




1. Introduction
Plants develop characteristic shoot architectures by extending branches at specific angles, creating distinct forms such as canopy structures or tree shapes (Nakamasu & Higaki, Reference Nakamasu and Higaki2019; Wang & Jiao, Reference Wang and Jiao2018). The shoot architecture is a critical trait that contributes to the acquisition of resources, such as light and water, and mechanical stability by mitigating loads from self-weight or environmental factors, such as snow. To adapt to favorable growth conditions, plants exhibit tropic responses, such as phototropism, the bending of stems toward light, and gravitropism, the bending of stems in relation to gravity (Morita, Reference Morita2010). In addition, plants have evolved mechanisms for organ straightening after bending, contributing to their structural integrity. For instance, wild-type Arabidopsis thaliana plants subjected to a clinostat-induced microgravity environment in darkness exhibit gradual straightening of the bent shoots over time through coordinated cellular and tissue-level responses (Okamoto et al., Reference Okamoto, Ueda, Shimada, Tamura, Kato, Tasaka, Morita and Hara-Nishimura2015). In contrast, stems of the myosin xif xik double mutant (hereafter xifk), which lacks functional MYOSIN XIf and XIk proteins, fail to straighten and instead are coiled on a clinostat, suggesting a critical role for these motor proteins in maintaining shoot posture and responding to gravitational stimuli (Okamoto et al., Reference Okamoto, Ueda, Shimada, Tamura, Kato, Tasaka, Morita and Hara-Nishimura2015). MYOSIN XIf and XIk are not only essential for stem straightening after bending but also play a critical role in regulating stress distribution within stems, ensuring mechanically favorable shapes (Tsugawa et al., Reference Tsugawa, Miyake, Okamoto, Toyota, Yagi, Morita, Hara-Nishimura, Demura and Ueda2023). This coordination of bending and straightening responses enables wild-type plants to maintain structural integrity, whereas myosin XI mutants exhibit disrupted stress regulation and impaired postural control.
Among the 35 myosin classes known in eukaryotes, plant myosins are categorized in classes VIII and XI (Odronitz & Kollmar, Reference Odronitz and Kollmar2007). In plants, actin–myosin XI complexes play critical roles in intracellular vesicle trafficking and organelle positioning (Schuh, Reference Schuh2011). Of the 13 myosin XI family members in A. thaliana, XI1, XI2, XIb, XI-i, and XIk have been identified as crucial participants in the movement of cellular organelles, including the endoplasmic reticulum, Golgi apparatus, peroxisomes, mitochondria, and nucleus (Peremyslov et al., Reference Peremyslov, Prokhnevsky and Dolja2010; Prokhnevsky et al., Reference Prokhnevsky, Avisar and Dolja2008; Tamura et al., Reference Tamura, Iwabuchi, Fukao, Kondo, Okamoto, Ueda, Nishimura and Hara-Nishimura2013; Ueda et al., Reference Ueda, Yokota, Kutsuna, Shimada, Tamura, Shimmen, Hasazawa, Dolja and Hara-Nishimura2010). Notably, MYOSIN XI2 and MYOSIN XIk are the primary motor proteins driving cytoplasmic streaming in vegetative organs (Peremyslov et al., Reference Peremyslov, Prokhnevsky and Dolja2010; Prokhnevsky et al., Reference Prokhnevsky, Avisar and Dolja2008). Although myosin xik (hereafter xik) and myosin xi2 (hereafter xi2) mutants exhibit impaired intracellular transport, these mutations generally do not result in notable growth defects, except inhibition of root hair elongation (Prokhnevsky et al., Reference Prokhnevsky, Avisar and Dolja2008; Ueda et al., Reference Ueda, Yokota, Kutsuna, Shimada, Tamura, Shimmen, Hasazawa, Dolja and Hara-Nishimura2010).
