


Introduction
Neurotrophin and NGF
Neurotrophins (NTs) are a family of small polypeptide growth factors that were initially discovered to play a role in the central and peripheral nervous systems (Ref. Reference Madhubala, Mahato, Khan, Bala and Mukherjee1). Subsequent studies have gradually revealed the widespread distribution of NT and its receptors in other tissues, including the digestive tract, lymph nodes, spleen, breast and reproductive system (Refs Reference Solé-Morata, Baenas, Etxandi, Granero, Forcales, Gené, Barrot, Gómez-Peña, Menchón, Ramoz, Gorwood, Fernández-Aranda and Jiménez-Murcia2, Reference Wang, Fan, Cao, Huang, Yao and Li3, Reference Ding, Li, Jaklenec and Hu4). Nerve growth factor (NGF), discovered by Levi Montalcini in 1952, was the earliest identified member of the NT family (Ref. Reference Levi-Montalcini5). To date, it is the most thoroughly studied NT and its main functions include regulating the growth, development, differentiation, survival and repairment of neurons after injury. In addition, NGF is also involved in multiple processes such as pain, inflammation, tumour proliferation and metastasis, as well as reproductive physiology. Clinical studies have found that NGF concentration has increased in diseases with chronic pain symptoms such as osteoarthritis, chronic headache, cancer-related pain and diabetes neuropathy. NGF gene mutations (R121W, V232fs and R221W) will lead to painless diseases, suggesting that NGF-mediated signals are continuously active in chronic nociceptive and neuropathic pain (Refs Reference Testa, Cattaneo and Capsoni6, Reference Wang, Ni, Xie, Li, Yuan, Meng, Zhao, Gao, Huang, Wang, Ma, Zhou, Feng, Chang, Yang, Yu, Yu, Zang, Zhang, Ji, Wang and Hao7). At present, monoclonal antibodies that block the NGF-tropomyosin receptor kinase A (TrkA) signalling pathway, such as Fasinumab (REGN475) and Tanezumab (PF-04383119), have advanced into clinical trials for the treatment of osteoarthritis-related pain (Ref. Reference Testa, Cattaneo and Capsoni6). Recently, Zaninelli et al. suggested that anti-NGF treatment reduced endometriosis-associated pain in mice (Ref. Reference Zaninelli, Fattori, Heintz, Wright, Bennallack, Sim, Bukhari, Terry, Vitonis, Missmer, Andrello, Anchan, Godin, Bree, Verri and Rogers8).
Structure of NGF
Human NGF gene is located on chromosome 1, and it first synthesizes two types of long and short NGF precursor after transcription and translation (Ref. Reference Chao, Bothwell, Ross, Koprowski, Lanahan, Buck and Sehgal9). After processing and hydrolysis, the long NGF precursor is transformed into 7.5S NGF, which is a complex composed of two α subunits (α-NGF), one β subunits (β-NGF) and two γ subunits (γ-NGF) with a relative molecular weight of around 140000 Da. In contrast, the short NGF precursor is converted into 2.5S NGF, which mainly contains β-NGF and can be dissociated from long NGF precursor. α-NGF does not have activity, β-NGF has biological activity, and γ-NGF is a unique serine proteinases, which may be related to the conversion of NGF precursor to mature NGF (Ref. Reference Yan, Yalinca, Paoletti, Gobbo, Marchetti, Kuzmanic, Lamba, Gervasio, Konarev, Cattaneo and Pastore10). β-NGF is a homodimer formed by noncovalent binding and parallel assembly of two identical peptide chains, with disulfide bonds present among six cysteine residues, which are crucial for maintaining the biological activity of NGF. Each peptide chain contains 118 amino acid residues, with a relative molecular weight of approximately 26518 Da and an isoelectric point of 9.3 (Ref. Reference Ratto, Leduc, Valderrama, van Straaten, Delbaere, Pierson and Adams11).
NGF receptors
The biological actions of NGF are mediated by binding to two different types of membrane surface receptors: TrkA, which belongs to the tyrosine kinase receptor superfamily and is a specific high-affinity receptor for NGF, and p75 neurotrophic receptor (p75NTR), which belongs to the tumour necrosis factor superfamily and is a non-specific low-affinity receptor for NGF (Refs Reference Reichardt12, Reference Chao and Hempstead13, Reference Skaper14, Reference Barbacid15).
Expression characteristics of NGF and its receptors in human ovaries
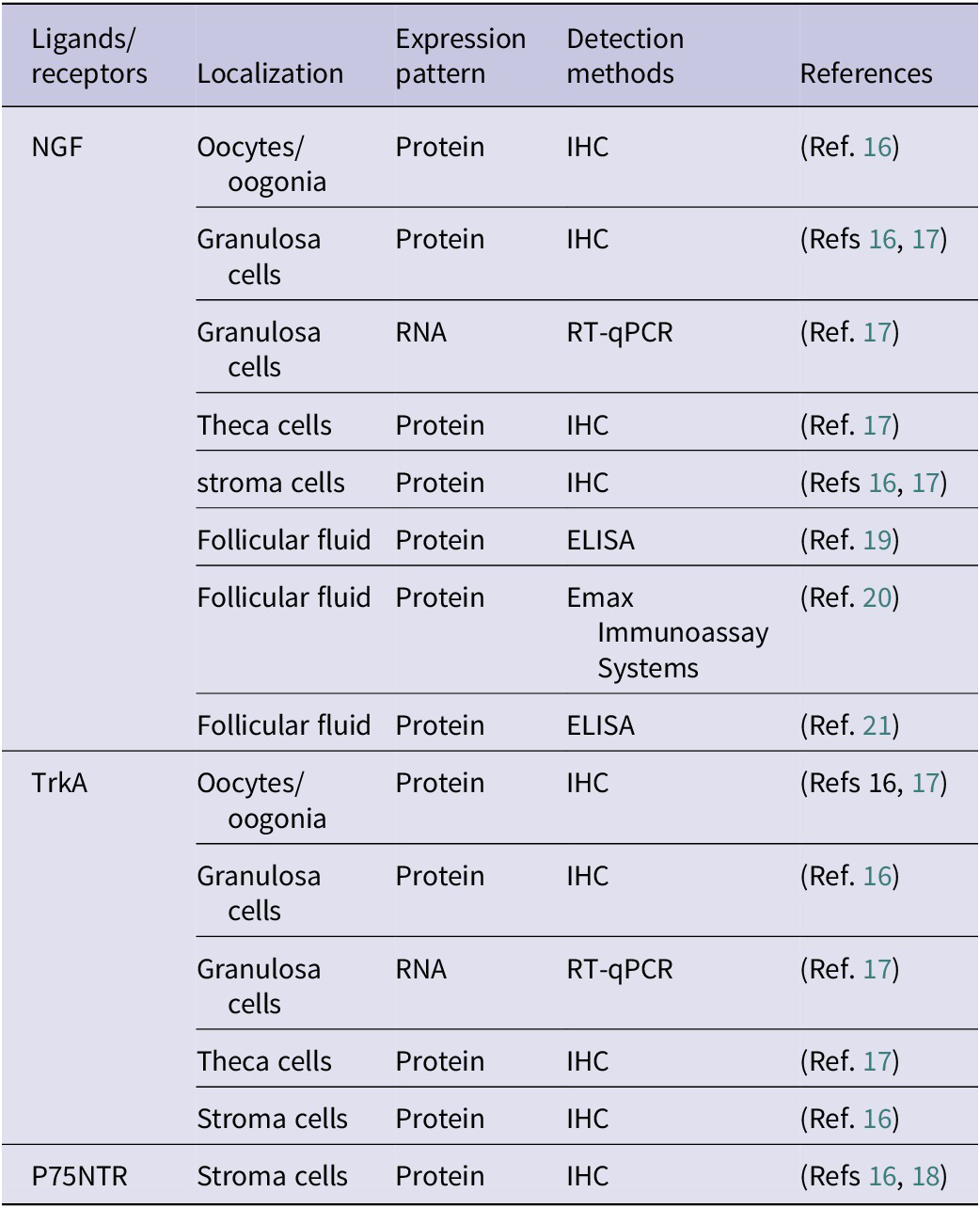
