodontia (DeMar and Breithaupt, Reference DeMar and Breithaupt2006; Longrich, Bhullar and Gauthier, Reference Longrich, Bhullar and Gauthier2012; Xing et al. Reference Xing, Niu and Evans2023). In the absence of skull roofing bones, it is not possible to make a definitive attribution of the dentary TRCT 4-14. However, its strong resemblance to the dentary identified by Codrea et al. (Reference Codrea, Venczel and Solomon2017 a) as a Barbatteiidae – a group of Teiioidea only known from the Maastrichtian of Romania – supports at least its teiioid relationship.
In this paper, we detail especially the study of the micromammals recovered in the Tricouté sites TRCT 1 and TRCT 4, which are represented by a sample of eleven dental specimens (Supplementary Table S1).
5. Systematic palaeontology
Class MAMMALIA Linnaeus, Reference Linnaeus1758
Subclass ALLOTHERIA Marsh, Reference Marsh1880
Order MULTITUBERCULATA Cope, Reference Cope1884
Suborder CIMOLODONTA McKenna, Reference McKenna, Luckett and Szalay1975
Family KOGAIONIDAE Rădulescu and Samson, Reference Rădulescu and Samson1996
Genus HAININA Vianey-Liaud, Reference Vianey-Liaud1979
Type species. Hainina belgica Vianey-Liaud Reference Vianey-Liaud1979 from the middle Paleocene of Hainin, Belgium.
Other species: H. pyrenaica Peláez-Campomanes et al. Reference Peláez-Campomanes, López-Martínez, Álvarez-Sierra and Daams2000; H. godfriauxi Vianey-Liaud, Reference Vianey-Liaud1979; H. vianeyae Peláez-Campomanes et al. Reference Peláez-Campomanes, López-Martínez, Álvarez-Sierra and Daams2000.
Hainina cassagnauensis n. sp. Gheerbrant
Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 1 (TRCT 1, Petites Pyrénées, France). Right P1 MHNT.PAL.2024.2.2. (a) occlusal view (stereo views). (b) Lingual view; (c) Labial view; (d) Distal view. (e) Mesial view. s.e.m. micrographs.

Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 1 and 4 (TRCT 1 and 4, Petites Pyrénées, France). (a–c). Right P3 MHNT.PAL.2024.2.1 (TRCT 1 site), in lingual (a), labial (b) and occlusal (stereo views) (c) views. (d) MHNT.PAL.2024.2.6 (TRCT 4 site), fragment of left ?P4 (or ?M1) in occlusal (stereo views) views. s.e.m. micrographs. The arrow indicates the orientation of the tooth row.

Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 4 (TRCT 4, Petites Pyrénées, France). (a) Right M1 MHNT.PAL.2024.2.4 in occlusal (stereo views) views. (b) Right M1 MHNT.PAL.2024.2.3, holotype, in occlusal (stereo views) views. (c) Details of the distal flank of MHNT.PAL.2024.2.3, showing subvertical wear grooves (arrows). (a–b) s.e.m. micrographs; (c), image from 3D digital model reconstructed from micro-CT scan. The arrow indicates the orientation of the tooth row.

Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 4 (TRCT 4, Petites Pyrénées, France). 3D digital models of the M1, reconstructed from micro-CT scans, showing the morphology of the pulp cavity (in green) and of the dentine (in yellow) at the base of the crown. (a–c), M1 MHNT.PAL.2024.2.3, holotype; (d–f), M1 MHNT.PAL.2024.2.4. (a–b), (d–e): occlusal views of the crown; (c, f): dorsal views of the tooth with roots broken apart, exposing the pulp cavity. Note the presence of well-distinct ‘pulp horns’, corresponding to digitation (extension) of the pulp cavity in the dentine in cone-shaped pits (in green) beneath the cusps. The enamel is depicted in red, and the dentine in yellow. c1-4: the four cusps of the central cusp row. ph: ‘pulp horns’ in the dentine.

(a–f) Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 4 (TRCT 4, Petites Pyrénées, France). (a–c) MHNT.PAL.2024.2.7, distal part of a right P4 in labial, lingual and occlusal views. (d–f) MHNT.PAL.2024.2.8, fragment of P4 in lateral (d, f) and occlusal view. G-I. Hainina cassagnauensis? MHNT.PAL.2024.2.10, possible lower incisor I1 in oblique lateral view. H-I, MHNT.PAL.2024.2.9, broken upper incisor (I2 or I3) in labial and lingual views. s.e.m. micrographs.

Hainina cassagnauensis n. sp. (Multituberculata, Kogaionidae) from the Maastrichtian of Tricouté 1 and 4 (Petites Pyrénées, France). Occlusal sketches of isolated teeth (in white) and tentative composite reconstruction of the upper premolar and molar series (with MHNT.PAL.2024.2.6 interpreted as a possible fragment of P4). (a) Tentative reconstruction of the right upper premolar and molar series, with P1 (MHNT.PAL.2024.2.2), P3 (MHNT.PAL.2024.2.1), ?P4 (MHNT.PAL.2024.2.6, in reversed view), M1 (MHNT.PAL.2024.2.3, holotype); (b) Right M1 MHNT.PAL.2024.2.4. (c) Tentative reconstruction of P4 (right tooth) from the fragmentary specimens MHNT.PAL.2024.2.7 and MHNT.PAL.2024.2.8. Teeth are aligned along the cusp rows axis. Shaded areas are reconstructed missing parts. The arrow indicates the orientation of the tooth row.

LSID urn:lsid:zoobank.org:act:ED374C34-1508-4FBA-86E1-8966DE4E28F4
Holotype. Specimen MHNT.PAL.2024.2.3 from TRCT 4 site (Figures 6b and 7a–c; Figure S1), isolated right M1, collection of the MHNT (Toulouse).
Type locality. Tricouté 4 (TRCT 4), Cassagnau Hill, Petites Pyrénées, Haute-Garonne, France.
Type horizon and age. Auzas Marls Formation, late Maastrichtian, latest Cretaceous.
Paratypes. MHNT.PAL.2024.2.1 (Figure 5a–c; Figure S3) from TRCT 1 site, right P3; MHNT.PAL.2024.2.4 (Figure 6a; Figure S2) from TRCT 4 site, right M1.
Referred material. MHNT.PAL.2024.2.2 (Figure 4) from TRCT 1 site, right P1; MHNT.PAL.2024.2.6 (Figure 5d) from TRCT 4 site, distal half of ?P4 (or less likely ?M1); MHNT.PAL.2024.2.7 (Figure 8a–c) from TRCT 4 site, distal part of a right P4; MHNT.PAL.2024.2.8 (Figure 8d–f) from TRCT 4 site, fragment of P4.
Tentatively referred material (H. cassagnauensis?). MHNT.PAL.2024.2.9 (Figure 8h–i) from TRCT 4 site, broken upper incisor (I2 or I3); MHNT.PAL.2024.2.10 (Figure 8g) from TRCT 4 site, broken lower incisor (I1).
Etymology. Species name derived from the geographical name of the Cassagnau Hill, where the type locality Tricouté TRCT 4 is located.
Diagnosis. A medium-sized kogaionid species, most closely related in size and morphology to the genus Hainina. It shares with Hainina the P3 shorter than M1 and with tooth cusp formula (3:5) within the same range (1–3:2–5), the M1 with high tooth cusp formula (3:4:5), especially in the lingual cusp row which is extended to the mesial margin of the tooth, and the lingual cusp row of M1 decreasing in height anteriorly. Among the species of Hainina, the dental morphology of H. cassagneauensis n. sp. is closer to H. pyrenaica, especially for P3 and M1. H. cassagneauensis n. sp. differs from H. pyrenaica by a larger size, the teeth with weakly wrinkled enamel, the M1 with better individualized cusps in the lateral rows, and the P3 with wider central valley. It differs from H. belgica by larger size and lower cusp formula of M1 (lingual row). It differs from H. vianeyae by smaller size and the M1 that is wider and has lingual cusp row less developed. It differs from H. godfriauxi by smaller size, shorter M1, narrower and more triangular P1.
5.a. Description
5.a.1. Upper dentition
P 1 . The specimen MHNT.PAL.2024.2.2 is a right P1 (Figure 4), the only known anterior upper premolar of multituberculate in the localities of Tricouté. Its morphology is close to that of the P1 of kogaionids, particularly of Hainina. Within the known sample of multituberculate teeth from Tricouté, this tooth departs by its large size (Figures 9 and 10). Its specific allocation is discussed below.
Graph showing the relative tooth dimensions (length (L) x width (W)) for the upper postcanine teeth (ranked by tooth locus) of H. cassagnauensis n. sp. from the Maastrichtian of Tricouté (Petites Pyrénées, France) and other kogaionid multituberculates (mean for each dental locus). Based on data from Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022; Table 3).

