



Archaeologies of colonialism have undergone major shifts in perspectives in the past two decades. One of the most significant developments has been the longitudinal studies of Indigenous persistence (Lightfoot and Gonzalez Reference Lightfoot and Gonzalez2018; Panich and Schneider Reference Panich and Schneider2014; Silliman Reference Silliman, Oland, Hart and Frink2012) and an increasing attention to how introduced species contributed to Indigenous socioeconomic change and independence (Bethke Reference Bethke2017; Fowles et al. Reference Fowles, Arterberry, Montgomery, Atherton, Douglass and Graves2017; Montgomery Reference Montgomery2019). Archaeologists have long acknowledged the profound impacts livestock have had on Spanish colonial ecology and society in North America, with particular attention to ethnogenesis, subsistence, landscape change, and labor (Allen Reference Allen2010; Freiwald and Pugh Reference Freiwald and Pugh2018; Jones Reference Jones, Giovas and LeFebvre2018; Pavao-Zuckerman et al. Reference Pavao-Zuckerman, Mayfield, Copperstone and Thomas Foster2018; Reitz et al. Reference Reitz, Pavao-Zuckerman, Weinand, Duncan and Thomas2010; Sunseri Reference Sunseri2017). The next step in these inquiries is to understand the complicated ways in which Native groups managed livestock within their cultural landscapes in response to multiple waves of colonial demands. The US Southwest region encompasses several Indigenous postcontact adoptions of Eurasian domesticated animals, yet each case reflects tremendous diversity shaped by local knowledge and conditions.
Animal-husbandry—the practices associated with the care, feeding, and breeding of animals—underlies any argument about colonial landscape change and Indigenous labor, and it is the cornerstone for all further interpretation. Species-specific strategies shaped how people used their time and energy and which parts of the landscape were most impacted by grazing. Time allocation reflects cultural priorities, and this is significant when examining native strategies to balance Spanish colonial demands with cultural and subsistence needs.
In the southern Southwest and northwestern Mexico, semiferal animal management should be considered a potential Indigenous adaptive strategy to Spanish colonial intrusions in the Southwest. “Semiferal” describes a state in which animals have minimal contact with humans and are left to graze and breed with little human interference, but they are still intermittently herded and culled. Semiferal practices reflect a lower time investment while also creating a product for colonial markets. To argue for semiferal practices as a strategy of Indigenous persistence, evidence for this type of management must first be identified in the archaeological record.
In this article, I examine the history of animal husbandry practices in southwestern North America and the zooarchaeological study of herding practices in the Spanish colonial period. Although semiferal cattle management is a widely assumed historical practice in western North America, little archaeological work has identified or defined the practice in excavated contexts. I use zooarchaeological and historical evidence as a case study for semiferal management at Mission Los Santos Ángeles de Guevavi in Nogales, Arizona, and I outline evidence for the archaeological identification of semiferal management. Finally, I discuss why animal-husbandry strategies at Spanish missions is pertinent to understanding the erasure of Native American ranching from mainstream narratives of southwestern history.
Feral Livestock in Archaeology
Archaeological work has often surveilled the boundaries of transformations in human–animal relationships, particularly as it relates to domestication and wildness. Although Ingold (Reference Ingold1980) distinguished the economic systems of hunting, pastoralism, and ranching, shifts from wild to domestic uses in some species can be difficult to identify archaeologically and are accompanied by substantial re-wilding and/or ferality. Species such as cattle, horses, goats, reindeer, and pigs shift into feral states easily and survive without human interference (Lazo Reference Lazo1994; Naundrup and Svenning Reference Naundrup and Svenning2015). Genetics, use wear, population structure, osteometrics, stable isotopes, artifacts, and ethnographic evidence have all been brought to bear on the question of whether an animal is wild or domestic (Balasse et al. Reference Balasse, Evin, Tornero, Radu, Fiorillo, Popovici, Andreescu, Dobney, Cucchi and Bălăşescu2016; Levine Reference Levine, Mills and McDonnell2005; Perry and Makarewicz Reference Perry and Makarewicz2019; Zeder Reference Zeder, Harlan and Gepts2012). Previous research on degrees of ferality have focused on the Eurasian origins of livestock and pastoralists (Chen et al. Reference Chen, Hu, Hu, Wang, Ma, Lü and Wang2016; Taylor Reference Taylor2017; Vermeersch et al. Reference Vermeersch, Riehl, Starkovich, Streit and Höflmayer2021). Researchers used multiple, reinforcing lines of evidence—ranging from genetics to the presence of bridles—to help identify domestic livestock from feral and wild counterparts. Much of this research is concentrated in Eurasia and Africa, and the de facto assumption in the Americas is that introduced livestock were in contexts controlled by humans.
