


Introduction
Agricultural intensification in North America is tied heavily to extensive, large-scale monocultures of field crops, such as soybean, Glycine max (Linnaeus) Merrill (Fabaceae) (Johnston Reference Johnston2014; Bigelow and Borchers Reference Bigelow and Borchers2017; Malaj et al. Reference Malaj, Freistadt and Morrissey2020). Such intensification leads to the reduction of natural and semi-natural habitats that support wild pollinators and diverse floral resources, contributing to global biodiversity loss (Kremen et al. Reference Kremen, Williams and Thorp2002; Deguines et al. Reference Deguines, Jono, Baude, Henry, Julliard and Fontaine2014; Raven and Wagner Reference Raven and Wagner2021). Given the limited floral diversity in many agricultural landscapes, both wild and managed pollinators might take advantage of mass-flowering soybean fields as a source of floral resources (Gill and O’Neal Reference Gill and O’Neal2015; Esquivel et al. Reference Esquivel, Parys and Brewer2021). Despite soybean being primarily self-pollinated, there is mounting evidence that bee visitation can enhance its yield (reviewed in Garibaldi et al. Reference Garibaldi, Schulte, Nabaes Jodar, Gomez Carella and Kremen2021 and Esquivel et al. Reference Esquivel, Parys and Brewer2021). However, the perception among growers that soybean does not benefit from insect pollination persists (de O. Milfont et al. Reference de O. Milfont, Rocha, Lima and Freitas2013), complicating the adoption of pollinator-friendly practices among field crop growers.
Although soybean flowers can autonomously self-pollinate, pod formation is often low, and pollination deficits are common (de O. Milfont et al. Reference de O. Milfont, Rocha, Lima and Freitas2013; Garibaldi et al. Reference Garibaldi, Schulte, Nabaes Jodar, Gomez Carella and Kremen2021; da Cunha et al. Reference da Cunha, Chacoff, Sáez, Schmucki, Galetto and Devoto2023). Both honey bees, Apis mellifera Linnaeus (Hymenoptera: Apidae), and wild bees can enhance soybean yields through pollination services, although these increases vary among cultivars and as a function of management practices and plant health (Esquivel et al. Reference Esquivel, Parys and Brewer2021; Adelabu et al. Reference Adelabu, Franke, Delves, Terzi, Membretti, Taylor and Delves2023). To date, most studies have focused on honey bees, but other insects – including wild bees, flies (Diptera), beetles (Coleoptera), and butterflies (Lepidoptera) – can also pollinate soybean (Garibaldi et al. Reference Garibaldi, Schulte, Nabaes Jodar, Gomez Carella and Kremen2021 and references therein; Levenson et al. Reference Levenson, Sharp and Tarpy2022). As in other crops, the complementary pollination provided by honey bees and wild pollinators can further enhance soybean productivity (de O. Milfont et al. Reference de O. Milfont, Rocha, Lima and Freitas2013).
Recent studies have documented non-Apis diversity in soybean fields using passive sampling techniques, such as pan trapping (e.g., Gill and O’Neal Reference Gill and O’Neal2015; Wheelock et al. Reference Wheelock, Rey and O’Neal2016; Torrez et al. Reference Torrez, Beauzay, St. Clair and Knodel2022). Although these methods are useful for collecting baseline data, passive sampling cannot on its own distinguish insects that visit crop flowers and potentially act as pollinators from those that are simply passing through without pollinating (Levenson et al. Reference Levenson, Sharp and Tarpy2022). Active netting of insects visiting flowers is a better approach to identifying crop-pollinating species, but such surveys remain rare. One notable exception documented 35 wild bee species foraging on soybean flowers in southeastern United States of America (Levenson et al. Reference Levenson, Sharp and Tarpy2022). However, findings from one region may not generalise across different regions or landscapes.
In addition to wild bees, other wild anthophilous insects, such as aculeate wasps (Hymenoptera: Apocrita, Aculeata), might also visit soybean flowers for nectar (Sholahuddin et al. Reference Sholahuddin, Wijayanti, Arniputri and Widyaningrum2019) and could incidentally contribute to pollination (Rader et al. Reference Rader, Bartomeus, Garibaldi, Garratt, Howlett and Winfree2016; Brock et al. Reference Brock, Cini and Sumner2021). Despite their potential, the role of wasps as crop pollinators, including in soybean, remains largely unstudied.
