


Introduction
The cutthroat eel family Synaphobranchidae comprises 13 genera and 57 species (Fricke et al. Reference Fricke, Eschmeyer and Fong2026) of mostly deep-sea fishes found in all oceans, some of which exceed 1 m in length. It differs from most other eel families in having ventrolateral to ventral gill openings and the posterior nostril close in front of the eye, at or below its midlevel. Synaphobranchid leptocephali are unique among eels in having an elongated vertical or diagonal eye (Robins & Robins Reference Robins, Robins and Böhlke1989). Robins & Robins (Reference Robins and Robins1976, Reference Robins, Robins and Böhlke1989) divided the family into three subfamilies: Synaphobranchinae, Simenchelyinae and Ilyophinae; the latter subfamily is the most diverse, with eight genera and 43 valid species, including eight species of Ilyophis (Fricke et al. Reference Fricke, Eschmeyer and Fong2026). Over the last 25 years, the Synaphobranchidae received a fair amount of attention through the preparation of Food and Agriculture Organization (FAO) species identification guides for the living marine resources of several ocean regions (Smith Reference Smith, Carpenter and Niem1999, Reference Smith and Carpenter2002, Reference Smith, Carpenter and De Angelis2016) and recent books on the fishes of Australia and New Zealand (Smith Reference Smith, Gomon, Bray and Kuiter2008, Smith & Stewart Reference Smith, Stewart, Roberts, Stewart and Struthers2015). Two cutthroat eels have been reported from the Southern Ocean, namely Diastobranchus capensis and Histiobranchus bathybius (e.g. Gon & Heemstra Reference Gon and Heemstra1990, Hanchet et al. Reference Hanchet, Stewart, McMillan, Clark, O’Driscoll and Stevenson2013); however, Southern Ocean specimens of the latter species are believed to be Histiobranchus australis as defined by Karmovskaya & Merrett (Reference Karmovskaya and Merrett1998).
In this paper, we report the first finding of a cutthroat eel tentatively identified as Ilyophis cf. maclainei in the Southern Ocean. The specimen was captured in a bottom trawl station on Astrid Ridge in the Kong Haakon VII Sea off Dronning Maud Land in March 2019.
Materials and methods
Retrieval of specimen
Trawling was conducted during a multi-gear scientific cruise aimed at collecting diverse oceanographic, ecosystem and biodiversity data in this area. The cruise was organized and led by the Norwegian Polar Institute with the Institute of Marine Research (Norway), the University of Bergen (Norway), the Arctic University of Norway, the University of Science and Technology (Norway) and the Stellenbosch University (South Africa) onboard the Norwegian research vessel Kronprins Haakon. Two bottom trawl stations were conducted during the survey; for further survey details, see Kauko et al. (Reference Kauko, Hattermann, Ryan-Keogh, Singh, de Steur and Fransson2021). At station 72, the trawl included a single ilyophine eel specimen, which was collected and delivered frozen to the University Museum of Bergen for further processing.
List of abbreviations
- BOLD = Barcode of Life Data System
- CSIRO = Commonwealth Scientific and Industrial Research Organisation
- HL = head length
- IMR = Institute of Marine Research
- NMNZ = National Museum of New Zealand
- RV = research vessel
- SAIAB = South African Institute of Aquatic Biodiversity
- TL = total length
- UM = University Museum of Bergen
- USNM = United States National Museum
- ZMUB = University Museum of Bergen, vertebrate collection
Morphometrics and meristics
Measurements and counts follow Böhlke (Reference Böhlke and Böhlke1989) and Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) (Tables I & II). Head length was measured from tip of snout to upper pectoral-fin base. Postorbital length was measured from the posterior edge of eye to upper pectoral-fin base. The adnasal pore is included in the infraorbital canal count. Vertebrae were counted from radiographs; predorsal and preanal vertebrae were counted starting from the vertebra positioned vertically below the first detectable fin ray anteriorly. Prepectoral, predorsal, preanus and postanus pores were counted using a microscope with 3–6× magnification. Pores were counted three times independently at different time points to review counting results. Detail images were taken and stacked using a Leica M205c stereomicroscope with a Leica DMC 5400 camera and Leica Application Suite (LAS 4.13), stacked with Zerene Stacker V-1.04.
