


Background
Delia radicum was first recorded in Canada in 1855 (Fletcher Reference Fletcher1886). Gibson and Treherne (Reference Gibson and Treherne1916) described D. radicum as causing year-to-year losses from Nova Scotia to British Columbia. The same authors also described control measures explored since 1910, which included carbolic acid, a kerosene emulsion, heavy manuring, glue and bran, and a pyrethrum insect powder. They concluded that none of these methods showed consistent benefit, with none being recommended as a control option.
The first published record of D. radicum in Manitoba was reported in 1958 (Allen Reference Allen1964), with Turnock et al. (Reference Turnock, Boivin and Whistlecraft1995) reporting records of D. radicum along the Red and Assiniboine rivers around 1930. Wilson (Reference Wilson1913) confirmed the presence of D. radicum in British Columbia in 1912. The origin of D. radicum in North America was a source of speculation prior to the advent of DNA analysis. Based on the historical evidence, D. radicum was thought to have been introduced via a single introduction to North America along the northeastern coast. Biron et al. (Reference Biron, Landry, Nénon, Coderre and Boivin2000) confirmed the genetic similarity of D. radicum in North America with populations in northwestern Europe, supporting the single introduction theory.
Despite its widespread presence in most provinces, D. radicum was noticeably absent from certain regions of Canadian agriculture at certain times in history. For example, major pests affecting field crops and gardens in Saskatchewan did not include D. radicum from 1922 to 1927 (King Reference King1928). While Tolman et al. (Reference Tolman, McLeod and Harris1986) was reporting losses of 55–60% from D. radicum in southwestern Ontario rutabaga, Turnock et al. (Reference Turnock, Timlick, Galka and Palaniswamy1992) noted that Manitoba experienced no yield losses in 1985–1988. By 2004, the picture had changed radically, with Manitoba and Saskatchewan reporting 96% of canola fields with damage and Alberta reporting 99% of surveyed fields with D. radicum damage (Soroka et al. Reference Soroka, Dosdall, Olfert and Seidle2004). In Alberta, D. radicum was recorded from canola fields in central and north–central Alberta throughout the 1980s (Liu and Butts Reference Liu and Butts1982; Liu Reference Liu1984; Griffiths Reference Griffiths1985) but not in the arid southeast. Although the Peace River region of Alberta was not included in these surveys (Liu and Butts Reference Liu and Butts1982; Liu Reference Liu1984; Griffiths Reference Griffiths1985), Broatch (Reference Broatch1993) later confirmed the presence of D. radicum throughout that region in the early 1990s. In 2005, the Crop Profile for Rutabaga described D. radicum damage in all rutabaga-growing regions as a “widespread yearly occurrence with high pest pressure” (Agriculture and Agri-Food Canada 2005). The occurrence and status of D. radicum have remained the same across the rutabaga-growing regions, as reported in subsequent updates of the Crop Profile (Agriculture and Agri-Food Canada 2021) until 2021, with occasional fluctuations between high to moderate pressure in cabbage and broccoli production in British Columbia and Quebec (Agriculture and Agri-Food Canada 2022).
Identification and life history
Difficulties accurately identifying D. radicum have exacerbated the challenges associated with managing this pest. Until recently, the most used key for Delia spp. identification was that developed by Brooks (Reference Brooks1951), which described the different life stages of Delia spp. pests in Canadian crucifer crops. Brook’s (Reference Brooks1951) key is comprehensive but poorly illustrated and names taxa using now-obsolete nomenclature. More recently, Savage et al. (Reference Savage, Fortier, Fournier and Bellavance2016) published a revision of the Delia genus, focusing on species present in Canada: D. antiqua (Meigen), D. floralis (Fallén), D. florilega (Zetterstedt), D. planipalpis (Stein), D. platura (Meigen), and D. radicum. Savage et al. (Reference Savage, Fortier, Fournier and Bellavance2016) provide keys (in English and French) to the adult, larval, pupal, and egg stages with high-resolution images to assist users. Morphological keys can be augmented by the use of DNA barcoding, which is especially useful for distinguishing between the larvae of the common Delia spp. (Savage et al. Reference Savage, Fortier, Fournier and Bellavance2016). Additionally, Savage et al. (Reference Savage, Fortier, Fournier and Bellavance2016) conducted observational studies to understand the association of the larval stages of Delia spp. with different host plants, revealing that D. radicum larvae were associated only with Brassica spp. host plants and not with Allium spp. host plants. The key and accompanying studies published by Savage et al. (Reference Savage, Fortier, Fournier and Bellavance2016) provide useful and practical tools for entomologists working with Delia spp., but accurately and rapidly identifying Delia spp. in the lab and field remains challenging. As a result of these challenges, molecular methods were developed (Van der Heyden et al. Reference Van der Heyden, Fortier and Savage2020; Mlynarek and Sim Reference Mlynarek and Sim2021). The high-resolution melting–polymerase chain reaction assay (HRM–PCR) was developed to discriminate between D. florilega and the two genetic lines of D. platura, whereas the polymerase chain reaction–restriction fragment length polymorphism (PCR–RFLP) method successfully distinguished between five Delia spp. and between the genetic lines of D. platura. Such tools offer an economical option to sequencing that can confirm species identification for research purposes. Development of this technology into a commercially available and user-friendly application would provide growers and pest management practitioners with the ability to accurately identify D. radicum.
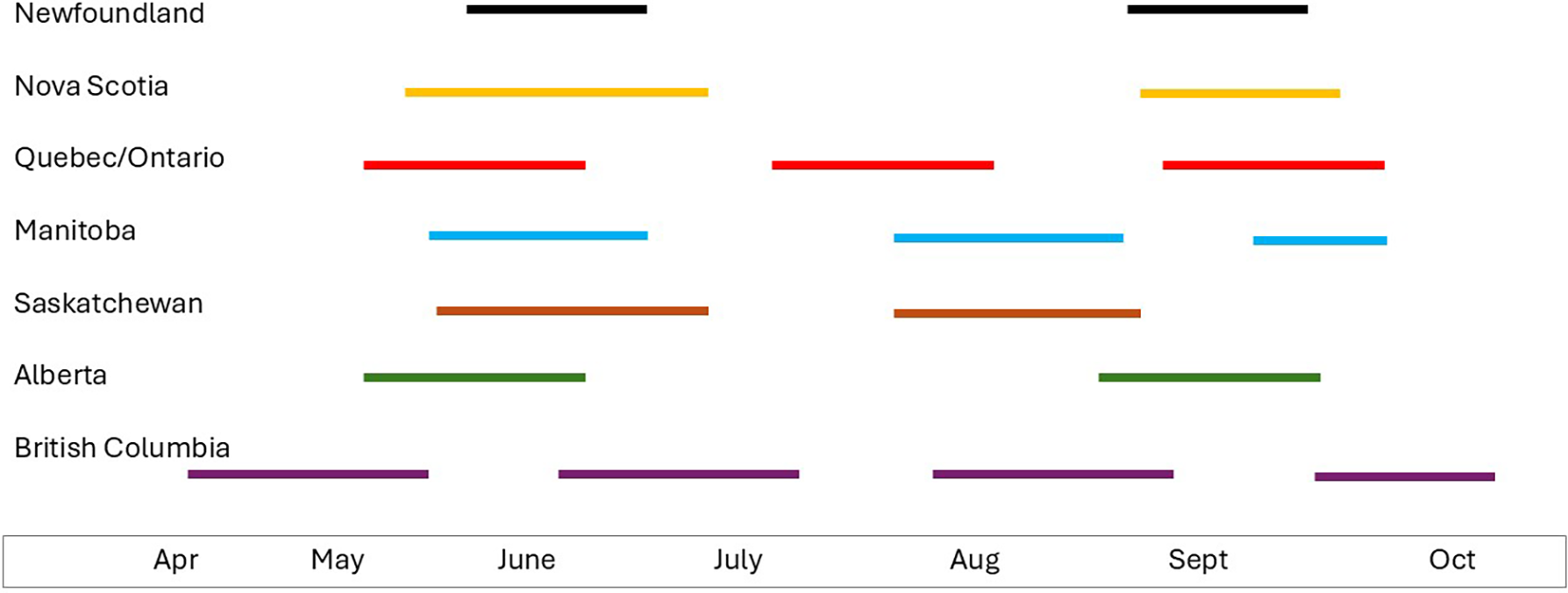
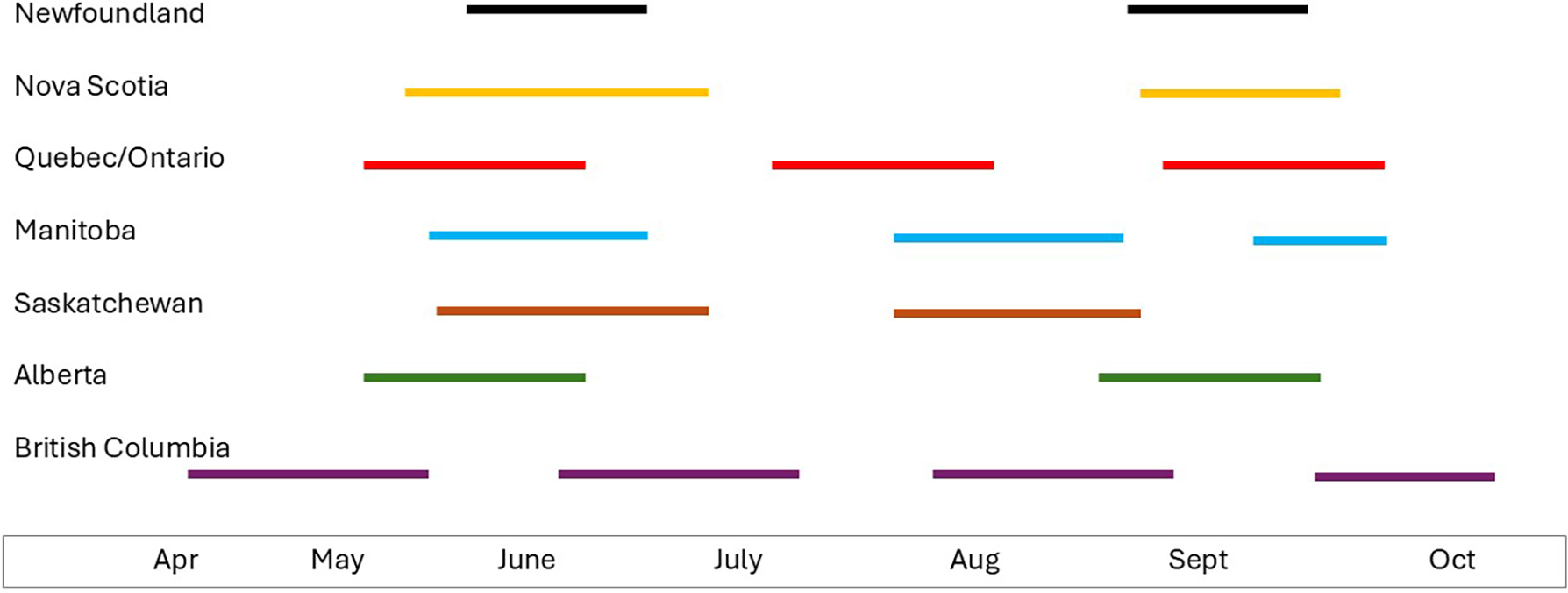