Eurasian domesticates have long been identified as a form of ecological colonialism (Crosby Reference Crosby2004), but Native peoples of the Americas adapted and adopted livestock husbandry into their lifeways. On the Plains, Indigenous groups integrated horses derived from Spanish stock into emergent pastoral economies (Bethke Reference Bethke2017; Montgomery Reference Montgomery2019). Southwestern Diné groups began independently adapting their practices around sheepherding early in the contact period (Campbell Reference Campbell2021). After 1650 in New England, groups such as the Natick and Punkapoag used domesticated livestock to fend off colonists’ incursions onto Native lands, and cattle became a survival tool as subsistence became circumscribed by settlers (Silverman Reference Silverman2003). In Peru, animal husbandry existed prior to contact, but Spanish appetites fueled the rapid growth of Eurasian herds and became integrated into Indigenous subsistence strategies (Covey Reference Covey and Staller2021; Kennedy and VanValkenburgh Reference Kennedy and VanValkenburgh2016).
Feral animals can be enormously productive in grassland ecosystems, and this phenomenon has been a significant factor both in colonial economics and the emergence of Indigenous animal husbandry in the Americas. In Argentina, feral cattle hunting and semiferal management helped establish colonial control of the region and provided hides and meat for mining centers (Cushner Reference Cushner1984). In Paraguay, Guaraní hunted cattle in the Río de la Plata region from feral mission herds, and Spanish accounts of this period imply that the cattle and the land were ownerless (Sarreal Reference Sarreal2011). Significantly, not all of these emergent relationships were formalized through active management, and feral herds could be as valuable as carefully managed ones. Loosely managed, semiferal herds were common across the Americas after contact, with wide-ranging economic impacts.
In the greater Southwest, Native labor was used to manage mission herds, and these herds provided seed stock for later presidios and towns occupied by mestizo, European, and Indigenous families (Kessell Reference Kessell2002). The criollo cattle breed typical of the Spanish period is known for its drought tolerance and its ability to both give birth without human aid and defend itself from predators (Anderson et al. Reference Anderson, Estell, Gonzalez and Cibils2015). As part of this process, however, Native peoples across Arizona, New Mexico, Texas, and California became adept at raising and processing livestock (Allen Reference Allen1998; Pavao-Zuckerman Reference Pavao-Zuckerman2008; Tarcan Reference Tarcan2005).
Toward a Zooarchaeology of Semiferal Animal Husbandry
Zooarchaeological analysis has been well equipped to look at human livestock management in the past. The identification of semiferal management practices in the archaeological record benefits from a mixed methodology using a variety of proxies. Zooarchaeological and isotopic analyses, including others such as histology and aDNA, are part of a suite of tools at researchers’ disposal. Culling profiles helps link human behavior and economies to material remains and elucidate patterns of dairying, wool, and meat procurement strategies (Helmer et al. Reference Helmer, Gourichon and Vila2007; Lyman Reference Lyman2008). Culling profiles of semiferal animal husbandry, however, would not appear distinct from meat-focused culling, with adult animals primarily being culled. Identification of semiferal animal husbandry therefore requires additional lines of evidence.
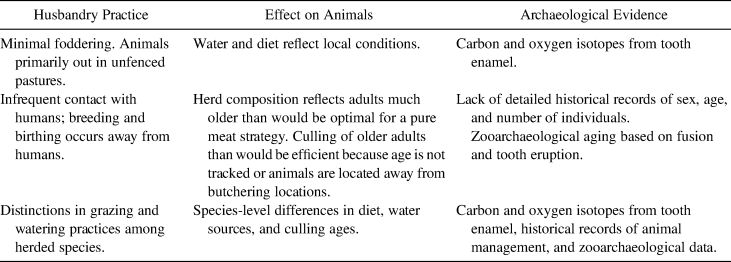
Archaeologically, what would free-range, semiferal cattle management look like? Semiferal ranching would be characterized by less surveillance and minimal interaction between the animals and people (Table 1). The continuum of human–animal relationships ranges from constant companionship to passing interactions, with many possibilities in between. Today, animals have tags, and animal managers track age, sex, lineage, and health individually, although the animals spend the most time unsupervised on the range. In the colonial period, without a detailed record and tagging system, it would have been difficult to monitor the exact ages of the animals. Pastoralists, whether reindeer herders, sheepherders, or dairy farmers, develop deep familiarity with individual animals without tagging systems. Minimal interaction makes it potentially difficult to access the animal when it has a reached its full size and weight, and a greater number of older animals would be culled than expected in a purely meat procurement profile. Also, feral animals’ diets will reflect the surrounding area and follow seasonal changes, and foddering will not be a major contribution to their diet. In terms of record keeping, lack of contact would hinder accurate tracking of herd size. Together, each facet of semiferal husbandry can be identified archaeologically.
