Refine search
Actions for selected content:
25644 results in Astronomy: General Interest
Type Ia supernova sub-classes and progenitor origin
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S357 / October 2019
- Published online by Cambridge University Press:
- 09 October 2020, pp. 1-15
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
Role of environment on AGN activity
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S356 / October 2019
- Published online by Cambridge University Press:
- 29 January 2021, p. 171
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
Galactic Mergers at redshift z ∼ 5, a sample of fast growing QSOs
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S356 / October 2019
- Published online by Cambridge University Press:
- 29 January 2021, p. 291
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
NGC7469 as seen by MEGARA at the GTC
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S356 / October 2019
- Published online by Cambridge University Press:
- 29 January 2021, pp. 306-309
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
Probabilistic evolution of pairs of trans-Neptunian objects in close orbits
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S364 / October 2019
- Published online by Cambridge University Press:
- 30 May 2022, pp. 165-170
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
The role of LoBALs in quasar evolution
-
- Journal:
- Proceedings of the International Astronomical Union / Volume 15 / Issue S356 / October 2019
- Published online by Cambridge University Press:
- 29 January 2021, pp. 285-289
- Print publication:
- October 2019
-
- Article
-
- You have access
- Export citation
Stellar accelerations and the galactic gravitational field
-
- Journal:
- Publications of the Astronomical Society of Australia / Volume 36 / 2019
- Published online by Cambridge University Press:
- 16 September 2019, e038
-
- Article
-
- You have access
- HTML
- Export citation
-
Typical stars in the Milky Way galaxy have velocities of hundreds of kilometres per second and experience gravitational accelerations of
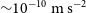 $\sim\!10^{-10}~{\rm m\,s}^{-2}$, resulting in velocity changes of a few centimetres per second over a decade. Measurements of these accelerations would permit direct tests of the applicability of Newtonian dynamics on kiloparsec length scales and could reveal significant small-scale inhomogeneities within the galaxy, as well increasing the sensitivity of measurements of the overall mass distribution of the galaxy. Noting that a reasonable extrapolation of progress in exoplanet hunting spectrographs suggests that centimetre per second level precision will be attainable in the coming decade(s), we explore the possibilities such measurements would create. We consider possible confounding effects, including apparent accelerations induced by stellar motion and reflex velocities from planetary systems, along with possible strategies for their mitigation. If these issues can be satisfactorily addressed, it will be possible to use high-precision measurements of changing stellar velocities to perform a ‘blind search’ for dark matter, make direct tests of theories of non-Newtonian gravitational dynamics, detect local inhomogeneities in the dark matter density, and greatly improve measurements of the overall properties of the galaxy.
$\sim\!10^{-10}~{\rm m\,s}^{-2}$, resulting in velocity changes of a few centimetres per second over a decade. Measurements of these accelerations would permit direct tests of the applicability of Newtonian dynamics on kiloparsec length scales and could reveal significant small-scale inhomogeneities within the galaxy, as well increasing the sensitivity of measurements of the overall mass distribution of the galaxy. Noting that a reasonable extrapolation of progress in exoplanet hunting spectrographs suggests that centimetre per second level precision will be attainable in the coming decade(s), we explore the possibilities such measurements would create. We consider possible confounding effects, including apparent accelerations induced by stellar motion and reflex velocities from planetary systems, along with possible strategies for their mitigation. If these issues can be satisfactorily addressed, it will be possible to use high-precision measurements of changing stellar velocities to perform a ‘blind search’ for dark matter, make direct tests of theories of non-Newtonian gravitational dynamics, detect local inhomogeneities in the dark matter density, and greatly improve measurements of the overall properties of the galaxy.
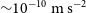
Contents
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp ix-xi
-
- Chapter
- Export citation
2 - Why Life Cannot Be Defined
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 33-62
-
- Chapter
- Export citation
Index
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 243-246
-
- Chapter
- Export citation
1 - The Enduring Legacy of Aristotle: The Battle over Life as Self-Organization or (Genetic-Based) Reproduction
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 8-32
-
- Chapter
- Export citation
Dedication
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp vii-viii
-
- Chapter
- Export citation
5 - Challenges for a Universal Theory of Life
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 105-131
-
- Chapter
- Export citation
Introduction
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 1-7
-
- Chapter
- Export citation
9 - A Shadow Biosphere: Alien Microbes on Earth?
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 195-216
-
- Chapter
- Export citation
Copyright page
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp vi-vi
-
- Chapter
- Export citation
4 - How Scientific Theories Develop
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 82-104
-
- Chapter
- Export citation
7 - Artificial Life: Could ALife Solve the N = 1N=1 Problem?
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 161-171
-
- Chapter
- Export citation
3 - What Is a Scientific Theory?
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 63-81
-
- Chapter
- Export citation
8 - Searching for Extraterrestrial Life Without a Definition or Universal Theory of Life
-
- Book:
- The Quest for a Universal Theory of Life
- Published online:
- 05 September 2019
- Print publication:
- 12 September 2019, pp 172-194
-
- Chapter
- Export citation