The life history of D. radicum was reviewed by Holliday et al. (Reference Holliday, Andreassen, Dixon, Kuhlmann, Mason and Gillespie2013) and is summarised here. Delia radicum overwinters as pupae located in the soil, with pupal development being completed in the spring when thermal requirements are met. Adult flies emerge, mate, and lay eggs on suitable host plants. Oviposition requires sequential landings on an appropriate host (Kostal and Finch Reference Kostal and Finch1994). Eggs hatch in 3–10 days, and larvae develop through three instars in 3–4 weeks (Holliday et al. Reference Holliday, Andreassen, Dixon, Kuhlmann, Mason and Gillespie2013). The larval stage is the life stage of concern for crops as they feed upon the roots of the host. In young plants, feeding can result in plant death, whereas in older plants, damage will depend on the crop type and severity of attack. In flowering brassicas, for example, broccoli or cauliflower, root damage weakens the plant, resulting in smaller heads, whereas in root crops, for example, rutabaga, feeding damage can render the root unmarketable. Across Canada, the number of D. radicum flights each year can vary from two, in Newfoundland, to four, in British Columbia (Bracken Reference Bracken1988; Coady Reference Coady1999; Holliday et al. Reference Holliday, Andreassen, Dixon, Kuhlmann, Mason and Gillespie2013; Fig. 1). In Ontario, three generations are commonly noted, but a partial fourth generation was observed in the 1970s (Nair and McEwen Reference Nair and McEwen1975). To further complicate the management situation, Biron et al. (Reference Biron, Langlet, Boivin and Brunei1998, Reference Biron, Coderre, Boivin, Brunel and Nénon2002) confirmed the presence of early- and late-emerging phenotypes resulting in extended emergence patterns and potential overlap of generations. The diversity in D. radicum life history across Canada highlights another challenge related to its management and the importance of researching and identifying different approaches to integrated pest management across Canada. Multiple generations over the growing season create additional challenges for pesticide applications. Application timing may conflict with need to access the crop for horticultural practices such as hand-weeding and, more importantly, may result in an increased number of applications during the growing season. Pesticide reviews commonly seek to reduce environmental exposure through a reduction in the number of sprays, which is difficult for areas experiencing multiple generations of D. radicum. Pesticide labels do not currently acknowledge the difference in the number of generations across Canada, and therefore, this is not considered in the products’ recommended use of the product; that is, there are no early-season versus later-season recommendations.
Schematic showing number and duration of flights of Delia radicum across Canada.
Monitoring methods
Monitoring in Canada for D. radicum to identify adult emergence and assess population pressure has focused on two main strategies: sticky traps and water pan traps. Yellow sticky traps set up in a grid around a rutabaga planting in British Columbia captured more D. radicum on the perimeter of the field, but correlation between trap capture and number of eggs or observed larval feeding damage could not be determined (Blackshaw et al. Reference Blackshaw, Vernon and Prasad2012). Water pan traps can be effective to determine adult emergence, but captures showed no correlation with numbers of eggs in Manitoba brassica crops (Bracken Reference Bracken1988). Assessing D. radicum population levels at harvest provides a means to evaluate populations over time. Damage at harvest is often assessed using a scale of 0 to 4 for brassica vegetables (King and Forbes Reference King and Forbes1954) or a modified scale of 0 to 5 for canola (Dosdall et al. Reference Dosdall, Herbut and Cowle1994).
Another option is to monitor oviposition. Dixon et al. (Reference Dixon, West, McRae and Spaner2002) investigated the use of felt traps to detect oviposition by D. radicum in rutabaga and cabbage. Traps were adequate for monitoring later-season oviposition, but D. radicum females preferred ovipositing on the soil in the early season. Until 2022, crop consultants in Saskatchewan monitored irrigated brassica vegetable crops for D. radicum eggs to time the application of chlorpyrifos as a soil drench, as per Ontario Ministry of Agriculture and Food and Rural Affairs recommendation (Government of Ontario 2025). Although labour intensive, this practice did help to ensure that insecticides were applied at the correct time to maximise efficacy. In British Columbia, a combination of monitoring for adults using yellow sticky cards and monitoring for eggs was used during 2011–2013 in rutabaga fields. This monitoring method was used to help growers better time their applications of chlorpyrifos and cypermethrin (product name RipcordTM 400 EC) against the hatching larvae; the latter is registered for emergency use only in British Columbia (van Herk et al. Reference van Herk, Vernon, Waterer, Tolman, Lafontaine and Prasad2017). Based on the results of these monitoring efforts, rutabaga growers in British Columbia essentially follow a calendar spray schedule because D. radicum activity (both adults and eggs) is detected throughout the growing season. More recently, Prasad and van Herk have conducted extensive egg sampling in rutabaga fields over a four-year period (2019–2022). Preliminary data from their studies found egg sampling in fields to correlate with damage observed at harvest; areas with high egg counts match areas where high damage is observed at harvest (R. Prasad, unpublished data). Our literature search did not identify any economic thresholds for any crop.
Management methods
Chemical
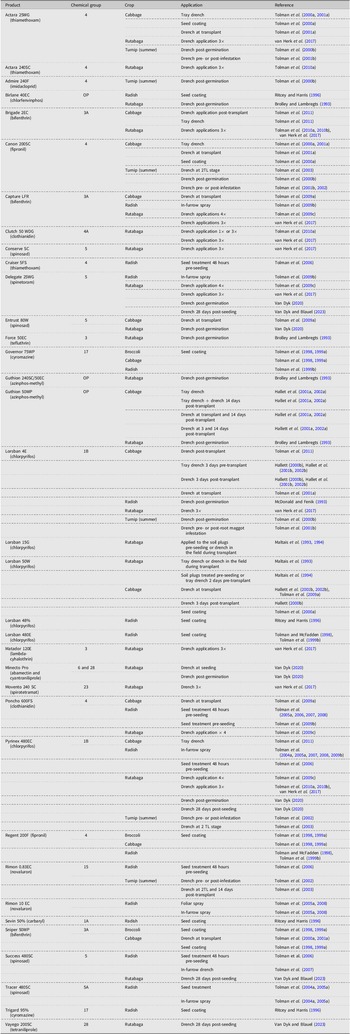
Insecticides are still the most commonly used tool for D. radicum management and have been widely researched. Insecticides against D. radicum target the root-feeding larval stage over the highly mobile adult stage. Insecticide products in several chemistry classes and groups have been investigated, including organophosphates, pyrethroids, neonicotinoids, anthranilic diamides, and biopesticides, applied either singly (Tables 1 and 2) or in combination (Table 3). Application methods targeting the larval stage have ranged from seed treatments (i.e., systemic or nonsystemic products applied directly to the seed prior to sowing), to in-furrow treatments at seeding, to treatment of seedling plugs with systemic insecticides prior to transplant, to the application of foliar sprays at various times during the growing season. The latter treatment is more accurately called a “soil drench”, whereby the product is applied with sufficient water volume to carry the active ingredient into the root zone. Seed treatments targeting D. radicum larvae require that the product have systemic activity to facilitate the transfer of the active ingredient into the developing root or that the product is washed off the seed into the soil and onto the developing roots, where it can be taken up by the plant or encountered by the larvae, that is, through ingestion or contact (Altmann Reference Altmann2003).
