


Introduction
Recurrent drought, intensified by climate change, represents a major constraint for perennial crops and threatens the long-term sustainability of tree-based agroecosystems worldwide. According to UNCCD projections, more than three-quarters of global land areas may experience increased drought risk by 2050 (UNCCD 2022). Rising temperatures and fluctuating rainfall increase the vulnerability of trees to this type of stress, particularly in drought-prone regions such as the Mediterranean.
Almond (Prunus dulcis) is one of the most economically important nut crops in these regions and has traditionally been cultivated under rainfed or deficit-irrigation systems (Egea et al. Reference Egea, Nortes, González-Real, Baille and Domingo2010; Alonso et al. Reference Alonso, Kodad, Gradziel, Badenes and Byrne2012; Prgomet et al. Reference Prgomet, Pascual-Seva, Morais, Aires, Barreales, Ribeiro, Silva, Barros and Gonçalves2020). In Mediterranean semi-arid environments, recurrent drought typically refers to successive growing seasons characterized by limited and irregular rainfall, resulting in repeated soil moisture deficits during critical phenological stages. Such conditions expose perennial crops to cumulative water stress effects across years rather than isolated short-term episodes, with carry-over impacts on subsequent seasons’ yield and tree survival (Prgomet et al. Reference Prgomet, Pascual-Seva, Morais, Aires, Barreales, Ribeiro, Silva, Barros and Gonçalves2020; Moldero et al. Reference Moldero, López-Bernal, Testi, Lorite, Fereres and Orgaz2021; Barreales et al. Reference Barreales, Capitão, Bento, Casquero and Ribeiro2023). The increasing frequency and intensity of drought events now challenge both orchard productivity and varietal sustainability (Moldero et al. Reference Moldero, López-Bernal, Testi, Lorite, Fereres and Orgaz2022; Fernandes de Oliveira et al. Reference Fernandes de Oliveira, Mameli, De Pau and Satta2023; Gohari et al. Reference Gohari, Imani, Talaei, Abdossi and Asghari2023). Improving drought resilience has therefore become a strategic priority for almond breeding and management programs (Yadollahi et al. Reference Yadollahi, Arzani, Ebadi, Wirthensohn and Karimi2011; Ayup et al. Reference Ayup, Yang, Gong and Guo2022; Barreales et al. Reference Barreales, Capitão, Bento, Casquero and Ribeiro2023; Calderón-Pavón et al. Reference Calderón-Pavón, García-Tejero, Noguera-Artiaga, Lipan, Sendra, Hernández, Herencia-Galán, Carbonell-Barrachina and Zuazo2025).
Plant genetic resources, including local landraces, hybrids and exotic varieties derived from regional prospection, constitute a critical reservoir of adaptive traits. These materials often harbour valuable characteristics such as drought tolerance, phenological plasticity and productivity that may be underrepresented in elite commercial cultivars (Kandemir and Saygili Reference Kandemir and Saygili2015; Karaat Reference Karaat2020). Their characterization is essential for pre-breeding strategies aimed at developing climate-resilient almond cultivars.
Previous studies have explored almond responses to water stress using morphological, physiological and biochemical traits, including leaf area, photosynthesis, stomatal conductance, chlorophyll content, nutrient status and stress metabolites (Gikloo and Elhami Reference Gikloo and Elhami2012; Hamzaoui et al. Reference Hamzaoui, Oukabli and Moumni2014; Zokaee-Khosroshahi et al. Reference Zokaee-Khosroshahi, Esna-Ashari, Ershadi and Imani2014; Ranjbar et al. Reference Ranjbar, Imani, Piri and Abdoosi2021; Yildirim et al. Reference Yildirim, Şan, Yildirim, Çeli̇K, Bayar and Karakurt2021). While these studies have provided valuable insights, most have relied on short-term experiments or limited genotype sets, making it difficult to assess trait stability across years and environmental variability. Moreover, the relative importance of these traits as reliable indicators of drought tolerance remains insufficiently clarified.
This knowledge gap is particularly critical for perennial species such as almond, where long-term performance and trait stability are essential selection criteria. Multi-year evaluations are therefore required to identify traits that are both discriminating and consistent under recurrent drought conditions.