The occlusal outline of the crown of MHNT.PAL.2024.2.2 is triangular and relatively wide (length/width ratio = 1.16) by comparison to other kogaionids, being closest to H. pyrenaica. This is likely correlated to the reduction of the posterior shelf. The labial side is more inflated than the lingual side, and this is illustrated by the apex of the labial cusp that is significantly shifted lingually (Figure 4d,e). The tooth is very bunodont with the presence of three large bulbous cusps, one on the labial side, two on the lingual side (cusp formula 1:2). The anterior cusp is the smaller and lower one. Its mesial flank is significantly canted distally. The disto-lingual cusp is only a little smaller and lower than the disto-labial cusp. The tooth is worn on the labial side at the cervix level. The cusps almost lack distinct vertical ridges, and the enamel is not wrinkled but smooth. Only one small vertical crest is present between the two lingual cusps, and it is more marked on the distal flank of the mesial cusp. There is no distal shelf or cingulum. The wear stage of the cusps is poorly advanced. In particular the apex of the cusps is still pointed, with little abrasion.
The tooth has two roots which are fused at apex, all along the preserved part in specimen MHNT.PAL.2024.2.2. The posterior root is larger than the anterior one. Its distal flank is slightly concave. CT scan observations provide some details on the root and the internal structure of the tooth. The anterior root has a small pulpar canal that is significantly compressed laterally. The posterior root bears a larger pulpar canal that is compressed antero-posteriorly. The two pulpar canals of the roots are widely confluent on a significant height below the crown, as are the roots. The enamel is thin (enamel preserved volume 0.18 mm3), whereas the dentine is very thick (dentine preserved volume 1.19 mm3), and it is much more voluminous (eight times larger) than the pulpar cavity and canal (preserved volume 0.13 mm3).
Dimensions: see Table 3.
Dimensions of the upper teeth of H. cassagnauensis n. sp. from the Maastrichtian of the Tricouté sites TRCT 1 and TRCT 4, Petites Pyrénées, France (in millimetres)
Comments. The morphology of MHNT.PAL.2024.2.2 is reminiscent of the kogaionid Hainina. However, the P1 MHNT.PAL.2024.2.2 is proportionally very large with respect to the P3 MHNT.PAL.2024.2.1 from the same locality (Figures 9a and 10), which could cast doubts as to whether it belongs to the same species. However, the size of MHNT.PAL.2024.2.2 remains within the range of the medium-sized kogaionids (Figure 10), like all other multituberculate teeth identified as H. cassagnauensis n. sp. in the Tricouté site. In addition, a high intraspecific variability is known in kogaionids such as Kogaionon, Barbatodon, Hainina godfriauxi and H. belgica (Codrea et al. Reference Codrea, Solomon, Venczel and Smith2017 b; De Bast and Smith, Reference De Bast and Smith2017; Solomon et al. Reference Solomon, Codrea, Venczel and Smith2022). For example, De Bast and Smith (Reference De Bast and Smith2017) have referred to Hainina godfriauxi a P1 that is large relative to P2 and other teeth of the species, based on their morphological affinity. We therefore consider that the larger size of some Tricouté specimens such as MHNT.PAL.2024.2.2 is related to a high intraspecific variation. Consequently, the P1 MHNT.PAL.2024.2.2 is here referred to the same species H. cassagnauensis n. sp. as the other multituberculate teeth found in Tricouté, and we identify only one species of multituberculate at the Tricouté site, based on known material (see Discussion).
P 2 . This tooth is unknown in the available material from Tricouté.
P 3 . This tooth is known by the single (right) specimen MHNT.PAL.2024.2.1 (Figure 5a–c). The labial and lingual margins of the crown are eroded by postmortem wear so that the elongated occlusal outline of the tooth (length/width ratio = 1.9) is a little exaggerated. The crown height decreases mesially. The labial flank is more inflated than the lingual flank that is more vertical. The enamel is weakly wrinkled by comparison to most other kogaionids: the vertical enamel ridges/crests radiating from cusp apex are few and poorly developed. There are two parallel cusp rows, with a cusp formula of 3:5. The three labial cusps are large and more pyramidal and inflated than the lingual cups, but they are slightly lower. The median labial cusp is the largest of the teeth. In the lingual cusp row, the four anterior cusps mesially decrease in height and volume, and the most distal cusp is the smallest of the tooth as in H. pyrenaica. The latter cusp is even smaller than in H. pyrenaica. The longitudinal valley between the two cusp rows is wide, flat and more widely open mesially. The mesial part of the tooth is broken, so it is not possible to check the presence of a cingulum joining the two anterior cusps. There are two roots. The anterior root is much larger (longer) than the posterior root.
There is no well-distinct wear facet. The stronger wear occurs as apical abrasion on the third cusp of the lingual row. In contrast, the crown (enamel) is significantly abraded on both sides by postmortem erosion.
Comments. The tooth cusp formula 3:5 seen in MHNT.PAL.2024.2.1 is characterized by a large number of lingual cusps as in Hainina (1–4:5), in particular H. pyrenaica (3:5), and it is distinct from Kogaionon (2–3:3) and Barbatodon (2:3). However, the tooth cusps formula 2:5 of ‘Litovoi tholocephalos’ (=Barbatodon transyvanicus) implies some overlapping variation of the genus Barbatodon with Hainina.
?P 4 . The identification of P4 in the available material from Tricouté is uncertain. One broken specimen, MHNT.PAL.2024.2.6 (Figure 5d), might correspond to the distal part of a P4 (Figure 9a). It preserves four large bunodont cusps and an inflated cingulum. The position and development of the cingulum is most reminiscent of that observed on the disto-lingual side of P4 of kogaionid multituberculates such as H. pyrenaica (see López Martínez and Pelaez-Campomanes, Reference López Martínez and Pelaez-Campomanes1999). Such homology of the cingulum of MHNT.PAL.2024.2.6 would indicate it is a left P4. The labial cusps are high, more separated and slightly more distal than the labial ones also fit with a P4 of the left tooth row. Additionally, the preserved size and length-to-width ratio of the broken tooth MHNT.PAL.2024.2.6 are proportional to the M1 MHNT.PAL.2024.2.3, which is consistent with an adjacent tooth (Figure 9a and see below), i.e. a P4. Other possible tooth locus homologies of MHNT.PAL.2024.2.6 are with M1 (anterior part of left M1) and with M2 (anterior part of right M2). However, MHNT.PAL.2024.2.6 differs from M1 by the wide central valley and a more inflated cingulum, among other traits. The wide central valley resembles the M2 of kogaionids, as well as the position and development of the cingulum. However, the topographical relations of the cusps of MHNT.PAL.2024.2.6 are somewhat different, for instance the labial cusps of M2 are larger and more separate than the lingual ones, whereas the reverse is true in MHNT.PAL.2024.2.6 (assuming its homology with a left M2, anterior part). In addition, comparison of MHNT.PAL.2024.2.6 with the M1 MHNT.PAL.2024.2.3 (holotype) and MHNT.PAL.2024.2.4 (see below) shows that the tooth is too narrow to correspond to a M2 of the same species. Consequently, MHNT.PAL.2024.2.6 is tentatively identified and described here as a distal fragment of a left P4 (shown in reversed view for the tooth row reconstruction in Figure 9a). The cingulum is inflated disto-lingually so that the occlusal outline is bulging disto-lingually. The cingulum orientation is oblique with a strong vertical tilt. The tooth has two cusp rows separated by a wide (U-shaped) central valley. Two labial and two lingual cusps are preserved. The cusps are inflated and well-individualized. They do not display distinct wrinkles on the enamel which is remarkably smooth. The two lingual cusps are more coalescent than the labial ones which are separated by a deeper notch. The two labial cusps are larger. There is a low and inflated distal crest linking the two distal cusps. It bears small secondary cuspules. The occlusal outline of the preserved part of the crown is straight labially, but oblique, narrowing mesially on the lingual side, in relation with the presence of the inflated lingual cingulum. Dimensions of MHNT.PAL.2024.2.6 are provided in Table 3. The wear in MHNT.PAL.2024.2.6 occurs only as slight apical abrasion on the labial cusps (no wear on the lingual side) which are higher than the lingual cusps. The morphology and development of the cusps, the smooth enamel and the size of this tooth fit well with other teeth of Tricouté identified as H. cassagnauensis n. sp., and especially with the M1 of the holotype MHNT.PAL.2024.2.3 as a possible adjacent tooth (see Figure 9a). CT scan observation shows the enamel to be thick.
M 1 . This tooth locus is represented by the two right teeth MHNT.PAL.2024.2.3 (holotype) and MHNT.PAL.2024.2.4 (Figure 6, Figures S1–S2). The occlusal outline is wide and sub-rectangular with a high width/length ratio (W/L = 0.72). The crown is slightly wider distally than mesially. The distal flank is convex, whereas the mesial flank is straight. The cusps are broad and more or less pyramidal. The tooth has three cusp rows separated by two wide and deep longitudinal valleys. The middle cusp row is oriented obliquely with respect to the longitudinal axis of the tooth, being directed mesio-lingually, as in other kogaionids. It bears the largest cusps. The cusp formula is 3:4:5. In the lingual cusp row, the mesialmost cusp is poorly inflated, but its presence is supported by a thickening of the dentine and, as for other cusps, by the expansion of the pulp cavity beneath it (well distinct in MHNT.PAL.2024.2.4; Figure 7). The cusps of the labial row decrease in size distally, whereas the opposite is true for the lingual cusp row (except for the small posteriormost cusp). This relative height ratio in the cusp rows is considered representative of the species despite significant wear of the teeth, as it is observed in the two specimens known from Tricouté material and because this pattern is also seen on the dentine surface (see Anemone et al. Reference Anemone, Skinner and Dirks2012) of the 3D digital models of the teeth from which enamel is removed. The labial row of cusps is low, and in MHNT.PAL.2024.2.4, it is strongly worn so the cusps are poorly distinct. The labial row of cusps is shorter than the lingual one. The two disto-lingual cusps of the lingual cusp row are partially fused. The two most mesial cusps, of the labial and middle cusp rows, are linked by a curved cingular-like ridge of enamel (Figure 6a). Even considering the advanced wear of the two teeth, the enamel appears moderately wrinkled: the vertical enamel ridges radiating around the cusps are present, but they are few and discreet.
Dimensions of M 1 MHNT.PAL.2024.2.3 (holotype) and MHNT.PAL.2024.2.4: see Table 3.
CT scan observation shows that the pulp cavity in these M1s was large (wide) (Figure 7). A remarkable feature is that it deepens within the dentine into several small, cone-shaped accessory hollow digits, forming pulp cavity digitations beneath the cusps. These digitations of the pulp cavity in the dentine are known as ‘pulp horns’ (Figure 7). All cusps are indeed hollowed out at their base by a pulp horn in the dentine. The size of the ‘pulp horns’ (deepness and volume) is as a whole proportional to that of the cusps. For example, the ventral base of the crown displays at its centre four distinct ‘pulp horns’ in the dentine beneath the four cusps of the central row (Figure 7). They are more developed than in other cusp rows, in proportion to the cusps size. It should be noted that the vertical axis of the ‘pulp horns’ may be slightly misaligned and offset in relation to the cusp they feed; as a result, some of the ‘pulp horns’ appear slightly offset in with respect to the cusps when viewed occlusally through transparency. The ‘pulp horns’ beneath the cusps are more developed in deepness and volume in MHNT.PAL.2024.2.3 (holotype) than in MHNT.PAL.2024.2.4, probably in relation to the age difference of the two individuals (the crown of MHNT.PAL.2024.2.4 has thicker dentine). The morphology of the pulp cavity is indeed a useful tool for the study of the relative development and homology of the many cusps present on the multituberculate teeth. The enamel is thicker on the occlusal surface (especially above the cusps) than on the lateral flank of the tooth.
Wear. There is an interdental wear facet on the mesial side of the crown of the two specimens. In addition, the distal side of MHNT.PAL.2024.2.3 (holotype) displays a remarkable wear pattern with the presence of large subvertical wear grooves (7–8 groves, the length varying from 0.06 to 0.25 µm and inter-groove distance varying from 0.11 to 0.18 µm) (Figure 6c). The wear grooves extend a little on both sides of the base of the crown. Wear facets with thin subhorizontal striae are also present on the lingual side of the cusps of the middle and labial rows.
Tentatively referred upper incisor. MHNT.PAL.2024.2.9 is a broken fragment of an unworn undetermined incisor (Figure 8h–i). The crown is characterized by a flat or slightly convex labial surface and an inflated convex lingual flank that bears an inflated vertical carina at mid-length. Two lateral crests are present. This morphology suggests an upper incisor, either I2 or I3. The minimal length, width and height of the preserved crown in specimen MHNT.PAL.2024.2.9 are respectively 2.00, 1.05 and 3.47 mm. These dimensions are consistent with those of other multituberculate teeth from Tricouté. The profile of the crown viewed mesially or lingually is asymmetric with the development of a long vertical crest on one side (lateral?) and a shorter more oblique crest on the opposite side (medial?). There is an accessory lateral cuspule at the base of the shorter crest. The enamel is not restricted to a labial band, but covers the entire tooth and is thicker on the labial side than on the lingual side. The enamel surface is slightly rugose, which is more common on I3. In this fragmentary specimen, the enamel-dentine boundary and the root at the base of the crown are not preserved. The broken crown exposes the apex of a large pulp cavity.
5.a.2. Lower dentition
The lower teeth of multituberculates from the Tricouté sites are scarce and damaged. They are mostly represented by fragments of P4 and a possible lower incisor. Their assignment to the same kogaionid species represented by the upper teeth described above is primarily based on their similar and homogeneous size which indicates compatible occlusion.
P 4 . This tooth is represented by the two fragments MHNT.PAL.2024.2.7 (Figure 8a–c) and MHNT.PAL.2024.2.8 (Figure 8d–f), which are of comparable size and might belong to the same species. They have the cimolodontan arcuate and blade-like shape with apical serrations. MHNT.PAL.2024.2.8 preserves only two serrations which are larger than in MHNT.PAL.2024.2.7. This tooth fragment MHNT.PAL.2024.2.8 corresponds to a middle or anterior portion of the P4 (Figure 9c), which bears the largest serrations. The minimal width of the crown seen in MHNT.PAL.2024.2.8 is 1.25 mm. MHNT.PAL.2024.2.7 is a distal portion of a right P4. It preserves a well-developed labio-distal platform, forming about half the labial width of the crown in its distal area. However, it is short and does not extend mesially beyond the level of the last tooth serration. It is separated lingually from the rest of the crown by a distinct crescent-shaped groove. The apex of the labio-distal platform is truncated by a flat abrasion wear surface likely for a single cuspule. The tooth fragment bears four well-distinct apical serrations, whose size slightly decreases with anterior position. The serration axis is tilted distally. An enamel ridge runs mesio-ventrally down each serration on both sides of the tooth. The ridges are parallel and are more pronounced labially and anteriorly. The minimal height measured of the fragment of the P4 MHNT.PAL.2024.2.7 is 2.43 mm. The width of the distal part of MHNT.PAL.2024.2.7 (with labio-distal platform is 1 mm. The width of MHNT.PAL.2024.2.7 is slightly less than that of MHNT.PAL.2024.2.8, but the dimensions of these two fragments, as preserved, remain consistent with their belonging to the P4 of the same species.
A tentative composite reconstruction of the upper tooth row (postcanine teeth) of the multituberculate species from Tricouté is proposed in Figure 9.
Tentatively referred Incisor. MHNT.PAL.2024.2.10 is a mesialmost fragment of an multituberculate incisor of which the apex is truncated by an oblique wear facet. This incisor is characterized by a laterally compressed shape (Figure 8g) and by the enamel worn off laterally and reduced to a labial band. Together, these traits best evoke a kogaionid I1 (e.g. Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018). The maximal length preserved of the incisor fragment MHNT.PAL.2024.2.10 is 1.72 mm.
5.b. Comparisons and discussion of the relationships of Hainina cassagnauensis n. sp
The isolated multituberculate teeth herein described from the Tricouté sites are relatively homogeneous in size and morphology, consistent with the presence of a single species. They all share, for instance, a more or less weakly wrinkled enamel. Comparisons show specialized dental features which are shared with the European cimolodontan family Kogaionidae, and in particular with the genus Hainina (Peláez-Campomanes et al. Reference Peláez-Campomanes, López-Martínez, Álvarez-Sierra and Daams2000; Kielan-Jaworowska and Hurum, Reference Kielan-Jaworowska and Hurum2001; Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018; Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). According to Csiki-Sava et al. (Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018) and Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022), the most significant kogaionid features of the Tricouté species are the M1 short and wide, with a short middle cusp row bearing four cusps. Additional kogaionid features are the elongated P3 (specimen MHNT.PAL.2024.2.1), and the M1 with three rows of cusps and with a long lingual row of cusps (more of 50% the length of the middle row of cusps in Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018). The enamel reduced to a labial band seen in specimen MHNT.PAL.2024.2.10 has also been described as a feature of the kogaionids anterior incisor (Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Brusatte and Norell2018). One character of the kogaionids that remains to be checked in the Tricouté species is the presence of gigantoprismatic enamel (Carlson and Krause, Reference Carlson and Krause1985; Fosse et al. Reference Fosse, Radulescu and Samson2001). Within the Kogaionidae, the morphology of the species from Tricouté is unambiguously closer to that of the Paleocene genus Hainina from Western Europe, than to that of the Late Cretaceous genera Kogaionon and Barbatodon (incl. Litovoi, a junior synonym following Smith et al. Reference Smith, Codrea, Devillet and Solomon2022) from Eastern Europe. According to Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022: 18), a distinctive feature of Hainina seen in the Tricouté species is the complex tooth morphology, especially for P3 and M1 (main known teeth in the Tricouté species) which have a high number of cusps. This is especially true for the lingual cusp row of the M1 of the Tricouté species which is elongated and has five cusps unlike the Cretaceous kogaionids Kogaionon and Barbatodon. The occurrence of five lingual cusps in P3 also differs from Cretaceous kogaionids, except for the species B. transylvanicus (incl. ‘Litovoi tholocephalos’) which, however, differs from the Tricouté species in other distinctive characters. For instance, the P3 of the Tricouté species is much shorter relative to M1 and it has three labial cusps instead of two (one cusp more). In addition, the Tricouté species is much smaller and its M1 has a more rounded shape. Although it is acknowledged that the number of tooth cusps varies considerably in Kogaionidae (and other multituberculates), particularly in the premolars, we argue that the high number of cusps observed in several dental loci of the Tricouté species (at least P3 and M1) is significant and is a derived trait that relates it closer to Hainina than to other genera from the Cretaceous. Additionally, the Tricouté species is smaller than most Cretaceous kogaionids (Figure 9), with the exception of B. oardensis and K. radulescui which are the closer Cretaceous species in size. However, the Tricouté species remains well distinct from these small Cretaceous species by significant morphological characters, and especially by characters shared with Hainina (see above) such as the higher tooth formula of P3 and M1, and in particular the long lingual cusp row of M1. The closest species of Hainina to that from Tricouté is H. pyrenaica Pelaez-Campomanes et al. Reference Peláez-Campomanes, López-Martínez, Álvarez-Sierra and Daams2000, from the early Paleocene (early Danian, MP1–5) of the Southern Pyrenees (Tremp Basin, Spain), which was previously the oldest known species of Hainina. Both species share very similar upper premolars and molars, especially P3 and M1. This includes the inflated cusps, the P1 lacking a posterior talon and the P3 that bears a high number of lingual cusps. However, the species from Tricouté differs by a number of traits. The Tricouté species is larger in tooth size than in H. pyrenaica, with a surface (L × W) ratio of 136% and 124% for M1 and P3. A noticeable morphological difference of the Tricouté species is the higher cusp formula of the lingual row of M1 (five cusps vs. four cusps), even though a significant intraspecific variation in tooth cusp formula (especially lateral rows of M1) is known in kogaionids (De Bast and Smith, Reference De Bast and Smith2017; Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). Other distinctive characters are seen in the Tricouté species: smoother enamel, better individualized cusps in the two lateral rows of M1, and P3 with posterior cusp less protruding posteriorly and with cusps more compressed laterally resulting in much wider central valley. Although we note again that several of the differences between the two species could be explained by the high intraspecific dental variation known in kogaionids (Smith et al. Reference Smith, Codrea, Devillet and Solomon2022), we contend that the combination of all distinctive characters reported here, based on the available material, supports the presence of a new species of Hainina at the Tricouté site, which is named Hainina cassagnauensis n. sp. H. cassagnauensis n. sp. is among the largest species of Hainina, together with H. godfriauxi Vianey-Liaud, Reference Vianey-Liaud1979 from the middle Paleocene (MP1–5) of Hainin (Belgium) and H. vianeyae Pelaez-Campomanes et al., Reference Peláez-Campomanes, López-Martínez, Álvarez-Sierra and Daams2000 from the late Paleocene (MP6a) of Cernay (France).
5.c. Phylogenetic analysis of the relationships of Hainina cassagnauensis n. sp
We performed a cladistic analysis of the relationships of H. cassagnauensis among mutituberculates with TNT 1.6 (Goloboff et al. Reference Goloboff, Farris and Nixon2008). Using the characters matrix of Smith et al. (Reference Smith, Codrea, Devillet and Solomon2022), we were able to code only 12 characters out of a total of 130, based on the poorly known material from Tricouté. In this matrix (see Section 2.d), the state of character 55 (M1, distal cusps in lingual row) in H. pyrenaica and H. belgica was re-coded 0 (cusps decreasing in height anteriorly). Two rounds of standard analysis were developed with TNT 1.6, the first obtaining 90 MPTs, and the second obtaining by overflow 15 000 MPTs with length of 467 steps, consistency index of 41.5 and retention index of 76.4 (see Supplementary Text S1). In the resulting MPTs H. cassagnauensis is unambiguously resolved as a Kogaionidae and a relative of the genus Hainina (Figure 11). The two nodes of the Kogaionidae and Hainina are among the best supported (Bremer Index = 2) in the resulting MPTs. Our analysis supports the relationships of the species from Tricouté with the Kogaionidae and with Hainina, in accordance with our detailed comparative study. The relationship of H. cassagnauensis to the Kogaionidae is supported by three unambiguous (but homoplastic) synapomorphies seen in H. cassagnauensis, in addition to two possible synapomorphies which are optimized. Two synapomorphies relate to the morphology of M1: K54-4 (RI = 75), M1 cusp formula 3-5:2-4:2-7 and K58-2 (DELTRANS optimization, RI = 58), M1 cusp lingual row (‘M1 posterolingual wing’) with 3 or four 4 cuspules (transformed to state 3, i.e. 5 cuspules in H. cassagnauensis). The other Kogaionidae synapomorphy is the character 124-1 (RI = 75) corresponding to the elongated penultimate premolar (P3). The relationship of H. cassagnauensis to Hainina seen in resulting MPTs is supported by a single synapomorphy: character 55-0 (RI = 71), height of the cusps in the lingual row of M1 decreasing anteriorly. Within the genus Hainina, H. pyrenaica is sister group to both H. cassagnauensis and H. belgica, but this is based on a single homoplastic character (character 58-3, five or more cusps on the lingual row of M1 at the node of H. cassagnauensis and H. belgica).
Phylogenetic relationships of the multituberculate H. cassagnauensis n. sp. from the Maastrichtian of Tricouté (Petites Pyrénées, France). Consensus tree of 15 000 MPTs (overflow) obtained from cladistic standard analysis with TNT1.6. Length = 467; CI = 41.5; RI = 76.4. The resulting topology supports the relationships of H. cassagnauensis n. sp. to the Kogaionidae and to Hainina.