Research conducted in Canada with pesticide products (active ingredient) no longer available for control of Delia radicum, highlighting chemistry, crop, and application method.
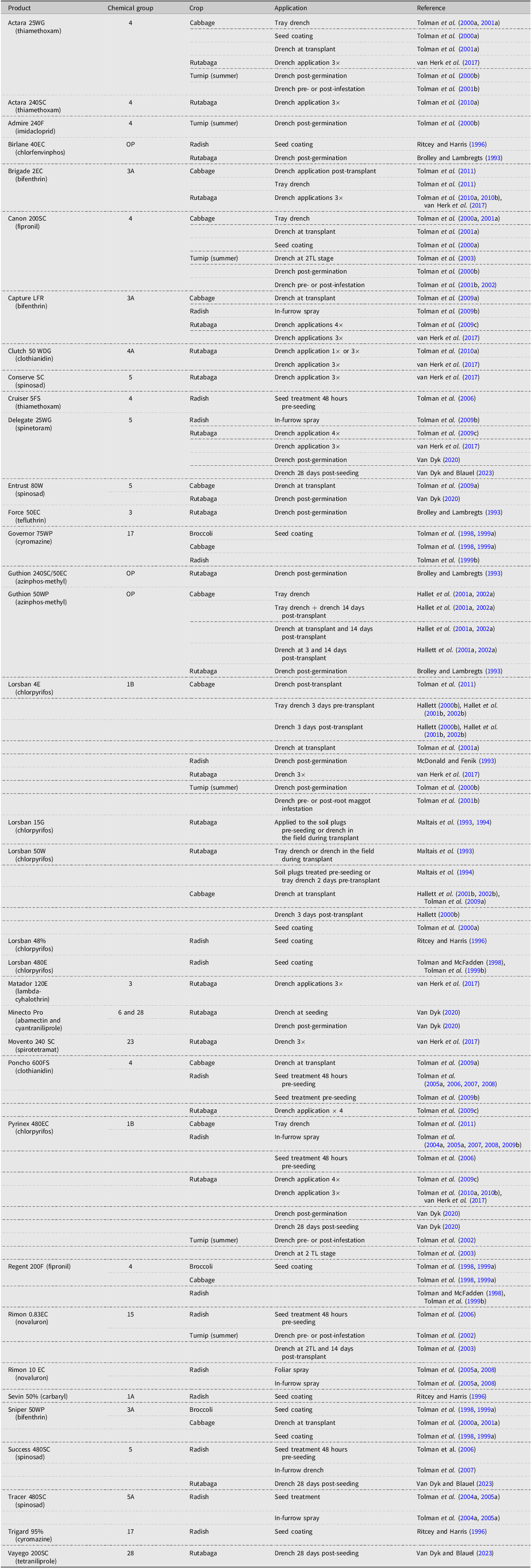
Note: 2TL refers to the second true leaf stage of plant development.
Research conducted between 1992 and 2022 using currently available products for control of Delia radicum in Canada, highlighting chemistry, crop, and application method.
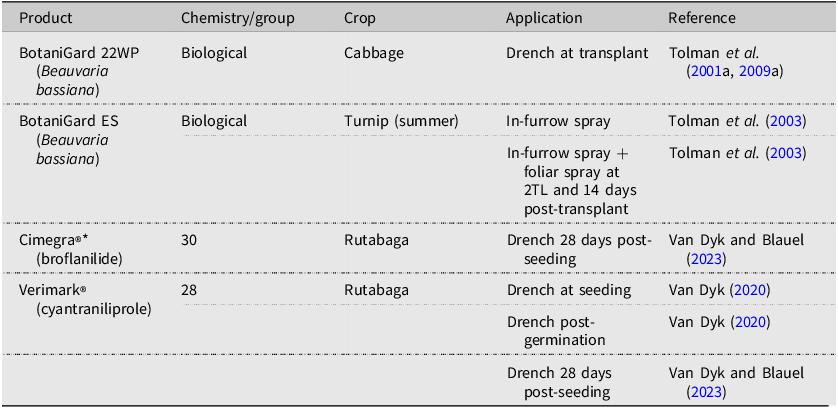
Note: 2TL refers to the second true leaf stage of plant development.
*Emergency registration
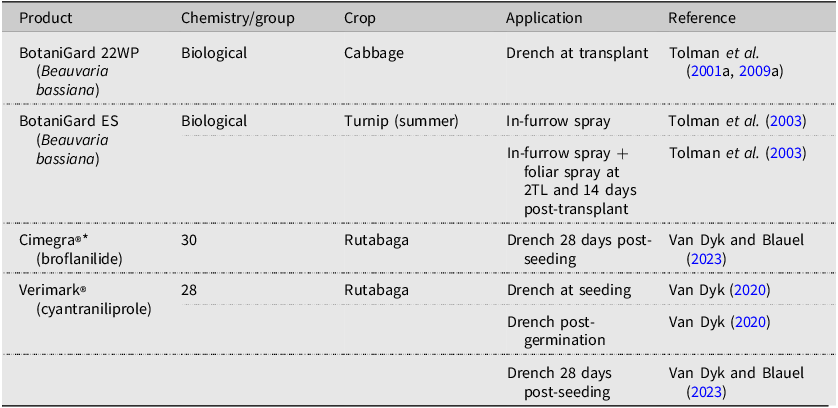
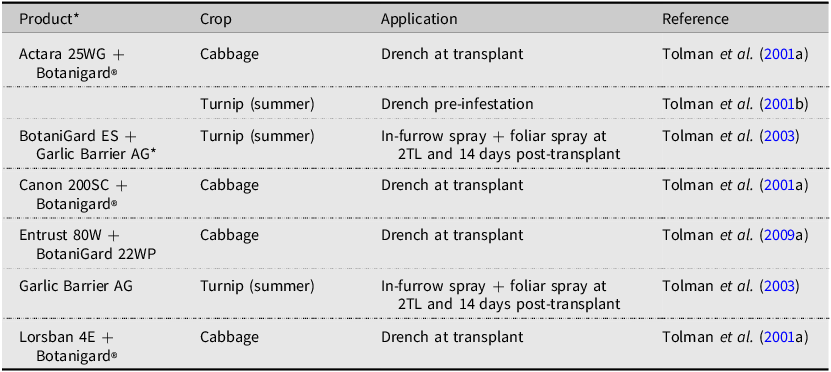
Botanical products and combinations tested for efficacy against Delia radicum in Canada.
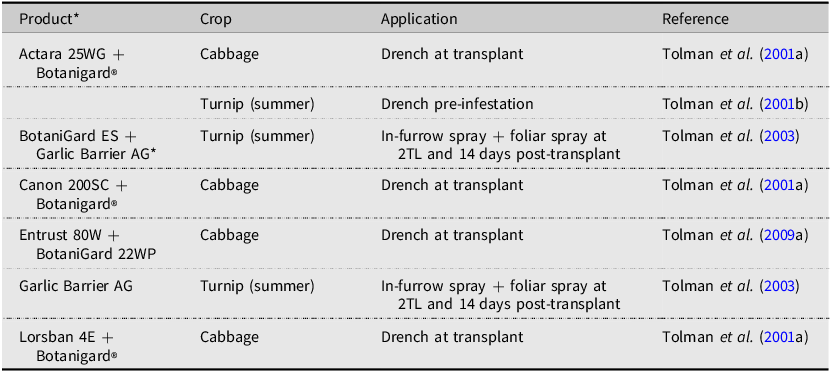
Note: 2TL refers to the second true leaf stage of plant development.