To address these limitations, the present study was designed as a 3-year screening of a diverse almond genetic collection, including local landraces, clones, hybrids and reference cultivars grown under recurrent drought. The objectives were to characterize inter-genotypic variability under persistent water stress, to identify stable and performance-related indicator traits of drought tolerance and to classify genotypes according to their adaptive responses, thereby supporting the use of almond genetic resources in pre-breeding and climate-resilient improvement programmes.
Materials and methods
Working approach
The screening was conducted over three consecutive cropping seasons (2022, 2023 and 2024), during which climate conditions varied (Fig. 1). The objective is to conduct an in-depth analysis of the impact of climatic variations on the almond collection and to identify the most determining parameters in the context of recurring droughts. During the first year of the trial (2022 season), yield and 23 agro-physiological, morphological and biochemical parameters were measured between May and August, corresponding to the main vegetative and productive period of the crop. This initial phase provided a complete data set to establish a baseline assessment of the impact of climatic conditions on almond tree performance. In the second and third years, the focus was only on the parameters identified as determining traits during the first year of the study. These parameters were measured during the same phenological window to ensure inter-annual comparability. Based on these traits, classified as stress indicators, the studied genotypes were classified into three groups (sensitive, medium and resistant). This approach allows the optimization of resources and efforts by concentrating monitoring on the most important indicators of almond tree resilience to challenging climatic conditions.
Schematic overview of the study workflow.

Plant material
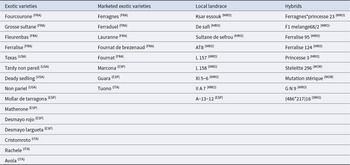
The genotypes selected for this study originate from a diverse collection of almond trees, including landraces, hybrids developed for their tolerance and well-established commercial varieties used as controls. The choice of these genotypes is based on their performance under water stress conditions, demonstrating their capacity to produce in a promising manner. This selection highlights the genetic diversity and the interest of these resources for varietal improvement (Table 1). Photographs illustrating tree architecture of almond genetic resources are provided in Supplementary (Fig. S4).
Almond genotypes included in the study, with their genetic status and geographic origin (Monastra and Raparelli Reference Monastra and Raparelli1997)

Geographical origin is indicated as a superscript after each genotype name: MOR (Morocco), FRA (France), ESP (Spain), ITA (Italy) and USA (United States).
Experimental conditions
The trials were conducted at the Ain Taoujdate experimental station (INRA Morocco), located in a semi-arid Mediterranean climate. Annual precipitation is approximately 320 mm in 2022, 132 mm in 2023 and 310 mm in 2024, with irregular and concentrated rainfall mainly during the winter period, which favours prolonged periods of drought.
The soil texture is sandy-clay, moderately low in organic matter (1.38%), with a relatively low water retention capacity, with a usable water reserve of 1.7 mm cm−1, reinforcing crop sensitivity to water stress, with a slightly alkaline pH (around 7.9), and low salinity (electrical conductivity 0.32 ms/cm) (Table 2). The orchard was managed under rainfed conditions throughout the study period, which allowed the evaluation of genotype responses to naturally occurring seasonal drought typical of semi-arid Mediterranean environments. Standard cultural practices (fertilization and plant protection) were applied uniformly to all trees according to local recommendations.
Physical and chemical properties of soil in the experimental orchard

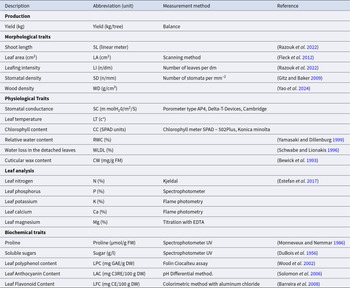
Parameters measured
Mature, healthy leaves from light-exposed branches were collected from each tree in the morning at mid-height. Sampling was carried out during the main vegetative period, May, of each experimental year, at comparable phenological stages across years. These samples were then placed in paper bags and stored in a cooler at low temperatures before being transferred to the laboratory. At the same time, the fruit was harvested at maturity to assess yield.
In the laboratory, the leaves were washed with distilled water to determine their water status (RWC, WLDL), calculate their LA, and measure their SD and WD. Some of the leaves were dried for 24 hours and then finely ground to analyse their nutritional status. Another portion of the leaves was then finely ground and extracted into ethanol (80%) before being stored in airtight bags at −20°C until analysis. These preparations ensured the quality and integrity of the samples for subsequent biochemical analysis. Additionally, SC and CC were measured directly in the orchard. Table 3 summarizes the parameters measured using the appropriate measurement methods.