Multiple studies have shown that NGF and its receptors are expressed in oocytes, granulosa cells and other cells within the human ovary from embryo to adulthood (Table 1), and play an important role in reproductive physiology processes, such as follicle development and ovulation. As early as the embryonic period at 19–33 weeks of gestation, NGF are expressed in oogenia/oocytes of human embryo ovaries (Ref. Reference Abir, Fisch, Jin, Barnnet, Ben-Haroush, Felz, Kessler-Icekson, Feldberg, Nitke and Ao16). In ovaries of adolescents/adults, NGF is expressed in granulosa cells of preantral follicles and antral follicles, as well as in all oocytes (Reference Abir, Fisch, Jin, Barnnet, Ben-Haroush, Felz, Kessler-Icekson, Feldberg, Nitke and Ao16, Reference Salas, Julio-Pieper, Valladares, Pommer, Vega, Mastronardi, Kerr, Ojeda, Lara and Romero17). In the embryonic period, TrkA is mainly expressed in oogenia/oocytes in most cases, with a few expressed in granulosa cells (Ref. Reference Abir, Fisch, Jin, Barnnet, Ben-Haroush, Felz, Kessler-Icekson, Feldberg, Nitke and Ao16). In ovaries of adolescents/adults aged 13–39, TrkA is expressed in granulosa cells of preantral follicles and antral follicles, theca cells of antral follicles and most oocytes (Refs Reference Abir, Fisch, Jin, Barnnet, Ben-Haroush, Felz, Kessler-Icekson, Feldberg, Nitke and Ao16, Reference Salas, Julio-Pieper, Valladares, Pommer, Vega, Mastronardi, Kerr, Ojeda, Lara and Romero17). P75NTR is expressed in some stroma cells of human foetal ovaries (Refs Reference Abir, Fisch, Jin, Barnnet, Ben-Haroush, Felz, Kessler-Icekson, Feldberg, Nitke and Ao16, Reference Anderson, Robinson, Brooks and Spears18). Additionally, NGF also exists in human follicular fluid (Refs Reference Sadeu, Doedée, Neal, Hughes and Foster19, Reference Buyuk and Seifer20, Reference Zhai, Yao, Rao, Wang, Song and Sun21).
Distribution of NGF and its receptors in the human ovaries

Table 1. Long description
From the top row downward, the table is divided by ligand or receptor. NGF is localized in oocytes or oogonia, granulosa cells, theca cells, stroma cells, and follicular fluid. Expression is detected as protein in all except granulosa cells, where RNA is also detected. Detection methods include I H C for all cell types, R T-q P C R for granulosa cell RNA, and E L I S A or Emax Immunoassay Systems for follicular fluid. References are 16, 17, 19, 20, and 21. TrkA is found in oocytes or oogonia, granulosa cells, theca cells, and stroma cells, with protein detected by I H C and RNA by R T-q P C R in granulosa cells. References are 16 and 17. P75 N T R is detected as protein in stroma cells by I H C, with references 16 and 18. Table footnotes define I H C as immunohistochemistry, N G F as nerve growth factor, P75 N T R as p75 neurotrophic receptor, R T-q P C R as quantitative reverse transcription polymerase chain reaction, and TrkA as tropomyosin receptor kinase A.
IHC: immunohistochemistry; NGF: nerve growth factor; p75NTR: p75 neurotrophic receptor; RT-qPCR: quantitative reverse transcription polymerase chain reaction; TrkA: tropomyosin receptor kinase A.
Both granulosa cells and theca cells of human ovaries can synthesize NGF, with theca cells serving as the primary site of secretion (Ref. Reference Salas, Julio-Pieper, Valladares, Pommer, Vega, Mastronardi, Kerr, Ojeda, Lara and Romero17). Experiments have shown that NGF concentration in follicular fluid is related to the follicle size. In sheep, large antral follicles (>4 mm in diameter) produce NGF, whereas small antral follicles and early atresia follicles produce minimal to undetectable levels (Ref. Reference Mattioli, Barboni, Gioia and Lucidi22). The NGF concentration in follicular fluid is regulated by gonadotrophin-releasing hormone (GnRH) and gonadotropins. GnRH causes a dramatic increase of NGF in large follicles, while gonadotrophins stimulate cultured granulosa cells and follicles to produce NGF. In the presence of both luteinizing hormone (LH) and follicle-stimulating hormone (FSH), the production of NGF showed a clear dose-dependent response in vitro based on the culture of sheep antral follicles, while no detectable levels of neurotrophic activity accumulated in medium when gonadotrophins were absent or added separately (Refs Reference Mattioli, Barboni, Gioia and Lucidi22, Reference Barboni, Mattioli, Gioia, Turriani, Capacchietti, Berardinelli and Bernabò23). In addition, some researchers reported that NGF and its receptors changed with seasons in squirrels. Stronger immunostaining of NGF, TrkA and p75 were observed in granulosa cells, thecal cells, interstitial cells and ovarian surface epithelium in the breeding season compared to the nonbreeding season (Refs Reference Li, Sheng, Bao, Huang, Li, Liu, Han, Watanabe, Taya and Weng24, Reference Bao, Li, Liu, Li, Sheng, Han and Weng25).