*Garlic Barrier AG: 500 mL reverse osmosis water, 10.0 mL Garlic Barrier AG insect repellent, 10.0 mL canola oil, and 8.0 mL Palmolive® hand soap
One challenge with the use of insecticides to manage D. radicum has been the insect’s development of resistance to certain active ingredients, resulting in the need to switch to different chemistries. Delia radicum populations resistant to cyclodiene organochlorine insecticides, for example, were reported in the late 1950s and early 1960s in British Columbia and western Washington (United States of America; Finlayson Reference Finlayson1962; Howitt and Cole Reference Howitt and Cole1962), Newfoundland (Morris Reference Morris1963), Quebec (Harris et al. Reference Harris, Manson and Mazurek1962), Prince Edward Island (Read Reference Read1964), and Nova Scotia (Fox Reference Fox1965). Research in the 1960s with organophosphate and carbamate groups found organophosphates to show promise (Finlayson et al. Reference Finlayson, Noble and Fulton1967) and provide an alternative where resistance was developing. In the 1980s, the efficacy of carbofuran (a carbamate) against D. radicum was reported to be declining in Newfoundland and British Columbia due to “antagonistic” soils (Mackenzie et al. Reference Mackenzie, Vernon and Szeto1987). Harris et al. (Reference Harris, Chapman, Morris and Stevenson1988) determined that the soil microbial community was actively degrading carbofuran, effectively removing the product from the soil. Problems with other organophosphates (e.g., chlorfenviphos, diazinon) included phytotoxicity (reduced germination, stunting) and suspected resistance (e.g., diazinon), supporting increased use of chlorpyrifos (MacKenzie et al. Reference Mackenzie, Vernon and Szeto1987). However, trials in Ontario during 2008 showed D. radicum to exhibit some tolerance to chlorpyrifos (e.g., Pyrinex® 480EC; Tolman et al. Reference Tolman, Minto, Steffler and McPherson2008). Observations from different Canadian growing regions prior to 2013 indicated that D. radicum populations were not being controlled with chlorpyrifos (e.g., LorsbanTM, Pyrinex®) to desirable levels. A survey of D. radicum populations collected from British Columbia in 2013, Newfoundland in 2015, and southern Ontario in 2016 was conducted to assess chlorpyrifos susceptibility relative to a laboratory insecticide–susceptible D. radicum strain (Scott et al. Reference Scott, Baptista, Countryman, Dixon, Hann and Krolikowski2018). The survey of these 12 D. radicum populations indicated reduced susceptibility to chlorpyrifos in 75% (9 of 12). Reduced susceptibility and resistance were related to the intensive use of LorsbanTM and other chlorpyrifos-containing products (Scott et al. Reference Scott, Baptista, Countryman, Dixon, Hann and Krolikowski2018). Chlorpyrifos showed strong efficacy against D. radicum and was used for over 30 years until it was deregistered in Canada in 2023 (Health Canada 2023). Currently, registered pesticide options for use against D. radicum are few (Table 2) but include cyantraniliprole (e.g., Verimark®) and Spinosad (e.g., Entrust SC, SuccessTM), applied either as a tray drench to seedlings prior to transplanting (e.g., broccoli, cabbage) or as a single, direct in-furrow spray (cyantraniliprole) or several broadcast granular applications at the base of plants (Spinosad). Recently, Emergency Use registrations were granted by the Pest Management Regulatory Agency (PMRA) for SuccessTM (PCP 26835) and Cimegra® (PCP 33666), active ingredient broflanilide, in rutabaga. Registrations for the adult (fly) stage of D. radicum include several formulations of cypermethrin for rutabaga and turnips (in British Columbia only).
Other chemical applications, such as elemental sulfur and sulfate sulfur, and insect growth regulators have also been evaluated for efficacy against D. radicum. Despite application at high rates, for example, 112 kg ha−1, no substantial reduction in damage with sulfur was observed relative to controls (Dosdall et al. Reference Dosdall, Yang and Conway2002). Field studies by Howard (Reference Howard1977), Philip et al. (Reference Philip, Steiner and Howard1977), and Ritcey et al. (Reference Ritcey, McGraw and McEwen1977) all found chitin synthesis inhibitors to reduce crop damage, with similar positive results in greenhouse trials (Turnbull Reference Turnbull1982). In lab studies, Young et al. (Reference Young, Gordon and Cornect1987) found Dimilin® (active ingredient: diflubenzuron, a chitin biosynthesis inhibitor) to be as effective as the commercial insecticide, carbofuran, when comparing treated rutabaga slices for D. radicum damage. Despite the efficacy observed in lab studies, we could find no field studies evaluating Dimilin® in Canada. Pesticides continue to be used throughout Canada for the control of D. radicum in vegetable crops, but numerous alternative options have also been explored. There are currently no registered products for use in canola production to manage losses from D. radicum.
Physical and cultural practices
Row covers
Various types of physical barriers have been evaluated for the management of D. radicum and other insect pests under field conditions in Canada. In the mid-1980s, the arrival of the agricultural spunbonded polyester and polypropylene fabrics allowed for their use as row covers with a role in crop protection by excluding pest insects from the crop (Millar and Isman Reference Millar and Isman1988). Two types of floating row covers and an uncovered treatment were compared in trials for early- and mid-season radish production (Rekika et al. Reference Rekika, Stewart, Boivin and Jenni2008). The floating row covers completely excluded D. radicum compared to control plots.
Impediments to the use of row covers have been their relatively high cost in contrast to the use of pesticides, along with the amount of handling required, including for the initial placement, removal for weeding, fertilising, and harvesting, and the final storage or disposal of the covers (Boiteau and Vernon Reference Boiteau, Vernon, Vincent, Panneton and Fleurat-Lessard2001). More recent use of row covers by organic growers to protect crops from insect pests has demonstrated that covers can work successfully against D. radicum. In addition, Dixon et al. (Reference Dixon, Fillmore, LeBlanc, Madore, Mellish and Owen2013) found that plots covered with Wondermesh® and ProtekNet® resulted in higher marketable yields of rutabaga than plots treated with chlorpyrifos insecticides.
Field trials using row covers conducted by Agriculture and Agri-Food Canada in 2014 and 2015 in British Columbia, Nova Scotia, and Newfoundland evaluated the efficacy of three mesh sizes (range: 0.60–1.33 mm) at protecting transplanted brassica seedlings from D. radicum, comparing these to both uncovered, untreated transplants and uncovered plants treated with chlorpyrifos drenches (the industry standard; Dixon and Anderson Reference Dixon and Anderson2016). All three mesh sizes significantly reduced D. radicum damage on broccoli and radish plants to a greater extent than conventional insecticide in both years. Broccoli grown under netting matured more quickly and had significantly higher marketable yields compared to uncovered plants in 2015. These studies corroborated earlier work with direct-seeded rutabaga in New Brunswick, Newfoundland, Nova Scotia, and Prince Edward Island in 2011 and 2012 (Owen Reference Owen2013). In those studies, row covers provided similar protection from D. radicum damage as chlorpyrifos did. The studies addressed several common concerns, including how to manage weeds (this varies with region and with weed species present), and the potential of reusing different row cover types for several years to justify the substantial initial expense. A larger-scale 12-ha (30-acre) demonstration trial in British Columbia occurred in 2019 and showed a dramatic reduction in D. radicum–related culls in turnip grown under Wondermesh® following one insecticide application prior to installation of the mesh. As part of this demonstration, rutabaga pieces and yellow sticky cards were placed along the field margin to confirm season-long D. radicum activity in the field. Wondermesh® has now been adopted in large-scale turnip and daikon radish production in British Columbia. Rutabaga growers, however, remain hesitant to use mesh covers due to anecdotal reports of misshapen rutabaga under the mesh, possibly a result of trapped D. radicum under the netting. In 2021 and 2022, seven commercial-scale demonstration trials were conducted in Quebec using a HIWERTM floating row cover system from Dubois Agrinovation (Saint-Rémi, Quebec) in Chinese cabbage, cabbage, rutabaga, and turnip. Similar to previous trials, results showed comparable yields between conventional and netting strategies; however, there was a large economic cost associated with the installation and removal of the netting (Kora and Bélec Reference Kora and Bélec2023).
Exclusion fences
Exclusion fencing capitalises on insect behaviour and provides an option for D. radicum control. Early studies found that many vegetable insect pests were trapped in high numbers within 30 cm of the ground or crop canopy (Vernon Reference Vernon1979). When these pest insects encounter obstacles, such as a screen interception trap, they tend to move upwards to become trapped at the apex of the structure. Fences constructed of fine netting material with a screen overhang at the top were found to impede the movement of insect pests into the host crop (Boiteau and Vernon Reference Boiteau, Vernon, Vincent, Panneton and Fleurat-Lessard2001). Exclusion fences that are at least 90 cm high have been shown to reduce the dispersal of Delia flies into small plantings of rutabaga and other brassica vegetables (Vernon and Mackenzie Reference Vernon and Mackenzie1998). The benefit of exclusion fences is that they permit mechanical or chemical weeding without the need for removal (Päts and Vernon Reference Päts and Vernon1999). In addition to fence height, a screen collection overhang at the top can improve exclusion. Bomford et al. (Reference Bomford, Vernon and Päts2000) found a significant reduction in the number of D. radicum captured on yellow sticky traps and percentage of radishes damaged when plots were fenced with a 25-cm overhang compared to unfenced control plots. Despite these reductions, exclusion barriers were not adopted by the agricultural community at the time, owing to the cost of fencing materials, labour required to install and maintain the fences, and reduced efficacy of the strategy in large fields.