Subclass THERIA Parker and Haswell, Reference Parker and Haswell1897
Infraclass EUTHERIA Gill, Reference Gill1872
Family cf. ZHELESTIDAE Nessov, Reference Nessov1985
Subfamily incertae sedis (probably new)
Comment. The two genera Azilestes and Valentinella depart from all other Cretaceous eutherians by their remarkably derived dental morphology, which would require their inclusion in a new suprageneric taxon, at least a new subfamily within the Zhelestidae which includes their closer eutherian relatives. Because of the still poorly known material, we provisionally refer Azilestes and Valentinella to an unidentified subfamily of Zhelestidae.
cf. Azilestes yvettae n. sp. Gheerbrant
The new eutherian mammal cf. Azilestes yvettae n. sp. from the Maastrichtian of Tricouté 4 (TRCT 4, Petites Pyrénées, France). Holotype, specimen MHNT.PAL.2024.2.5, left M1 or M2 in occlusal view. (a) SEM stereo views; (b–c) 3D digital image reconstructed from microtomographies (CT scan); (b) unretouched specimen; (c) specimen reconstructed to its original shape by virtual reassembly of the fragmented parts.

Tentative reconstruction of the occlusion of the upper molar MHNT.PAL.2024.2.5 of cf. Azilestes yvettae n. sp. (holotype) with the lower molar (M1) of Azilestes ragei (holotype), based on occlusal sketches at the same scale. Both the morphology and size are consistent with a compatible occlusion of the two specimens and are in accordance with close systematic affinity. We note, however, that the best fit for occlusion with Azilestes ragei lower molar would be for a little larger upper tooth, by about 5%. It indicates that the holotype of cf. Azilestes yvettae n. sp. is slightly smaller than the holotype of Azilestes ragei.

Comparison of the occlusal sketches of the upper molar MHNT.PAL.2024.2.5 (holotype) of cf. Azilestes yvettae n. sp. (a) from the Maastrichtian of Tricouté with the fragment of upper molar UP-VLP-07-04 (b), a broken upper molar hypocone referred to Valentinella vitrollense from the Maastrichtian of Vitrolles.