Oocyte maturation and folliculogenesis
Follicular assembly and follicle development
The effects of NGF on follicle development are mostly obtained from mouse models, with a few taken from other animals such as rats and sheep (Figure 1). Follicular assembly is a process characterized by primary oocytes enveloped by pre-granulosa cells and primordial follicles formation (Ref. Reference Wang, Ge, Zhai, Liu, Sun, Liu, Li, Lei, Dyce, de Felici and Shen26). The main period of mouse follicular assembly occurs from postnatal day 0 to days 3–5. At birth, the ovary contains a large number of oocytes, which are connected by intercellular bridges, forming oocyte nests. Within 5 days after birth, the oocyte nests break down, the intercellular bridges disintegrate, and pre-granulosa cells surround individual oocytes, thereby forming primordial follicle. Around day 5 after birth, the primordial follicle pool in mice is essentially established, and some primordial follicles begin to be activated and develop into primary follicles (Refs Reference Lei and Spradling27, Reference Tingen, Kim and Woodruff28). Gene editing animal models have found that NGF exerted an effect in the reproductive physiology of primordial follicular assembly and early follicle development. Dissen et al. found that the number of unassembled and naked oocytes in 7-day-old ovaries from NGF −/− mice significantly increased, while the number of primary follicles and secondary follicles decreased significantly, with normal serum FSH and LH levels, indicating that NGF participates in follicular assembly in a gonadotropin-independent manner and increases the number of primary follicles and secondary follicles (Ref. Reference Dissen, Romero, Hirshfield and Ojeda29). Kerr conditionally in vitro cultured ovaries from 1-day-old NGF −/− mice for 4 days in the presence of NGF. The number of primordial follicles increased and follicular assembly was rescued after 4 days of culture, proving that the reduction of follicles is reversible (Ref. Reference Kerr, Garcia-Rudaz, Dorfman, Paredes and Ojeda30). Furthermore, in the phase of late follicle development researchers has found that NGF maintained the number of preantral follicles in ovarian tissue cultured in vitro by downregulating the expression of pro apoptotic genes Bcl-2-associated X protein (Bax) and tumour protein 53 (p53), and the ovarian injury model also confirmed the relationship between NGF and selective follicle activation (Refs Reference Roh and Pi31, Reference He, Peng, Wu, Yang, Liu, Zhang, Su, Kong, Dou and Li32). On the other hand, Zhang et al. reported that reduced acetylation of NGF accelerates its ubiquitin-mediated degradation, and that low levels of NGF protein help maintain dormancy of primordial follicles (Ref. Reference Zhang, Tong, Zhu, Liang, Zhang, Hu, He, Hu, Shen, Niu, Zhang, Yu, Jin, Lei, Zeng, Wu, Cheng, Xiao, Guo, Zhao, Xu, Pan and Chen33). This supports a positive role for NGF in follicular development.
Involvement of NGF in multiple reproductive processes, such as follicular assembly and follicle development, oocyte maturation, ovulation and ovarian steroidogenesis. Note: NGF: nerve growth factor; GVBD: germinal vesicle breakdown.

Figure 1. Long description
At the center, N G F is linked by a double-headed arrow to ovary. Top left quadrant, labeled ovarian steroidogenesis, shows N G F from granulosa or theca cells in an N G F-overexpressed mouse, increasing estradiol, progesterone, and testosterone. Top right quadrant, labeled follicular assembly and follicle development, shows N G F knockout mouse leading to increased naked oocytes, and a sequence from primordial follicle to primary and secondary follicle with plus signs. Bottom right quadrant, labeled oocyte maturation, shows physiological concentrations of N G F promoting first polar body extrusion, nuclear maturation, and G V B D, with a plus sign from oocyte to mature oocyte, and pathological elevated concentrations leading to polycystic ovary. Bottom left quadrant, labeled ovulation, shows N G F inducing ovulation in an induced ovulator (alpaca) and more synchronous ovulation in a spontaneous ovulator (cow). All quadrants are color-coded and connected to the central N G F–ovary axis.
However, another research cultured ovarian cortex fragments of goats and pointed that NGF improved the survival of preantral follicles and maintained the morphology of normal follicles at 1 ng/ml concentration, while at high concentration of NGF (100 and 200 ng/ml), there was a significant reduction in the percentage of normal follicles and the disrupted integrity of follicular structure, manifesting cytoplasmic vacuolization, disruption of basal membrane integrity and disorganization of granulosa cells (Ref. Reference Chaves, Alves, Duarte, Araújo, Celestino, Matos, Lopes, Campello, Name, Báo and Figueiredo34).
Additionally, Buyuk revealed a positive relation between the number of oocytes retrieved and TrkA mRNA level in human mural and cumulus granulosa cells, providing a novel method for ovarian reserve prediction (Ref. Reference Buyuk, Santoro, Cohen, Charron and Jindal35).
To determine which receptor mediates the effects of NGF on follicular assembly, researchers constructed TrkA−/− mouse model and found that the number of primordial follicles in 2-day-old ovaries was reduced significantly. When ovaries from 1-day-old TrkA−/− mice were cultured for 4 days in the presence of NGF, no increase in primordial follicle numbers were detected in comparison with untreated TrkA−/− ovaries, indicating that the facilitatory effect of NGF on follicular assembly is mediated by receptor TrkA (Ref. Reference Kerr, Garcia-Rudaz, Dorfman, Paredes and Ojeda30). Furthermore, the number of secondary follicles rather than primary follicles decreased in TrkA−/− mice, indicating a defect in primary follicle recruitment to the proliferative pool and involvement of NGF mediated by TrkA in the early follicle development process (Ref. Reference Kerr, Garcia-Rudaz, Dorfman, Paredes and Ojeda30). Ovaries from p75NTR−/− mice formed follicles in a normal manner, and the number of primary follicles and secondary follicles increased, indicating that p75NTR may act as an inhibitory regulatory signal during follicle development (Ref. Reference Ojeda, Romero, Tapia and Dissen36).
The currently known molecules that may be involved in the regulation of follicular assembly and development by NGF include cadherin 11 and pituitary tumour transforming gene (PTTG). Cadherin 11 is a cell adhesion molecule, and in situ hybridization shows that oocytes express a large amount of calcium binding protein mRNA before follicle formation. During the follicular formation stage within 24 h after birth, oocytes are surrounded by pre-granulosa cells, and the expression of cadherin 11 mRNA decreases, indicating that the loss of adhesion between oocytes helps to isolate individual oocytes into primordial follicles. Another factor that may be involved in follicle formation is PTTG, a proto-oncogene that increases mRNA expression 24 h after birth and is expressed at a fixed level after follicle formation (Ref. Reference Ojeda, Romero, Tapia and Dissen36). In addition, reduction of proliferation markers, proliferating cell nuclear antigen and 5-bromo-2′-deoxyuridine, was observed in NGF−/− mice, suggesting that NGF may stimulate proliferation signals (Ref. Reference Dissen, Romero, Hirshfield and Ojeda29).
Oocyte maturation
Oocyte maturation is a complex process involving a series of events (Ref. Reference Coticchio, Dal Canto, Mignini Renzini, Guglielmo, Brambillasca, Turchi, Novara and Fadini37). In this review, we focused specifically on the role of NGF in regulating this process (Figure 1).