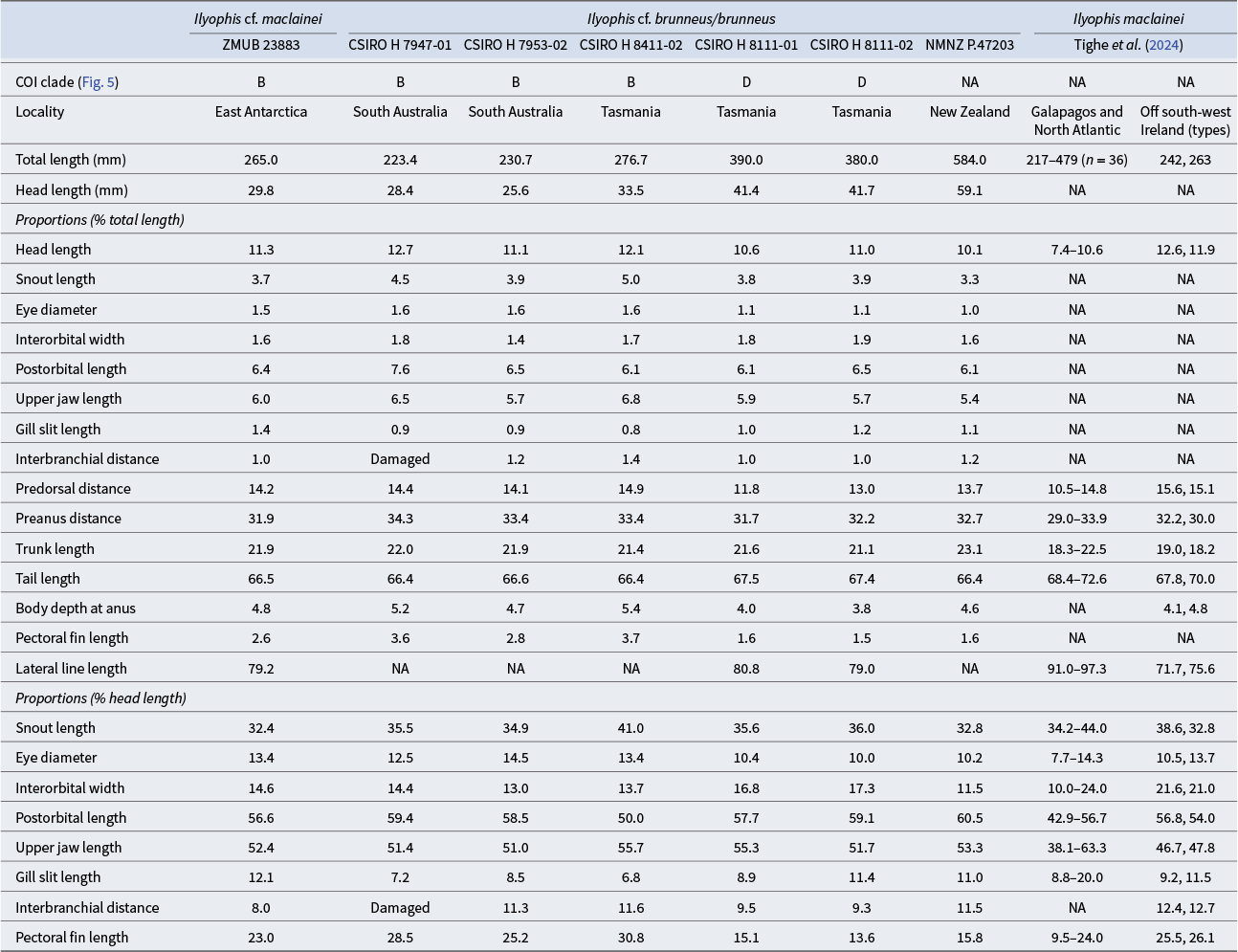
Proportional measurements data for the Antarctic Ilyophis cf. maclainei and comparative material of Ilyophis maclainei, Ilyophis cf. brunneus and Ilyophis brunneus.

COI = cytochrome oxidase I; NA = not available.
