Refine search
Actions for selected content:
10186 results in Genetics
Population Genetics, Molecular Evolution, and the Neutral Theory. Selected Papers of Motoo Kimura. Edited by Naoyuki Takahata. University of Chicago Press, Chicago. 1994. 686 pages. Price: Cloth US$80.00 £63.95 Paper US$29.95 £23.95. ISBN 0 226 43562 8.
-
- Journal:
- Genetical Research / Volume 66 / Issue 2 / October 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 179-180
-
- Article
-
- You have access
- Export citation
Mitotic mutants of Aspergillus nidulans
-
- Journal:
- Genetical Research / Volume 26 / Issue 3 / December 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 237-254
-
- Article
-
- You have access
- Export citation
The use of maternally coded gene products in Drosophila
-
- Journal:
- Genetical Research / Volume 55 / Issue 2 / April 1990
- Published online by Cambridge University Press:
- 14 April 2009, pp. 87-96
-
- Article
-
- You have access
- Export citation
The parasexual cycle in Verticillium albo-atrum
-
- Journal:
- Genetical Research / Volume 5 / Issue 2 / July 1964
- Published online by Cambridge University Press:
- 14 April 2009, pp. 305-315
-
- Article
-
- You have access
- Export citation
Non-randomness of nucleotide bases in mRNA codons
-
- Journal:
- Genetical Research / Volume 27 / Issue 3 / June 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 353-362
-
- Article
-
- You have access
- Export citation
Cytokines. By M. J. Clemens. Medical Perspectives Series (Editors: A. P. Read and T. Brown). βios Scientific Publishers, Oxford, England. 1991. 122 + x pages. Paperback. £11.95/US$24.00. ISBN 1 872 748 70 8.
-
- Journal:
- Genetical Research / Volume 61 / Issue 2 / April 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 155
-
- Article
-
- You have access
- Export citation
Natural selection and gene substitution*
-
- Journal:
- Genetical Research / Volume 13 / Issue 2 / April 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 127-141
-
- Article
-
- You have access
- Export citation
Variability and restrictions against inbreeding and unequal family size in control populations of Tribolium
-
- Journal:
- Genetical Research / Volume 7 / Issue 1 / February 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 122-129
-
- Article
-
- You have access
- Export citation
A tomato selection experiment
-
- Journal:
- Genetical Research / Volume 2 / Issue 3 / November 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 361-372
-
- Article
-
- You have access
- Export citation
GRH volume 16 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 16 / Issue 1 / August 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
The genetic control of triosephosphate isomerase of hexaploid wheat and other Triticeae species
-
- Journal:
- Genetical Research / Volume 45 / Issue 2 / April 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 127-142
-
- Article
-
- You have access
- Export citation
Books received
-
- Journal:
- Genetical Research / Volume 20 / Issue 3 / December 1972
- Published online by Cambridge University Press:
- 14 April 2009, p. 337
-
- Article
-
- You have access
- Export citation
Mutations in Man. Edited by G. Obe. Berlin: Springer-Verlag. 1984. 327 pages. DM116. U.S. $45.50. ISBN 3 540 13113 2.
-
- Journal:
- Genetical Research / Volume 45 / Issue 3 / June 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 347-348
-
- Article
-
- You have access
- Export citation
The Mus musculus musculus type Y chromosome predominates in Asian house mice
-
- Journal:
- Genetical Research / Volume 50 / Issue 3 / December 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 195-198
-
- Article
-
- You have access
- Export citation
Variability in genetic parameters among small populations
-
- Journal:
- Genetical Research / Volume 29 / Issue 3 / June 1977
- Published online by Cambridge University Press:
- 14 April 2009, pp. 193-213
-
- Article
-
- You have access
- Export citation
-
For a model in which quantitative traits are assumed to be determined solely by additive genes at many loci, formulae are developed for the variance among replicated small populations of size N, maintained without selection, of the additive genetic variance, heritability, genetic correlations and similar parameters. The base population is assumed to be in linkage equilibrium, but it is argued that most of the variation in the within-line additive variance (VAt at generation t) is due to linkage disequilibrium caused by sampling. If
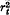 is the squared correlation of gene frequencies averaged over all pairs of loci at time t, the coefficient of variation (CV) of VAt equals
is the squared correlation of gene frequencies averaged over all pairs of loci at time t, the coefficient of variation (CV) of VAt equals 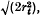 , with similar formulae for other parameters.
, with similar formulae for other parameters.The formulae are evaluated for models of loci distributed uniformly along the chromosome but much of the disequilibrium is due to loci on different chromosomes. For unlinked loci CV(VAt) reaches √4/(3(N)), and for mammalian models, this value is not greatly exceeded. The variance in successive generations has a correlation of at least one-half due to the maintenance of linkage disequilibrium. The magnitude of this variance in parameters and their autocorrelation with time shows that accurate predictions cannot be made about genetic parameters in the base population from single replicate results.
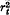 is the squared correlation of gene frequencies averaged over all pairs of loci at time t, the coefficient of variation (CV) of V
is the squared correlation of gene frequencies averaged over all pairs of loci at time t, the coefficient of variation (CV) of V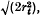 , with similar formulae for other parameters.
, with similar formulae for other parameters.Gregor Mendel: The First Geneticist. By V. Orel. Oxford University Press, 1996. pp. x + 363. Price £29.50, Hardback. ISBN 0 19 8547 74 9
-
- Journal:
- Genetical Research / Volume 68 / Issue 3 / December 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. 264-265
-
- Article
-
- You have access
- Export citation
Inheritance of alleles at ten enzymatic loci of the freshwater snail Stagnicola elodes (Basommatophora: Lymnaeidae)
-
- Journal:
- Genetical Research / Volume 49 / Issue 3 / June 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 201-206
-
- Article
-
- You have access
- Export citation
Non-autonomy in achaete mosaics of Drosophila
-
- Journal:
- Genetical Research / Volume 27 / Issue 1 / February 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-22
-
- Article
-
- You have access
- Export citation
‘Midget’, a new dwarfing gene in the house mouse dependant on a genetic background of small body size for its expression
-
- Journal:
- Genetical Research / Volume 2 / Issue 2 / July 1961
- Published online by Cambridge University Press:
- 14 April 2009, pp. 272-282
-
- Article
-
- You have access
- Export citation
The distribution of transposable elements within and between chromosomes in a population of Drosophila melanogaster. I. Element frequencies and distribution
-
- Journal:
- Genetical Research / Volume 60 / Issue 2 / October 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 103-114
-
- Article
-
- You have access
- Export citation