The objective of this study was to identify wild hymenopteran species (bees and aculeate wasps) that visit soybean flowers and carry soybean pollen in southeastern Canada (Ontario and Quebec), thereby likely contributing to soybean pollination. Identifying the wild pollinator fauna visiting soybean flowers could help raise awareness among field crop growers about the role of wild pollinators in soybean production. It also provides a baseline to guide efforts to enhance pollinator conservation in landscapes dominated by field crops.
Methods
Study sites
The study was conducted at 19 soybean fields along the Ottawa River in Renfrew County, Ontario, and the Outaouais region of Quebec, Canada (Fig. 1). This region is composed primarily of agricultural lands, dominated by dairy and beef cattle and field crop farms. Field sites ranged in size from 4 ha to 40 ha, and all but three were conventionally managed. Field margins were often narrow but supported a variety of herbaceous flowering plants and woody vegetation. No honey bee hives were located directly adjacent to the sampled fields.
Map of the 19 soybean study sites in Ontario and Quebec, Canada, where pollinator sampling was conducted.

Pollinator sampling and identification
Pollinator surveys were conducted from 18 July to 9 August 2023, when soybean was flowering. Each soybean field was surveyed once during this period. Sampling was conducted between 10:00 and 17:00, local time, on days that were favourable for bee activity (clear to partly cloudy skies, temperatures above 15 °C by 10:00). At each field, two observers walked parallel to the crop rows, 1 m inside the field from the margin, at a steady pace for 30 minutes. All bees and aculeate wasps observed actively foraging on soybean flowers were collected using a handheld net, with time stopped while handling specimens. Insects were placed in a cooler during transport to the laboratory and were then kept in a freezer at −20 °C until sampled for pollen (see Soybean pollen sampling and identification section, below).
Following pollen sampling, each insect was pinned and identified. Bees were identified to species when possible, using relevant taxonomic keys (Packer et al. Reference Packer, Genaro and Sheffield2007; Gibbs Reference Gibbs2010, Reference Gibbs2011; Williams et al. Reference Williams, Thorp, Richardson and Colla2014; Ascher and Pickering Reference Ascher and Pickering2020). Wasps were identified to tribe (Bohart and Menke Reference Bohart and Menke1976) or species (Buck et al. Reference Buck, Marshall and Cheung2008), depending on the level of certainty achievable for each specimen based on the authors’ level of expertise. Specimens are stored in the Forrest Laboratory collection at the University of Ottawa (Ottawa, Ontario, Canada).
Soybean pollen sampling and identification
We sampled pollen from each insect’s corbiculae or scopae (female bees only), as well as from other parts of their body (all specimens). Corbicular and scopal pollen was removed using fine-tipped forceps and transferred into a Petri dish. The full pollen load was then crushed and thoroughly mixed using a metal pick. We subsampled a small portion (approximately one-quarter the size of a grain of rice) and mounted it in glycerine jelly tinted with basic fuchsin (fuchsin gel), based on Kearns and Inouye (Reference Kearns and Inouye1993). Similarly, we analysed pollen sampled from other parts of each insect’s body by rubbing a 1-mm3 cube of fuchsin gel on the thorax, abdomen, legs, and head of the insect before melting it on a slide with an alcohol lamp. All tools were cleaned with 70% ethanol between specimens to minimise pollen contamination.