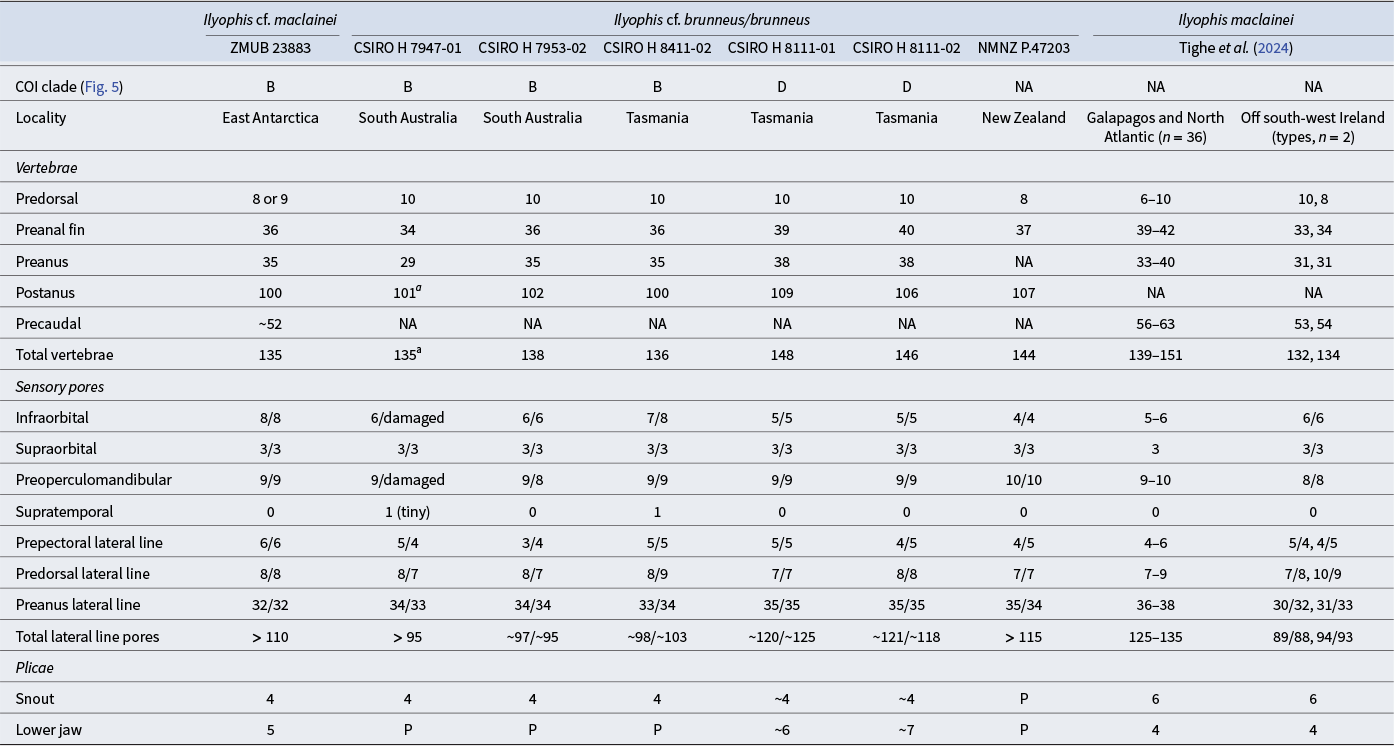
Meristic data for the Antarctic Ilyophis cf. maclainei and comparative material of Ilyophis maclainei, Ilyophis cf. brunneus and Ilyophis brunneus.

NA = not available.
a Tail tip damaged, possibly +1 vertebra.
P = plicae present but not counted.
Ilyophis cf. maclainei ZMUB 23883, 265 mm TL, off Dronning Maud Land, 68°28´S, 11°48´E, bottom trawl, 1515–1500 m, IMR Antarctic Cruise 2019 (2019702), station 72, RV Kronprins Haakon, 16 March 2019.
Comparative material identified as Ilyophis brunneus or Ilyophis cf. brunneus
CSIRO H 7947-01, 223+ mm TL (BOLD ID FOAP254-17, BW-A13560), Great Australian Bight, South Australia, 33°30.97´S, 130°15.90´E, beam trawl, 978–1013 m, IN2015_C02/382, RV Investigator, 15 December 2015; CSIRO H 7953-02 (BOLD ID FOAP082-16, BW-A13573), 231 mm TL, Great Australian Bight, South Australia, 34°37.50´S, 130°16.80´E, beam trawl, 2007–2067 m, IN2015_C02/449, RV Investigator, 18 December 2015; CSIRO H 8411-02, 277 mm TL (BOLD ID FOAO1929-19, BW-A15011), south-east of Pedra Branca seamount, Tasmania, 44°17.81´S, 147°06.90´E, beam trawl, 1392–1448 m, IN2018_V06/011, RV Investigator, 25 November 2018; CSIRO H 8111-01, 390 mm TL (BOLD ID FOAO1048-18, BW-A14241) and CSIRO H 8111-02, 380 mm TL (BOLD ID FOAO2508-20, BW-A15668), east of Seymour, Freycinet Commonwealth Marine Reserve, Tasmania, 41°43.83´S, 149°07.18´E, beam trawl, 2751–2820 m, IN2017_V03/004, RV Investigator, 18 May 2017. NMNZ P.47203, 584 mm TL, New Zealand.