Table 1. Semiferal Practices Identifiable in the Archaeological and Historical Records.
Case Study: Mission Los Santos Ángeles de Guevavi
A few miles northeast of Nogales, Arizona—10 km (6 miles) north of what is today the modern US-Mexican border, are the ruins of Mission Los Santos Ángeles de Guevavi. Mission Guevavi was located in the northern Pimería Alta of New Spain. Today, the National Park Service (NPS) administers the site. Guevavi belonged to the mission system founded by the Jesuit Father Eusebio Kino after 1691, with the intent of converting Tohono O'odham ancestors to Catholicism and Spanish ways of life. In 1768, following the Jesuit Expulsion, the Franciscan Order took over the mission network. Franciscan missionaries replaced the Jesuits, but their presence at the mission was short and plagued by raiding. At Guevavi, however, the Jesuit period is preserved because the mission was emptied by 1775, and it moved north. Excavations at Guevavi (Figure 1) provided an opportunity to study the first 70 years of colonization and the introduction of animal husbandry to the Pimería Alta.
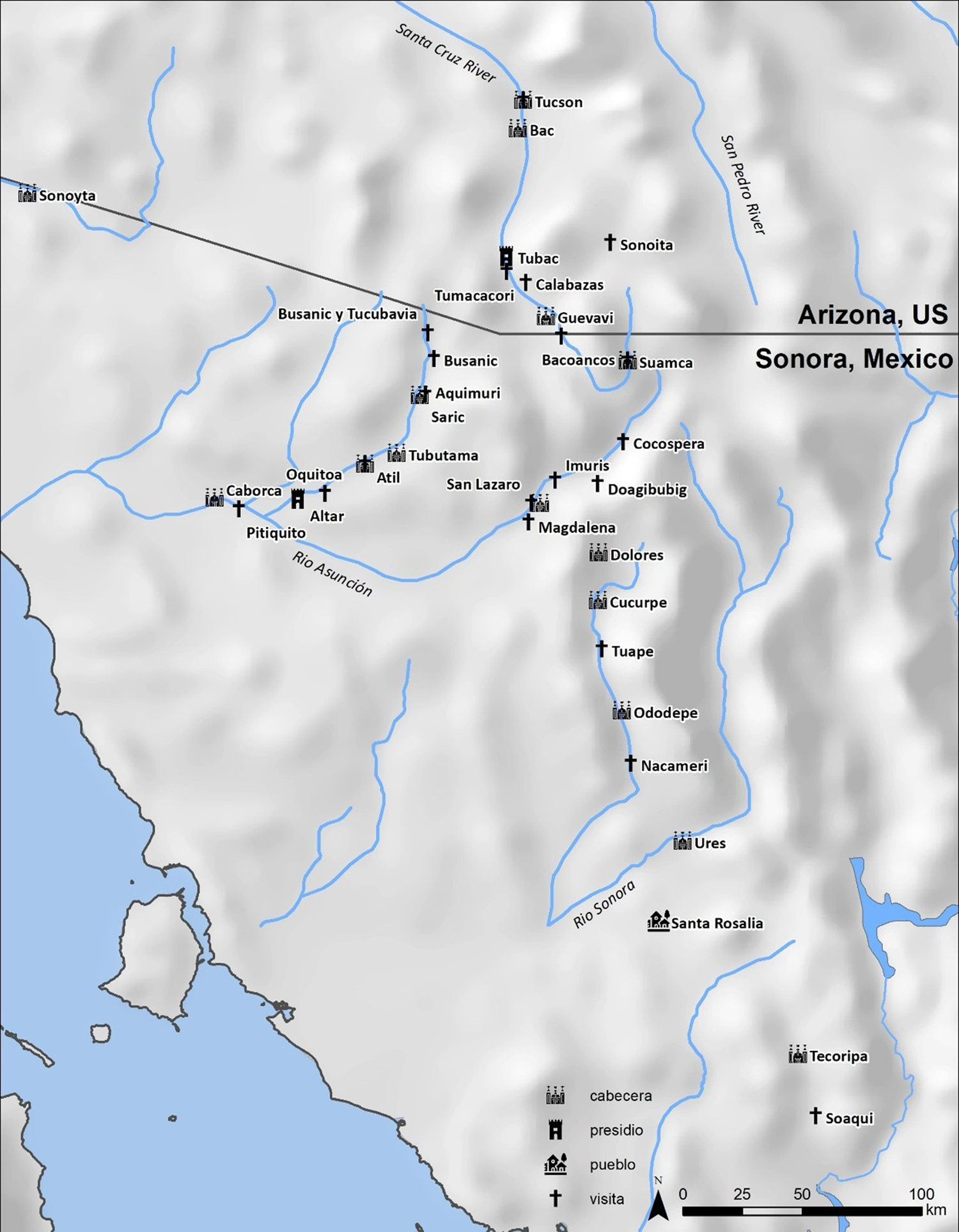
Figure 1. Map of the Pimería Alta mission systems. Map by Kathryn MacFarland.