Subsequent Agriculture and Agri-Food Canada research on exclusion fences in 2009 resulted in an affordable, commercially available design (Telstar Eco Fence; Telstar Window Service, Agassiz, British Columbia), that was field-tested in commercial rutabaga fields in British Columbia, Saskatchewan, and Newfoundland in 2010 and 2011 (van Herk Reference van Herk2016). This fence was similar to the earlier prototypes but used black, 1-mm-mesh nylon window screen, is slightly higher (1.3 m), and has an inverted (45° angle), outward-facing 0.3-m overhang to intercept flies entering the field. Individual 30-m sections can be installed or dismantled by a single person in less than 50 minutes, are easily rolled up for storage, and, under optimal conditions, can be reused for more than five years (Vernon et al. Reference Vernon, Blackshaw and Prasad2011). Weekly monitoring of D. radicum adults, eggs, and damage in commercial plantings of rutabaga enclosed with this fence design in British Columbia in 2009 showed a substantial reduction in the number of female flies entering fields (by up to 96%). In addition, most female flies, eggs, and damage observed in the enclosed field were largely restricted to the perimeter adjacent to the fence (Vernon et al. Reference Vernon, Blackshaw and Prasad2011). This work suggests that exclusion fences can effectively reduce the number of female flies reaching crops and can be used to manage D. radicum in commercially high-value commodities such as rutabaga. Similar reductions of other co-occurring, low-flying anthomyiids (i.e., Delia platura) have also been documented (Blackshaw et al. Reference Blackshaw, Vernon and Prasad2012).
Several important factors can reduce the efficacy of exclusion fences, including the presence of shrubs or other vegetation immediately outside the fence, in-field cruciferous weeds (alternate hosts and potential early-season sources of infestation from within the field), and optimal timing of fence installation (before or shortly after planting). For example, Blackshaw et al. (Reference Blackshaw, Vernon and Prasad2012) compared damage from D. radicum inside the fence where the area outside the fence was managed, or not, and noted that regular mowing of the grass and weeds outside the fence reduced D. radicum damage inside the fence. In summary, it appears that exclusion fences work best and are most cost-effective when pest pressure is low to moderate, when the previous crop was not cruciferous, when fields are more or less square (to reduce the perimeter–area ratio) and larger than 1 ha. The larger size ensures that, should a pesticide spray along the perimeter of the field just inside the fenced area be needed, this will be increasingly cost-effective; that is, the proportion of the field requiring spray would decrease (Vernon Reference Vernon2012; van Herk Reference van Herk2016). For example, a 10-m band sprayed along the outer perimeter of a (square) 1-ha field requires 40% of the field to be sprayed. This proportion decreases to 20% and 10% for square fields that are 4 and 16 ha, respectively.
Agronomic practices
Seeding density, seeding timing, and crop rotation have been explored for management of D. radicum. Delayed seeding of canola in Alberta to later in May reduced damage from D. radicum but also reduced yield, likely due to the shorter growing season (Dosdall et al. Reference Dosdall, Herbut, Cowle and Micklich1996). For a practice such as delayed seeding to be effective, it must be coupled with a short-season variety to avoid reduced yields owing to decreasing daylength. The development of short-season varieties in crops such as canola may allow delayed planting to become a viable control strategy to reduce D. radicum impact. Seeding in the fall or earlier in spring did not reduce damage from D. radicum, likely owing to its extended emergence spanning the early crop stages, which are most vulnerable to attack (Dosdall et al. Reference Dosdall, Clayton, Harker, O’Donovan and Stevenson2006). Increasing the seeding rate in both studies resulted in reduced basal stem diameter of the crop. Crop rotation was studied in western Canada as a potential strategy to reduce D. radicum in canola. The highest canola yields were obtained when canola was rotated with pea and barley in a three-year rotation (Dosdall et al. Reference Dosdall, Harker, O’Donovan, Blackshaw, Kutcher, Gan and Johnson2012). Although three-year or four-year rotations are not widely practised, alternation of canola with wheat or a pulse crop is now more common in canola production.
Behavioural modification
Efforts to alter D. radicum behaviour have focused on the females to manipulate where oviposition occurs. Attracting D. radicum females to a secondary crop and away from the main crop or deterring oviposition have received some attention in Canada. Hallett (Reference Hallett2000a) tested sinapic acid and monoterpene combinations to repel D. radicum in cabbage and rutabaga. Sinapic acid combined with a three- or six-component monoterpene blend showed the lowest damage and highest yields in rutabaga but not cabbage. An additional study of sinapic acid and monoterpene combinations found less damage with a six-component blend in both cabbage and rutabaga but no increase in yield (Hallett et al. Reference Hallett, Sopher and Heal2001c).
Another form of behavioural manipulation, explored by Coady (Reference Coady1999), Dixon et al. (Reference Dixon, Coady, Larson and Spaner2004), and Parsons et al. (Reference Parsons, Dixon and Colbo2007), is intercropping, which could interfere with the pre-oviposition flights of female D. radicum described by Kostal and Finch (Reference Kostal and Finch1994). Undersowing rutabaga with a nonhost crop such as white clover, Trifolium repens Linnaeus (Fabaceae), resulted in the undersown plots and control plots experiencing similar levels of D. radicum damage and lower yields in the undersown plots (Coady Reference Coady1999; Dixon et al. Reference Dixon, Coady, Larson and Spaner2004). It was concluded that competition between rutabaga and the white clover was the primary reason for the observed reduction in rutabaga yield, not D. radicum damage (Dixon et al. Reference Dixon, Coady, Larson and Spaner2004). When intercropped with lettuce, D. radicum egg numbers were reduced, and cauliflower yield and quality were retained (Parsons et al. Reference Parsons, Dixon and Colbo2005, Reference Parsons, Dixon and Colbo2007).
For intercropping to be successful as a strategy, the choice of nonhost, or more-preferred host (for trap cropping), is key. Population pressure, likewise, needs to be such that D. radicum has adequate substrate for oviposition (in the case of trap cropping) or can locate alternate hosts outside the cropped area. One factor that may impact population pressure in any given year may be proximity to other brassica hosts (or crops). Despite anecdotal evidence suggesting that planting brassica vegetable crops near a canola field results in higher damage, we could find no studies in Canada on this topic. We likewise identified no preference studies conducted in an intercropping or trap-cropping context in the Canadian D. radicum literature.
Host plant resistance
As it is challenging to manage D. radicum with chemicals, there has been significant interest in and research investment made to identify germplasm and to develop resistant or tolerant varieties of brassica oilseeds and vegetables, both in Canada and internationally. Rutabaga lines bred for resistance to D. radicum in the early 1960s had consistently damage-free roots compared to susceptible commercial varieties, owing to differential egg-laying (Pond et al. Reference Pond, Dionne, White and Moore1962). Woods and Dosdall (Reference Woods and Dosdall1997) created 200 breeding lines from crosses of rutabaga and canola with backcrossing; some showed promise when compared against known susceptible varieties, for example, AC Sunshine. Tolman et al. (Reference Tolman, Kott, Mayo and Murray2004b) evaluated canola, mustard, and rutabaga breeding lines in mineral soil in Ontario. Observed damage was less than 10% in the canola and mustard lines, while the rutabaga lines showed severe damage.
Field, laboratory, and greenhouse studies have identified several wild relatives of cultivated brassicas that are resistant via antibiosis (insects fail to survive on host), antixenosis (insects are not attracted to the host), or both, or are tolerant to D. radicum. These include Sinapis alba Linnaeus (Dosdall et al. Reference Dosdall, Good, Keddie, Ekuere and Stringam2000; Jyoti et al. Reference Jyoti, Shelton and Earle2001), Brassica fruticulosa Cirillo, Hirschfeldia incana (Linnaeus) Lagrèze-Fossat (formerly Brassica incana), Brassica villosa Bivona-Bernardi, Brassica macrocarpa Gussone, and Brassica spinescens Pomel (Ellis et al. Reference Ellis, Pink, Barber and Mead1999). Intergeneric hybrids of B. napus × S. alba have also demonstrated resistance to D. radicum in Canada (Dosdall et al. Reference Dosdall, Good, Keddie, Ekuere and Stringam2000). Kott and Dosdall (Reference Kott and Dosdall2004) used introgression and selection to develop D. radicum–resistant, canola-quality germplasm. A follow-up study found white mustard and canola × white mustard crosses to have low damage from D. radicum (Tolman et al. Reference Tolman, Kott, Mayo and Murray2005b). Malchev et al. (Reference Malchev, Fletcher and Kott2010) also transferred resistance in canola, originally from S. alba, to rutabaga and developed a biochemical selection protocol using glucosinolate profiles to assist in breeding programmes for D. radicum resistance. In addition to studies conducted in Canada, various cultivars, varieties, accessions, and hybrids of cultivated brassicas have been screened for resistance to D. radicum in Europe, with some expressing variable levels of resistance or tolerance (e.g., Santolamazza-Carbone et al. Reference Santolamazza-Carbone, Velasco and Cartea2017; Niemann et al. Reference Niemann, Szwarc, Bocianowski, Weigt and Mrówczyński2020).