Description, abbreviation and measurement method of the evaluated parameters

For each year of assessment, the coefficient of variation (CV, %) was calculated for each measured trait in order to quantify inter-genotypic variability. The CV was computed as the ratio of the standard deviation of a trait to the mean of that trait, expressed as a percentage (Pélabon et al. Reference Pélabon, Hilde, Einum and Gamelon2020)
 \begin{equation*}C{V_{t,Y}} = \frac{{SD}}{{Mean}} \times 100\end{equation*}
\begin{equation*}C{V_{t,Y}} = \frac{{SD}}{{Mean}} \times 100\end{equation*}Statistical analysis
The data collected were analysed using an analysis of variance (ANOVA) to compare the means of the various parameters. Mean comparisons between genotypes were performed using LSD test at P < 0.05 and P < 0.01. Next, a principal component analysis (PCA) was performed to identify the main factors influencing stress effects. Based on these results, a hierarchical ascending classification (HAC) was performed to group genotypes that exhibited similar responses to water stress. All these analyses were carried out using SPSS v22 software.
Results
First-year analysis
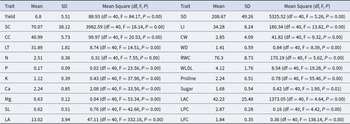
A descriptive statistical analysis was performed for the 22 traits measured during the first year of evaluation (Table 4). The results revealed substantial variability among genotypes for most of the studied parameters. Mean values and associated standard deviations highlighted pronounced differences in physiological, morphological, and biochemical traits. Yield averaged 6.8 kg per tree, with high variability (standard deviation of 5.51), ranging from 0 to 20.0 kg. Mean values for physiological traits, especially SC (70.07) and CC (40.99), also showed wide variation, as reflected by their relatively large standard deviations.
Descriptive analysis and analysis of variance of traits of the first year

Trait abbreviations: yield, SC: stomatal conductance, CC: chlorophyll content, LT: leaf temperature, N: leaf nitrogen, P: leaf phosphorus, K: leaf potassium, Ca: leaf calcium, Mg: leaf magnesium, SL: shoot length, LA: leaf area, SD: stomatal density, LI: leafing intensity, CW: cuticular waxes, WD: wood density, RWC: relative water content, WLDL: water loss from detached leaves, proline, sugar, LAC: leaf anthocyanin content, LPC: leaf polyphenol content, and LFC: leaf flavonoid content. ** Significant at 0.01.
The analysis of variance (Table 4) indicated that all the measured traits were highly significant (P < 0.01), demonstrating a strong genotypic effect and confirming the existence of substantial variability within the population. The high value of mean square, of SD (MS = 5325.52), SC (MS = 3982.59) and LAC (1373.05), highlights their discriminating potential. The other traits showed lower MS values but remained significant.
In addition, Pearson correlation analysis was conducted to explore the relationships among the measured parameters (Table S1). The results showed that chlorophyll content (CC) was positively correlated with yield (r = 0.39, P < 0.05) and CS (r = 0.51, P < 0.01). LA also exhibited a correlation with yield, reflecting its association with productive performance. WD showed significant negative correlations with both leaf area (r = –0.43, P < 0.01) and leaf nitrogen content (r = –0.38, P < 0.01). Moreover, WLDL and RWC were also showed associations with production and growth performance.
PCA highlighted the main traits contributing to genotypic differentiation under drought (Table S2). Nine principal components were retained, explaining a large proportion of the total variance. Among them, the first two components accounted for the most meaningful variation, mainly associated with CC, CS, N, LA, WD, SD, RWC, WLDL and yield.
Based on these three complementary approaches, inter-genotypic variance, correlations with performance, and contribution to PCA. These traits were selected as priority discriminant parameters, justifying their use for monitoring and classification in multi-year screening.