Although recent quantitative analyses and mathematical modeling have advanced our understanding of the mechanical stability of primary stems under gravitational forces in myosin XI mutants (Tsugawa et al., Reference Tsugawa, Miyake, Okamoto, Toyota, Yagi, Morita, Hara-Nishimura, Demura and Ueda2023), shoot architecture in A. thaliana is shaped not only by the primary stem but also by lateral branches. Lateral branches are critical for optimizing light capture, enhancing mechanical stability, and supporting reproductive strategies. Therefore, analyzing the entire plant structure, including lateral branches, is essential to fully comprehend how shoot architecture responds to environmental and developmental cues. Understanding shoot architecture at the whole-plant level requires a detailed comparison of wild-type and myosin XI mutant plants, taking into account the contributions of lateral branches. To achieve this, it is important to quantitatively evaluate the temporal changes of lateral branch morphology. Two-dimensional imaging alone is insufficient for this purpose, as lateral branch morphology is inherently three-dimensional. Accurate morphological analysis requires advanced techniques, such as three-dimensional reconstruction. We previously developed a method based on a volume intersection method to reconstruct the three-dimensional architecture of Arabidopsis primary stems and lateral branches and to monitor their temporal changes, in a so-called four-dimensional analysis, providing a framework for quantitative evaluation (Kunita et al., Reference Kunita, Morita, Toda and Higaki2021).
In this study, we aimed to evaluate the contributions of MYOSIN XIf and XIk to the formation of lateral branches using a four-dimensional reconstruction system and quantitative analysis of lateral branch morphology. Employing this system, we quantified lateral branch morphology in wild-type and myosin XI mutant plants, focusing on critical metrics, such as branch angle and straightness, with the latter specifically introduced to evaluate phenotypic differences in the mutants. This approach enables a comprehensive analysis of how myosin XI mutations affect lateral branch morphology, providing novel insights into their roles in shaping plant shoot architecture at the whole-plant level.
2. Materials and methods
2.1. Plant materials
Wild-type Arabidopsis thaliana Columbia-0 (Col-0) and the myosin XI null mutants xif, xik, and the xifk double mutant were used in this study (Okamoto et al., Reference Okamoto, Ueda, Shimada, Tamura, Kato, Tasaka, Morita and Hara-Nishimura2015; Ueda et al., Reference Ueda, Yokota, Kutsuna, Shimada, Tamura, Shimmen, Hasazawa, Dolja and Hara-Nishimura2010). Seeds were sown on potting soil (Jiffy-7; Sakata Seed Corp., Yokohama, Japan) and incubated in a growth chamber (Plantflec LH-241PFP-S; NK System, Tokyo, Japan) under a 16-h light/8-h dark cycle with illumination from an 86.2 μmol m−2 s−1 light-emitting diode at a constant temperature of 23.5°C for approximately 10 days. The seedlings were then transplanted into vermiculite and grown under the same conditions. Plants were watered with a solution containing 0.5 g/L Hyponex fertilizer (Hyponex Japan, Osaka, Japan). Plants aged 40–50 days were used for imaging. Prior to imaging, leaves near the base of the lateral branches were carefully removed with precision scissors. The removal of the cauline leaves does not affect the angle of the branch base. The rosette leaves were covered with black cloth to minimize interference. After imaging, the black cloth was removed and the plants were returned to the growth chamber.
2.2. Image acquisition and three-dimensional reconstruction
Image acquisition and three-dimensional reconstruction were performed following the method described by Kunita et al. (Reference Kunita, Morita, Toda and Higaki2021). A digital single-lens reflex camera (EOS Kiss X10, Canon, Tokyo, Japan) equipped with a fixed-focus lens (EF28IS, Canon) was used to capture images. Plants were mounted on a rotating platform (RT-5; TAITEC, Saitama, Japan) with a calibration target for camera pose estimation, ensuring precise alignment. The platform was rotated at a speed of approximately 2.5 rpm, and videos were recorded for 3 min with the camera tilted at an angle of −8.5°. The recording settings were standardized to 30 fps with a resolution of 1,080 × 1,920 pixels, corresponding to a spatial resolution of 0.25 mm per pixel. Video frames were extracted as still images and used for three-dimensional reconstruction.