Camelina sativa (Linnaeus) Crantz is a poor host for D. radicum (Soroka et al. Reference Soroka, Olivier, Grenkow and Séguin-Swartz2015). In field studies in western Canada, plots of C. sativa, B. napus, and rutabaga were grown and evaluated for D. radicum damage at harvest (Soroka et al. Reference Soroka, Olivier, Grenkow and Séguin-Swartz2015). In this study, up to 50% of B. napus roots were damaged, and the rutabaga experienced severe damage from D. radicum larvae, but only three of 26 C. sativa plants had larval feeding damage, and that damage was minor (i.e., all sampled plants were classified as 0 or 1 on the scale developed by Dosdall et al. (Reference Dosdall, Herbut and Cowle1994), where 0 indicates no damage and 1 indicates very light feeding; Soroka et al. Reference Soroka, Olivier, Grenkow and Séguin-Swartz2015). Given how well D. radicum can fly, these results could provide evidence of preference for B. napus and rutabaga over C. sativa when planted in close proximity.
There are no D. radicum–resistant brassica varieties (either oilseed or vegetable crops) commercially available for farmers to grow. This could be for several reasons. First, Jyoti et al. (Reference Jyoti, Shelton and Earle2001) noted that where resistance has been observed, high levels of resistance could rarely be attributed to both antibiosis and antixenosis, and breeding programmes could not be justified because resistance is more likely to break down when conferred by only one mechanism or characteristic. Furthermore, resistant species or varieties rarely possess the marketable qualities of their susceptible counterparts, adding to the complexity of breeding programmes to achieve commercial-quality cultivars. For example, Santolamazza-Carbone et al. (Reference Santolamazza-Carbone, Velasco and Cartea2017) identified several turnip accessions with resistance to D. radicum, but these all had small, white (not purple) roots that are not desired by consumers. In the case of oilseed crops, Tansey and Dosdall (Reference Tansey and Dosdall2011) reported that newly developed resistant canola lines varied in their resistance to major canola pests, including D. radicum, cabbage seedpod weevil, Ceutorhynchus obstrictus (Marsham) (Coleoptera: Curculionidae), and flea beetle, Phyllotreta cruciferae (Goeze) (Coleoptera: Chrysomelidae). These and other insect pests of canola have overlapping distributions and pose different challenges to integrated pest management programmes on a large scale in Canada’s prairie region. Ideally, resistant B. napus hybrids would impart resistance to multiple pests, not just one, which may have stymied attempts to select and commercialise D. radicum–resistant lines. In addition, in suitable agronomic conditions, canola has impressive abilities to compensate for damage to the root system caused by D. radicum (Dosdall et al. Reference Dosdall, Clayton, Harker, O’Donovan and Stevenson2004; Soroka and Dosdall Reference Soroka and Dosdall2011).
Biological control
As chemical insecticides are becoming more challenging to use and physical or cultural practices offer only some protection (with significant trade-offs in terms of labour), biological control options for D. radicum management are an increasingly important avenue for research and development.
Predators and parasitoids
Generalist predators including members of the Carabidae and Staphylinidae prey upon the egg, larval, and pupal life stages of D. radicum (reviewed by Andreassen et al. Reference Andreassen, Kuhlmann, Mason and Holliday2007; Holliday et al. Reference Holliday, Andreassen, Dixon, Kuhlmann, Mason and Gillespie2013) and potentially account for egg mortality rates reaching 30% (Finch Reference Finch1996). Prasad and Henderson (Reference Prasad and Henderson2002) conducted plot studies to evaluate the effects of mulches on D. radicum damage to organically produced broccoli and found that the activity density of carabid and staphylinid beetles was greater in mulched plots than in control plots. They also found that D. radicum mortality was greater in mulched plots than in control plots, likely due to predation by carabid and staphylinid beetles (Prasad and Henderson Reference Prasad and Henderson2002). The relationship between predator density and predation of D. radicum eggs (and probably other life stages) is not linear, however, likely due to the complexity of the food web in agroecosystems (Prasad et al. Reference Prasad, Henderson and Snyder2002). Beetle banks and other conservation approaches, especially those that focus on increasing predator density in the field, could help to increase D. radicum mortality (Prasad and Henderson Reference Prasad and Henderson2002; Prasad et al. Reference Prasad, Henderson and Snyder2002). In Canada, the rove beetles, Aleochara bilineata Gyllenhal and Aleochara verna Say (Coleoptera: Staphylinidae), are likely the most effective predators of D. radicum eggs and larvae during their adult stage, whereas the larval stages of A. bilineata and A. verna are predators of D. radicum pupae (Wadsworth Reference Wadsworth1915; Colhoun Reference Colhoun1953).
Surveys of the various life stages of D. radicum in western Canada revealed no egg parasitoids of D. radicum (Hemachandra et al. Reference Hemachandra, Holliday, Mason, Soroka and Kuhlmann2007). The most recent survey found five parasitoids of the larval and pupal stages of D. radicum in Canada, including Aphaereta minuta (Nees) (Hymenoptera: Braconidae), Trybliographa rapae (Westwood) (Hymenoptera: Eucoilidae), one species from the genus Trichopria (Hymenoptera: Proctotrupidae), and two species from the genus Phygadeuon (Hymenoptera: Ichneumonidae) (Hemachandra et al. Reference Hemachandra, Holliday, Mason, Soroka and Kuhlmann2007). The D. radicum–parasitoid community in Canada is similar to that of Europe, with three notable additional parasitoids in Europe: Phygadeuon trichops Thomson (Hymenoptera: Ichneumonidae), Aleochara bipustulata (Linnaeus), and Aleochara brevipennis Gravenhorst (Coleoptera: Staphylinidae) (Hemachandra et al. Reference Hemachandra, Holliday, Mason, Soroka and Kuhlmann2007). Of the latter, both P. trichops and A. bipustulata were released in Canada in the 1950s but did not establish (McLeod Reference McLeod1962). Work by Hemachandra et al. (Reference Hemachandra, Holliday, KIimaszewski, Mason and Kuhlmann2005) confirmed that A. bipustulata is not present in Canada, either because it did not establish or because it was misidentified. Because A. bipustulata is absent in Canada, it was studied for use in a subsequent importation biological control programme to manage D. radicum (Andreassen et al. Reference Andreassen, Kuhlmann, Mason and Holliday2009; reviewed by Holliday et al. Reference Holliday, Andreassen, Dixon, Kuhlmann, Mason and Gillespie2013). Although the efficacy testing showed promise, to our knowledge, a petition for import and release has not yet been submitted (Canadian Food Inspection Agency 2025), and A. bipustulata has not been released into Canadian agroecosystems.
Aleochara bilineata is the most prevalent predator of D. radicum in Canadian agroecosystems (Read Reference Read1962; Turnock et al. Reference Turnock, Boivin and Whistlecraft1995; Dixon et al. Reference Dixon, Coady, Larson and Spaner2004; Hemachandra Reference Hemachandra2004; Broatch et al. Reference Broatch, Dosdall, O’Donovan, Harker and Clayton2010). In an Albertan field study conducted in 2005 and 2006, for example, Hummel et al. (Reference Hummel, Dosdall, Clayton, Harker and O’Donovan2009) observed predation rates of up to nearly 82% at some sites, and Turnock et al. (Reference Turnock, Boivin and Whistlecraft1995) suggested that predation of D. radicum by A. bilineata could reach 94%, although they also found that rates varied widely between study sites in Manitoba, Quebec, Ontario, and Newfoundland. Compared to predation by A. bilineata, parasitism of D. radicum by T. rapae tends to be lower and is also variable between provinces. For example, parasitism by T. rapae ranged from approximately 2.2% to 14.6% in Alberta (Hummel et al. Reference Hummel, Dosdall, Clayton, Harker and O’Donovan2009). Therefore, although parasitism by Hymenoptera may help to manage D. radicum populations, it appears that rove beetles may offer stronger control.
Parasitoids of related Dipteran pests might also accept D. radicum as hosts and could provide new avenues for parasitoid research, although this approach has not recently been evaluated. One example from the literature is a study conducted by Floate et al. (Reference Floate, Soroka and Spooner1999), who investigated parasitoids of the house fly, Musca domestica Linnaeus (Diptera: Muscidae), for their ability to use D. radicum as hosts for oviposition and development. Although Muscidifurax raptorellus Kogan (Diptera: Muscidae), Muscideifurax zaraptor Kogan (Diptera: Muscidae), and Trichomalopsis sarcophagae Gahan (Hymenoptera: Pteromalidae) can use D. radicum as hosts, all three species demonstrated a clear preference for M. domestica and, in greenhouse studies reviewed by Soroka et al. (Reference Soroka, Kuhlmann, Floate, Whistlecraft, Holliday, Boivin, Mason and Huber2001), parasitism of D. radicum by these parasitoids was negligible. Parasitoids from other, more closely related Dipteran pests may prove more promising as biological control agents of D. radicum.