The effect of NGF on oocyte maturation mainly came from mammals such as mice and sheep, and there is relatively limited research in humans. NGF may act as a paracrine signal and indirectly promote or inhibit oocyte maturation. Immunohistochemistry and western blot investigation gave fulfil proofs that TrkA is located on cumulus cells rather than oocytes (Refs 21, Reference Barboni, Mattioli, Gioia, Turriani, Capacchietti, Berardinelli and Bernabò23). At physiological concentrations, NGF promotes oocyte maturation. Through mRNA/miRNA sequencing and functional analysis, NGF was identified as a favourable gene for oocyte developmental potential in mouse cumulus–oocyte complexes (Ref. Reference Zhang, Wang, Zhang, He, Luo, Gong, Fu, Yuan, Zhang and Tan38). Barboni et al. demonstrated that NGF could alter the meiotic arrest of sheep oocytes in vitro, and dose dependently reduce the number of oocytes in the germinal vesicle phase at concentrations of 1–100 ng/ml (Ref. Reference Barboni, Mattioli, Gioia, Turriani, Capacchietti, Berardinelli and Bernabò23). Similarly, Wang et al. found that the optimal concentration of NGF for promoting sheep oocyte maturation, which is manifested by nuclear maturation and the rate of first polar body extrusion, is 3 ng/ml at a range of 0–300 ng/ml in vitro (Ref. Reference Wang, Ren, Zhou, Han, Wang and Liang39). The role of NGF may vary at different stages of oocyte and embryo development. For example, supplement of NGF during oocyte maturation or fertilization could induce early cleavage, while adding NGF during embryo culture had no significant effect (Ref. Reference Crispo, dos Santos-Neto, Vilariño, Mulet, de León, Barbeito and Menchaca40). Furthermore, NGF may act as a mediator during follicle development, manifesting elevated NGF mRNA expression about 5 h before ovulation after injection of gonadotrophin in rats (Refs Reference Dissen, Hill, Costa, Les Dees, Lara and Ojeda41, Reference Mayerhofer, Dissen, Parrott, Hill, Mayerhofer, Garfield, Costa, Skinner and Ojeda42).
Conversely, pathological elevations of NGF concentration can inhibit oocyte maturation. This theory is mainly derived from the discovery of abnormally elevated NGF concentration in follicular fluid of patients with polycystic ovary syndrome (PCOS), which is characterized by impaired oocyte maturation (Refs Reference Sadeu, Doedée, Neal, Hughes and Foster19, Reference Dissen, Garcia-Rudaz, Paredes, Mayer, Mayerhofer and Ojeda43, Reference Gulino, Giuffrida, Leonardi, Marilli and Palumbo44). Zhai et al. found that NGF concentration in follicular fluid of PCOS patients (22 ± 2.63 ng/ml) was significantly higher than that of non-PCOS patients (7.18 ± 2.42 ng/ml). Further research found that decrease of oocyte maturation competence caused by elevated NGF was associated with glycolysis abnormalities (Ref. Reference Zhai, Yao, Rao, Wang, Song and Sun21).
Ovulation
Based on the upstream stimuli that induce ovulation, mammals can be classified into two distinct groups: induced ovulators and spontaneous ovulators. Induced ovulators, such as rabbits, cats and camels, are characterized by triggered ovulation. Initially, researchers attributed this response to neural signals from mechanical stimulation of the female reproductive tract. However, more recent studies have revealed that seminal fluid contains bioactive factors that act as the primary stimulus for the pre-ovulatory LH surge, thereby challenging the traditional view (Ref. Reference Ratto, Berland, Silva and Adams45). By contrast, spontaneous ovulators, such as cows, horses and pigs, exhibit oestrogen-dependent ovulation. In these species, rising estradiol levels increase hypothalamic GnRH pulse frequency and amplitude, ultimately surpassing a threshold to elicit the LH surge (Ref. Reference Jo, Brännström, Akins and Curry46).
β-NGF and ovulation
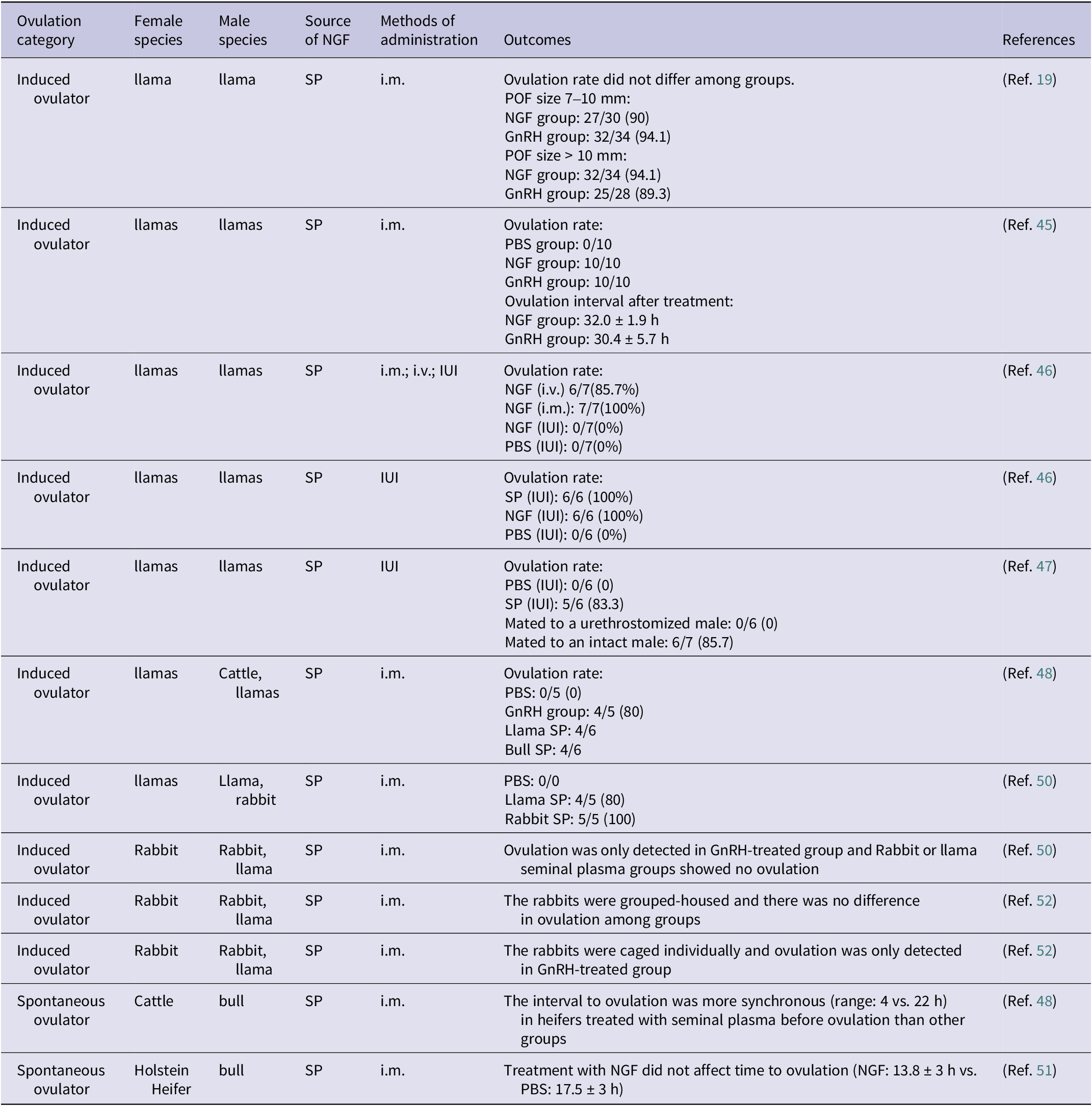
Previous studies have referred to the substance present in the seminal plasma that can induce ovulation as ovulation-inducing factor (OIF) (Ref. Reference Adams, Ratto, Huanca and Singh47). In 2012, Ratto et al. successfully identified and characterized the molecular structure of OIF in llamas and bulls, proving it is β-NGF (Ref. Reference Ratto, Leduc, Valderrama, van Straaten, Delbaere, Pierson and Adams11). β-NGF in the seminal plasma (spβ-NGF) can affect ovulation and corpus luteum development in females (Table 2). To date, however, no equivalent studies have been carried out in humans.