In 2013, the Guevavi mission midden was disturbed by rodent activity and was targeted by the NPS for archaeological mitigation. The midden deposit there offered a unique opportunity to explore the early years of missionization prior to the maturation of the colonial economies a century later. The feature that produced the vertebrate samples reported here was excavated as part of the 2013 Mission Guevavi Archaeological Field School, a collaborative effort between the University of Arizona, Arizona State Museum, Tumacácori National Historic Park, and Desert Inc. The excavation analyses are reported in Thiel and Pavao-Zuckerman (Reference Thiel and Pavao-Zuckerman2016). The present faunal materials came from two mission-period 2 × 2 m units, with smaller, later reoccupation by Yaqui miners in the 1810s, several decades after the mission's abandonment but still within the Spanish colonial period. This later occupation contributed little to faunal materials analyzed here, because the bulk of the animal remains were found alongside mission-period artifacts and below the levels where the later mining slag accumulated (Supplemental Text 1). Severe rodent disturbance to the units mixed the older mission-period levels with upper levels, and it was impossible to extricate this smaller portion from the earlier materials, so faunal materials are conservatively interpreted as Spanish colonial from 1700 to 1810. The bulk of the occupation likely dates to the 1720s–1775. Mission Guevavi fauna represents the largest analyzed colonial-period sample in the region to date.
Tohono O'odham Subsistence Practices
Prior to contact, Tohono O'odham groups moved seasonally, hunted, and planted crops in various regions along the floodplain of the Santa Cruz River, and many of those practices endured through the Spanish colonial period and beyond (Fish et al. Reference Fish and Fish2008; Hill Reference Hill2018; McIntyre and Arizona Historical Society Reference McIntyre and Historical Society2008). Prior to and following contact, Tohono O'odham farmers adapted to variable summer rainfall by cultivating maize, squash, and beans and collecting wild resources such as goosefoot, cholla, prickly pear, saguaro fruit, agave, yucca, and mesquite pods (Fontana and Schaefer Reference Fontana and Schaefer1989; Weber et al. Reference Weber, Ariffin, Nabhan, Idouraine and Kohlhepp1996). Tohono O'odham made use of local game such as deer, bighorn sheep, pronghorn, rabbits, and rodents (Dean Reference Dean2003; Diehl and Waters Reference Diehl, Waters, Kennett and Winterhalder2006).
The Tohono O'odham integrated animal husbandry into their subsistence strategies and belief systems following the Spanish colonial period, allowing cattle to go feral and hunting them (Kozak and Lopez Reference Kozak and Lopez1999; Marak and Tuennerman Reference Marak and Tuennerman2013:109). Tohono O'odham, compared to Akimel O'odham to the north, integrated ranching into their lifeways earlier because of mission contact and had their own herds during and after the Spanish colonial period. The farming Akimel O'odham communities were not missionized, but they were not untouched by Spanish colonialism in the south. By the nineteenth century, Akimel O'odham began ranching, and cattle (and sheep to a lesser extent) were a major part of the faunal assemblage found at the late nineteenth-century site of Blackwater Village (Loendorf et al. Reference Loendorf, Fertelmes, DeJong, Kyle Woodson and Lewis2019). During the historical period, Tohono and Akimel O'odham images of branded cattle and brands appeared in rock art (Bostwick and Krocek Reference Bostwick and Krocek2016), indicating integration of the animals into other cultural facets.
