Refine search
Actions for selected content:
10186 results in Genetics
The lactose system in Klebsiella aerogenes V9A: 1. Characteristics of two Lac mutant phenotypes which revert to wild-type
-
- Journal:
- Genetical Research / Volume 20 / Issue 2 / October 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-191
-
- Article
-
- You have access
- Export citation
The addition of the chromosomes of Aegilops umbellulata to Triticum aestivum (var. Chinese Spring)
-
- Journal:
- Genetical Research / Volume 9 / Issue 1 / February 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 111-114
-
- Article
-
- You have access
- Export citation
Genetic control of DNA specificity in Pseudomonas aeruginosa
-
- Journal:
- Genetical Research / Volume 12 / Issue 1 / August 1968
- Published online by Cambridge University Press:
- 14 April 2009, pp. 99-102
-
- Article
-
- You have access
- Export citation
Our Uncertain Heritage: Genetics and Human Diversity. By L. Hartl Daniel. London: Harper and Row. 1985. 468 pages. £35.75. ISBN 0 06042684 5.
-
- Journal:
- Genetical Research / Volume 47 / Issue 1 / February 1986
- Published online by Cambridge University Press:
- 14 April 2009, p. 81
-
- Article
-
- You have access
- Export citation
GRH volume 29 issue 1 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 29 / Issue 1 / February 1977
- Published online by Cambridge University Press:
- 14 April 2009, p. b1
-
- Article
-
- You have access
- Export citation
Random selective advantages of genes and their probabilities of fixation*
-
- Journal:
- Genetical Research / Volume 21 / Issue 3 / June 1973
- Published online by Cambridge University Press:
- 14 April 2009, pp. 215-219
-
- Article
-
- You have access
- Export citation
-
The question of what is meant by random fluctuations in selection intensities in a finite population is re-examined. The model presented describes the change in the frequency of a gene in a haploid population of size M. It is assumed that in any generation the adaptive values of A and a are equally likely to be 1 + s: 1 or 1: 1 + s. If s is the selective advantage and x the frequency of gene A, then the first two moments of the change in frequency are found to be m(Δx) = x(1 − x)(1 − 2x) θ/2M and
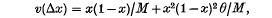
where E(s2) = θ/M. The ultimate probability of fixation is computed, showing that variability in selection increases the chance of fixation of a rare gene. A more general form for m(Δx) also is obtained. This form is compared with the equation currently used in describing random fluctuations in selection intensities.
Genetic aspects of fertility and endocrine organ size in rats*
-
- Journal:
- Genetical Research / Volume 16 / Issue 3 / December 1970
- Published online by Cambridge University Press:
- 14 April 2009, pp. 251-259
-
- Article
-
- You have access
- Export citation
Is subline differentiation a continuing process in inbred strains of mice?
-
- Journal:
- Genetical Research / Volume 19 / Issue 1 / February 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. 53-59
-
- Article
-
- You have access
- Export citation
Partial complementation by murine t haplotypes: deficit of males among t6/tw5 double heterozygotes and correlation with transmission-ratio distortion
-
- Journal:
- Genetical Research / Volume 57 / Issue 1 / February 1991
- Published online by Cambridge University Press:
- 14 April 2009, pp. 55-59
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 54 / Issue 3 / December 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 247-248
-
- Article
-
- You have access
- Export citation
Human Gene Mutation. By David N. Cooper and Michael Krawczak. BIOS Scientific Publishers. 1993. 402 pages. Price £49.50/US $99.00. ISBN 1 8872748 41 4.
-
- Journal:
- Genetical Research / Volume 62 / Issue 2 / October 1993
- Published online by Cambridge University Press:
- 14 April 2009, p. 161
-
- Article
-
- You have access
- Export citation
Damage in DNA of an infecting phage T4 shifts reproduction from asexual to sexual allowing rescue of its genes
-
- Journal:
- Genetical Research / Volume 49 / Issue 3 / June 1987
- Published online by Cambridge University Press:
- 14 April 2009, pp. 183-189
-
- Article
-
- You have access
- Export citation
Index of Subjects and Authors
-
- Journal:
- Genetical Research / Volume 66 / Issue 3 / December 1995
- Published online by Cambridge University Press:
- 14 April 2009, pp. 285-286
-
- Article
-
- You have access
- Export citation
A nuclear gene modifying instability of fertility restoration in cytoplasmic male sterile rice
-
- Journal:
- Genetical Research / Volume 60 / Issue 3 / December 1992
- Published online by Cambridge University Press:
- 14 April 2009, pp. 195-200
-
- Article
-
- You have access
- Export citation
Genome Analysis: A Practical Approach. Edited by K. E. Davies. Oxford: IRL Press Ltd.1988. 192 pages. $27·00/US$54·00. ISBN 1 85221 109 1.
-
- Journal:
- Genetical Research / Volume 53 / Issue 2 / April 1989
- Published online by Cambridge University Press:
- 14 April 2009, pp. 149-150
-
- Article
-
- You have access
- Export citation
Controlling elements in the mouse X-chromosome: I. Interaction with the X-linked genes
-
- Journal:
- Genetical Research / Volume 14 / Issue 3 / December 1969
- Published online by Cambridge University Press:
- 14 April 2009, pp. 223-235
-
- Article
-
- You have access
- Export citation
Books received
-
- Journal:
- Genetical Research / Volume 32 / Issue 3 / November 1978
- Published online by Cambridge University Press:
- 14 April 2009, p. 315
-
- Article
-
- You have access
- Export citation
GRH volume 20 issue 2 Cover and Back matter
-
- Journal:
- Genetical Research / Volume 20 / Issue 2 / October 1972
- Published online by Cambridge University Press:
- 14 April 2009, pp. b1-b4
-
- Article
-
- You have access
- Export citation
Occurrences and linkage relations of the mutant ‘extra-toes’ in the mouse
-
- Journal:
- Genetical Research / Volume 9 / Issue 3 / June 1967
- Published online by Cambridge University Press:
- 14 April 2009, pp. 383-385
-
- Article
-
- You have access
- Export citation
Evidence that HT mutant strains of bacteriophage P22 retain an altered form of substrate specificity in the formation of transducing particles in Salmonella typhimurium
-
- Journal:
- Genetical Research / Volume 27 / Issue 2 / April 1976
- Published online by Cambridge University Press:
- 14 April 2009, pp. 315-322
-
- Article
-
- You have access
- Export citation