Mitochondrial genome reconstruction
We sampled muscle tissue from the Antarctic specimen (ZMUB 23883) prior to 75% ethanol preservation and fixed it in 96% EtOH. We extracted the DNA from the muscle tissue (2 mg) using the EchoLUTION Tissue DNA Kit (BioEcho Life Sciences, Cologne, Germany). We adapted the manufacturer protocol by increasing the lysis time to 60 min and retrieving 80 μl of eluted DNA. After DNA extraction, we followed the SRSLY® PicoPlus kit protocol (Claret Bioscience, Scotts Valley, CA, USA) to construct single-stranded DNA libraries. We proceeded to 10 indexing polymerase chain reaction (PCR) cycles and eluted the final DNA libraries in 20 μl low tris-EDTA (TE; 0.1 M). We converted the DNA library with the Adept Library Compatibility Kit for sequencing on a 2 × 150 bp low-output AVITI™ lane (Element Biosciences, San Diego, CA, USA). We trimmed the generated raw reads for adapter sequences as well as for low-quality (below 10) nucleotides at both 5´ and 3´ ends using Cutadapt v.4.7 (Martin Reference Martin2011). We assessed read quality before and after processing with FastQC v.0.11.7 (Andrews Reference Andrews2010).
We reconstructed the mitochondrial genome using the baiting and iterative mapping approach implemented in MITObim V1.9.1 (Hahn et al. Reference Hahn, Bachmann and Chevreux2013). We first merged the paired reads using FLASh V1.2.11 (Magoč & Salzberg Reference Magoč and Salzberg2011). Then, we used the complete mitochondrial genome of I. brunneus (GenBank accession: NC013634; Inoue et al. Reference Inoue, Miya, Miller, Sado, Hanel and Hatooka2010) as a reference to reconstruct the full mitochondrial sequence. To annotate the assembled mitochondrial genome, we used the MitoAnnotator pipeline from MitoFish V4.09 (Zhu et al. Reference Zhu, Sato, Sado, Miya and Iwasaki2023). We estimated the nucleotide composition of the entire mitogenome with MEGA X V10.1 (Kumar et al. Reference Kumar, Stecher, Li, Knyaz and Tamura2018). Finally, we depicted the result as a circular map using Chloroplot (Zheng et al. Reference Zheng, Poczai, Hyvönen, Tang and Amiryousefi2020).
Phylogenetic placement using mitochondrial DNA
To place our specimen phylogenetically using mitochondrial DNA (mtDNA) sequence data, we extracted part of the mitochondrial cytochrome oxidase I (COI) gene from the complete mitochondrial assembly. This locus is widely used in DNA barcoding of fish species, and appropriate reference sequences are available from BOLD (https://boldsystems.org/). BOLD sequence and specimen identifiers have been added for comparative material in consecutive order.
We downloaded 125 synaphobranchid sequences from the BOLD Data Portal and aligned this dataset with our newly generated sequence of the Antarctic specimen using MAFFT V-7 online (Katoh & Standley Reference Katoh and Standley2013). The aligned dataset was then converted to Phylip format and analysed with Maximum Likelihood (ML) using RaxML (Stamatakis Reference Stamatakis2014) performing 100 bootstrap replicates after a thorough ML search under the GTR Gamma substitution model. A COI sequence of the pelican eel Eurypharynx pelecanoides (BOLD ID ANGBF31318-19) was chosen as outgroup. After identifying relevant clusters, the dataset was trimmed to contain 28 sequences of Ilyophis, Simenchelys and potentially misidentified sequences assignable to Ilyophis. This trimmed dataset was then used to compute a haplotype network using Hapsolutely V-0.2.2. (Vences et al. Reference Vences, Miralles, Brouillet, Ducasse, Fedosov and Kharchev2021). Maximum parsimony was selected for the tree inference and Fitchi (Matschiner Reference Matschiner2016) was used for graphical production of the haplotype network. The corresponding figure was created in the freeware Inkscape V-0.92 (www.inkscape.org).
Results
Ilyophis cf. maclainei ZMUB 23883 (Figs 1–3)
Description
Body long and slender, tapering posteriorly beyond anus, rapidly tapering in last third of body; maximum body depth near middle of trunk, ~20 times in TL; body depth at anus slightly smaller, ~21 in TL. Trunk length 4.6, predorsal distance 7.0 and preanal distance 3.1, all in TL. Head fairly small, ~9 in TL, its maximum depth at a distance of somewhat less than snout length behind eye, 1.5 in HL; snout moderately long, 3.1 in HL; eye small, covered with translucent skin, its diameter 2.4 in snout length and 7.5 in HL; interorbital width somewhat larger than eye diameter, 6.8 in HL; upper jaw long, slightly over half HL; postorbital length slightly shorter than upper jaw; gill small, 1.1 in eye diameter and 8.3 in HL; interbranchial width, measured at lower (anterior) end of gill 1.3 in gill length and 10.9 in HL. Anterior nostril tubular, its aperture similar in size to posterior nostril; posterior nostril round with raised thickened rim and with membranous skin covering most of its lower half.