Semiferal cattle ranching was a primary economic activity for the Tohono O'odham, and grazing lands were often held in common—a practice that dated to the mission period—until the 1930s Depression, when the US government ordered the creation of grazing districts, thereby seriously disrupting traditional grazing and water rights (Parman Reference Parman1992). Cattle, in particular, became central to local and Indigenous cuisine in the form of dried meat, stews such as chile colorado, and tallow. Today, beef is considered part of family cooking by members of the Tohono O'odham Nation (Fazzino Reference Fazzino2008), and ranching remains a major economic activity on the reservation.
Methods
Faunal remains in this study came from the historical midden (Feature 26) associated with Mission Guevavi on land administered by Tumacácori National Historic Park. The collection is currently housed at the National Park Service Western Archeological Conservation Center in Tucson, Arizona. The specimens reported were sieved through a 6.35 mm (¼″) screen. Zooarchaeological analysis followed the methods described in Reitz and Wing (Reference Reitz and Wing2008) and are originally reported in Mathwich (Reference Mathwich2016). Contextual and additional excavation information about Feature 26 is available in Supplemental Text 1.
Faunal Analysis Methods
Vertebrate remains from the Mission Guevavi Field School were identified by the author using standard zooarchaeological methods (Reitz and Wing Reference Reitz and Wing2008). Identifications were made using the comparative skeletal collections housed at the Stanley J. Olsen Laboratory of Zooarchaeology, Arizona State Museum, University of Arizona.
A variety of observations, such as element and taxonomic identification, which can be replicated by later investigators, were collected as part of the analysis (Reitz and Wing Reference Reitz and Wing2008). Specimens were identified to taxon, element represented, portion recovered, and symmetry (left vs. right). Indicators for sex, age at death, and modifications were noted where observed, and these were recorded. Age ranges resulted from observations of epiphyseal fusion and tooth eruption above the gumline. For age, the presence of wear is significant because it means a tooth fully erupted.
In this article, “caprine” is the term used to describe the remains from animals that could be domestic sheep or goat. During the Spanish colonial period, however, sheep were the most common small domestic bovid (Kessell Reference Kessell1970; Officer Reference Officer1987). Goats were present, but in far smaller numbers. Mission inventories of the region in 1768 record 10,069 sheep and 251 goats, or 2.4% of all small livestock (Estado del Rectorado de la Pimería 1767). In recognition of this possibility, this article employs a more conservative category of “caprine,” but these are mostly likely sheep.
The Minimum Number of Individuals (MNI) is reported in this study, and this was derived from Minimum Number of Elements (MNE). MNE was calculated by taking into consideration element portions, side, size, and fusion; teeth were excluded from MNE. There were occasions where the lower taxonomic category was smaller than the MNI for the corresponding higher taxonomic level. MNI for the lower taxonomic category was reported in parentheses but was not included in subsequent calculations. MNI increases with the smaller, discrete proveniences used for analysis, resulting in problems of aggregation and accuracy (Domínguez-Rodrigo Reference Domínguez-Rodrigo2012; Grayson Reference Grayson1978; Lyman Reference Lyman2019), but MNI are valuable when looking at the degree of fragmentation. Fragmentation can be indicative of disposal patterns, marrow extraction, and grease extraction (Outram Reference Outram2001). The Guevavi assemblage and contemporary Pimería Alta assemblages reflect substantial fragmentation associated with grease rendering (Pavao-Zuckerman Reference Pavao-Zuckerman2011). For these reasons, MNI was reported, but it was contextualized with Number of Identified Specimens (NISP) and biomass based on the allometric relationships reported in Reitz and Wing (Reference Reitz and Wing2008) in order to balance MNI's overestimation of smaller taxa.