LSID urn:lsid:zoobank.org:act:5D82C390-D45F-401E-B171-43C0C22C45F0
Holotype. MHNT.PAL.2024.2.5 (Figures 12, 14a), damaged left upper molar, M1 or M2
Type locality. Tricouté 4 (TRCT 4), Cassagnau Hill, Petites Pyrénées, Haute-Garonne, France.
Type horizon and age. Auzas Marls Formation, late Maastrichtian, latest Cretaceous.
Etymology. Species dedicated to the memory of our esteemed colleague and friend Yvette Tambareau (1938-2008), in particular for her contributions and extensive knowledge of the geology and stratigraphy of the Cretaceous and Palaeogene formations of the Petites Pyrénées, as well as her invaluable assistance with field research for microvertebrates in these formations.
Diagnosis. Upper molars basically tritubercular but with well-developed and individualized postcingulum and hypocones, the latter being high and pointed. Stylar shelf reduced. Bunodonty moderately developed: Cusps and crests still high and moderately sharp, protofossa vast and deep. Postmetacrista reduced. Conules labially set and with an internal crest (premetaconule and postparaconule cristae). Metaconule doubled. Hypocone flanked lingually by an accessory small hypoconular cusp.
Among Cretaceous mammals, cf. A. yvettae n. sp. is most closely related in tooth size and morphology to the European eutherians Azilestes ragei and Valentinella vitrollense. Together with these two species, it is among the largest Cretaceous eutherians. Together with Valentinella vitrollense, cf. A. yvettae n. sp. is the only known Cretaceous eutherian with a large and high hypocone. cf. A. yvettae n. sp. is only a little smaller than Azilestes ragei, and significantly smaller than V. vitrollense. It additionally differs from A. ragei by less developed bunodonty, especially in the narrow and still sharp protocone. It also differs from V. vitrollense (referred specimen UP-VLP-07-04) in the presence of a more individualized hypocone lingual lobe (hypocone and postcingulum well-developed and separated from protocone), the protocone narrower, and the hypocone more lingual.
5.d. Description of cf. Azilestes yvettae n. sp
The specimen MHNT.PAL.2024.2.5 is an anterior upper molar, either an M1 or an M2. It lacks the roots (broken) and its crown exhibits evidence of damage in the form of several cracks and breaks. The individual pieces of the crown separated from one another by cracks were digitally reassembled from a virtual 3D model generated from micro-CT scan of the specimen. The resulting reconstruction of the tooth is illustrated in Figure 12.
The occlusal outline of the tooth is slightly extended transversely (ratio L/W 77%) and more triangular than rectangular with a labial border longer (2.74 mm from anterior part of preparacrista to metastyle) than the lingual border (1.71 mm at level of hypocone and protocone). The distal flank of the tooth is slightly concave in occlusal view. When the tooth is aligned transversely along the paracone-protocone axis, the labial flank of the tooth appears oblique, which is a feature found in posterior molars, i.e. in M2 rather than M1. However, the transversely poorly extended occlusal outline is more reminiscent of the anterior molar M1.
The tooth is weakly bunodont, much less than for instance Protungulatum donnae Sloan and Van Valen, Reference Sloan and Van Valen1965: the cusp and crests are moderately sharp and high (including the protocone), and the protofossa is deep. The crown is basically tritubercular, but with well-developed postcingulum and hypocone. The conules are well distinct as inflated cusps, and they bear an internal crest. Although the mesostylar area is broken, the ectoflexus was probably weak based on the development of the ectocingulum on both of its sides. The stylar shelf is narrow to absent. It is reduced to a well-developed and inflated ectocingulum that is broken on the mesial part. The ectocingulum is inflated and high, in labial view it appears only a little lower than metacone height. The metastyle is small but distinct as a minute inflated cuspule (e.g. distal view), and the metastylar area is reduced. No other stylar cusps are distinct, but the mesostylar and parastylar area parts are broken. The ectocingulum is in connection with the postcingulum at the level of the metastyle, and both form a continuous belt around the labial and distal borders of the tooth.
The parastylar area is broken but it might have been salient mesio-labially (Figure 12). The ectoloph, i.e. preparacrista, centrocrista and postmetacrista, are aligned close to the longitudinal axis. Preparacrista and postmetacrista are short, with no horizontal segment. The paracone and metacone are high and a little compressed labio-lingually. The paracone is significantly larger and higher than the metacone. It is actually the largest cusp of the tooth. The metacone is slightly more lingually placed than the paracone resulting in an ectoloph slightly oblique with respect to the longitudinal axis. The paracone and metacone are set well apart to each other, with a wide notch in-between, although they are fused at their basis to about the mid-height of the metacone (labial view). As a result, the centrocrista is straight longitudinally (ectoloph rectodont) and high. In distal view, the metacone appears to be slightly inclined lingually, which lends it a convergent look with the protocone. The internal flank of the paracone and metacone, lingually to the centrocrista, is slightly concave. This is related to the presence at their lingual base of a small but distinct crest/carina, which is linked to the conules respectively via the preparaconule and postmetaconule cristae. A narrow paracingulum is present at the mesio-lingual base of the paracone. It does not extend labially beyond the lingual side of the paracone and the preparacrista. The metacingulum is even shorter and more lingual; it ends lingually far from the postmetacrista. The paraconule is inflated, but compressed mesio-distally with oval apical wear surface. Its base is confluent with the paracone, and it displays a slight postparaconule crista. The metaconule is doubled: it consists of two mesio-distally compressed cusps, which are closely appressed. The labial metaconular cusp is longer transversely, while the lingual one is larger and more inflated. The labial metaconular cusp bears the premetaconule crista. The two metaconular cusps are separated by a small subvertical notch of the distal flank. The notch separating the protocone from the metaconule (lingual cusp; see Supplementary Figure S4) is larger. The metaconule is separated from the metacone by the deep notch of the metacingulum. As a result, the postmetaconule crest does not join the base of the metacone.
The protofossa is deep, vast and wider than longer. The protocone is sharp and compressed mesio-distally, especially at apex. The protocone is nearly as high as the paracone. It is inclined labially, with a long lingual slope (see Luo, Reference Luo1991). It is also tilted mesially and placed anteriorly, much closer to the level of the paracone than to that of the metacone. Its lingual flank is not convex but concave, which contributes to the wide extension of the protofossa. The postcingulum is high (higher than mid-height of the crown) and extended labio-lingually from the metastyle to the posterior part of the protocone. It bears an apical crest, ending lingually by the hypocone. The postcingulum and its hypocone are separated from the protocone by a deep and wide intermediate groove. The unworn hypocone is high, narrow and pointed rather than bulbous. It is doubled with a much smaller and lower accessory lingual cusp. The main hypocone cusp is located at the lingual end of the postcingulum crest, slightly more lingual than the metaconule (lingual cusp). The accessory hypocone cusp is located lingually to the protocone. It is mesially shifted with respect to the main hypocone cusp, and it bears a small abrasive wear surface. The lingual part of the postcingulum and the hypocone together form a developed posterior lingual lobe that is well differentiated in the occlusal outline. Although not preserved, the precingulum was probably thin.
Wear. The tooth MHNT.PAL.2024.2.5 does not show significant functional wear (few and weak wear facets and striae), which is indicative of a young individual. Wear facets of the centrocrista for shearing with oblique cristid and hypocristid (facets 3 and 4 of Crompton, Reference Crompton, Kermack and Kermack1971; facets PA-d and ME-m of Schultz et al. Reference Schultz, Menz, Winkler, Schulz-Kornas, Engels, Kalthoff, von Koenigswald, Ruf, Kaiser, Kullmer, Südekum and Martin2018) are restricted to the apex of the postparacrista and premetacrista, respectively. A small wear facet occurs on the mesial flank of the preprotocrista, close to the apex of the protocone, for occlusion with distal part of metaconid (facet 5 of Crompton, Reference Crompton, Kermack and Kermack1971; facet PR-d of Schultz et al. Reference Schultz, Menz, Winkler, Schulz-Kornas, Engels, Kalthoff, von Koenigswald, Ruf, Kaiser, Kullmer, Südekum and Martin2018). The mesio-lingual flank of the paracone shows a few subvertical wear striae, indicative of a strong vertical component in the Phase I of the power stroke of mastication. Slight abrasion wear surfaces occur at the apex of the paracone, paraconule, and accessory hypocone.
Pattern of occlusion. Study of the morpho-functional occlusal pattern of the upper molar MHNT.PAL.2024.2.5 allows a short discussion and a tentative inference on the corresponding lower molar pattern (Crompton, Reference Crompton, Kermack and Kermack1971; Butler, Reference Butler1972; Crompton and Kielan-Jaworowska, Reference Crompton, Kielan-Jaworowska, Butler and Joysey1978; Butler, Reference Butler, Smith and Tchernov1992). The deep and vast protofossa best matches in occlusion with lower molar with high and large hypoconid and a large (wide) talonid. The long (with higher paracone than metacone) and sharp postparacrista, bearing a distinct wear facet 3 (Pa-d in Schultz et al. Reference Schultz, Menz, Winkler, Schulz-Kornas, Engels, Kalthoff, von Koenigswald, Ruf, Kaiser, Kullmer, Südekum and Martin2018), suggests a well-developed (long) and shearing oblique cristid. The rectilinear (longitudinal) centrocrista best occludes with a poorly oblique and mainly labial oblique cristid. The pointed hypocone should occlude with the paraconid and/or trigonid basin.
Dimensions. See Table 4.
Dimensions of cf. Azilestes yvettae n. sp. from the late Maastrichtian of the Tricouté 4 site, Petites Pyrénées (in millimetres). * estimated measurement