Entomopathogenic fungi
Entomopathogenic fungi, primarily of the order Hypocreales, are important mortality factors affecting insects in the field, and their use as biocontrol agents has been investigated in diverse taxa (Shah and Pell Reference Shah and Pell2003). The results of fungal infection are often dramatic, and affected insects have the potential to serve as nuclei for further infection of targeted hosts, prolonging the efficacy of treatments. In addition, with advances in sterile culture technique, the production of infective material to be formulated into pesticide products is relatively inexpensive, and the products are attractive for use in organic or low-chemical input agricultural systems.
The use of such fungal biopesticides has been explored in the management of root maggot flies, primarily in Europe (Vänninen et al. Reference Vänninen, Hokkanen and Tyni-Juslin1999; Chandler and Davidson Reference Chandler and Davidson2005) and the United States of America (Vänninen et al. Reference Vänninen, Hokkanen and Tyni-Juslin1999; Bruck et al. Reference Bruck, Snelling, Dreves and Jaronski2005). The efficacy of fungal biopesticides in field conditions is often not superb (Herbst et al. Reference Herbst, Razinger, Ugrinović, Škof, Schroers, Hommes and Poehling2017). Published Canadian research has dealt primarily with the natural history of wild entomopathogens (Nair and McEwen Reference Nair and McEwen1973) or the screening of many candidates in laboratory bioassays (Myrand et al. Reference Myrand, Buffet and Guertin2015). BotaniGard 22WP was registered in Canada in 2009 and has been field-tested alongside and in combination with insecticides in mineral soils (Tolman et al. Reference Tolman, Dries and McFadden2001a, Reference Tolman, Dries and McFadden2001b, Reference Tolman, Dries and Muth2003, Reference Tolman, Steffler, Alhemzawi and McPherson2009a). When combined with a spinosad product, cabbage showed little D. radicum damage (Tolman et al. Reference Tolman, Steffler, Alhemzawi and McPherson2009a), whereas combining BotaniGard® with a garlic barrier as an in-furrow application provided poor control in summer turnip (Tolman et al. Reference Tolman, Dries and Muth2003). Combining BotaniGard with insecticides gave variable results. When paired with LorsbanTM or Canon 200SC, cabbage had excellent protection (Tolman et al. Reference Tolman, Dries and McFadden2001a), while pairing with Actara® resulted in low protection of summer turnip (Tolman et al. Reference Tolman, Dries and McFadden2001b). A significant challenge with the use of fungal biocontrol agents against D. radicum is the need to target the larvae, which spends considerable time belowground. Future research may discover a way to target D. radicum for fungal infection that results in quick kill of the larvae and protection of brassicas in the field, but the science is not there yet.
Entomopathogenic nematodes
Numerous species of entomopathogenic nematode, predominantly within the Steinernematidae and Heterorhabditidae families (Order: Rhabditida), have been explored and cultivated for the purpose of biological control. Historically, the evaluation of nematodes against D. radicum has occurred under laboratory conditions, with field studies showing variable results (Bracken Reference Bracken1990). Efforts in 2017 (McDonald et al. Reference McDonald and Vander Kooi2018a, Reference McDonald and Vander Kooi2018b) in Ontario muck soils found Steinernema feltiae (Filipjev) to control D. radicum in cabbage and radish as well as LorsbanTM. Trials combining nematode species S. feltiae and S. carpocapsae (Weiser) gave similar protection to that observed with Verimark® (Van Dyk and Blauel Reference Van Dyk and Blauel2023). Renewed interest in using nematodes against D. radicum found S. feltiae and S. kraussei (Steiner) to be highly efficacious (Patuwatha Withanage et al. Reference Patuwatha Withanage, Briar and Edeogu2024). Efforts to improve the performance of entomopathogenic nematodes have sought to isolate strains already adapted to the region where they are intended to be used (Shapiro-Ilan et al. Reference Shapiro-Ilan, Han and Dolinski2012). Sharifi-Far et al. (Reference Sharifi-Far, Shapiro-Ilan, Brownbridge and Hallett2018) explored a combined approach in which a strain of Heterorhabditis bacteriophora (Poinar) with suitable traits was identified and then inbred to ensure the trait was retained. Although recent studies have shown entomopathogenic nematodes to be equally as effective as chemical options, the cost associated with this technology has hindered adoption by growers. Research efforts to reduce the cost of rearing or strategies that minimise the area to be treated are needed.
Sterile insect technique
In Canada, the sterile insect technique is not widely used as a control method, despite it being highly effective, species-specific, environmentally friendly, and used in other areas of the world (Dyck et al. Reference Dyck, Hendrichs and Robinson2021). In British Columbia, codling moth, Cydia pomonella (Linnaeus) (Lepidoptera: Tortricidae), has been successfully managed since the 1960s (Proverbs et al. Reference Proverbs, Newton and Campbell1982; Thistlewood and Judd Reference Thistlewood and Judd2019), and onion maggot, Delia antiqua, has been managed in Quebec with the sterile insect technique since 2011, with some growers in Ontario also using the technique (Fournier and Brodeur Reference Fournier and Brodeur2012; Fortier Reference Fortier2021; Fortier et al. Reference Fortier, Brodeur, Fournier, Vankosky and Martel2024). Use of the sterile insect technique against D. radicum began in Quebec in 2012 and 2013 (Fournier and Fortier Reference Fournier and Fortier2014), where sterilisation parameters were first validated, for example, greater than 90% sterility in males achieved with an irradiation dose of 35 Gy (Cobalt-60) applied to pupae at least eight days old. Irradiated males exhibited longevity and sexual competitiveness comparable to non-irradiated males. Small-scale releases of sterile D. radicum then occurred in three 1-ha radish plots, with control plots located within the same field for comparison. A total of 290 000 sterile flies were released in each plot between April and June 2013. Although eggs collected from the sterile insect technique plots had a sterility rate of 79.4 ± 0.6% compared to 14.2 ± 7.3% in the control plots, crop damage did not differ between treated and control plots. It is suspected that other Delia species (e.g., D. platura) contributed to this damage, as egg counts from this species could reach 90% of the eggs observed near the plant stem. It must be recognised that in areas where D. radicum and D. platura co-occur, they often overlap within a field. It has been observed that Delia platura will commence egg-laying later than D. radicum and will continue to lay eggs after D. radicum has ceased (S. Blatt, unpublished data). For areas where these species overlap, the sterile insect technique may be less effective in years of high D. platura pressure.
Between 2014 and 2016, a second set of experiments occurred in daikon radish and broccoli (Fortier et al. Reference Fortier, Fournier and Sanchez2017). Distance between the sites was found to be critical as, in 2014 and 2015, the fields were divided into two 1-ha sections separated by a 1-ha buffer zone. High pest pressure in broccoli fields in 2015, combined with the migration of fertile flies from outside the field, resulted in increased damage levels observed in the sterile insect technique release sections. In 2016, entire fields were treated and compared with conventionally managed fields on the same farm. This approach improved the sterile–wild insect ratio compared to that of the 2015 season; however, warm, dry spring conditions likely reduced egg and larval survival, contributing to very low damage and the lack of differences between treatments. In the daikon sites, the sterile insect technique reduced damage from D. radicum to 16.5 ± 3.6% and 1.0 ± 1.0% in treated fields, compared to 31.0 ± 5.9% and 39.0 ± 4.3% in control fields (Fortier et al. Reference Fortier, Fournier and Sanchez2017). For the sterile insect technique to achieve effective control, sterile insects must be released in sufficient numbers during periods of peak wild population activity. Research from the 1980s and 1990s found spring emergence of D. radicum to follow a bimodal distribution, with “early” and “late” phenotypes (Biron et al. Reference Biron, Langlet, Boivin and Brunei1998). Adult monitoring using sticky traps is essential to characterise the spring emergence to ensure optimal timing of sterile fly releases.
From 2019 to 2022, the efficacy of the sterile insect technique was studied across different crops (e.g., daikon radish, radish, broccoli, cauliflower, and Chinese cabbage) and across various agro-environmental conditions. Nearly 70 fields (sterile insect technique–treated and controls) were monitored in Quebec under commercial conditions and demonstrated the effectiveness of the technique with significant reductions in crop damage (Fortier et al. Reference Fortier, Bush-Beaupre and Savage2025). Infestation rates were 5.6% in radish and 2.1% in daikon for sterile insect technique–treated fields, compared to 19.4% and 11.8% in untreated fields. Results in cauliflower, broccoli, kale, and Chinese cabbage varied by site and crop type, with damage levels ranging from similar to higher or lower in sterile insect technique–treated fields compared to conventional controls. Based on these studies conducted in flowering brassica species, the presence of Delia platura and/or D. florilega in a large proportion of infested plants, coupled with the high cost of sterile fly production, currently makes the sterile insect technique less appealing to producers of crops such as broccoli and cauliflower. Evaluation of the sterile insect technique in brassica root crops, for example, rutabaga, has not occurred owing to the potential for damage to the marketable root.