Lateral line long, apparently incomplete (difficult to determine its posterior end due to damaged skin), its length ~1.3 in TL; total pores at least 110.
Pectoral fin small, inserted on lower half of body, its length 4.3 in HL and 38.5 in TL; pectoral-fin rays 14 (right fin, dissected).
Dorsal-fin origin above proximal part of pectoral fin and above 8th or 9th vertebra; predorsal distance 7.0 and trunk length 4.6, both in TL; anal-fin origin below 37th vertebra, anus below 36th vertebra, preanus distance 3.1 in TL (Fig. 1b,d).
Ilyophis cf. maclainei ZMUB 23883. a. Left lateral view of the freshly collected specimen after death. b. Right lateral view of the ethanol-preserved specimen. White line in front of the anal opening is a plastic string holding the specimen for photographing. c. Arrangement of lateral scales. d. Anal opening.

Head without scales. Body covered with non-imbricate slender scales arranged in basket-weave pattern with rounded to somewhat pointed ends; most scales’ width ~5 times in their length (Fig. 1c). Scale density decreases in the last fifth of body length, perhaps due to abrasion.
Head sensory canal pores (Fig. 2a–d): first and second supraorbital pores close to each other and in front of anterior nostril; third pore slightly behind anterior nostril; first infraorbital pore (= adnasal pore) adjacent to posterior edge of anterior nostril; second pore below and slightly anterior to first pore; third pore placed close above margin of upper jaw, at approximately a third of distance between tip of snout and posterior nostril; fourth and fifth pores are below posterior nostril and posterior edge of pupil, respectively; sixth to eighth pores are spread around posterior edge of orbit, the last two pores placed higher than middle of orbit; the first seven preoperculomandibular pores are spread at more or less equal distances along the mandible from the tip of lower jaw to a vertical below rictus (Fig. 2b). Preoperculomandibular pores 8 and 9 in preopercular region on posterior head, pore 9 only visible on the left lateral side of the specimen (Fig. 2d).
Ilyophis cf. maclainei (ZMUB 23883): head cephalic system using image stacking. a. & c. Right lateral side of head. a. shows cephalic supraorbital canal pore III, infraorbital pores 1–8 and preopercular-mandibular pores 1–7; c. is used to visualize preopercular-mandibular pore 8, pore 9 not visible. b. & d. Left lateral side of head. b. shows cephalic supraorbital canal pores I–III, infraorbital pores 1–5 and preopercular-mandibular pores 1–7 (pore 5 not visible due to damage); d. is used to visualize preopercular-mandibular pores 8 and 9, the latter not visible in c. Infra- and preopercular-mandibular pores indicated by Arabic numerals, supraorbital pores indicated by Roman numerals.

Mouth large, filled with food remains which are difficult to remove without damaging the teeth. Mouth gape reaching to approximately one eye diameter behind eye (Figs 1 & 2). Teeth large, not compound, conical and recurved. Maxilla with two rows, some anterior teeth enlarged; intermaxillary teeth at least seven, arranged in a circular tooth patch in front of but separated from vomerine teeth by a small gap, and including the largest teeth as well as small ones; vomer with two rows of teeth, the anterior teeth enlarged. Each side of dentary with a medial row of large and well-spaced teeth followed laterally by two to three irregular rows of smaller close-set teeth (Fig. 3a,b).
Ilyophis cf. maclainei (ZMUB 23883): a. close-up of upper jaw; b. close-up of lower jaw.

Colour
The freshly captured Antarctic specimen (Fig. 1a) is generally brown with dark brown head and blackish abdominal region. The dorsal and anal fins are dark with a pale edge; the pectoral fin is brown and the caudal fin is blackish with a pale tip. The nostrils and pores of the head sensory canals are pale. The lateral line is darker than the body, but its pores are pale; it becomes gradually paler behind anus then barely noticeable farther posteriorly. This colour pattern remains largely unchanged in the preserved specimen (Fig. 1b). In the preserved specimen, the pectoral fins are dark with pale edges, the tip of the lower jaw is pale; there is a pale area behind the eye (Fig. 2); the mouth is dark, but the tissue surrounding the teeth series is pale (Fig. 3); and the anal opening has a pale rim surrounded by naked skin of darker pigment (Fig. 1d).