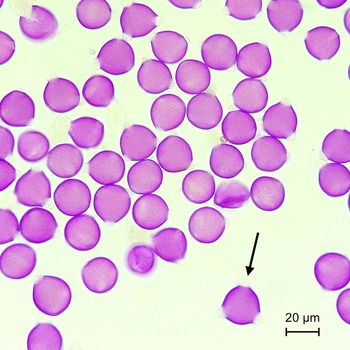
We used a compound microscope (Nikon, Tokyo, Japan) at 400× magnification to identify soybean pollen using a pollen reference library from Auer (Reference Auer2020), as well as pollen sampled from soybean flowers collected at our study sites (Fig. 2). For corbicular and scopal pollen, we systematically examined the first 200 pollen grains encountered on each slide. We counted pollen grains along parallel vertical field-of-view transects, starting with transects close to the midpoint of the sample and progressing outwards. Then, we visually estimated the number of soybean pollen grains out of the total 200 grains examined and categorised the proportions using the following scale: at least 25%, less than 25%, trace (approximately 1%), or none (no soybean pollen observed). Given the variation in bee species’ sizes and pollen-carrying structures, we did not attempt to measure pollen quantity. Instead, we focused on the proportion of soybean pollen within each sample, using it as an indicator of this crop’s significance as a forage resource. For pollen sampled from other parts of an insect’s body, soybean pollen was scored as either present or absent across the entire slide. This was done to ensure the detection of soybean pollen on specimens that do not actively collect pollen but might carry it incidentally (e.g., male bees and wasps). We defined major soybean pollen carriers as insects with 25% or more soybean pollen in their corbicular or scopal pollen loads, regardless of the presence of soybean pollen on other parts of their bodies. Insects with less than 25% soybean pollen in their corbicular or scopal loads, or those without corbicular or scopal loads (e.g., male bees and wasps) but carrying soybean pollen on other parts of their bodies, were defined as minor soybean pollen carriers. Those carrying no soybean pollen either in their corbicular or scopal pollen loads or on their bodies were defined as noncarriers.
Fuchsin-stained soybean pollen grains sampled from a bee’s scopa. Arrow indicates typical shape and size of triporate soybean pollen.

Results
From a total of 19 person-hours of observation, we collected 77 wild bees and four wasps from soybean flowers across 19 different fields (ranging from 0 to 21 specimens and from 0 to 5 species per field). These soybean flower visitors comprised 18 bee species from eight genera and three families, and four wasp species from two families (Table 1). Most of the bees collected were generalist bumble bees, followed by ground-nesting bees (i.e., bees that excavate underground nests; Supplementary material, Dataset S1).
List of wild bees and wasps collected while foraging on soybean flowers, with number of individuals classified as major, minor, or non-carriers of soybean pollen

Of the 18 bee species collected from soybean flowers, only five – Augochlorella aurata (Smith) (Hymenoptera: Halictidae), Bombus impatiens Cresson (Hymenoptera: Apidae), Bombus ternarius Say (Hymenoptera: Apidae), Halictus confusus Smith (Hymenoptera: Halictidae), and Lasioglossum versatum Robertson (Hymenoptera: Halictidae) – were found at more than three sites, and together, they accounted for 59.7% of total bee abundance. Bumble bee workers were the most frequently collected soybean flower visitors across all sites. In addition to bees and wasps, other insects (ants, flies, and small beetles) were observed on soybean flowers but were not collected.
The majority of insects collected from soybean flowers (74 of 81; 91.4%) carried some soybean pollen on their bodies. Indeed, only one flower visitor species, Bombus fervidus (Fabricius) (Hymenoptera: Apidae), could not be confirmed as a soybean pollen carrier: the sole specimen collected, a male, had no soybean pollen on his body. However, the proportion of soybean pollen actively collected by female bees varied among species. In six bee species, at least some individuals were identified as major soybean pollen carriers. These included four bumble bee species (Hymenoptera: Apidae), B. impatiens, B. rufocinctus Cresson, B. sandersoni Franklin, and B. ternarius, one sweat bee species (Hymenoptera: Halictidae), Lasioglossum admirandum (Sandhouse), and one mining bee species (Hymenoptera: Andrenidae), Andrena wilkella (Kirby). Most specimens collected (46 of 81; 56.8%) were classified as minor soybean pollen carriers, either because less than 25% of their corbicular or scopal pollen loads was soybean (13 of 60 female bees) or because soybean pollen was detected only on other parts of their bodies (14 of 60 female bees, 15 of 17 male bees, and 4 of 4 wasps).