The ovulation induction effect of β-NGF in induced and spontaneous ovulation animals

Table 2. Long description
The table has seven columns: Ovulation category, Female species, Male species, Source of NGF, Methods of administration, Outcomes, and References. From the top row, the first section lists induced ovulators, mainly llamas and rabbits, with seminal plasma as the NGF source. Administration methods include intramuscular, intravenous, and intrauterine. Outcomes detail ovulation rates and intervals for NGF, GnRH, and PBS groups. For example, in llamas with preovulatory follicle size 7 to 10 mm, NGF group had 27 out of 30 ovulations (90 percent), GnRH group 32 out of 34 (94.1 percent); with follicle size greater than 10 mm, NGF group had 32 out of 34 (94.1 percent), GnRH group 25 out of 28 (89.3 percent). In another row, ovulation rate was 0 out of 10 for PBS, 10 out of 10 for NGF, and 10 out of 10 for GnRH; ovulation interval was 32.0 plus or minus 1.9 hours for NGF and 30.4 plus or minus 5.7 hours for GnRH. Other rows show that intrauterine NGF or PBS did not induce ovulation, while intramuscular NGF did. In rabbits, ovulation was only detected in GnRH-treated groups, not in seminal plasma groups. The spontaneous ovulator section includes cattle and Holstein heifers, showing that seminal plasma treatment led to more synchronous ovulation intervals (range 4 versus 22 hours), but NGF did not significantly affect time to ovulation compared to PBS (13.8 plus or minus 3 hours versus 17.5 plus or minus 3 hours). References are listed for each row. Abbreviations are defined below the table: GnRH is gonadotrophin-releasing hormone, i.m. is intramuscular, i.v. is intravenous, IUI is intrauterine, NGF is nerve growth factor, PBS is phosphate buffered saline, POF is preovulatory follicle, SP is seminal plasma.
GnRH: gonadotrophin-releasing hormone; i.m.: intramuscular administration; i.v.: intravenous administration; IUI: intrauterine administration; NGF: nerve growth factor; PBS: phosphate buffered saline; POF: preovulatory follicle; SP: seminal plasma.
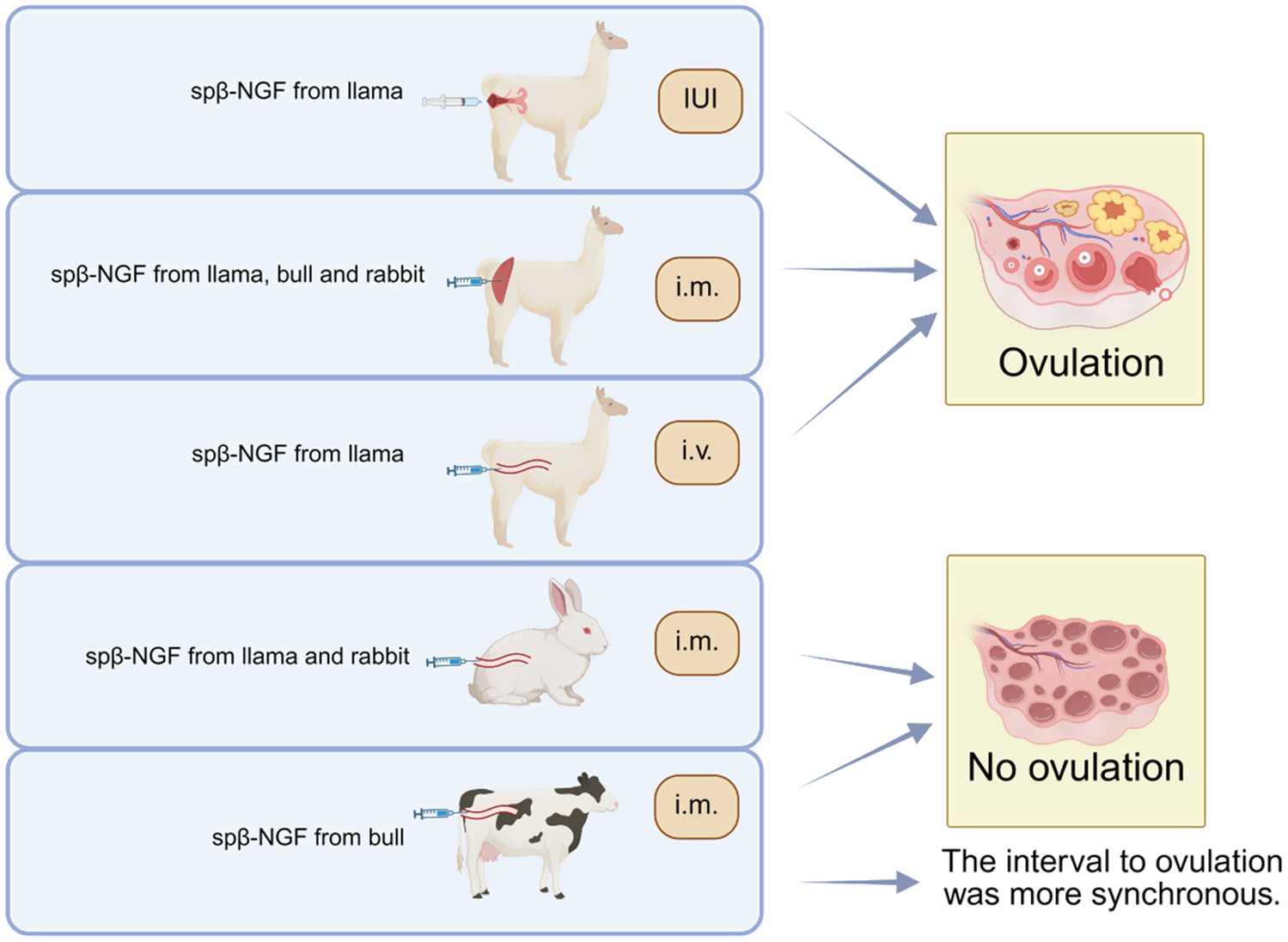
Silva et al. found that intramuscular injection of llama spβ-NGF-induced ovulation in female llama regardless of the follicle size prior to ovulation (Refs Reference Silva, Ulloa-Leal, Norambuena, Fernández, Adams and Ratto48, Reference Ulloa-Leal, Bogle, Adams and Ratto49). Subsequently, further experiments were conducted and verified that both intravenous injection and intramuscular injection of spβ-NGF-induced ovulation. However, intrauterine infusion of spβ-NGF at a concentration of 2 mg/2 ml failed to induce ovulation, whereas high volume and concentration of 20 mg/4 ml of spβ-NGF induced 100% ovulation rate (Ref. Reference Silva, Fernández, Ulloa-Leal, Adams, Berland and Ratto50). Notably, simple physical stimulation during sexual intercourse did not induce ovulation in llamas, while intrauterine infusion and deposition of seminal plasma in reproductive tract were sufficient to cause ovulation, with circulating β-NGF increasing and reaching peak about 15 minutes after treatment, positively correlated with LH elevation (Ref. Reference Ratto51). Interestingly, Tribulo et al. found that intramuscular injection of spβ-NGF from both cattle- and llamas-induced ovulation in female llamas, reflecting the ovulation-induced effect of spβ-NGF among different species (Ref. Reference Tribulo, Bogle, Mapletoft and Adams52). A similar phenomenon about induced ovulation was also observed in rabbits (Ref. Reference Maranesi, Petrucci, Leonardi, Piro, Rebollar, Millán, Cocci, Vullo, Parillo, Moura, Mariscal, Boiti and Zerani53). In rabbits, intramuscular treatment with homologous seminal plasma or murine β-NGF resulted in a significant increase in the number of large haemorrhagic anovulatory follicles, suggesting a direct local effect of these molecules with LH-like action on LH receptors (Ref. Reference Silva, Niño, Guerra, Letelier, Valderrama, Adams and Ratto54). Moreover, pre-treatment with lumbar anaesthesia or cyclooxygenase inhibitors prior to artificial insemination suppressed the plasma LH surge, attenuated the systemic rise in β-NGF which was not synchronous with LH increase, and decreased ovulation rates, confirming the nerve involvement of spβ-NGF in induced ovulation (Ref. Reference Maranesi, Petrucci, Leonardi, Piro, Rebollar, Millán, Cocci, Vullo, Parillo, Moura, Mariscal, Boiti and Zerani53).