The three-dimensional reconstruction was performed using the volume intersection method (Cheung et al., Reference Cheung, Baker and Kanade2005; Kutulakos & Seitz, Reference Kutulakos and Seitz2000; Tsai, Reference Tsai1987). The camera projection matrix for each captured image was estimated using a simplified calibration method with a calibration target, following the approach described by Baumberg et al. (Reference Baumberg, Lyons and Taylor2005), Datta et al. (Reference Datta, Kim and Kanade2009), and Kunita et al. (Reference Kunita, Morita, Toda and Higaki2021). Specifically, the calibration target was aligned with the X–Y plane of the world coordinate system (Z = 0), and the camera matrix was estimated by mapping the world coordinates of 60 red circular markers on the calibration target to their corresponding positions in the captured images. Silhouette images were then generated by segmenting plant regions from the captured images. For each silhouette image, a cone was constructed with the camera’s optical center as the vertex and the silhouette region as the cross-section. The three-dimensional structure of the plant was reconstructed by computing the intersection of cones derived from images captured at multiple angles. The reconstruction accuracy depended on the number and distribution of captured images. To account for partial occlusions, a cross-ratio threshold was applied to define valid intersections. The cross ratio was defined as the percentage of images contributing to the intersection volume, with a threshold of 95% applied in this study (Kunita et al., Reference Kunita, Morita, Toda and Higaki2021). Volume rendering was performed using Imaris (Bitplane, Belfast, UK).
2.3. Quantification of branch angles and straightness
The reconstructed three-dimensional images were analyzed to quantify lateral branch morphology. Skeletonization of plant structures was performed using the default algorithm in ImageJ software (Schneider et al., Reference Schneider, Rasband and Eliceiri2012). The primary stem was defined as the shortest path connecting manually designated endpoints at the base and tip. Lateral branches were identified as paths originating from branching points with more than two voxel connections on the primary stem.
The angle between the primary stem and each lateral branch,
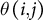 $\theta \left(i,j\right)$
, was calculated as follows:
$\theta \left(i,j\right)$
, was calculated as follows:
 $$\begin{align*}\theta \left(i,j\right) = {\cos}^{-1}\left(\frac{Vstem(i)\cdotp Vbranch(j)}{\parallel Vstem(i)\parallel \parallel Vbranch(j)\parallel}\right)\end{align*}$$
$$\begin{align*}\theta \left(i,j\right) = {\cos}^{-1}\left(\frac{Vstem(i)\cdotp Vbranch(j)}{\parallel Vstem(i)\parallel \parallel Vbranch(j)\parallel}\right)\end{align*}$$
Here,
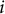 $i$
is the number of branching points, and
$i$
is the number of branching points, and
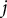 $j$
denotes the voxel index along the lateral branch. The primary stem vector,
$j$
denotes the voxel index along the lateral branch. The primary stem vector,
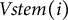 $Vstem(i)$
, was calculated as the vector connecting the average coordinates of consecutive voxels along the primary stem, from branching points
$Vstem(i)$
, was calculated as the vector connecting the average coordinates of consecutive voxels along the primary stem, from branching points
 $i-1$
to
$i-1$
to
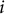 $i$
and from
$i$
and from
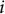 $i$
to
$i$
to
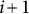 $i+1$
. To calculate the angle with the vertical vector,
$i+1$
. To calculate the angle with the vertical vector,
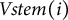 $Vstem(i)$
was set as (0,0,1). The lateral branch vector,
$Vstem(i)$
was set as (0,0,1). The lateral branch vector,
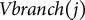 $Vbranch(j)$
, was computed based on the voxels between
$Vbranch(j)$
, was computed based on the voxels between
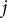 $j$
and
$j$
and
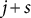 $j+s$
, where
$j+s$
, where
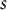 $s$
was set to 20, corresponding to 20-mm intervals.
$s$
was set to 20, corresponding to 20-mm intervals.