5.e. Comparisons of cf. Azilestes yvettae n. sp
The morphology of the specimen MHNT.PAL.2024.2.5 is unambiguously that of an eutherian upper molar. This upper molar is characterized by a basically tritubercular (tribosphenic) morphology associated with well-developed postcingulum and hypocone that form a well-separated and high hypocone lobe.
The specimen MHNT.PAL.2024.2.5 represents the first subcomplete mammalian upper molar with such a well-developed hypocone lobe to be discovered in the Late Cretaceous. The only other mammalian material known from the Late Cretaceous with well-individualized and high hypocone is the broken upper tooth (a lingual fragment) UP-VLP-07-04 from the late Maastrichtian of Europe (locality of Vitrolles-La Plaine), which was referred to the eutherian Valentinella vitrollense Tabuce et al. Reference Tabuce, Vianey-Liaud and Garcia2004 on the basis of its size (Tabuce et al. Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013). Upper molars with postcingulum and hypocone occur in some other late Cretaceous eutherian mammals such as Gypsonyctops and the Zhelestidae (Archibald and Averianov, Reference Archibald and Averianov2012). However, the hypocone is much smaller in these taxa, being reduced to a ‘hypocone shelf’ (i.e. flat and not inflated). Even early ‘condylarths’ such as Protungulatum (P. gorgun and P. donnae), Oxyprimus, Ragnarok, Mimatuta and Baiconodon and others have a weak, much smaller hypocone, generally associated with a lower postcingulum (e.g. Archibald, Reference Archibald1982; Luo, Reference Luo1991; Lofgren, Reference Lofgren1995; Dewar, Reference Dewar2003).
In addition, MHNT.PAL.2024.2.5 is characterized by a large size with respect to most known Late Cretaceous eutherians. Its size is close to that of the largest known Late Cretaceous eutherians such as V. vitrollense, A. ragei Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021), Protungulatum combsi Archibald et al. Reference Archibald, Zhang, Harper and Cifelli2011, Cimolestes magnus Clemens and Russell, Reference Clemens and Russell1965 and C. stirtoni Clemens, Reference Clemens1973.
Comparison with Azilestes ragei Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021
Comparison of MHNT.PAL.2024.2.5 with Azilestes ragei is difficult and indirect because this species is only known from the lower teeth and jaw. However, as illustrated in Figure 13, the occlusal pattern of the M1 (holotype) of A. ragei shows interesting relationships with that of MHNT.PAL.2024.2.5. Comparison of the occlusal sketches at the same scale of the upper molar MHNT.PAL.2024.2.5 and lower molar of A. ragei shows that they fit well both in size and morphology (see also above description of the pattern of occlusion), beyond a little smaller size of MHNT.PAL.2024.2.5. There are indeed significant related morpho-functional features. The hypoconid of the M1 of Azilestes ragei occludes well with the centrocrista and with the protofossa of MHNT.PAL.2024.2.5, and conversely the protocone of the upper molar MHNT.PAL.2024.2.5 occludes well with the talonid basin of M1 of A. ragei. Interestingly, simulation of the occlusion shows that the cingular-like hypoconulid and postcristid of the lower molars of A. ragei occlude well within the postcingulum valley between the postprotocrista and the hypocone of the upper molar MHNT.PAL.2024.2.5, suggesting that these two features of opposing molars are morphologically and functionally related, i.e. coupled during occlusion. In other words, it suggests that the cingular hypoconulid and postcristid of lower molars of A. ragei and the postcingulum and hypocone of upper molar MHNT.PAL.2024.2.5 are homologous functional (shearing) structures. These features evoke an early stage of a specialized distal loph-like cingulum. It should be noted that the lower molar occlusal pattern and wear pattern of A. ragei has already been interpreted as corresponding to upper molar with a developed and functional hypocone (see Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021), especially with features such as the cingular-like hypoconulid and postcristid, the presence of an incipient hypolophid and the presence of a wear facet on the distal side of the entoconid which occludes with mesial side of hypocone (wear facet ed-d of Schultz et al. Reference Schultz, Menz, Winkler, Schulz-Kornas, Engels, Kalthoff, von Koenigswald, Ruf, Kaiser, Kullmer, Südekum and Martin2018; wear facet 8 of Butler, Reference Butler1952.
Both the morphology and the size suggest indeed a close taxonomic affinity of the upper molar MHNT.PAL.2024.2.5 with A. ragei, at least at the generic level. However, MHNT.PAL.2024.2.5 displays some morphological differences, in addition to a little smaller size. The most noticeable is a less bunodont morphology of MHNT.PAL.2024.2.5, and in particular a protocone that is less robust (sharper and less inflated) by comparison to the hypoconid of A. ragei. These differences are significant at species level, although the comparison clearly needs to be further developed on additional and more complete material.
Comparison with Valentinella vitrollense Tabuce et al. Reference Tabuce, Vianey-Liaud and Garcia2004
Unfortunately, the detailed comparison of V. vitrollense is also difficult and limited because this species is known by a damaged material displaying no details or very little of the occlusal morphology of the teeth, and because the upper dentition is poorly known, only by an upper premolar and a fragment of upper molar with specimen UP-VLP-07-04 figured by Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013, Figure 7c,d). The large size of MHNT.PAL.2024.2.5 is shared with Valentinella, as with Azilestes. The referred broken upper molar UP-VLP-07-04 of Valentinella displays additional important resemblances such as a high hypocone and postcingulum. A high hypocone is unknown in other Late Cretaceous eutherian mammals, except possibly in Azilestes as inferred from our analysis of the occlusal pattern (see above). Other noticeable resemblances of Valentinella and MHNT.PAL.2024.2.5 are the high and labially inclined protocone and the long lingual slope of the hypocone.
Although our comparison with Azilestes is indirect, given that it is known only from lower teeth, it shows closer morphological affinity of MHNT.PAL.2024.2.5 with Azilestes than with Valentinella. V. vitrollense is larger with respect to the species represented by MHNT.PAL.2024.2.5, consistently with its larger size with respect to Azilestes ragei (Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). It remains difficult to give an exact estimation of the size difference of V. vitrollense and MHNT.PAL.2024.2.5, as the former is mostly known by lower teeth. Further comparison of the upper molar fragment UP-VLP-07-04 referred to V. vitrollense evidences several distinctive morphological features of MHNT.PAL.2024.2.5 (Figure 14). The most important is the hypocone (main and accessory cusps) larger and more separated from protocone by a wider and deeper postcingular valley: both the hypocone and postcingulum form a distinctive hypocone lingual lobe in MHNT.PAL.2024.2.5 not seen in V. vitrollense. This construction of the upper molar MHNT.PAL.2024.2.5 is actually more consistent functionally with the occlusal pattern of lower molars of Azilestes ragei (see above). Other differences of the specimen MHNT.PAL.2024.2.5 from the referred upper molar UP-VLP-07-04 of V. vitrollense include the followings: presence of an accessory hypocone, crown less robust and bunodont, protocone placed more mesially, labial crest of hypocone straight and not concave as in UP-VLP-07-04, hypocone located more lingual and less inclined labially, with apex slightly more lingual than that of the protocone (in UP-VLP-07-04 it its more aligned with protocone apex).
Comparison with Zhelestidae
Azilestes and Valentinella have been tentatively referred to the family Zhelestidae (Tabuce et al. Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013; Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). Their familial position remains uncertain because of their original specialized features such as the cingular-like hypoconulid and postcristid and the presence of a hypolophid in Azilestes, and the high hypocone and postcingulum in Valentinella (Tabuce et al. Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013; Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). In addition, the reduced dental formula of Azilestes markedly departs from that of the Zhelestidae and even from all known Cretaceous eutherian mammals.
At familial level, the basic molar pattern of MHNT.PAL.2024.2.5 most closely resembles the Zhelestidae (including both the Zhelestinae and Lainodontinae) among all known Late Cretaceous eutherians. The zhelestid morphological affinity of the upper molar MHNT.PAL.2024.2.5 lies especially in the narrow stylar shelf, reduced preparacrista, postmetacrista, stylar cusps, large protofossa, distinct paraconule and metaconule and located close to the paracone and metacone, shallow ectoflexus, high protocone with long and labially inclined lingual flank, presence of lingual cingulum and robust more or less bunodont crown. These characters are considered derived among eutherians (Nessov et al. Reference Nessov, Archibald and Kielan-Jaworowska1998; Gheerbrant and Astibia, Reference Gheerbrant and Astibia1999, Reference Gheerbrant and Astibia2012; Archibald and Averianov, Reference Archibald and Averianov2012). However, the upper molar MHNT.PAL.2024.2.5 differs from those of most zhelestids by specialized features shared with Azilestes and Valentinella, in particular the high hypocone and postcingulum (inferred in Azilestes ragei; see Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). Among the Cretaceous zhelestines from Central Asia, Eoungulatum most closely approaches the condition of the developed hypocone and postcingulum seen in MHNT.PAL.2024.2.5, though in a less advanced stage, and it also shares a labially inclined protocone (Nessov et al. Reference Nessov, Archibald and Kielan-Jaworowska1998). MHNT.PAL.2024.2.5 differs from zhelestids by other features such as the doubled metaconule, the presence of an accessory hypocone, the reduced postmetacrista, the absence of metacingulum behind the metacone and the less inflated protocone.
Comparison with ‘condylarths’
Besides the Zhelestidae, the specimen MHNT.PAL.2024.2.5 displays some advanced features known in ‘condylarths’ (stem euungulates), particularly in arctocyonids, such as the labially inclined protocone (e.g. Luo, Reference Luo1991). However, MHNT.PAL.2024.2.5 departs from early ‘condylarths’ in other important features, such as a larger and higher hypocone and postcingulum (shelf-like hypocone in early arctocyonids), reduced postmetacrista, larger and deeper protofossa, much smaller and less inflated protocone (especially at lingual basis), and less bunodont crown and cusps. Paradoxically, some (the first two) of these distinctive characters of MHNT.PAL.2024.2.5 are more advanced traits. It should be noted that the upper molar MHNT.PAL.2024.2.5 is slightly smaller than those of Protungulatum donnae, for which the body mass estimate is 225 g (Wilson, Reference Wilson2013).
5.f. Discussion of cf. Azilestes yvettae n. sp
Among the known Late Cretaceous eutherians mammals, the Tricouté species represented by specimen MHNT.PAL.2024.2.5 is most closely related to the European genera Azilestes and Valentinella tentatively referred to the Zhelestidae. The most remarkable (derived) shared feature of Azilestes, Valentinella and MHNT.PAL.2024.2.5 are the large size and the well-developed hypocone and postcingulum (inferred in Azilestes ragei). Further comparison suggests closer relationship of the Tricouté species with Azilestes than with Valentinella. This is indicated by a similar size and more developed and better individualized hypocone than in Valentinella. The well-developed hypocone in MHNT.PAL.2024.2.5 is morpho-functionally reminiscent of the cingular-like hypoconulid and postcristid seen in Azilestes (Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021): both features are more consistent in terms of the occlusion pattern. As a result, the size and pattern of occlusion of the tooth MHNT.PAL.2024.2.5 argue for referral of the Tricouté species to Azilestes. At the species level, MHNT.PAL.2024.2.5 differs from Azilestes ragei by a less bunodont morphology, especially in the narrow and still sharp protocone. Consequently, we identify MHNT.PAL.2024.2.5 as belonging to the new species most closely related to Azilestes: cf. Azilestes yvettae n. sp. The generic referral of the new eutherian species from Tricouté represented by MHNT.PAL.2024.2.5 is tentative and provisional, pending additional material for more direct comparison (i.e. comparison based on teeth of the same row, either upper or lower). In this respect, a key feature of the Tricouté species to be known is its tooth formula.
5.g. Phylogenetic analysis of cf. Azilestes yvettae n. sp
We investigated the phylogenetic relationships of cf. Azilestes yvettae n. sp. through a cladistic analysis using TNT 1.6 (Goloboff et al. Reference Goloboff, Farris and Nixon2008) and the character matrix of Gheerbrant and Teodori (Reference Gheerbrant and Teodori2021), used for the study of Azilestes ragei and which was modified from Archibald and Averianov (Reference Archibald and Averianov2012) and Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013) (see Material and Methods and Supplementary Text S2). In all our parsimony analyses, the relationships of cf. Azilestes yvettae n. sp. are recovered with low Bremer support, primarily due to the still limited knowledge, with many unknown characters being optimized.
Standard unconstrained analysis. Two rounds of standard analyses were developed without topological constraint, the first obtaining 60 MPTs and the second obtaining 10 000 MPTs by overflow. The resulting MPTs have a length of 2245 steps (CI = 25.3, RI = 57.7). The strict consensus tree (Figure 15a) places cf. Azilestes yvettae n. sp. as the sister group to Valentinella vitrollense within the clade Zhelestidae. In contrast, Azilestes ragei is not included in the Zhelestidae, but is placed as the sister group to the Glires (Rodentia + Lagomorpha). As previously commented by Gheerbrant and Teodori (Reference Gheerbrant and Teodori2021), such a topology relating Azilestes ragei to the Glires is based on some remarkable shared derived characters such as the reduced dental formula (two premolars) that are most likely convergent. In resulting MPTs, Lainodon is a stem Zhelestidae together with Sheikhdzheilia, and the Zhelestidae are the sister group of Paranyctoides as in previous studies (e.g. Archibald and Averianov, Reference Archibald and Averianov2012). The relationship of cf. Azilestes yvettae n. sp. to the Zhelestidae is here supported by several important synapomorphies: stylar shelf narrow (<25 % transverse width of molar crown; character 64-2, RI= 76); postcingulum developed, extended labially past the metaconule (character 97-2, RI= 70); ectoflexus narrow (character 75-2, RI = 71, ACCTRAN optimization); postmetacrista weak (character 82-2, RI = 75, DELTRAN optimization); conular region wide antero-posteriorly (character 90-2, RI = 80, DELTRAN optimization); protocone high, close in height to paracone and metacone (character 95-2, RI = 72, ACCTRAN optimization).
Phylogenetic relationships of the eutherian mammal cf. Azilestes yvettae n. sp. from the Maastrichtian of Tricouté (Petites Pyrénées, France). Consensus tree of 10 000 MPTs (overflow) obtained from the cladistic analysis with TNT1.6 of the character matrix of Gheerbrant & Teodori (Reference Gheerbrant and Teodori2021), that is modified from Archibald and Averianov (Reference Archibald and Averianov2012) & Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013). (a) Unconstrained standard analysis. Length = 2245; RI = 57.7; CI = 25.3. (b) Constrained standard analysis, in which the three Late Cretaceous European taxa Azilestes ragei, cf. Azilestes yvettae n. sp., and Valentinella are constrained to belong to the same clade. Length = 2251. RI = 57.6; CI = 25.3.