Mitochondrial genome
The sequencing output consisted of a total of 7 429 481 raw paired reads. After removing low-quality reads and merging paired reads, we retrieved 6 305 009 reads, which were used as input for the mitogenome reconstruction. The assembled and circularized mitogenome (GenBank accession number: PX673979) has an average coverage of 131× per nucleotide and is 16 733 bp long, comprising 13 protein-coding genes, 22 transfer RNAs (tRNAs), two ribosomal RNAs (rRNAs) and one control region (or ‘d-loop’). Most genes are located on the heavy, positive strand of the genome, while nad6 and eight tRNAs (tRNA-Glu, tRNA-Pro, tRNA-Gln, tRNA-Ala, tRNA-Asn, tRNA-Cys, tRNA-Tyr and tRNA-Ser) stand on the light, negative strand. The nucleotide composition of the mitochondrial genome is: A = 31.5%, T = 26.1%, G = 16.5%, C = 25.9%, with a biased A + T content (57.6%), similar to other species from the same family (Fig. 4).
Mitochondrial genome map of Ilyophis cf. maclainei ZMUB 23883. Genes encoded by the heavy strand are shown inside the circle, and those encoded by the light strand are shown outside the circle. Each gene type is depicted by a different colour. The pale grey inner circle represents the GC content of the mitogenome. ATP = adenosine triphosphate; COX = cytochrome oxidase; GC = guanine and cytosine; NADH = nicotinamide adenine dinucleotide hydrogen; rRNA = ribosomal RNA; tRNA = transfer RNA.

Phylogenetic placement using COI
The aligned dataset used for the phylogenetic placement of the Antarctic specimen comprised a total of 128 sequences and was trimmed to 651 nucleotides for each specimen sequence (Supplementary Material 1). The phylogenetic inference derived from the data generally shows weak node support values for ‘deeper’ phylogenetic nodes. In our dataset, the genera Meadia, Histiobranchus, Ilyophis, Synaphobranchus and Dysomma appear paraphyletic. Ilyophis sequences are placed in four different clades (Fig. 5a, clades A–D) and a single lineage (specimen Ilyophis sp. FOAO1074-18, BW-A14270); their phylogenetic interrelationships remain unclear given the weak bootstrap support (74%) and the presence of Simenchelys parasitica between two Ilyophis clades (Fig. 5a, clades C and D), as well as a potentially misidentified I. cf. brunneus within the Meadia spp. clade (Fig. 5a, clade A), rendering it paraphyletic. I. brunneus/cf. brunneus is present in five clades, suggesting a species complex and/or specimen misidentifications due to specimens of variable genetic diversity morphologically keying to I. brunneus. The Antarctic specimen reported here (ZMUB 23883) is placed in a monophyletic and well-supported clade containing specimens of I. brunneus, I. cf. brunneus, and Ilyophis sp. (Fig. 5a, clade B). Its sister clade (Fig. 5a, clade C), comprising specimens from the north-west Pacific (Taiwan and China), seems to be yet another cryptic lineage. Interestingly, clades B and D include specimens from both the Atlantic (Walvis and mid-Atlantic ridges) Ocean and from around Tasmania, implying that two morphologically similar species may inhabit these locations. The sister clade to clades B and C contains only sequences of S. parasitica from circumglobal locations.
Ilyophis cf. maclainei (ZMUB 23883): phylogenetic placement. a. Maximum Likelihood-based phylogenetic reconstruction with bootstrap support indicated at nodes as black dots or numbers if below 95% support (100 replicates) based on partial cytochrome oxidase I data. Estimate performed under the GTR Gamma substitution model. Eurypharynx pelecanoides chosen as outgroup. b. Haplotype network reconstruction of clade B as shown in a. based on the Jukes-Cantor model. Black dots on lines connecting haplotypes indicate mutational steps; unique haplotypes dominate. Barcode of Life Data System records are indicated after species names.

The haplotype network reconstruction of the trimmed dataset, including sequences of the aforementioned Ilyophis and Simenchelys clades, shows high haplotype diversity, with only two haplotypes occurring more than once. The clades are separated by more than 30 substitutions on average. The Antarctic specimen shows a unique haplotype separated by two substitutions from its closest conspecifics (Fig. 5b).