To quantitatively evaluate the degree of straightening in the lateral branches, we introduced straightness, a metric that quantifies the branch linearity. Straightness at the
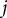 $j$
th voxel on the lateral branch was calculated as the ratio of the straight-line distance
$j$
th voxel on the lateral branch was calculated as the ratio of the straight-line distance
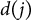 $d(j)$
from the branching point to the
$d(j)$
from the branching point to the
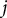 $j$
th voxel to the trace path length
$j$
th voxel to the trace path length
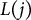 $L(j)$
from the branching point. A straightness value closer to 1 indicates greater branch linearity.
$L(j)$
from the branching point. A straightness value closer to 1 indicates greater branch linearity.

Figure 1. Representative images of Arabidopsis thaliana wild type (Col-0) and myosin xif, xik, and xif xik (xifk) mutants. (a–d) Photographs captured with the target pattern (red) for three-dimensional (3D) reconstruction. Images of the rotating plant were captured using a fixed camera to obtain plant images from various angles, and the plant architecture was reconstructed in 3D using the volume intersection method (Kunita et al., Reference Kunita, Morita, Toda and Higaki2021). Wild type (Col-0) (a), myosin xif (B), xik (c), and xif xik (xifk) mutants (d) are shown. (e–h) 3D reconstructions of the aboveground parts of (a–d). (i–l) 3D views of a single lateral branch extracted from (e–h).
3. Results
3.1. Three-dimensional morphological analysis of lateral branch phenotypes in myosin XI mutants
Morphological observations of lateral branches revealed distinct differences among the examined A. thaliana genotypes. Wild-type plants and the xif mutant displayed lateral branches with a characteristic S-shaped curvature, characterized by pronounced bending at both the base and tip (Figure 1a, b). In contrast, lateral branches of the xik mutant exhibited reduced curvature at both the base and tip, resulting in greater linearity in overall morphology (Figure 1c). The xifk double mutant displayed a unique phenotype, with branches appearing predominantly linear yet marked by fine, repetitive bends, resembling a previously coiled wire that had been partially straightened (Figure 1d). These observations indicate that mutations in myosin XI genes profoundly affect lateral branch morphology.
To quantitatively evaluate these phenotypic differences, we employed a three-dimensional reconstruction system as described by Kunita et al. (Reference Kunita, Morita, Toda and Higaki2021). Plants were mounted on a rotating platform equipped with a calibration target, and videos were recorded for 3 min (Figure 1a–d). Individual frames from the videos were extracted and processed using the volume intersection method to generate three-dimensional stack images, which were then rendered to reconstruct the three-dimensional structure of the lateral branches (Figure 1e–l). Lateral branch morphology was quantified using the metrics of the angle relative to the primary stem or the gravity vector and straightness, as described in the Materials and Methods section. Briefly, branching points and the primary stem axis were identified, lateral branches were divided into unit lengths, and angles were measured between the primary stem axis and the gravity vector, respectively, and each segment of the lateral branch. Angles approaching 90° indicated horizontal growth, whereas angles greater than 90° reflected drooping branches. By measuring angles at multiple points along the entire length of each lateral branch, we created spatial profiles of angle distribution across the branches (Figure 2a–d and Supplementary Figure S1A–D). These spatial distribution profiles revealed that wild-type branches exhibited large angles at the base and tip, with smaller angles in the middle, resulting in a vault-like shape consistent with the observed S-shaped curvature (Figure 2a). The xif and xik mutants displayed spatial distributions similar to the wild type; however, the xik mutant showed a general tendency toward higher angle values throughout the branches (Figure 2b, c). In contrast, the xifk double mutant exhibited a disrupted vault-like shape, indicating that the S-shaped curvature of the branches was compromised (Figure 2d). Similar results were obtained from the angle plots of the lateral branch relative to the gravity vector (Supplementary Figure S1A–D). Statistical analysis revealed that the branches of the xik mutant and the xifk double mutant exhibited significantly higher angle values compared with those of the wild type, with the increase being more pronounced in the double mutant (Figure 2e and Supplementary Figure S1E). These results suggest a greater degree of branch drooping in these mutants.