Multi-year analysis
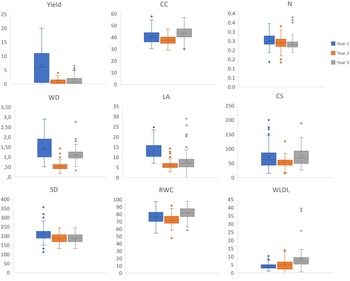
Box-plot analysis (Fig. 2) of the nine traits measured over three cropping seasons showed a gradual decline in yield over the years, from 6.7 kg/tree in the first year to 3.9 and then 3.34 kg. This decreasing trend was accompanied by a reduction in variability. CC and N remain stable overall, at around 40 ± 3 and 2.5 ± 0.15, respectively, with consistent inter-genotypic variability across years.
Box plot distribution of traits over three years.

In contrast, LA, SD and WD exhibited a marked reduction between the first and second years, followed by partial recovery during the third year, although substantial variability persisted. SD decreased by approximately 12% in the second year, then stabilized in the third year. In addition, RWC remained almost stable throughout the 3 years (around 77% on average), while maintaining significant variability within each year.
The observed increase in WLDL is because initial water stress before detachment often weakens the cell structure, reducing the leaf’s ability to retain moisture and leading to more rapid dehydration. This graphical analysis confirms that inter-annual climatic conditions influence the expression of most traits. This observation is confirmed by the analysis of variance (Table S3).
The coefficients of variation calculated for the different traits highlighted clear differences in inter-genotypic variability across years (Table S3). Physiological traits such as chlorophyll content, leaf nitrogen concentration, stomatal density, and relative water content generally exhibited low to moderate CV values (<14.5%), indicating limited dispersion among genotypes. In contrast, yield, leaf area and water loss-related parameters showed higher CV values, reflecting greater variability.
PCA was performed for each year to identify the traits contributing most strongly to genotype differentiation (Fig. S2 and S3). In the first year, the first component explained approximately 30% of the total variance, mainly influenced by WD, N, LA, and CC. In the second year, characterized by increased water stress with 59% less rainfall than the first year (Fig. S1), the trait structure shifted, with WD, N, CC and yield becoming the most discriminating variables. In the third year, the results indicated a trend similar to that of the first year, with some changes in the importance of the traits. Across all years, CC, N, WD and yield consistently exhibited high loadings on PC1.
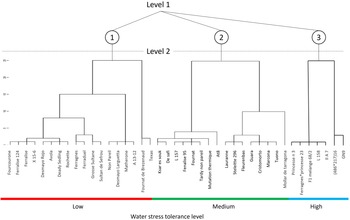
The classification of genotypes was established on two levels, based on traits considered discriminating according to their correlation with overall production performance and their inter-annual stability (Fig. 3). The first level relied on CC, considered a fundamental indicator of plant vitality and metabolic activity. The second level integrates both agronomic and structural traits, namely yield, N, WD and LA, which proved relevant for refining differentiation between genotypes.
Ward’s classification of 41 genotypes into 2 levels. Level 1 is based on CC. Level 2 integrates leaf nitrogen, wood density, yield and leaf area to refine genotype classification.

Hierarchical clustering was used to group genotypes according to their similarity based on discriminating traits. Three main groups emerge consistently. The first group was partly composed of well-known commercial varieties. These varieties exhibit low CC and low yields. The second group displayed intermediate trait values, whereas the third group showed superior performance under drought conditions, combining high CC and good yield.
To illustrate contrasting phenotypic traits among drought-response groups, a subset of nine almond genotypes was selected for photographic documentation. These genotypes were chosen based on the hierarchical classification results, with three genotypes representing each response group (tolerant, intermediate and sensitive). For each selected genotype, photographs of the whole tree, leaves and fruits were taken under field conditions (Fig. 4).
Representative phenotypic traits of nine almond genetic resources selected from the hierarchical classification. Three genotypes were chosen from each drought response group (tolerant (A), intermediate (B), and sensitive (C)). For each genotype, tree architecture, leaf morphology and fruit traits are shown.

Discussion
Recurrent drought represents a major constraint for almond cultivation, particularly in Mediterranean and semi-arid environments, where productivity and long-term orchard sustainability depend on the capacity of genotypes to maintain functional stability under repeated water deficit (Fernandes de Oliveira et al. Reference Fernandes de Oliveira, Mameli, De Pau and Satta2023). In this context, identifying reliable phenotypic traits that consistently discriminate genotypes across years is a central challenge for the characterization and valorization of almond genetic resources.