Comparisons
Morphometric and meristic data of the Antarctic Ilyophis specimen are compared with data (Tables I & II) from three Australian specimens, also members of clade B (Fig. 5a), identified originally as I. cf. brunneus (CSIRO H 7947-01 (FOAP254-17); CSIRO H 7953-02 (FOAP082-16); CSIRO H 8411-02 (FOAO1929-19)). In addition, the data of the Antarctic specimen are compared with data from two Tasmanian specimens, identified as I. brunneus (CSIRO H 8111-01 (FOAO1048-18); CSIRO H 8111-02 (FOAO2508-20)), from clade D (Fig. 5a); data from a specimen identified as I. brunneus (NMNZ P.47203) from New Zealand (no DNA sequence data available); as well as data from Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) for the latter species, including the holotype, and for the two type specimens of I. maclainei (no genetic data for aforementioned type specimens). The meristic and morphometric data of the Antarctic specimen place it between I. brunneus and I. maclainei. It is closer to the latter species in the total number of vertebrae (Table II) but is different in having more preanus (35 vs 31, respectively) and preanal fin vertebrae (36 vs 33 and 34). It also has more infraorbital pores (8 vs 6 in I. maclainei), more preoperculomandibular pores (9 vs 8, respectively) and more prepectoral pores (6 vs 4 or 5). As for body proportions (Table I, as percentage TL and HL), the predorsal distance of the Antarctic specimen (7.0 times in TL) is shorter than in I. maclainei (6.4 and 6.6 in TL), its trunk length is larger (4.6 vs 5.2 and 5.5 in TL), its interorbital width is narrower (6.8 vs 4.6 and 4.8 in HL), its upper jaw is longer (1.9 vs 2.1 in HL), its gill slit is somewhat longer (8.3 vs 8.6 and 10.9 in HL) and its interbranchial width is smaller (10.9 vs 7.8 and 8.1 in HL). The Antarctic specimen is generally similar to the three Australian specimens (FOAP254-17; FOAP 082-16; FOAO 1929-19) in the comparative material, initially identified as I. cf. brunneus, which are also members of clade B (Fig. 5). It differs from these three Australian specimens in having a slightly shorter preanus distance (3.1 vs 2.9–3.0 in TL, respectively), longer gill slits (8.3 vs 11.8–14.7 in HL), shorter interbranchial distance (12.5 vs 8.6 and 8.9 in HL) and smaller pectoral fin (4.3 vs 3.2 and 4.0 in HL). The New Zealand and two clade D Tasmanian specimens we examined distinctly differ from the Antarctic specimen and the other three clade B Australian specimens in having more preanal-fin vertebrae (37–40 vs 34–36, respectively), more postanus vertebrae (107–109 vs 100–102) and a higher total number of vertebrae (144–148 vs 135–138), as well as fewer infraorbital pores (4–5 vs 6–8), smaller eyes (9.6–9.9 vs 6.9–8.0 in HL) and a shorter pectoral fin (6.3–7.3 vs 3.2–4.3 in HL). The counts data of Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) for specimens of I. brunneus include the values we had for the two Tasmanian specimens of clade D and the New Zealand specimens for most meristic characters (Table II); the New Zealand specimens had 37 preanal-fin vertebrae as opposed to 39–42 in Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) and four infraorbital pores (vs 5–6), and both the New Zealand and the two Tasmanian clade D specimens had fewer preanus pores (34–35 vs 36–38). In the morphometric characters, the Antarctic and the Australian clade B specimens have somewhat larger heads and shorter tail sections (Table I) than the I. brunneus of Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024). The broad range of the morphometric data Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) (Table II) presented as %HL for I. brunneus (Table I) precludes meaningful comparison.
Comparison of Gilbert’s (Reference Gilbert1891) description of the 380 mm TL (15 inches) holotype of I. brunneus, also designated by him as the type species of the genus Ilyophis, confirmed that the Antarctic specimen is a species of this genus. However, the smaller (265 mm TL) Antarctic specimen differs from the holotype, as described by Gilbert (Reference Gilbert1891), in the following characters: the interbranchial distance is 0.76 of the gill slit length (Table I) but equal to the latter in the holotype; the pectoral fin is longer (4.35 vs 6.0 in HL, respectively); the dorsal and anal fins are dark with pale edges, the caudal fin has a pale tip and the pectoral fin has a pale edge vs all fins being dark in the holotype. Gilbert’s (Reference Gilbert1891) description of the holotype’s dentition is in general agreement with the Antarctic specimen, but comparison is difficult due to the absence of illustration in Gilbert’s (Reference Gilbert1891) description. For example, he considered the jaw teeth small, but in the Antarctic specimen they certainly are not, although they are distinctly smaller than the vomerine teeth. Gilbert (Reference Gilbert1891) may have used ‘small’ in a relative sense. We did not observe significant differences in dentition among all the examined specimens.