Despite the challenges, the sterile insect technique has been gaining interest in Quebec. One grower expanded their use of the sterile insect technique in daikon radish between 2017 and 2024 to include their entire daikon acreage. As the wild population decreased over these five years, dropping from an average of 4.0 to that of 0.2 flies per trap per day, the number of released sterile flies also decreased from an average of 520 000 flies per hectare in 2015 to 44 000 flies per hectare in 2019 (Fortier Reference Fortier2021). This successfully eliminated insecticide use against D. radicum and reduced crop damage from 30–50% in 2013–2015 to approximately 1–3% in 2017–2019 (Lucie Leblanc, agronomist, personal communication). It is suspected that the location of this farm played a role in the success of the trial, as it is somewhat isolated from neighbouring crops and the grower uses protective netting against the first generation of D. radicum to complement their use of the sterile insect technique.
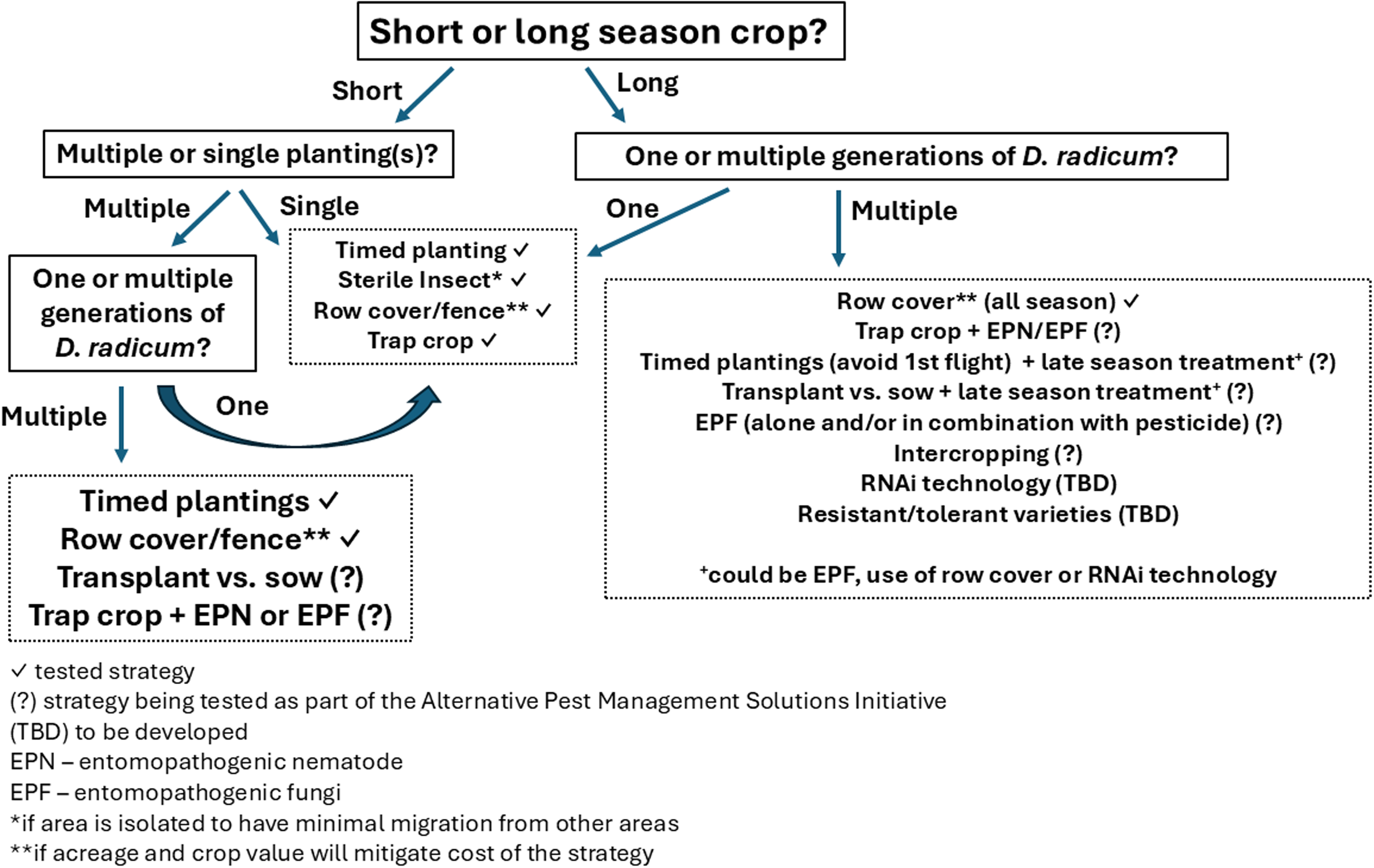
Future directions
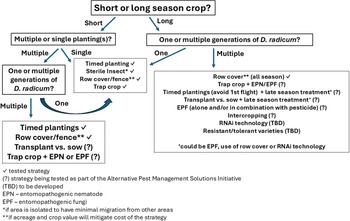
The previously described integrated pest management tactics all have merit for use by brassica growers in Canada. There are three main challenges associated with the recommendation of a single strategy for D. radicum management across Canada: (1) variability in the life history and seasonal phenology; (2) variability in population density, subsequent pest pressure, and insecticide susceptibility; and (3) variability in economic impact to different oilseed and vegetable crops grown across Canada. Additionally, the wide range of brassica crops (e.g., marketable product, economic value) and other pest species attacking brassicas further adds to the complexity of selecting a single management strategy for all provinces and commodities. Figure 2 provides an overview of the variables to be considered when selecting a management strategy for D. radicum and highlights where effective strategies have been demonstrated and where research gaps exist. The decision to plant a short-season versus a long-season crop is perhaps the single most important factor affecting which management strategy will be possible. Short-season crops, such as broccoli, may encounter only a single generation of D. radicum, unlike longer-season crops that may experience two or three generations of attack, such as rutabaga in the western provinces. While any of the described strategies can show efficacy against a single generation of D. radicum, multiple generations of the pest may require a combination of tactics. For example, timed planting or trap crops could address the first generation of D. radicum in a long-season crop but would need to be paired with a late-season application of a biological control, that is, entomopathogenic fungi or entomopathogenic nematodes, to address later generations. Research on combining strategies has only recently been examined in Canada. In 2022, projects to evaluate the efficacy of integrated pest management tactics applied in novel combinations for Delia management were initiated by Agriculture and Agri-Food Canada’s Pest Management Centre, partly supported through the Alternative Pest Management Solutions Initiative. Broccoli and rutabaga represent short- and long-season high-value crops and were ideal subjects for these studies. Rutabaga trials in Saskatchewan combined intercropping and a different rutabaga variety, while in Ontario, the trials explored netting, entomopathogenic nematodes, and sterile insect release, tested singly and in combination. Broccoli trials occurred in Newfoundland, Nova Scotia, and British Columbia and evaluated trap cropping, entomopathogenic nematodes, and entomopathogenic fungi, tested singly and in combination. The trials are ongoing. It is recognised that combining strategies to protect a crop could be challenging for many growers owing to labour needs or other costs associated with the implementation of the strategy.
Flow chart showing tested and potential management strategies (boxes with dotted lines) for Delia radicum with consideration of crop and pest variables (boxes with solid lines).
For at least the last 30 years, insecticides with the active ingredient chlorpyrifos (e.g., LorsbanTM) have been widely and fairly successfully used to protect against D. radicum feeding damage and yield loss in brassica vegetable plantings in Canada. With the deregistration of chlorpyrifos in 2023 (Health Canada 2023), finding novel, alternative, or integrated management techniques is imperative. With limited pesticide chemistry options available in Canada, other chemistries have been examined in the United States of America (Joseph and Zarate Reference Joseph and Zarate2015) that may warrant further evaluation in the field in Canada. These include neonicotinoids (i.e., clothianidin), pyrethroids (i.e., zeta-cypermethrin, bifenthrin, and fenpropathrin), and electron transport inhibitors (i.e., tolfenpyrad). Going forwards, integrated pest management programmes that incorporate or combine the tactics reviewed here are likely to provide the best and most sustainable suppression of D. radicum. Field research to test different integrated pest management strategies and to demonstrate the most effective approaches is needed to encourage farmer adoption of novel integrated pest management strategies. Conducting cost–benefit analyses during the research and demonstration phases is also important, and these analyses should also consider possible long-term benefits to the agroecosystem. This is because integrated pest management programmes can be more complicated and costly to implement but can require decreasing investment over time with increasingly effective pest suppression, as demonstrated by the sterile insect technique (e.g., Fournier and Fortier Reference Fournier and Fortier2014; Fortier et al. Reference Fortier, Fournier and Sanchez2017). New molecular technologies for crop breeding and insect pest management (e.g., RNA interference technologies) should also be developed and incorporated into integrated pest management programmes to manage D. radicum.
Acknowledgements
The authors thank Cezarina Kora and Caitlin Watt for thoughtful suggestions on the manuscript.
Competing interests
The authors declare that they have no competing interest.

 Open access
Open access