In spontaneously ovulating animals such as cows, intramuscular injection of spβ-NGF from bull does not affect the timing of ovulation (Ref. Reference Stewart, Stella, Cunha, Dias, Canisso, Mercadante, Cardoso, Williams, Pohler and Lima55). However, it has been shown to increase the diameter of preovulatory follicles, and treatment with bovine seminal plasma prior to ovulation can enhance the synchrony of ovulation interval (Ref. Reference Tribulo, Bogle, Mapletoft and Adams52) (Figure 2).
Role of NGF in both induced ovulators and spontaneous ovulators. Note: spβ-NGF: β-NGF in the seminal plasma; IUI: intrauterine administration; i.m.: intramuscular administration; i.v.: intravenous administration.

Figure 2. Long description
From top to bottom, the first row shows a llama receiving intrauterine administration of sp beta N G F from llama, labeled I U I, with an arrow pointing to ovulation. The second row shows a llama receiving intramuscular administration of sp beta N G F from llama, bull, and rabbit, labeled i dot m dot, with an arrow pointing to ovulation. The third row shows a llama receiving intravenous administration of sp beta N G F from llama, labeled i dot v dot, with an arrow pointing to ovulation. The fourth row shows a rabbit receiving intramuscular administration of sp beta N G F from llama and rabbit, labeled i dot m dot, with an arrow pointing to no ovulation. The fifth row shows a cow receiving intramuscular administration of sp beta N G F from bull, labeled i dot m dot, with an arrow pointing to no ovulation and a note that the interval to ovulation was more synchronous.
The mechanism of β-NGF-induced ovulation
In induced-ovulation species, multiple studies have demonstrated that NGF triggered ovulation concurrently with a synchronous surge in circulating LH by modulating the hypothalamic–pituitary axis. The primary evidence supporting this mechanism includes three key observations: (1) NGF is absorbed into the systemic circulation through the endometrial tissue, (2) both intravenous and intramuscular administration of NGF reliably induce ovulation accompanied by increased LH levels and (3) the temporal pattern of circulating NGF elevation closely parallels the LH surge (Refs Reference Stewart, Stella, Cunha, Dias, Canisso, Mercadante, Cardoso, Williams, Pohler and Lima55, Reference Adams, Ratto, Silva and Carrasco56, Reference El Allali, El Bousmaki, Ainani and Simonneaux57, Reference Silva, Recabarren, Recabarren, Adams and Ratto58).
Further supporting a central mechanism, pre-treatment with a GnRH antagonist was shown to block OIF-mediated LH release in llamas (Ref. Reference Silva, Smulders, Guerra, Valderrama, Letelier, Adams and Ratto59). Intracerebroventricular injection of β-NGF elevated plasma LH levels in male hypogonadotropic hypogonadal mice, and promoting GnRH release from TrkA-expressing GT1–7 immortalized hypothalamic cells in vitro. These findings indicate that β-NGF’s neuromodulatory action on GnRH neurons represents a conserved mechanism across species and sexes (Ref. Reference Luo, Yang, Zhang, Su, Yu, Lin, Chen, Zhang, Xiang, Xue, Ge and Huang60). The findings also demonstrate that NGF induces a preovulatory LH surge through GnRH neuronal activity.
Hypothalamic GnRH neurons synthesize and transport the hormone to axonal terminals, which project to the median eminence (ME) (Refs Reference Campbell, Han and Herbison61, Reference Herde, Iremonger, Constantin and Herbison62). Here, their terminals associate with the fenestrated capillaries of the hypophyseal portal system, facilitating GnRH delivery to the anterior pituitary (Refs Reference Silverman, Jhamandas and Renaud63, Reference Goldsmith, Thind, Song, Kim and Boggant64, Reference Moore, Prescott, Marshall, Yip and Campbell65). Recent neuroanatomical studies in llamas reveal distinct spatial organization of GnRH neurons. Quantitative analysis shows that the medio-basal hypothalamus contains the highest density of GnRH neurons (Ref. Reference Carrasco, Singh and Adams66). Localization of TrkA receptor adjacent to the third ventricle’s hypothalamic periventricular zone suggests potential β-NGF transport from circulation into cerebrospinal fluid via barrier penetration (Ref. Reference Carrasco, Singh and Adams67). This neuroanatomical arrangement implies the medio-basal hypothalamic regions representing principal target sites for β-NGF activity (Ref. Reference Carrasco, Singh and Adams66). Although TrkA and p75NTR are widely expressed in the llama hypothalamus, subsequent research demonstrated minimal TrkA receptor expression (2.5%) in GnRH neurons and absent p75NTR immunoreactivity, suggesting that β-NGF likely influences GnRH activity through indirect neural pathways rather than direct receptor-mediated stimulation (Ref. Reference Carrasco, Singh and Adams67).