Figure 2. Spatial distribution of lateral branch angles in Arabidopsis relative to the primary stem. (a–d) Spatial distribution of lateral branch angles in the wild type (Col-0) (a) and myosin xif (b), xik (c), and xif xik (xifk) mutants (d). Each branch is represented by a distinct color (N = 11). (e) Mean angles of lateral branches relative to the primary stem. Box plots show the angles of lateral branches relative to the primary stem. ** P < 0.01, * P < 0.03 (Mann–Whitney U-test, N = 11 individuals).

Figure 3. Spatial distribution of lateral branch straightness in Arabidopsis. (a–d) Spatial distribution of lateral branch straightness in the wild type (Col-0) (a) and myosin xif (b), xik (c), and xif xik (xifk) mutants (d). Each branch is represented by a distinct color (N = 11). (e) Mean straightness of lateral branches. Box plots show the straightness of lateral branches. ** P < 0.01, * P < 0.03 (Mann–Whitney U-test, N = 11 individuals).

Figure 4. Temporal changes in lateral branch angles in Arabidopsis wild type (Col-0) and myosin XI mutants. (a–d) Spatial distribution of lateral branches plotted using one branch per individual plant from three different individuals in the wild type (Col-0) (a) and myosin xif (b), xik (c), and xif xik (xifk) mutants (d). Color intensity represents the number of observation days (Days 1, 2, and 3). (e–h) Temporal evolution of branch angle in the wild type (Col-0) (e) and myosin xif (f), xik (g), and xif xik (xifk) mutants (h). These plots represent the mean values of the results shown in (a–d). Color intensity represents the replicate number (#1, 2, and 3).

Figure 5. Temporal changes in lateral branch straightness in Arabidopsis wild type (Col-0) and myosin XI mutants. (a–d) Spatial distribution of lateral branches plotted using one branch per individual plant from three different individuals in the wild type (Col-0) (a) and myosin xif (b), xik (c), and xif xik (xifk) mutants (d). Color intensity represents the number of observation days (Days 1, 2, and 3). (e–h) Temporal evolution of branch straightness in the wild type (Col-0) (e) and myosin xif (f), xik (g), and xif xik (xifk) mutants (h). These plots represent the mean values of the results shown in (a–d). Color intensity represents the replicate number (#1, 2, and 3).
Straightness, defined as the ratio of the straight-line distance from the branch base to the tip to the total path length, provided distinct insights into branch morphology. Higher straightness values indicate greater branch linearity. In wild-type and xif branches, straightness values typically ranged from 0.80 to 0.85, except near the base (Figure 3a, b). In contrast, the xik and xifk mutants predominantly exhibited values between 0.85 and 0.90, although the xifk double mutant showed greater variability (Figure 3c, d). These values reflect greater linearity in the branch morphology. Statistical analysis further revealed that the branches of the xik mutant and xifk double mutant showed significantly higher straightness values compared with those of the wild type, indicating greater linearity in these mutants (Figure 3e).
3.2. Temporal analysis of lateral branch morphology
The three-dimensional reconstruction system used in this study is a non-destructive method that causes negligible damage to the plants, allowing for temporal acquisition of three-dimensional images and enabling four-dimensional data analysis (Kunita et al., Reference Kunita, Morita, Toda and Higaki2021). To investigate the growth of lateral branches, we tracked and analyzed their three-dimensional structures over a 3-day period, capturing images at 24-h intervals (Supplementary Figure S2). This approach allowed us to quantitatively evaluate the contribution of MYOSIN XIf and XIk to changes in branch morphology over time. Results are presented for three replicates per genetic background (Figures 4a–d, 5a–d, and Supplementary Figure S3A–D). The x-axis represents the branch length, illustrating branch elongation during the observation period. In the experimental system, the branch length included the floral organs at the branch tip. Individual differences in branch elongation within each genotype were greater than the difference between genotypes (Figures 4a–d, 5a–d, and Supplementary Figure S3A–D). Therefore, no significant difference in the elongation rate of lateral branches between genotypes was apparent.