Standard constrained analyses. We carried out several other cladistic analyses on the same character matrix in order to further test and elucidate the relationships between Azilestes ragei, cf. Azilestes yvettae n. sp. and Valentinella vitrollense. To this end, the relationships of the European taxa have been constrained in different ways, both between themselves and with the Zhelestidae.
In the parsimony analysis constraining a sister group relationship of Azilestes ragei and cf. Azilestes yvettae n. sp. (two rounds of standard analysis, the first obtaining 120 MPTs and the second obtaining by overflow 10 000 MPTs with length of 2250 steps; see Supplementary Text S2), both species are recovered as sister group of the glires. This relationship to the glires is mainly the result of the convergence of the reduced tooth (premolar) formula seen in A. ragei (see Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). In this analysis, Valentinella is included, in a polytomy, within the Zhelestidae.
We developed another parsimony analysis constraining the grouping of Azilestes ragei, cf. Azilestes yvettae n. sp. and Valentinella in a same clade (two rounds of standard analysis, the first obtaining 30 MPTs and the second obtaining by overflow 10 000 MPTs with length of 2251 steps). In the strict consensus of resulting MPTs (Figure 15b), this clade is recovered within a polytomy together with the Eulipotyphla (Erinaceidae and Soricidae) and Mistralestes (Mistralestes, (Azilestes ragei, cf. Azilestes yvettae n. sp., Valentinella), Eulipotyphla). The main synapomorphies supporting such a relationship of Azilestes ragei, cf. Azilestes yvettae n. sp. and Valentinella to the Eulipotyphla correspond to a procumbent lower canine (character 26-1, RI = 80), moderately wide upper molar with a L/W ratio between 75 and 99% (character 63-1, RI = 70), a postcingulum extending to labial margin (character 97-3, RI = 73; with a reversal to state 97-2 in soricids) and an oblique cristid mesially ending posteriorly to the protoconid (character 115-3, RI = 69). The most important of these is the procumbent lower canine which, however, remains poorly known both in Azilestes ragei and cf. Azilestes yvettae n. sp. The relationship of the Late Cretaceous European eutherians to the Eulipotyphla obtained here is actually not supported by our detailed comparisons of the dental anatomy of both Azilestes ragei and cf. Azilestes yvettae n. sp. This topology differs from all our other analyses and is less supported and rejected here.
We also developed a parsimony analysis constraining the inclusion of the three European taxa Azilestes ragei, cf. Azilestes yvettae n. sp. and Valentinella within the clade Zhelestidae (two rounds of standard analysis, the first obtaining 30MPTs and the second obtaining by overflow 10 000 MPTs with length of 2253 steps). In the strict consensus of resulting MPTs (Figure 16a and Supplementary Text S2 – Constrained analysis 3), Azilestes ragei, cf. Azilestes yvettae n. sp., Valentinella and all the zhelestids are in polytomy, in a node that is sister group of Paranyctoides. In the majority rule tree, Azilestes ragei and cf. Azilestes yvettae n. sp. are sister group, and both are sister group successively to Lainodon and Sheikhdzheilia (Sheikhdzheilia (Lainodon (Azilestes ragei, cf. Azilestes yvettae n. sp.))) (see Supplementary Text S2, Constrained analysis 3). It supports the hypothesis that Azilestes belongs to the European zhelestid clade of the Lainodontinae. However, this is based on a single lainodontine character, the lingually set molar paraconid, still unknown in cf. Azilestes yvettae n. sp. In contrast, and surprisingly, Valentinella is placed in the Asian Zhelestinae instead of the European Lainodontinae (majority rule tree; see Supplementary Text S2, Constrained analysis 3). The relationships of cf. Azilestes yvettae n. sp. to the Zhelestidae is here based on the same synapomorphies (upper molar characters) as reported above for the standard unconstrained analysis. Although this analysis supports the generic relationship of the Tricouté species (specimen MHNT.PAL.2024.2.5) with Azilestes, this remains based on only one synapomorphy (character 62-1, molar cusps inflated) which is homoplastic, being also known in Valentinella!
Phylogenetic relationships of the eutherian mammal cf. Azilestes yvettae n. sp. from the Maastrichtian of Tricouté (Petites Pyrénées, France). Consensus tree of 10 000 MPTs (overflow) obtained from the cladistic analysis with TNT1.6 of the character matrix of Gheerbrant and Teodori (Reference Gheerbrant and Teodori2021) that is modified from Archibald and Averianov (Reference Archibald and Averianov2012) and Tabuce et al. (Reference Tabuce, Tortosa, Vianey-Liaud, Garcia, Lebrun, Godefroit, Dutour, Berton, Valentin and Cheylan2013). (a) Constrained standard analysis, in which the three European taxa Azilestes ragei, cf. Azilestes yvettae n. sp., and Valentinella are constrained to belong to the Zhelestidae. Length = 2253. RI = 57.5; CI = 25.3. (b) Constrained standard analysis, in which Azilestes ragei and cf. Azilestes yvettae n. sp. are constrained to belong to the Zhelestidae to the exclusion of Valentinella. Length = 2254. RI = 57.5; CI = 25.2.

Another analysis which constrains the inclusion of Azilestes ragei and cf. Azilestes yvettae n. sp., within the clade Zhelestidae to the exclusion of Valentinella, also supports the generic affinity of the Tricouté species with Azilestes, and the belonging of this genus to the European Lainodontinae clade of Zhelestidae (Figure 16b and Supplementary Text S2, Constrained analysis 4). The strict consensus tree resulting from two rounds of standard analysis (the first obtaining 30 MPTs and the second obtaining by overflow 10 000 MPTs with length of 2254 steps) recovers the following similar topology: (Lainodon (Azilestes ragei, cf. Azilestes yvettae n. sp.)). This is based on the same shared characters as for preceding analysis (62-1, inflated molar cups shared by Azilestes ragei and cf. Azilestes yvettae n. sp.; 107-1 molar paraconid lingual shared by Azilestes ragei, cf. Azilestes yvettae n. sp., and Lainodon). By contrast, in this analysis Valentinella is not related to the Zhelestidae, but placed as a stem carnivoran, together with Mistralestes, an unconvincing relationship based of the comparative anatomy of the known material.
Overall, our cladistic analysis best supports the relationship of the eutherians Azilestes ragei and cf. Azilestes yvettae n. sp. to the Zhelestidae. Interestingly, it also supports in some instances (constrained analyses of Azilestes within Zhelestidae) a generic relationship of the Tricouté species with Azilestes. However, the exact relationships of these two species, Azilestes ragei and cf. Azilestes yvettae n. sp., and in fact of all known Late Cretaceous European eutherians, to each other and within the family Zhelestidae, still need to be tested and clarified on additional and better preserved material.
Mammalia indet.
The microvertebrate material recovered from Tricouté also includes some very fragmentary mammalian specimens (at least three specimens) that are undeterminable. One of them, catalogued as MHNT.PAL.2024.2.11, is an anterior tooth of small size (minimum dimensions: H = 2.9 mm; L = 0.58 mm; W ∼ 0.8 mm), either an incisor or a canine (Figure 17). The tooth is tall, thin and slightly curved, and its crown is sharp and compressed laterally. It has a well-developed lateral wear facet. The tooth is eroded by deep and elongated pits which possibly are evidence of acid attack digestion (i.e. predation).
Mammalia indet. from the Maastrichtian of Tricouté 4 (Petites Pyrénées, France). (a–b) MHNT.PAL.2024.2.11, anterior tooth, canine or incisor. Lateral views. s.e.m. micrographs.