Discussion
Although the first North American study on wild bee communities within soybean crops dates back several decades (Rust et al. Reference Rust, Mason and Erickson1980), the integration of pollinator management into soybean production has gained attention only recently (Garibaldi et al. Reference Garibaldi, Schulte, Nabaes Jodar, Gomez Carella and Kremen2021). Indeed, few studies have identified the wild insect visitors of soybean flowers, and even fewer have examined those that carry soybean pollen. To our knowledge, this study is the first to address this knowledge gap in Canada.
Given the considerable sampling effort in this study, the relatively low number of bees and wasps collected could indicate that soybean is less attractive to wild pollinators than surrounding wildflowers or that pollinator abundances are generally low in soybean-dominated landscapes. Because we did not sample adjacent field margins concurrently, we cannot disentangle these two possibilities. Regardless, a crop’s low attractiveness to pollinators does not necessarily reflect the extent to which it benefits from their services, particularly in partially self-pollinating crops (Esquivel et al. Reference Esquivel, Parys and Brewer2021). To clarify both the attractiveness of soybean to pollinators and the potential contribution of pollinators to soybean production in eastern Canada, future research should include simultaneous sampling of crop fields and adjacent habitats, as well as experimental assessments of pollinator contribution to yield.
In our study, bees were the most abundant visitors of soybean flowers, with 77 wild bee specimens collected across 19 soybean fields, compared to only four wasps. Although a few insects from other taxa were observed, they were not abundant. Factors such as small size (e.g., ants), foraging behaviour, or flight patterns (e.g., hover flies (Diptera: Syrphidae)) might have made these insects harder to detect. Future surveys should include other pollinator groups to better assess the diversity of soybean flower visitors.
The pollen load assessment revealed that all but one of the specimens collected (91.4%) carried soybean pollen, suggesting potential involvement in pollination. However, the presence of pollen on an insect’s body does not guarantee effective pollen transfer (Gill and O’Neal Reference Gill and O’Neal2015), and further research is needed to assess pollination efficacy.
The bee assemblages identified were broadly consistent with those reported from surveys using passive sampling in soybean fields across eastern North America (Rust et al. Reference Rust, Mason and Erickson1980; Cunningham-Minnick et al. Reference Cunningham-Minnick, Peters and Crist2019; Levenson et al. Reference Levenson, Sharp and Tarpy2022; Torrez et al. Reference Torrez, Beauzay, St. Clair and Knodel2022; Gaudreault et al. Reference Gaudreault, Naujokaititis-Lewis, Lapen and Sargent2023). In particular, bumble bees, Bombus griseocollis (De Geer), B. impatiens, and B. vagans Smith, sweat bees, Augochlorella aurata, Halictus ligatus Say, and H. rubicundus (Christ), multiple species of Lasioglossum (Dialictus), and the small carpenter bee Ceratina calcarata Robertson (Hymenoptera: Apidae) were all found in high abundance and across multiple studies and were also observed carrying soybean pollen in the present study, supporting the existence of a soybean-associated bee community (Gill and O’Neal Reference Gill and O’Neal2015; Wheelock et al. Reference Wheelock, Rey and O’Neal2016; Cunningham-Minnick et al. Reference Cunningham-Minnick, Peters and Crist2019). Broader geographic and temporal sampling of soybean fields would likely reveal greater diversity.
Previous studies suggest that increases in soybean yields attributed to insect pollination likely stem from the complementary effects of a diverse pollinator assemblage rather than the activity of any single species (Cunningham-Minnick et al. Reference Cunningham-Minnick, Peters and Crist2019; Levenson et al. Reference Levenson, Sharp and Tarpy2022). Nonetheless, the most important pollinators are likely those most frequently observed foraging on soybean flowers. In the present study, bumble bees were the most common flower visitors, followed by various species of ground-nesting bees, indicating that these two groups likely play a key role in soybean pollination. Although nectar is the primary soybean floral reward for bees (Severson and Erickson Reference Severson and Erickson1984), our pollen load analyses show that certain species, such as A. wilkella, B. impatiens, B. rufocinctus, B. sandersoni, B. ternarius, and L. admirandum, collect substantial amounts of soybean pollen even in the presence of nearby wildflowers. In each of these species, at least one sampled individual carried pollen loads composed of more than 80% soybean pollen. However, larger sample sizes and sampling across a broader array of habitats are needed to determine whether some species exhibit a particular attraction to this crop.