There are several potential pathways through which seminal plasma-derived β-NGF might act at the hypothalamic level. First of all, the kisspeptin neuronal system represents a key regulatory component of GnRH and LH release, with most GnRH neurons expressing kisspeptin receptors (Refs Reference Clarkson, Han, Liu, Lee and Herbison68, Reference Ojeda, Lomniczi and Sandau69, Reference Rodríguez, Blázquez and Guerra70). A greater relative abundance of kisspeptin fibres was detected in association with GnRH fibres in the ME region (Refs Reference Ratto, Berland, Silva and Adams45, Reference Ainani, Chhaibi, Achaâban, Piro, Ouassat, Tibary, el Allali and Simonneaux71). Second, circumventricular organs, where specialized ependymal cells unction as selective gateways and enable bidirectional communication between the bloodstream and neural tissues, may therefore act as a gateway for β-NGF entering and acting on the llama GnRH systems (Refs Reference Rodríguez, Blázquez and Guerra70, Reference Hawkins and Davis72, Reference Prevot, Hanchate, Bellefontaine, Sharif, Parkash, Estrella, Allet, de Seranno, Campagne, d’Anglemont de Tassigny and Baroncini73). Consistent with this, Ratto et al. identified β-NGF immunoreactivity in the tanycytes of the ME in llamas, where GnRH nerve terminals are concentrated (Ref. Reference Ratto, Berland, Silva and Adams45). Experimental evidence in rats demonstrates that tanycyte ablation disrupts GnRH secretion into portal vessels, consequently blocking the LH surge and ovulation, indicating NGF may affect GnRH releasing through tanycytes (Refs Reference Rodríguez, Blázquez and Guerra70, Reference Prevot, Hanchate, Bellefontaine, Sharif, Parkash, Estrella, Allet, de Seranno, Campagne, d’Anglemont de Tassigny and Baroncini73, Reference Rodríguez, González and Delannoy74). Additionally, steroid hormones differentially regulate GnRH neuronal activity and subsequent LH secretion in response to β-NGF stimulation. While progesterone levels do not significantly affect GnRH neuronal system, estradiol demonstrates potent modulatory effects. Ovariectomized llamas show markedly attenuated β-NGF-induced LH secretion by approximately 67%, an effect reversible by estradiol replacement therapy (Refs Reference Silva, Recabarren, Recabarren, Adams and Ratto58, Reference Silva, Smulders, Guerra, Valderrama, Letelier, Adams and Ratto59, Reference Carrasco, Singh, Ratto and Adams75). These findings highlight estradiol’s facilitatory role in β-NGF-mediated ovulation induction.
Ovarian steroidogenesis
Estradiol
Experiments in vitro demonstrated that cultured human granulosa cells showed increased estradiol secretion in the presence of NGF through elevated FSHR mRNA and formation of functional FSHR receptor (Ref. Reference Salas, Julio-Pieper, Valladares, Pommer, Vega, Mastronardi, Kerr, Ojeda, Lara and Romero17). Consistently, a mouse model with ovarian-specific overproduction of NGF (17NF mice), showed an elevated production of estradiol in response to gonadotropins, associated with enhanced expression of Cyp19a1, which encodes the enzyme catalyzing the synthesis of estradiol (Ref. Reference Garcia-Rudaz, Dorfman, Nagalla, Svechnikov, Söder, Ojeda and Dissen76).
Progesterone
Experiments have found that spβ-NGF/NGF increased progesterone concentration whether in vivo of llamas or in primary granulosa and theca cells from llamas and cattle respectively, accompanied by upregulated transcription of Star, Hsd3b, Cyp11a1 and downregulated expression of Cyp19a1. The changes in gene expression of steroidogenic enzyme contributed to the rapid transition from estradiol synthesis to progesterone synthesis, and exerted luteal nutrition and promoted angiogenesis effects (Refs Reference Dissen, Parrott, Skinner, Hill, Costa and Ojeda77, Reference Silva, Ulloa-Leal, Valderrama, Bogle, Adams and Ratto78, Reference Valderrama, Goicochea, Silva and Ratto79, Reference Valderrama, Ulloa-Leal, Silva, Goicochea, Apichela, Argañaraz, Sari, Paiva, Ratto and Ratto80). Further research revealed that spβ-NGF activated the ERK1/2 signalling pathway and produced the aforementioned effects (Ref. Reference Valderrama, Ulloa-Leal, Silva, Goicochea, Apichela, Argañaraz, Sari, Paiva, Ratto and Ratto80). In addition, a mouse model with ovarian-specific NGF overproduction demonstrated consistent progesterone production in response to gonadotropins, associated with enhanced expression of Cyp17a1 (Ref. Reference Garcia-Rudaz, Dorfman, Nagalla, Svechnikov, Söder, Ojeda and Dissen76). However, studies in human granulosa cells cultured in vitro have shown opposite results, probably due to different receptors distribution in granulosa cells among species (Ref. Reference Salas, Julio-Pieper, Valladares, Pommer, Vega, Mastronardi, Kerr, Ojeda, Lara and Romero17).
Testosterone/Androgen
NGF promoted testosterone and androgen production. In bovine TrkA-transfected theca cells and bovine follicle wall pieces, which contained both theca cells and granulosa cells, NGF increased testosterone and androgen production, which could be due to an increase in the theca cell number and upregulated gene expression of steroidogenic enzyme HDS17B, by which androstenedione is converted to testosterone in the theca cells (Refs Reference Dissen, Parrott, Skinner, Hill, Costa and Ojeda77, Reference Stewart, Gao, Flaws, Mercadante, Dias, Canisso and Lima81). Consistently, 17NF mouse showed an elevated production of testosterone in response to gonadotropins, associated with enhanced expression of Hsd17b, which encodes the enzyme catalyzing the synthesis of testosterone (Ref. Reference Garcia-Rudaz, Dorfman, Nagalla, Svechnikov, Söder, Ojeda and Dissen76). METTL 14 is a kind of writer in the process of m6A. NGF was shown to activate the PI3K/AKT signalling pathway via METTL14, thereby regulating testosterone synthesis in porcine theca cells (Ref. Reference Luo, Zhao, Zhang, Chen, Chen, Shen, He, Cao, Chen, Wang, Wang, Zong, Zhou and Li82). These findings may further clarify the mechanisms underlying NGF-mediated testosterone synthesis and offer novel therapeutic targets for diseases associated with excessive testosterone production in female animals.