6. Discussion
The Tricouté localities from the Petites Pyrénées (Northern Pyrenees) have yielded some of the latest Mesozoic continental vertebrates from Europe. The fauna from Tricouté is characterized by the original association of multituberculate and eutherian mammals. Such an association was previously unknown in Western Europe, and Europe as a whole until a preliminary report in Serbia (Marković et al. Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025). Mammals from Tricouté include two species. The best-documented one is a new multituberculate, Hainina cassagnauensis n. sp., known by about ten more or less complete teeth. In addition, the Tricoute site 4 has yielded one upper molar of a poorly documented eutherian, that is identified as cf. Azilestes yvettae n. sp.
The multituberculate Hainina cassagnauensis n. sp. shares the specialized dental morphology of the endemic European family Kogaionidae, to which it is referred. Cretaceous kogaionids were previously only known in Eastern Europe, especially in the palaeogeographic area of the Hațeg Island (e.g. Benton et al. Reference Benton, Csiki, Grigorescu, Redelstorff, Sander, Stein and Weishampel2010). In Western Europe, kogaionids were previously only known after the K/Pg boundary, during the Paleocene, with the earliest occurrence represented by Hainina pyrenaica from the early Danian of the Spanish Pyrenees (Lopez-Martinez and Pelaez-Campomanes, Reference López Martínez and Pelaez-Campomanes1999). As a result, Hainina cassagnauensis n. sp. represents the earliest known species of the genus Hainina and the first known Late Cretaceous multituberculate from Western Europe. In addition, Hainina cassagnauensis n. sp. belongs to one of the only – if not the only known – continental vertebrate genus crossing the K/Pg boundary in Europe. The only other mentioned K/Pg vertebrate genus in Europe is the turtle Dortoka, which Cadena and Joyce (Reference Cadena and Joyce2015) considered Ronella (Gheerbrant et al. Reference Gheerbrant, Codrea, Hosu, Sen, Guernet, de Lapparent de Broin and Riveline1999) to be a junior synonym. The mammal lineage of the genus Hainina is documented at the K/Pg transition through the rapid succession of the species H. cassagnauensis n. sp. and H. pyrenaica in the local context of the Pyrenees. Actually, H. cassagnauensis n. sp. and H. pyrenaica are likely trans-K/Pg chronospecies, which represents one of the scarce documented instances in the fossil record of terrestrial vertebrates. Both the fossil record and current phylogenies argue for an origin of the kogaionids in the Cretaceous of Eastern Europe (Hațeg Island) (e.g. Csiki-Sava et al. Reference Csiki-Sava, Vremir, Meng, Vasile, Brusatte and Norell2022; Solomon et al. Reference Solomon, Codrea, Venczel and Smith2022). The species Hainina cassagnauensis n. sp. supports that the Western European genus Hainina originated from a stem kogaionid from the Hațeg Island by the end of the Cretaceous. Palaeobiogeographically, this is remarkably consistent with the assignment of the squamate dentary TRCT 4-14 from Tricouté to the family Barbatteiidae, previously only known from the Late Cretaceous of Romania: both the kogaionid Hainina and the barbatteiid squamate from Tricouté might have belonged to the same latest Cretaceous dispersal event between Eastern and Western Europe. It is interesting to note that the earliest kogaionids known in Western Europe are found in the Pyrenean region, which belonged to the Ibero-Armorican Island of the European Archipelago (=Tethyan Europe for some authors such as Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010). Later, in the Paleocene, Hainina expanded its distribution from the Pyrenees (Tremp basin, Spain) to Northern Europe (Hainin, Belgium; Cernay-Berru, France) and Southern Europe. This context suggests an initial dispersal of the stem group of Hainina into Western Europe along a southerly route through the European Archipelago, followed by a spread of Hainina through Western Europe. The occurrence of Hainina in the latest Paleocene of the Transylvanian Jibou-Rona sites (Gheerbrant et al. Reference Gheerbrant, Codrea, Hosu, Sen, Guernet, de Lapparent de Broin and Riveline1999) raises the hypothesis of a likely autochthonous origin of this genus in Eastern Europe, i.e. in the Hațeg Island, which is widely regarded as the centre of the initial radiation of the Kogaionidae family. The alternative hypothesis involves the biogeographic scenario of a later (Palaeogene) dispersal of the genus Hainina back to the kogaionid cradle of Eastern Europe (Csiki and Grigorescu, Reference Csiki and Grigorescu2001), which is much less likely (e.g. Solomon et al. Reference Solomon, Codrea, Venczel and Smith2022). Further study of the Hainina species from Jibou-Rona should provide conclusive evidence on the origin of this genus.
Although only known from an upper molar, cf. Azilestes yvettae n. sp. documents a new eutherian species characterized by an original specialized dental pattern. Comparison of cf. Azilestes yvettae n. sp. reveals closest morphological affinity with the poorly known latest Cretaceous genera Valentinella and especially with Azilestes, both of which are endemic European genera tentatively referred to the Zhelestidae. Valentinella and Azilestes share convergent ungulate-like specialized traits previously unknown in Cretaceous eutherians, such as a developed hypocone, incipient bilophodonty and a large size. It suggests that Valentinella and Azilestes belong to a new, at least suprageneric, clade. A well-developed hypocone was previously only known in Cenozoic crown placentals (e.g. O’Leary et al. Reference O’Leary, Bloch, Flynn, Gaudin, Giallombardo, Giannini, Goldberg, Kraatz, Luo, Meng, Ni, Novacek, Perini, Randall, Rougier, Sargis, Silcox, Simmons, Spaulding, Velazco, Weksler, Wible and Cirranello2013), but the general molar pattern of Valentinella and Azilestes is reminiscent of a zhelestid-like ancestral morphotype. European zhelestids such as the lainodontines are themselves derived from early Late Cretaceous Central Asian zhelestids as Sheikhdzheilia (Gheerbrant and Astibia, Reference Gheerbrant and Astibia1999; Gheerbrant and Astibia, Reference Gheerbrant and Astibia2012; Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). Whatever their familial relationships, the specialized species represented by cf. Azilestes yvettae n. sp., Azilestes ragei, and Valentinella vitrollense support a significant diversity of the latest Cretaceous European eutherians, despite their still scarce fossil record.
Both multituberculates and eutherians known from the late Cretaceous of Europe belong to old endemic clades that evolved from Early/Late Cretaceous Asian stem groups, respectively, eobaatarid-like stem cimolodontids and Sheikhdzheilia-like stem zhelestids (Gheerbrant and Astibia, Reference Gheerbrant and Astibia2012; Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). It implies for these mammals long ghost lineages extending over most of the Late Cretaceous in the European Archipelago (Aptian-late Campanian for the Kogaionidae; Cenomanian-Campanian for the Zhelestidae). In this respect, the kogaionid multituberculates and the zhelestid eutherians are ancient relict lineages in Europe, with an unknown early evolutionary history. Other long ghost lineages and remarkable relict taxa of non-mammalian vertebrates are well known in the Hațeg Island fauna (Pereda Suberbiola et al. Reference Pereda Suberbiola, Corral, Astibia Ayerra, Badiola Kortabitarte, Bardet, Berreteaga Escudero, Buffetaut, Delgado Buscalioni, Cappetta, Cavin, Díez Díaz, Gheerbrant, Ortega Coloma, Pérez García, Poyato Ariza, Rage, Sanz García and Torices Hernández2015; Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010), confirming the prevalent endemic context of evolution in the European Archipelago during the Late Cretaceous. The relationships of kogaionids and zhelestids to Asian stem groups suggest ancient palaeobiogeographic relations during the Cretaceous between central Asia and Europe and also within the European Archipelago. The best supported hypothesis is that they result from westward dispersals to the European Archipelago (Gheerbrant and Astibia, Reference Gheerbrant and Astibia2012; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015; Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). However, an alternative vicariant hypothesis cannot be yet ruled out and deserves to be mentioned here. The vicariant hypothesis assumes an Early Cretaceous ancestral stock with a wide Eurasian or even Laurasian distribution, which subsequently evolved and diverged in isolated insular areas of the European Archipelago during the Late Cretaceous. Few Early Cretaceous taxa with restricted Eurasian distribution are known, but Early Cretaceous European faunas such as Angeac-Charente (France) are consistently characterized by the predominance of higher clades (families) with Laurasian distribution (Allain et al. Reference Allain, Vullo, Rozada, Anquetin, Bourgeais, Goedert, Lasseron, Martin, Pérez-García, Fabrègues, Royo-Torres, Augier, Bailly, Cazes, Despres, Gailliègue, Gomez, Goussard, Lenglet, Vacant, Mazan and Tournepiche2022). As per this hypothesis, the vicariant groups inherited from the Laurasian ancestral stock belong to the Late Cretaceous ‘old European core’ (Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015). Such a palaeobiogeographic vicariant scenario may indeed explain the divergence of (1) the Asian Zhelestinae and the European Lainodontinae, both related to a Sheikhdzheilia-like stem group, and(2) the Hațeg kogaionid multituberculates from a basal stock of eobaatarid-like cimolodonts with wide distribution (Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). By the way, the genus Hainina tells a much later event in the evolution and palaeobiogeography of European kogaionids. Its origin is best explained by the dispersal of an ancestral kogaionid, most likely a stem Hainina species, from Eastern Europe at the end of the Cretaceous.
Another issue raised by our new fossil discoveries concerns the previously known segregated palaeobiogeographic pattern of the mammals in Europe during the Cretaceous, with the multituberculates restricted to Eastern Europe and the eutherians to western Europe. The discovery reported here of kogaionids from the Cretaceous of Western Europe breaks this picture, showing for the first time the sympatry of multituberculates and eutherians in Western Europe, at least by the end of the Cretaceous. It adds to the preliminary fossil data reported from Serbia by Marković et al. (Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025) which also challenges this picture for Eastern Europe. The coeval of multituberculates and eutherians in Western Europe by the end of the Cretaceous is likely linked to the late (Maastrichtian) dispersal of the kogaionid Hainina. Previous picture of the geographic segregation of Late Cretaceous mammals in Europe may, in fact, be due to a fossilization bias for some depositional environments, particularly in Eastern Europe. It may reflect distinct predominant palaeoenvironments and palaeoecologies in Western and Eastern Europe, with differences in group adaptation and relative abundance leading to differential preservation. Such a preservation bias is the most likely for the eutherians in Eastern Europe. This is supported by the origin of the Lainodontinae from a mid-Cretaceous Central Asian Sheikhdzheilia-like zhelestid stem group, and this is now evidenced in Serbia by Marković et al. (Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025). Interestingly, Csiki-Sava et al. (Reference Csiki-Sava, Vremir, Meng, Vasile, Brusatte and Norell2022) suggested that Cretaceous Transylvanian kogaionids probably inhabited semi-arid, dry environments (with a possible burrowing habit), based on the sedimentology of the sites and the particular adaptation of their iron-enriched teeth. Such biases of fossil preservation obviously do not alter the strong faunal provincialism signal in the European Archipelago during the Late Cretaceous.
7. Conclusion
Our new palaeontological field research at the Petites Pyrénées latest Cretaceous sites (Tricouté and Cassagnau) yielded a diversified continental vertebrate fauna comprising 16-18 species (Table 2). From the Tricouté sites 1 and 4, we report especially the first occurrence of an alestid characiform, that of a teiioid squamate previously unknown in Western Europe, and two new mammals. The localities of Tricouté are of particular interest because they have yielded both micro-and macrovertebrates, either in the same local sites or in adjacent, closely correlated ones. Among the macrovertebrates, the most frequent are the lambeosaurine hadrosaurs found at the Tricouté 2 site (Laurent, Reference Laurent2003; = Marignac-Laspeyres in Le Loeuff et al. Reference Le Loeuff, Buffetaut, Cavin, Laurent, Martin, Martin and Tong1994), the Tricouté 3 site (Laurent, Reference Laurent2003; Prieto-Márquez and Carrera Farias, Reference Prieto-Marquez and Carrera Farias2012), and the Cassagnau 1–2 sites (Laurent et al. Reference Laurent, Bilotte and Le Loeuff2002; Laurent, Reference Laurent2003). The Maastrichtian Tricouté vertebrate assemblage includes taxa with mixed palaeobiogeographic affinities, among which are endemic European mammals and squamates (teiioids), Laurasian hadrosaurs and thoracosaurs, and Gondwanan taxa such as an alestid characiform (this work) and a possible amphisbaenian squamate (Gheerbrant et al. Reference Gheerbrant, Abrial and Cappetta1997 and this work). The occurrence of endemic mammals reported here in the Tricouté sites is well consistent with the insular palaeogeographic context of the European Archipelago during the Late Cretaceous (Pereda Suberbiola, Reference Pereda-Suberbiola2009; Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015).
The best-documented mammal from Tricouté is a new kogaionid multituberculate, Hainina cassagnauensis n. sp. It is the first known late Cretaceous multituberculate in Western Europe and the earliest and most basal species of the genus Hainina that was previously only known from the Paleocene. Until this discovery and a recent preliminary report in Serbia (Marković et al. Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025), Cretaceous kogaionid multituberculates were only known on the Hațeg Island, in Eastern Europe, consistently with the strong provincialism of the vertebrate faunas within the European Archipelago, which evolved in several isolated local landmasses such as the Ibero-Armorican Island and the Hațeg Island (e.g. Pereda-Suberbiola, Reference Pereda-Suberbiola2009; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015; Csiki-Sava et al. Reference Csiki-Sava, Vremir, Vasile, Brusatte, Dyke, Naish, Norell and Totoianu2016). The endemism of the Hațeg Island fauna is even more striking in the composition of the reptilian assemblage and in the presence of dwarf dinosaurs (Stein et al. Reference Stein, Csiki, Rogers, Weishampel, Redelstorff, Carballido and Sander2010; Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010; Grellet-Tinner et al. Reference Grellet-Tinner, Codrea, Folie, Higa and Smith2012; Csiki-Sava et al. Reference Csiki-Sava, Vremir, Vasile, Brusatte, Dyke, Naish, Norell and Totoianu2016; Mayr et al. Reference Mayr, Codrea, Solomon, Bordeianu and Smith2020; Maidment et al. Reference Maidment, Butler, Brusatte, Meade, Augustin, Csiki-Sava and Ősi2026). The intra-European provincialism results from a vicariant evolution of an ancestral vertebrate fauna common to the entire European Archipelago, which represents the ‘old European core’ (e.g. Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015). Our new discoveries in the Petites Pyrénées, along with other fossil evidences, support that this strong and ancient European provincialism was broken by faunal interchanges at the end of the Cretaceous within the European Archipelago. Together with the barbatteiid teiioid squamate from Tricouté 4, and possibly the rhabdodontid dinosaurs (Weishampel et al. Reference Weishampel, Jianu, Csiki and Norman2003; Ősi et al. Reference Ősi, Prondvai, Butler and Weishampel2012; Augustin et al. Reference Augustin, Bastiaans, Dumbravă and Csiki-Sava2022; but see Maidment et al. Reference Maidment, Butler, Brusatte, Meade, Augustin, Csiki-Sava and Ősi2026), the kogaionid Hainina cassagnauensis n. sp. provides indeed one of the best-known fossil evidence for an East-West dispersal across the European Archipelago. This dispersal event correlates with a faunal change in Western Europe (Ibero-Armorican Island) during the latest Cretaceous, which is known as the ‘Maastrichtian Dinosaur Turnover’ (Le Loeuff et al. Reference Le Loeuff, Buffetaut, Cavin, Laurent, Martin, Martin and Tong1994; Fondevilla et al. Reference Fondevilla, Riera, Vila, Sellés, Dinarès-Turell, Vicens, Gaete, Oms and Galobart2019; Díez Díaz, Reference Díez Díaz2022). It is characterized by the disappearance and decline of several non-avian dinosaur taxa (Le Loeuff et al. Reference Le Loeuff, Buffetaut, Cavin, Laurent, Martin, Martin and Tong1994; Buffetaut Reference Buffetaut2005; Csiki-Sava et al. Reference Csiki-Sava, Vremir, Vasile, Brusatte, Dyke, Naish, Norell and Totoianu2016) and the sudden appearance (in abundance) of hadrosaurs (Laurent Reference Laurent2003; Prieto-Márquez et al. Reference Prieto-Márquez, Dalla Vecchia, Gaete and Galobart2013; Fondevilla et al. Reference Fondevilla, Riera, Vila, Sellés, Dinarès-Turell, Vicens, Gaete, Oms and Galobart2019), which are present in the Tricouté sites. In terms of stratigraphic distribution, the new kogaionid multituberculate from Tricouté provides evidence for one of the best-documented continental vertebrate lineages to cross the K/Pg boundary in Europe: The genus Hainina, particularly with the rapid and local succession, in the Pyrenean area, of the closely related species H. cassagnauensis n. sp. and H. pyrenaica.
Another mammal species discovered in the Tricouté sites is known from an eutherian upper molar. Although still poorly known, this species provides important new data on Cretaceous European eutherians. It displays an original specialized morphology that departs from most known Cretaceous eutherians. In fact, its specialized dental morphology is remarkably reminiscent of some other, still enigmatic Cretaceous European genera such as Azilestes and Valentinella, with which it shares a large size and other euungulate-like features such as the well-developed hypocone previously unknown in Mesozoic eutherians. Our detailed comparison shows that the Tricouté eutherian is more closely related to the genus Azilestes to which it is referred in open nomenclature as a new species, cf. Azilestes yvettae n. sp. In Europe, Mesozoic eutherians are only known from the latest Cretaceous (Campano-Maastrichtian stages) and mostly from the SW part of the European Archipelago, i.e. in the Ibero-Armorican Island (but see Marković et al. Reference Marković, Milivojević, Butler, Barrett, Wills, van de Weerd, Wessels and Radović2025). They have a poor fossil record limited to less than 60 specimens, mostly isolated teeth, from about 10 localities. However, and paradoxically, they display a significant diversity, comprising at least 5 genera and 7 species (Gheerbrant and Teodori, Reference Gheerbrant and Teodori2021). The best identified ones are represented by (1) the lainodontine zhelestids (Lainodon, Labes), (2) the specialized genera Azilestes and Valentinella, and (3) the enigmatic genus Mistralestes. The discovery of cf. Azilestes yvettae n. sp. adds to the significant diversity of the European eutherians from the Cretaceous, especially adaptively with its original ungulate-like specializations, otherwise only known in Azilestes ragei and Valentinella. One important question remains unanswered: how many endemic radiations do these Cretaceous European eutherians represent? The lainodontines clearly belong to an ancient endemic branch of zhelestids (Gheerbrant and Astibia, Reference Gheerbrant and Astibia2012). The relationship of Azilestes and Valentinella are more ambiguous, being obscured by their original specialized characters and their few known fossils. However, their general molar morphotype is most reminiscent of the Zhelestidae to which they are provisionally referred. Consequently, it suggests an ancient endemic radiation of a single main eutherian clade in Europe, the zhelestids, but with several well-distinct suprageneric lineages, including (1) the conservative lainodontines and (2) the specialized genera Azilestes and Valentinella. This is a preliminary tentative hypothesis which remains to be tested. Future fossil discoveries may indeed alternatively provide support that Azilestes and Valentinella belong to an independent European radiation from the Zhelestidae. Mistralestes most likely represents another distinct European eutherian lineage.
Current phylogenetic studies suggest that the two main groups of mammals found in the Tricouté sites and in the late Cretaceous of Europe, i.e. the kogaionid multituberculates and the zhelestid eutherians, are relict endemic clades of early/middle Cretaceous Asian origin (Gheerbrant and Astibia, Reference Gheerbrant and Astibia2012; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015; Smith et al. Reference Smith, Codrea, Devillet and Solomon2022). They belong to the Cretaceous «old European faunal core» and are strong evidence of the ancientness of the endemism in the European Archipelago, together with several other non-mammalian continental vertebrates (Pereda-Suberbiola, Reference Pereda-Suberbiola2009; Weishampel et al. Reference Weishampel, Csiki, Benton, Grigorescu and Codrea2010; Csiki-Sava et al. Reference Csiki-Sava, Buffetaut, Ősi, Pereda-Suberbiola and Brusatte2015; Pereda Suberbiola et al. Reference Pereda Suberbiola, Corral, Astibia Ayerra, Badiola Kortabitarte, Bardet, Berreteaga Escudero, Buffetaut, Delgado Buscalioni, Cappetta, Cavin, Díez Díaz, Gheerbrant, Ortega Coloma, Pérez García, Poyato Ariza, Rage, Sanz García and Torices Hernández2015).
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S0016756826100673
Acknowledgements
We thank J.M. Pacaud (MNHN), P. Richir (MNHN) and C. de Muizon (CNRS, CR2P-MNHN) who helped with the field work at the Tricouté sites. We thank the successive mayors of Marignac-Laspeyres G. Cazaulon and J. L. Lassere and also Mr. Meme who kindly authorized and provided essential support for the excavation of the Tricouté sites in the Cassagnau Hill. We thank F. Goussard (CR2P, MNHN) for his great help with the 3D digital reconstruction and modelling of the Tricouté mammal teeth from microtomographies (Figures 6, 7, 12b,c; Figures S1–S3). We thank M. Bellato and P. Wills for the realization of the CT scans at the AST-RX technical platform (‘ID-Fix’ scanner, UMS 2700, MNHN). R. Vullo (CNRS, Geosciences Rennes) and D. Dutheil (CR2P, MNHN) helped us to identify the characiform tooth TRCT 4-13. We are very grateful to our colleague R. Vacant (CNRS, UMR7207-CR2P, MNHN) who helped us with the delicate and long work of extraction and preparation of the vertebrate remains from the sandstone matrix, especially with the acid attack process. J. Marchand (SAGA-MNHN, deceased) and A. Kriloff (formerly MNHN) provided great help with the difficult and time-consuming job of sorting and picking up the microvertebrate remains (including mammal teeth) from the concentrated residue resulting from the acid attack process. We thank P. Loubry (CR2P, MNHN) for the SEM photographs (Figures 3a–c, 5–6; 8, 12a and 17) and L. Cazes (CR2P, MNHN) for the optic photographs (Figure 3d–h). We thank A. Lethiers (UMR7207-CR2P, Sorbonne University) for redrawing the geological map (Figure 1). We thank R. Tabuce (CNRS, ISEM, University of Montpellier) for providing photographs of the specimen UP-VLP-07-04 of Valentinella vitrollense. We thank E. Buffetaut (CNRS) for his insightful comments on the palaeobiogeography of Cretaceous European faunas. Z. Kielan-Jaworowska (1925–2015) provided comments in April 2001 on some of the Tricouté multituberculate teeth which helped with their initial identification. We also thank T. Smith (IRSN, Brussels) for comments on the multituberculate tooth MHNT.PAL.2024.2.6. We thank the two referees for their useful comments and corrections, which greatly improved the manuscript of this article. Field work was supported by grants from the UMR7207-CR2P.
Author contributions
EG initiated and developed the fieldwork, studied the material and wrote the first draft of the paper. YL and RA contributed to writing the introduction (incl. geological context), to identifying, studying and commenting on the non-mammalian fauna (especially the herpetofauna) and to writing the general discussion. All authors contributed to writing, revising and editing the paper.
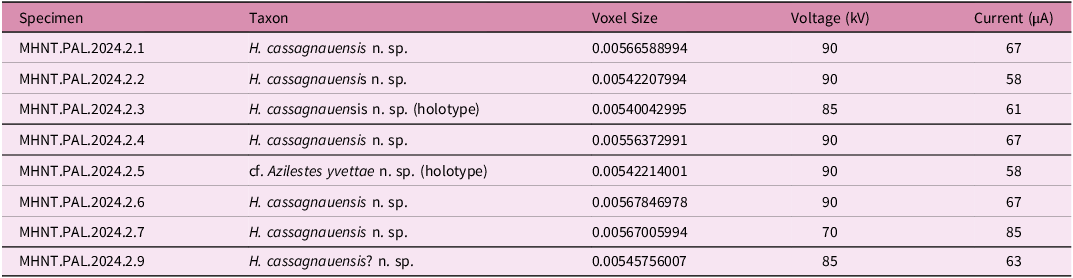