This study applied a progressive multi-year screening strategy to identify drought-tolerant genotypes and a reduced set of informative phenotypic traits. During the first year, a wide range of agro-physiological, morphological and biochemical parameters was intentionally evaluated to capture the broad spectrum of almond responses to drought. Comparable exploratory screens in diverse genetic resources panels have highlighted the same type of large inter-genotypic variability (Hamzaoui et al. Reference Hamzaoui, Oukabli and Moumni2014). Based on the convergence of ANOVA, correlation and multivariate analyses, the initial set of parameters was reduced to a limited number of key traits, without losing key information (Dale and Beyeler Reference Dale and Beyeler2001; Al-Ashkar et al. Reference Al-Ashkar, Al-Suhaibani, Abdella, Sallam, Alotaibi and Seleiman2021).
Yield is a key integrative trait that consistently declines under water stress in almond and other crops (Lipiec et al. Reference Lipiec, Doussan, Nosalewicz and Kondracka2013; Sofi et al. Reference Sofi, Rehman, Ara and Gull2018; Kanbar et al. Reference Kanbar, Chege, Lantos, Kiss and Pauk2020; Khan et al. Reference Khan, Wang, Akbar and Alhoqail2025). Its moderate and consistent association with chlorophyll content (r ≈ 0.40) suggests that maintaining photosynthetic capacity contributes to performance stability rather than acting as a standalone determinant of tolerance. Similar observations have been reported in drought-adapted genotypes maintaining pigment stability and gas exchange activity under stress (Nemeskéri et al. Reference Nemeskéri, Molnár, Vígh, Nagy and Dobos2015; Correia et al. Reference Correia, Braga, Martins, Correia, Pinto and Canhoto2023). While water deficit is known to alter photosynthetic activity (Pandey et al. Reference Pandey, Ansari, Atri, Singh, Gupta and Bhat2018; Coşkun Reference Coşkun2025; Toprak and Coşkun Reference Toprak and Coşkun2025), previous findings were often based on annual crops or short-term responses. In contrast, this study demonstrates the stability (CV < 12%) and discriminatory value (r ≈ 0.7 with PC1) of chlorophyll content over multiple years in a diverse almond collection, reinforcing its relevance as a stable physiological indicator of drought response.
Water stress alters nutrient uptake (Seleiman et al. Reference Seleiman, Al-Suhaibani, Ali, Akmal, Alotaibi, Refay, Dindaroglu, Abdul-Wajid and Battaglia2021). Nitrogen concentration decreased under stress, affecting leaf growth and reproductive biomass production as previously highlighted by Bakhtiari et al. (Reference Bakhtiari, Mousavi, Yadegari, Haghighati and Martínez-García2025). The association between N and growth (r = 0.4; P < 0.05) and physiological performance (r = −0.4; P < 0.01) reinforces the well-established observation that foliar nitrogen is often correlated with traits related to growth and biomass accumulation (Pan et al. Reference Pan, Wang, Yan, Wu, Guo, Peng, Wu, Li, Wang, Su and Liu2024), suggesting that nitrogen efficiency may represent an additional functional component contributing to genotype differentiation within drought-adapted germplasm.
The negative correlation between leaf area and yield (Table S1) indicates that smaller leaves may represent an adaptive advantage under water deficit. This structural reduction is consistent with studies showing adaptive leaf plasticity in tolerant cultivars (Mahalle et al. Reference Mahalle, Chetia, Dey, Sarma, Baruah, Kaldate, Verma and Modi2021; Fernandes de Oliveira et al. Reference Fernandes de Oliveira, Mameli, De Pau and Satta2023; Qiao et al. Reference Qiao, Hong, Jiao, Hou and Gao2024). Wood density (WD) was also negatively correlated with both LA and N (Table S1), suggesting a potential trade-off between hydraulic safety and growth capacity. Genotypes with higher WD may possess thicker xylem walls, enhancing resistance to cavitation during severe drought, as previously described by Greenwood et al. (Reference Greenwood, Ruiz-Benito, Martínez-Vilalta, Lloret, Kitzberger, Allen, Fensham, Laughlin, Kattge, Bönisch, Kraft and Jump2017). The repeated contribution of leaf area and wood density to multivariate differentiation across years indicates that structural characteristics may also participate in the long-term adaptive strategies distinguishing tolerant and sensitive genotypes.