Reproductive system disease
Polycystic ovary syndrome
PCOS is the most common reproductive endocrine disorder, with a prevalence of 6–15% (Ref. Reference Yang83). Experimental evidence from rodent models demonstrates a strong association between NGF dysregulation and PCOS pathogenesis. In rats, steroid administration significantly upregulated intraovarian NGF and p75NTR synthesis, resulting in polycystic ovaries marked by multiple follicular cysts (Ref. Reference Lara, Dissen, Leyton, Paredes, Fuenzalida, Fiedler and Ojeda84). Similarly, ovarian-specific transgenic NGF overexpression in mice recapitulated key PCOS features, including both reproductive and metabolic abnormalities (Refs Reference Dissen, Garcia-Rudaz, Paredes, Mayer, Mayerhofer and Ojeda43, Reference Wilson, Chen, Dissen, Ojeda, Cowley, Garcia-Rudaz and Enriori85). Histopathological analysis revealed that NGF overproduction disrupted normal folliculogenesis, causing follicular arrest and promoting antral follicle apoptosis (Ref. Reference Dissen, Garcia-Rudaz, Paredes, Mayer, Mayerhofer and Ojeda43). These consistent findings across species strongly implicate ovarian sympathetic hyperactivation in PCOS development and progression (Ref. Reference Wilson, Chen, Dissen, Ojeda, Cowley, Garcia-Rudaz and Enriori85). Clinical observations further support this mechanistic link. IVF patient analyses demonstrated significantly elevated NGF levels in patients with PCOS, with approximately twofold higher follicular fluid concentrations and sixfold greater granulosa cell expression compared to controls (Ref. Reference Dissen, Garcia-Rudaz, Paredes, Mayer, Mayerhofer and Ojeda43). However, conflicting data from Buyuk failed to replicate these NGF elevation findings, highlighting potential methodological or population differences in clinical PCOS studies (Ref. Reference Buyuk and Seifer20). Recently, clinical evidence from Robeva’s study demonstrated that significantly elevated NGF in women with PCOS showed a positive correlation with plasma-free normetanephrine levels, suggesting that NGF may contribute to PCOS pathogenesis through enhanced sympathetic noradrenergic activity (Ref. Reference Robeva, Elenkova, Kirilov and Zacharieva86). Furthermore, NGF may promote the occurrence and development of PCOS pathological features by enhancing androgen and testosterone secretion. In in vitro experiments, NGF acted on bovine theca cells transfected with TrkA, resulting in excessive production of androgen and testosterone (Ref. Reference Stewart, Gao, Flaws, Mercadante, Dias, Canisso and Lima81). Consistent with this, in in vivo experiments, 17NF mice with ovarian local overexpression of NGF produced excessive testosterone under the stimulation of gonadotropins (Ref. Reference Garcia-Rudaz, Dorfman, Nagalla, Svechnikov, Söder, Ojeda and Dissen76). However, there is currently no experimental evidence to show that excessive NGF can induce a PCOS disease model.
Diminished ovarian reserve
Diminished ovarian reserve (DOR) refers to a condition characterized by a reduction in the quantity and quality of oocytes stored in the ovaries, leading to infertility, assisted reproductive technology failure and miscarriage. Based on previous literature, the diagnosis is primarily based on decreased AMH levels and antral follicle counts, and elevated FSH levels (Ref. Reference Cohen, Chabbert-Buffet and Darai87). The prevalence of DOR has increased significantly in recent years (Refs Reference Federici, Rossetti, Moleri, Munari, Frixou, Bonomi and Persani88, Reference Guo, Liu, Xu, Qi, Xu, Wu, He, Qin and Chen89, Reference Golezar, Ramezani Tehrani, Khazaei, Ebadi and Keshavarz90, Reference Lagergren, Hammar, Nedstrand, Bladh and Sydsjö91). Women with DOR exhibit significantly higher follicular fluid NGF levels compared to those with normal ovarian function. This finding is supported by clinical data from IVF patients, particularly in advanced maternal age populations (Ref. Reference Palumbo, Giuffrida, Gulino, Leonardi, Cantarella and Bernardini92). However, this apparent NGF elevation may represent a compensatory mechanism rather than a primary pathogenic factor, as evidenced by reduced expression of NGF receptors TrkA and p75NTR in cumulus cells from DOR patients undergoing assisted reproductive technology. These in vitro findings suggest that impaired NGF receptor signalling, rather than NGF availability itself, may contribute to the low ovarian reserve (Ref. Reference Buyuk, Santoro, Cohen, Charron and Jindal35).
Endometriosis
Endometriosis, which affects approximately 10% of reproductive-aged women, is a chronic gynaecological disorder characterized by ectopic implantation of endometrial tissue (Refs Reference Viganò, Parazzini, Somigliana and Vercellini93, Reference Afshari-Stasiak, Kobierzycki, PIotrowska, Rycerz, Wilczyński and Szubert94). While extensive research has been conducted, the exact pathogenesis and molecular mechanisms underlying this prevalent condition remain incompletely understood (Ref. Reference Bulun, Monsivais, Kakinuma, Furukawa, Bernardi, Pavone and Dyson95). Notably, immunohistochemical analyses revealed significantly stronger immunostaining for both NGF and its receptor TrkA in endometriotic lesions from patients experiencing deep dyspareunia compared to those without this symptom (Refs Reference Peng, Zhan, Alotaibi, Alkusayer, Bedaiwy and Yong96, Reference Barcena de Arellano, Arnold, Lang, Vercellino, Chiantera, Schneider and Mechsner97, Reference Wessels, Wu, Leyland, Wang and Foster98). In mouse, anti-NGF treatments reduced endometriosis-associated pain (Ref. Reference Zaninelli, Fattori, Heintz, Wright, Bennallack, Sim, Bukhari, Terry, Vitonis, Missmer, Andrello, Anchan, Godin, Bree, Verri and Rogers8). These findings suggest that NGF signalling may contribute specifically to endometriosis-associated sexual pain. Supporting this notion, clinical investigations have demonstrated elevated follicular fluid NGF levels in endometriosis patients relative to women with male factor infertility (Ref. Reference Buyuk and Seifer20).
Conclusions
NGF is a pivotal regulator of female reproductive functions, including follicular assembly, follicle development, oocyte maturation, ovulation and steroidogenic regulation. In particular, the role of NGF in inducing ovulation by acting on the hypothalamus has garnered considerable scholarly attention. Clinically, dysregulated NGF signalling has been implicated in the pathogenesis of several reproductive disorders, such as PCOS, DOR and endometriosis. These investigations may yield novel diagnostic and therapeutic approaches for managing NGF-associated reproductive conditions. Nevertheless, current research has two major limitations that warrant emphasis: (1) the underlying mechanisms by which NGF participates in reproductive physiology remain incompletely understood and (2) the clinical translation of NGF-based applications is still at an early stage. Future investigations should prioritize these two aspects to advance our understanding and optimize the clinical utility of NGF in reproductive health.
Author contribution
Y.R.L.: Conceptualization – Writing – Original Draft Preparation – Reviewing and Editing; S.H., X.Z.: Writing – Review and Editing – Supervision; W.G.: Supervision – Funding Acquisition; L.N.Y, L.H.W., S.Y.G.: Review and Editing; J.Y., R.Y.: Conceptualization – Supervision – Funding Acquisition.
Funding statement
This work was supported by the National Natural Science Foundation of China (R. Y., grant numbers 82571891 and 82171632); (W. G., grant number 82201805) and Peking University Clinical Scientist Training Program supported by ‘the Fundamental Research Funds for the Central Universities’ (W. G., grant number BMU2025PYJH029).
Competing interests
The authors declare none.
Disclaimer/Publisher’s Note
The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
Ethical standards
The authors assert that this work does not involve human or animal experimental procedures.


 Open access
Open access