Refine search
Actions for selected content:
10186 results in Genetics
Effects of actinomycin D and puromycin upon excystment of Didinium nasutum
-
- Journal:
- Genetical Research / Volume 27 / Issue 2 / April 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 291-296
-
- Article
-
- You have access
- Export citation
Modification of selection limits for egg number
-
- Journal:
- Genetical Research / Volume 3 / Issue 2 / July 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. 210-225
-
- Article
-
- You have access
- Export citation
The recognition of an alien chromosome segment translocated to a wheat chromosome
-
- Journal:
- Genetical Research / Volume 10 / Issue 3 / December 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 295-301
-
- Article
-
- You have access
- Export citation
Genetic analysis of a new haplotype of the histidine decarboxylase gene complex in C57BL/6 mice
-
- Journal:
- Genetical Research / Volume 47 / Issue 2 / April 1986
- Published online by Cambridge University Press:
- 14 April 2009, pp. 131-134
-
- Article
-
- You have access
- Export citation
Crossing over between closely linked markers spanning the centromere of chromosome 3 in Drosophila melanogaster*†
-
- Journal:
- Genetical Research / Volume 26 / Issue 2 / October 1975
- Published online by Cambridge University Press:
- 14 April 2009, pp. 173-185
-
- Article
-
- You have access
- Export citation
A model relating the replication and expression of colicin factor E2-P9
-
- Journal:
- Genetical Research / Volume 20 / Issue 3 / December 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 331-334
-
- Article
-
- You have access
- Export citation
Abstracts of papers presented at the 9th Mammalian Biochemical Genetics Workshop meeting held at the Linnean Society Rooms, Piccadilly, London on 24 and 25 November 1982
-
- Journal:
- Genetical Research / Volume 41 / Issue 3 / June 1983
- Published online by Cambridge University Press:
- 14 April 2009, pp. 303-309
-
- Article
-
- You have access
- Export citation
Evidence for polarity of chromosome replication in F− strains of Escherichia coli
-
- Journal:
- Genetical Research / Volume 8 / Issue 1 / August 1966
- Published online by Cambridge University Press:
- 14 April 2009, pp. 119-124
-
- Article
-
- You have access
- Export citation
Mapping of ochre suppressors in Escherichia coli
-
- Journal:
- Genetical Research / Volume 11 / Issue 1 / February 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. 15-20
-
- Article
-
- You have access
- Export citation
-
Genetic mapping of suppressors of the ilv-188 mutation is described. Suppressors of this mutation have been mapped at four ochre suppressor loci: sup L, sup M, sup N and sup O. The
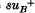 ochre suppressor described by Brenner & Beckwith is shown not to suppress ilv-188 whereas the
ochre suppressor described by Brenner & Beckwith is shown not to suppress ilv-188 whereas the 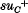 suppressor described by the same authors suppresses ilv-188 and may represent the same locus as supO suppressors. The mapping by P1 transduction of the sup L, sup M and sup N loci is described.
suppressor described by the same authors suppresses ilv-188 and may represent the same locus as supO suppressors. The mapping by P1 transduction of the sup L, sup M and sup N loci is described.
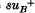 ochre suppressor described by Brenner & Beckwith is shown not to suppress ilv-188 whereas the
ochre suppressor described by Brenner & Beckwith is shown not to suppress ilv-188 whereas the 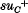 suppressor described by the same authors suppresses ilv-188 and may represent the same locus as supO suppressors. The mapping by P1 transduction of the sup L, sup M and sup N loci is described.
suppressor described by the same authors suppresses ilv-188 and may represent the same locus as supO suppressors. The mapping by P1 transduction of the sup L, sup M and sup N loci is described.Effect of Y chromosome and H-2 complex derived from Japanese wild mouse on sperm morphology
-
- Journal:
- Genetical Research / Volume 53 / Issue 1 / February 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 17-19
-
- Article
-
- You have access
- Export citation
GRH volume 34 issue 2 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 34 / Issue 2 / October 1979
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f4
-
- Article
-
- You have access
- Export citation
GRH volume 68 issue 1 Cover and Front matter
-
- Journal:
- Genetical Research / Volume 68 / Issue 1 / August 1996
- Published online by Cambridge University Press:
- 14 April 2009, pp. f1-f2
-
- Article
-
- You have access
- Export citation
Books Received
-
- Journal:
- Genetical Research / Volume 68 / Issue 3 / December 1996
- Published online by Cambridge University Press:
- 14 April 2009, p. 267
-
- Article
-
- You have access
- Export citation
Phenotypic characterization of R-factor tetracycline resistance determinants
-
- Journal:
- Genetical Research / Volume 24 / Issue 3 / December 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 333-343
-
- Article
-
- You have access
- Export citation
The Genetics of Populations. By Jay L. Lush. Edited by Arthur B. Chapman and Robert R. Shrode, with an addendum by James F. Crow. Iowa State University. 1994. Available from R. Willham, 239 Kildee, Iowa State University, Ames, IA 50011-3150. 900 pages. Price US$45.00. No ISBN number.
-
- Journal:
- Genetical Research / Volume 65 / Issue 1 / February 1995
- Published online by Cambridge University Press:
- 14 April 2009, p. 76
-
- Article
-
- You have access
- Export citation
Optimum selection strategies: studies with Drosophila melanogaster
-
- Journal:
- Genetical Research / Volume 46 / Issue 1 / August 1985
- Published online by Cambridge University Press:
- 14 April 2009, pp. 101-105
-
- Article
-
- You have access
- Export citation
The distribution of transposable elements within and between chromosomes in a population of Drosophila melanogaster. III. Element abundances in heterochromatin
-
- Journal:
- Genetical Research / Volume 64 / Issue 3 / December 1994
- Published online by Cambridge University Press:
- 14 April 2009, pp. 183-197
-
- Article
-
- You have access
- Export citation
Aberrant ascus genotypes from crosses involving mutants at the g locus in Sordaria fimicola
-
- Journal:
- Genetical Research / Volume 24 / Issue 3 / December 1974
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-250
-
- Article
-
- You have access
- Export citation
Y-chromosomal and other factors in the development of testis size in mice
-
- Journal:
- Genetical Research / Volume 50 / Issue 3 / December 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 205-211
-
- Article
-
- You have access
- Export citation
The spread of genes in random mating control populations
-
- Journal:
- Genetical Research / Volume 3 / Issue 1 / February 1962
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-10
-
- Article
-
- You have access
- Export citation