Notably, 22% of bee specimens were males, most of which carried soybean pollen on their bodies, despite not collecting pollen actively. Although male bees can be effective pollinators of certain crops (Cane et al. Reference Cane, Sampson and Miller2011), their contribution to soybean yields remains unclear. Similarly, the few wasps and the brood-parasitic (“cuckoo”) bee, Sphecodes sp. (Hymenoptera: Halictidae), that were collected could provide incidental pollination through nectar foraging. Although wasps are minor pollen carriers, their presence highlights the need for further investigation into the role of non-bee insects in soybean pollination.
Given the reported benefits of insect-mediated pollination for soybean yield in other regions (e.g., de O. Milfont et al. Reference de O. Milfont, Rocha, Lima and Freitas2013; Garibaldi et al. Reference Garibaldi, Schulte, Nabaes Jodar, Gomez Carella and Kremen2021), the present findings suggest that conservation efforts that support bumble bees and ground-nesting bees are likely to be particularly relevant for growers in Canada. Many ground-nesting bees nest not only in field margins but also within arable fields, and adding soil-based nesting resources within soybean fields has been shown to enhance both bee abundance and crop yield (Cunningham-Minnick et al. Reference Cunningham-Minnick, Peters and Crist2019; Tschanz et al. Reference Tschanz, Vogel, Walter, Keller and Albrecht2023). Other strategies to maintain or enhance pollinator habitat could include providing continuous forage throughout the season (Pywell et al. Reference Pywell, Warman, Carvell, Sparks, Dicks and Bennett2005; Purvis et al. Reference Purvis, Meehan and Lindo2020) and minimising pesticide use – especially pesticides that are persistent in soil and might affect nesting or overwintering bees (Willis Chan et al. Reference Willis Chan, Prosser, Rodríguez-Gil and Raine2019; Willis Chan and Raine Reference Willis Chan and Raine2021; Rondeau et al. Reference Rondeau, Baert, McArt and Raine2022a, Reference Rondeau, Willis Chan and Pindar2022b; Rondeau Reference Rondeau2024; Rondeau and Raine Reference Rondeau and Raine2024a, Reference Rondeau and Raine2024b, Reference Rondeau and Raine2024c). Although additional research is needed to confirm the benefits of pollinator management to soybean production in eastern Canada specifically, promoting pollinator-friendly practices might contribute to broader agroecosystem health.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.4039/tce.2025.10034.
Acknowledgements
The authors thank the field crop growers who granted access to their fields. The authors also thank Christine Rieux, Claire Cossette, and Maria José Maezo for assistance with site selection, Amélie Gervais for help with bee identification, and Sébastien Rousseau for support creating the study site map. They appreciate the constructive feedback provided by the editor and two anonymous reviewers to earlier versions of this manuscript. This project was funded by a research grant from the Weston Family Foundation. S.R. was supported by postdoctoral fellowships from the Natural Sciences and Engineering Research Council of Canada (NSERC; PDF-5783-2023), the L’Oréal–UNESCO for Women in Science Program, the Fonds de recherche du Québec – Nature et technologies (2023–2024 – B3X-332009), and the Weston Family Foundation. L.F.B. was supported by an Undergraduate Student Research Award (USRA) from NSERC. N.E.R. is supported as the Rebanks Family Chair in Pollinator Conservation by the Weston Family Foundation.
Competing interests
The authors declare that they have no conflicts of interest to disclose.
Author contributions
S.R.: conceptualisation, methodology, investigation, writing – original draft, writing – review and editing, visualization, supervision, project administration, and funding acquisition. L.F.B.: methodology, investigation, visualisation, and writing – review and editing. N.E.R.: conceptualisation, methodology, supervision, funding acquisition, and writing – review and editing. J.R.K.F.: conceptualisation, methodology, validation, resources, supervision, funding acquisition, and writing – review and editing.
Data availability
The data that support the findings of this study are available in Supplementary material, Dataset S1.





 Open access
Open access