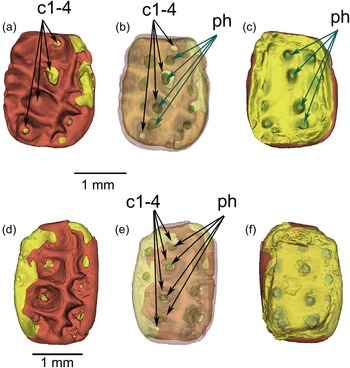



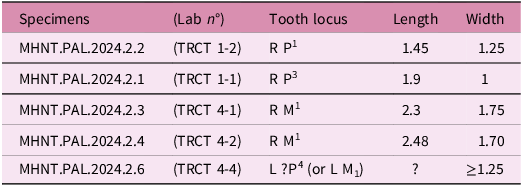


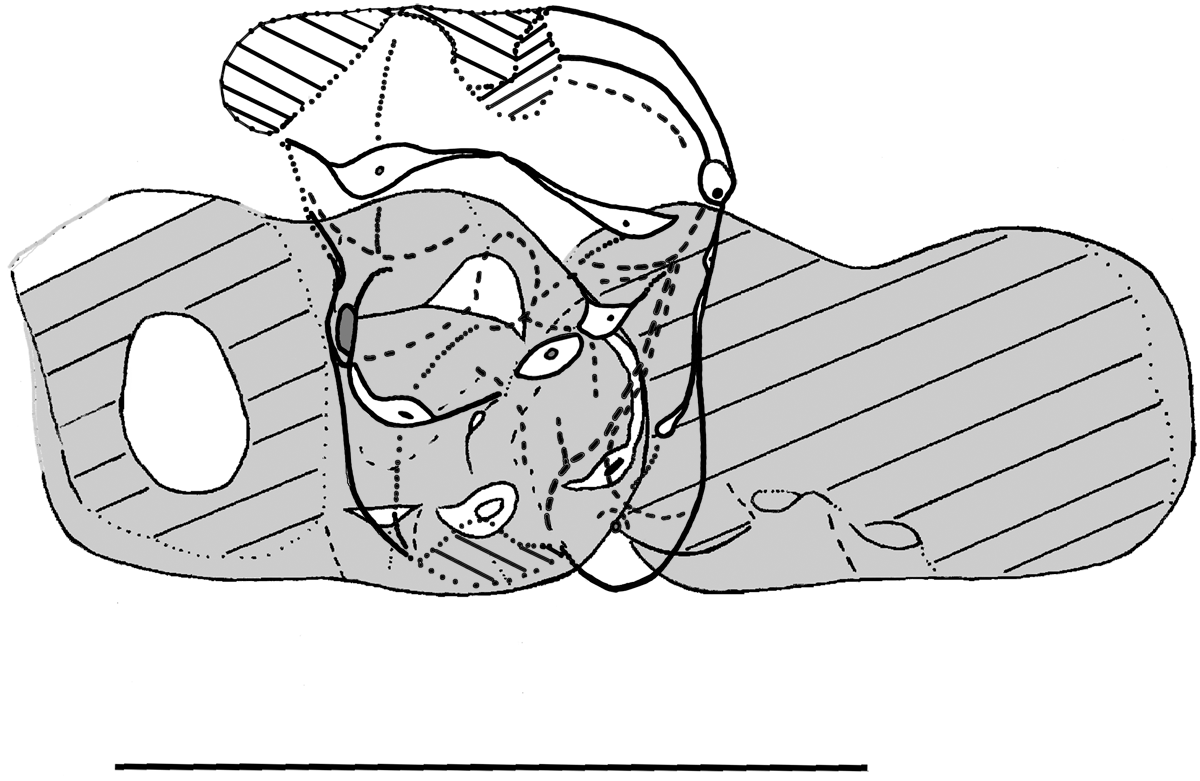



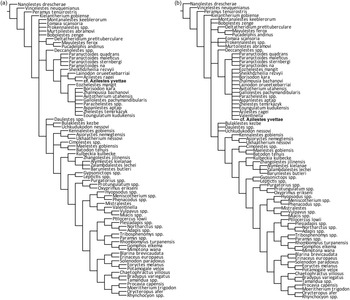




 Open access
Open access