The angle plots relative to the primary stem revealed that lateral branches of wild-type plants and the xif mutant exhibited a vault-like shape in their angle distribution during the second day, corresponding to branch lengths of 20–40 mm (Figure 4a, b). This vault-like shape was characterized by large angles at the base and tip, with smaller angles in the middle, corresponding to the S-shaped curvature observed in the branches themselves. In contrast, branches of the xik mutant displayed a similar vault-like angle distribution, but exhibited higher angle values in the middle region compared with those of the wild type and xif mutant, particularly as observed in xik replicate #1 (Figure 4c). This pattern suggested a tendency for the branches to droop, as indicated by the downward direction of the graphs in Figure 4. For the xifk double mutant, the angle distribution deviated from a vault-like shape, exhibiting pronounced fluctuations in angle values throughout the branch (Figure 4d). This pattern suggested instability in the overall direction of branch elongation. The angle plots relative to the gravity vector showed essentially the same trend (Supplementary Figure S3A–D). Time-lapse straightness distribution analysis revealed a reduction in straightness as branches elongated in all observed wild-type plants (Figure 5a). In contrast, certain branches in the xif, xik, and xifk mutants showed increased straightness during elongation (Figure 5b–d).
To further investigate differences in the morphological behavior of lateral branch elongation, we compared the mean angle and straightness values across genotypes. While wild-type plants and the xif mutant exhibited a decrease in angles over time, the xik mutant and xifk double mutant did not show a consistent trend of increasing angles (Figure 4e–h and Supplementary Figure S3E–H). Straightness consistently decreased in all three wild-type plants during elongation, whereas the myosin XI mutants displayed greater variability, with some branches showing increases and others decreasing in straightness over time (Figure 5e–h).
4. Discussion
This study leveraged advanced three-dimensional reconstruction and temporal imaging techniques to provide a comprehensive four-dimensional analysis of lateral branch morphology in A. thaliana. By quantifying the straightness, in addition to the angle relative to the primary stem (Kunita et al., Reference Kunita, Morita, Toda and Higaki2021), we succeeded in capturing the characteristics of the branch shape and distinguishing each myosin XI mutant. The results suggested that MYOSIN XIf and XIk are required for the regulation of branch architecture and growth dynamics. These findings shed light on the molecular mechanisms underlying branch morphogenesis and establish a foundation for exploring the regulatory networks governing shoot development.
The temporal analysis provided critical insights into the dynamics of lateral branch elongation. Lateral organs in shoots and roots extend at a specific angle relative to the gravity direction, known as the gravitropic setpoint angle (GSA) (Digby & Firn, Reference Digby and Firn1995). Wild-type plants exhibited progressive decreases in angles of branches relative to the primary stem as well as to the gravity vector during elongation from ~2 to 6 cm for 3 days. The straightness of wild-type branches consistently decreased over time, reflecting controlled curvature formation (Figure 5e). This is how the characteristic S-shaped curvature with vault-like angle distributions is formed and the GSA is established (Figure 1a, e, i; Figure 5a, b). The S-shaped branches with the GSA likely optimize light capture, reduce mechanical stress by minimizing torque from branch weight, and facilitate seed dispersal from higher positions, potentially increasing reproductive success (Figure 2a).