Discussion
The Antarctic ilyophine eel reported here comprises the southernmost record of the synaphobranchid subfamily Ilyophinae. The two other synaphobranchid eels known to occur in the Southern Ocean, D. capensis and H. australis, are classified in the subfamily Synaphobranchinae and have been reported (the latter species sometimes as H. bathybius) mostly from the sub-Antarctic islands (45–49°S) of the Indian Ocean (Gon & Heemstra Reference Gon and Heemstra1990, Duhamel et al. Reference Duhamel, Gasco and Davaine2005) and from south (54°S) of the Falkland Islands (Brickle & Laptikhovsky Reference Brickle and Laptikovsky2002). Hanchet et al. (Reference Hanchet, Stewart, McMillan, Clark, O’Driscoll and Stevenson2013) reported the former species from a higher latitude (71°S) in the Ross Sea. Duhamel et al. (Reference Duhamel, Belchier, Hulley, Romain, Koubbi, Vacchi, De Broyer, Koubbi, Griffiths, Raymond, D’Udeken d’Acoz and van de Putte2014) presented a map with records of synaphobranchid eels from the Ross Sea region, albeit without species details.
The wide range of the morphometric characters of I. brunneus presented as %HL (Table I) by Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) (Table II) could be explained by allometric growth or the presence of more than one species in their material. It should be noted that apart from the holotype of I. brunneus (type locality Galapagos Islands), all the material of this species used by these authors had been collected in the North Atlantic Ocean long before the advent of DNA-based molecular taxonomy. The same is true for the material used by Robins & Robins (Reference Robins, Robins and Böhlke1989), who illustrated a 440 mm TL specimen from the Bahamas that has a pectoral fin distinctly smaller than the pectoral fin of the Antarctic specimen.
The majority of the meristic and morphometric data of the Antarctic specimen ZMUB 23883 and the eight specimens of our comparative material fall within the range of most characters provided by Tighe et al. (Reference Tighe, Smith, Merrett, Frable and Zajonz2024) for I. brunneus and I. maclainei (Tables I & II). Unfortunately, the sample size of the comparative material of our study and of I. maclainei (two specimens) is too small to identify the species morphologically with certainty, and, as far as we know, no DNA sequence data are available for the latter species. Comparison of the Antarctic specimen with the description of the holotype of I. brunneus by Gilbert (Reference Gilbert1891) also was inconclusive. However, the phylogenetic analysis provides some clues when considered together with the morphological data. There is a distinct difference in the vertebral counts (except the predorsal count) and preanus lateral-line pores between the members of clades B and D of the phylogenetic tree (Fig. 5a), most of which were originally identified as I. cf. brunneus or I. brunneus. These differences - the total vertebral count in particular (Table II) - suggest that clade D is a better candidate to represent the true I. brunneus.
The Antarctic specimen and the three Australian specimens of clade B (Fig. 5a) align with I. maclainei in most of the morphological characters we studied (Tables I & II). Notable exceptions include interorbital width (13.0–14.6 %HL vs 21.0 and 21.6 %HL in I. maclainei) and upper jaw length (51.0–55.7 %HL vs 46.7 and 47.8 %HL). Given the small number of specimens in clade B and I. maclainei, as well as the absence of genetic material for the latter species, we preferred to err on the side of caution and identified the Antarctic specimen as I cf. maclainei. Further studies combining existing data with molecular and morphological data, ideally also including DNA sequence information of type material, from more Southern Hemisphere specimens are necessary to resolve this taxonomic conundrum. To that end, the full mitochondrial genome generated in this study, whose gene order and content (Fig. 4) are highly similar to those of other Synaphobranchidae (Inoue et al. Reference Inoue, Miya, Tsukamoto and Nishida2004, Reference Inoue, Miya, Miller, Sado, Hanel and Hatooka2010), will serve as a future resource for more in-depth molecular analyses of Ilyophis taxonomy.
Supplementary material
To view supplementary material for this article, please visit http://doi.org/10.1017/S0954102025100564.
Acknowledgements
We would like to express our sincere thanks to Anne Helene Soldberg Tandberg (UM), Ingvar Byrkjedal (retired, UM) and Gunnar Langhelle (retired, UM) for their help processing the specimen, to Ulrich K Schliewen (SNSB-ZSM) and Per Gunnar Fjelldal (IMR) for the help radiographing ZMUB 23883 as well as to Andrew Stewart and Jeremy Barker (NMNZ) for making a specimen and its radiograph available for examination. Kathrine Kongshaven (UM), Bjørn Reidar Olssen (UM) and Sergey Bogorodsky (SAIAB) kindly assisted with imaging. The late David G. Smith (USNM) kindly shared his encompassing knowledge of eels. Anne Kari Sveistrup (IMR) and other cruise participants are thanked for their invaluable help sampling at sea. Alastair Graham and Sharon Appleyard assisted with registration and COI sequencing of the CSIRO specimens, respectively.
Competing interests
The authors declare none.
Author contributions
NS and OG designed the study. Specimen and metadata collection by EHH. Morphological data collection by NS, JJP and OG. Molecular data collected and analysed by NS and SS. NS and OG drafted the manuscript with additions from and reviews by all authors.








 Open access
Open access