The relatively low interannual variability observed for CC, N, WD and yield, together with their consistent contribution to PCA differentiation across years (Fig. 2; Fig. S2–S3), supports their usefulness in multi-year screening. Coefficient of variation analysis thus served as a confirmatory tool supporting the robustness of previously identified key traits. Selecting parameters based on their correlations, stability, and repeated contribution to genotype discrimination ensures that only variables with high discriminating ability and relevance to performance are retained for multi-year evaluation (Füzy et al. Reference Füzy, Kovács, Cseresnyés, Parádi, Szili-Kovács, Kelemen, Rajkai and Takács2019). These traits should therefore not be interpreted independently but rather as complementary components of a multi-trait framework allowing robust genotype differentiation under recurrent drought conditions.
The integration of trait stability, functional relevance, and multivariate contribution enabled the establishment of a two-level hierarchical classification of genotypes. Level 1 is based on chlorophyll content, which captures a broad physiological gradient of plant functioning under drought and corresponds largely to the first principal component of the multivariate analysis, whereas Level 2 integrates yield, nitrogen content, wood density and leaf area to refine genotype differentiation by incorporating structural and nutritional dimensions of drought adaptation. This classification consistently separated sensitive, intermediate and drought-adapted groups across years (Fig. 3). Interestingly, several widely cultivated commercial varieties were classified as sensitive, whereas tolerant genotypes were predominantly represented by locally adapted selections and advanced breeding materials such as Princesse n° 3’, ‘Ferragnes*princesse 23’, ‘F1 melange 68/2’, ‘L 158’, ‘II A 7’, ‘(486*217)16’ and ‘GN9’. This finding highlights the importance of genetic diversity derived from local selection and hybrids for improving drought adaptation, supporting previous observations that underscore the value of genetic recombination and local adaptation in enhancing drought resilience (Acquaah Reference Acquaah2012).
This framework is particularly relevant for pre-breeding and genetic resource utilization, providing a practical basis for early-stage genotype selection targeting rainfed Mediterranean environments. Identified tolerant genotypes may serve either as parental material in breeding programs. However, validation in different environments and with molecular support (Cabrita et al. Reference Cabrita, Apostolova, Neves, Marreiros and Leitão2014; Hamzaoui et al. Reference Hamzaoui, Oukabli and Moumni2014) remains necessary before their practical use in improvement programs. Overall, the study demonstrates that progressive multi-year phenotypic screening can efficiently reduce trait complexity while preserving biological relevance, offering a reproducible framework for the characterization and utilization of almond genetic resources under drought-prone conditions.
Conclusions
This study provides a multi-year evaluation of almond genetic resources under rainfed semi-arid conditions. It identifies a reduced set of phenotypic traits consistently associated with drought tolerance in this environment. By combining inter-annual stability, discriminating power, and functional relevance, chlorophyll content, leaf nitrogen, wood density, yield and leaf area emerged as key indicators for differentiating genotypes within a diverse collection of genetic material. Unlike short-term, multi-trait approaches, the proposed framework integrates complementary traits to enhance the robustness of genotype classification under recurrent drought at a single site. This represents a practical step forward for multi-genotype screening and characterization of almond genetic resources in similar Mediterranean rainfed systems.
The identification of potentially tolerant genotypes within local selections and breeding material highlights the value of underutilized genetic resources for drought adaptation. However, these classifications should be considered preliminary and context-dependent. The genotypes identified nonetheless represent promising parental material for pre-breeding programs and candidates for further evaluation in a rainfed Mediterranean environment. Future research should validate the proposed combination of traits at other sites and environmental gradients before proposing them as generalizable selection criteria.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/S147926212610063X.
Author contributions
Conceptualization, H.A., R.R., L.H. and J.B.; Methodology, H.A., A.N. and R.R. Data acquisition, H.A and N.H. Writing, H.A. and R.R. Supervision, R.R. and L.H. and J.B. All authors have approved the final version of the manuscript.
Funding statement
This research did not receive any specific grant.
Competing interests
The authors report there are no competing interests to declare.
Availability of data and materials
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
AI statement
Artificial intelligence-based tools were used during the preparation of this manuscript solely for language editing and grammatical improvement (Deepl and grammarly); no scientific content was generated.






 Open access
Open access