In contrast, at a broader scale, some branches in the xik and xifk mutants exhibited increasing straightness during elongation (Figure 5g, h). As a result, the characteristic curvature observed in wild-type branches did not develop in the xik and xifk mutants (Figure 5c, d), indicative of impaired curvature regulation. The xik mutant formed branches with drooping morphologies, characterized by a higher angle relative to the primary stem (Figures 2c, e and 3c, e), demonstrating that MYOSIN XIk plays a dominant role in maintaining the upward growth and curvature of lateral branches. Plants sense the direction of gravity by the sedimentation of amyloplasts and subsequent translocation of LAZY1-like proteins from amyloplasts to the plasma membrane (Morita, Reference Morita2010; Nishimura et al., Reference Nishimura, Mori, Shikata, Nakamura, Hashiguchi, Abe, Hagihara, Yoshikawa, Toyota, Higaki and Morita2023). Primary inflorescence stems of the myosin xi1 xi2 xik triple mutant have been reported to exhibit deficient amyloplast sedimentation and gravitropic bending, but the defects are rescued by expressing MYOSIN XIk:YFP (Talts et al., Reference Talts, Ilau, Ojangu, Tanner, Peremyslov, Dolja, Truve and Paves2016). In addition, the amyloplasts exhibit lower motility in the xik endodermal cells than in the wild-type cells (Tsugawa et al., Reference Tsugawa, Miyake, Okamoto, Toyota, Yagi, Morita, Hara-Nishimura, Demura and Ueda2023), probably because of the suppression of cytoplasmic streaming in xik (Avisar et al., Reference Avisar, Abu-Abied, Belausov and Sadot2012; Peremyslov et al., Reference Peremyslov, Prokhnevsky, Avisar and Dolja2008; Ueda et al., Reference Ueda, Yokota, Kutsuna, Shimada, Tamura, Shimmen, Hasazawa, Dolja and Hara-Nishimura2010). Considering these reports, MYOSIN XIk may be required for gravity sensing through the amyloplast dynamics, which promote upward bending of the lateral branches.
Branch morphology was more severely disrupted in the xifk double mutant, with branches exhibiting irregular angle distributions (Figure 2d) and variability in straightness (Figure 3d). Because the primary stems in the xifk double mutant failed to grow straight (Figure 1d, h), lateral branches of the xifk double mutant are subject to mechanical stress differing from those of the wild type and the xif and xik single mutants. However, the angle plot of the lateral branches relative to the gravity vector was similar to that relative to the primary stem (Figure 2d and Supplementary Figure S1D). Simultaneous inactivation of MYOSIN XIk and MYOSIN XIf causes a defect in the organ straightening process required for restoration of a bent shape (Okamoto et al., Reference Okamoto, Ueda, Shimada, Tamura, Kato, Tasaka, Morita and Hara-Nishimura2015). Furthermore, mathematical modeling has revealed that an organ-angle-dependent gravitropic response and an organ-curvature-dependent proprioceptive response are weak in the xifk stem compared with those in the wild-type stem (Tsugawa et al., Reference Tsugawa, Miyake, Okamoto, Toyota, Yagi, Morita, Hara-Nishimura, Demura and Ueda2023). The pronounced angle fluctuations in xifk branches (Figure 4d and Supplementary Figure S3D) further emphasize the involvement of MYOSIN XIf and XIk in coordinating growth direction, particularly under rapid elongation. In summary, the branch morphology is established by MYOSIN XIk-dependent upward bending, and MYOSIN XIf- and XIk-dependent organ straightening, to stabilize growth direction and maintain curvature.
These findings might accord with reports linking myosin XI proteins to intracellular trafficking processes, such as vesicle transport and cytoplasmic streaming, which are essential for maintaining cellular polarity and directional growth (Peremyslov et al., Reference Peremyslov, Prokhnevsky and Dolja2010; Prokhnevsky et al., Reference Prokhnevsky, Avisar and Dolja2008; Ueda et al., Reference Ueda, Yokota, Kutsuna, Shimada, Tamura, Shimmen, Hasazawa, Dolja and Hara-Nishimura2010). The observed phenotypes can be partially attributed to disruptions in actin–myosin interactions, which play critical roles in cellular organization and growth. MYOSIN XIk is known to facilitate vesicle trafficking, including the delivery of cell wall components, which may be necessary for generating the mechanical properties that support branch curvature. The pronounced instability observed in the xifk mutant suggests that MYOSIN XIf may provide additional mechanical support or contribute to sensing environmental cues, such as gravitational stimuli. It is worth exploring the hypothesis that the actin filament bundles function as physical sensors of gravitational stimuli (Blancaflor, Reference Blancaflor2013).
Future studies should focus on unraveling the molecular mechanisms by which MYOSIN XIf and XIk interact with the actin cytoskeleton, and their involvement in vesicle dynamics and phytohormone regulatory systems that govern cell wall metabolism. Investigating other myosin XI isoforms could yield a more comprehensive understanding of their collective contributions to shoot architecture. Moreover, integrating the non-destructive quantitative morphometry system employed in this study with advanced biomechanical approaches may shed light on how motor proteins coordinate the upward growth of lateral branches, providing insights not only from a biological perspective but also from a physical standpoint.
Supplementary material
The supplementary material for this article can be found at http://doi.org/10.1017/qpb.2025.10007.
Data availability statement
The data pertaining to this article will be shared upon reasonable request to the corresponding author.
Acknowledgements
This work was supported by the Japan Society for the Promotion of Science KAKENHI to T.H. (JP18H05492, JP20H03289) and H.U. (JP18H05496 and JP24K09506), and the Hirao Taro Foundation of KONAN GAKUEN for Academic Research to H.U. We thank Ms. Hitomi Okada (Kumamoto University) and Ms. Remi Kawakami (Kumamoto University) for their assistance with plant maintenance. We thank Robert McKenzie, PhD, from Edanz (https://jp.edanz.com/ac) for editing a draft of this manuscript.
Author contributions
T.H. contributed to the study conception and design. Material preparation and data collection and analysis were performed by all authors. The first draft of the manuscript was mainly written by T.H., H.U., and D.Y. All authors read and approved the final manuscript. All authors approved the submitted version.
Funding statement
This work was supported by the Japan Society for the Promotion of Science KAKENHI to T.H. (18H05492, 20H03289) and H.U. (JP18H05496 and JP24K09506), and the Hirao Taro Foundation of KONAN GAKUEN for Academic Research to H.U.
Competing interest
The authors declare no competing interests.
Open peer review
To view the open peer review materials for this article, please visit http://doi.org/10.1017/qpb.2025.10007.








 Open access
Open access
Comments
Prof. Olivier Hamant
Editor-in-Chief
Quantitative Plant Biology
30 November 2024
Dear Prof. Hamant,
Please find attached our manuscript titled “Four-dimensional morphological analysis of lateral branches in Arabidopsis thaliana,” which we would like to be considered for publication as an Original Research Article in Quantitative Plant Biology in the Special Collection on ‘Plant Morphogenesis: Quantitative Aspects and Emerging Novel Concepts.’
Our study investigates the roles of myosin XI motor proteins (MYOSIN XI-F and XI-K) in regulating the morphology of lateral branches in Arabidopsis thaliana. Using a combination of three-dimensional reconstruction and temporal imaging, we developed a comprehensive four-dimensional framework to analyze branch morphology in wild-type and myosin XI mutant plants. Specifically, we introduced and quantified two critical metrics: branch angle and straightness.
Our findings reveal distinct roles for MYOSIN XI-F and XI-K in establishing and maintaining branch morphology. The wild type and the myosin xif mutant formed S-shaped branches, characterized by upward adjustment in branch angles over time. In contrast, the myosin xik mutant formed straight and drooping branches with minimal angle adjustments, whereas myosin xif xik double mutants showed irregular directional changes with fluctuating angles. These results suggest that MYOSIN XI-K is essential for gravitropic response to upward bending, whereas both MYOSIN XI-F and XI-K are required for organ-straightening processes to stabilize growth direction and maintain curvature.
We believe that these findings will resonate with the readership of Quantitative Plant Biology, particularly researchers interested in plant morphogenesis, biomechanics, and quantitative analyses of shoot architecture. Our study provides a novel, integrative framework to analyze lateral branch development, thereby advancing our understanding of how plants coordinate growth and structural stability at the whole-plant level.
We confirm that this manuscript has not been published elsewhere, is not under consideration by another journal, and has been approved by all authors for submission to Quantitative Plant Biology. The authors declare no conflicts of interest.
Please address all correspondence to:
Takumi Higaki
Graduate School of Science and Technology
Kumamoto University, 2-39-1 Kurokami, Chuo-ku
Kumamoto, 860-8555
Japan
Phone: +81-96-342-3975
E-mail: thigaki@kumamoto-u.ac.jp
We look forward to hearing from you at your earliest convenience.
Yours sincerely,
Takumi Higaki, PhD