Isotopic Assays
Isotopic assays from tooth enamel carbonate were analyzed to obtain information about animal diet. Tooth enamel carbon is used to examine dietary proportions of plants over the tooth's development. Oxygen isotopes from tooth carbonate help researchers understand the types of water sources an animal consumes. Other isotopes, such as nitrogen and strontium, have been valuable in elucidating animal management practices and movement of animal products (Guiry et al. Reference Guiry, Harpley, Jones and Smith2014, Reference Guiry, Noël and Fowler2021). The carbon and oxygen originated from tooth enamel bioapatite rather than collagen proteins. Carbon and oxygen were chosen specifically for their ability to shed light on vegetation communities and environmental water sources (Cerling et al. Reference Cerling, Harris and Passey2003; Passey et al. Reference Passey, Robinson, Ayliffe, Cerling, Sponheimer, Denise Dearing, Roeder and Ehleringer2005). The methods were originally reported in Mathwich and colleagues (Reference Mathwich, Pavão-Zuckerman and Ruff2019), and they are summarized here. Samples of 1–3 mg of enamel were drilled from the tooth, maintaining the overall physical integrity and identifiability. Teeth were sampled using a small drill, producing a shallow, vertical trench bulk sample in the outer enamel of the tooth crown, which reflected the average consumption over the tooth's development. Bovid molar crowns take about a year to form (Hillson Reference Hillson2005). Only adult molars were used in this study.
Oxygen and carbon isotopes were extracted from the CO3 component of biological apatite in the tooth enamel and calculated using the offsets from Kohn and Cerling (Reference Kohn and Cerling2002:465). All δ13C and δ18O samples were processed in the Stable Isotope Laboratory Carbonate Lab on the campus of the University of Arizona. Powder samples were pretreated to isolate the structural carbonate-apatite and to remove organics and secondary carbonate. A weak solution of 0.1 Μ acetic acid in distilled water was added to the sample and soaked overnight, and then removed with a pipette. Although it is no longer recommended (Pellegrini and Snoeck Reference Pellegrini and Snoeck2016), this aggressive pretreatment was standard practice at the Environmental Isotope Laboratory when the analysis was conducted in 2015. Samples were then rinsed with ultra-pure water and allowed to dry overnight. The δ18O and δ13C of carbonates were measured using an automated carbonate preparation device (KIEL III) coupled to a gas-ratio mass spectrometer (Finnigan MAT 252). Powdered samples were reacted with dehydrated phosphoric acid under vacuum at 70°C. The isotope ratio measurement was calibrated based on repeated measurements of NBS-19 and NBS-18, and precision is ±0.1‰ for δ18O and ±0.08‰ for δ13C (1σ).
The fractionation for bovids reported in Passey et alia (Reference Passey, Robinson, Ayliffe, Cerling, Sponheimer, Denise Dearing, Roeder and Ehleringer2005) that was used to calculate δ13C enamel-diet was αenamel-diet = 1.0146. Note that the δ13C values have been corrected for modern fossil-fuel consumption (1.5‰; Friedli et al. Reference Friedli, Lötscher, Oeschger, Siegenthaler and Stauffer1986; Leuenberger et al. Reference Leuenberger, Siegenthaler and Langway1992; Tieszen and Fagre Reference Tieszen and Fagre1993; Treydte et al. Reference Treydte, Frank, Saurer, Helle, Schleser and Esper2009). The composition of stable carbon is reported in δ (per mil) notation relative to the global Vienna Pee Dee Belemnite (VPDB) standard. Rsample and Rstandard refer to the ratios of 13C/12C and, as a sample or standard, they are reported relative to VPDB standard (Kendall and McDonnell Reference Kendall and McDonnell1998). The C3 endmember used to calculate the percentage of C4 in the animal diets was derived from Kohn (Reference Kohn2010), and the modern C4 endmember of −12.5‰ VPDB was from Cerling and colleagues (Reference Cerling, Harris and Passey2003). For the equations, voltages, standard deviations, endmembers, and data used, see Supplemental Table 1.
Historical Sources
The historical data presented here were translated into English from the original Spanish by Kessell (Reference Kessell1970) and the author in consultation with the Office of Ethnohistoric Research at the Arizona State Museum, Tucson. These documents were written by Jesuit priests sponsored by the Spanish colonial government, and they represent counts of herd sizes at the missions. These documents, which represent a missionary perspective, are concerned with the material holdings and property of the missions, and they were not meant to be demonstrative of livestock management. The author's use of them for livestock management is her own reading of the documents.
Results
The results of the above analyses were used to evaluate the presence of semiferal management of cattle and to contrast it to the management of sheep at Mission Guevavi. Several patterns emerged within these results relating to differences between the species. First, the isotopic ratios of tooth enamel from cattle and caprines indicate differences in both water and grazing sources. Second, the faunal ages at death for sheep were younger than three years, whereas those of cattle appear to be older than three years. Third, the historical sources demonstrate different conceptual groupings of domesticated species but offer sparse information about the actual management of the animals.
Stable Isotope Analysis
From 180 inventoried whole teeth, a total of 13 specimens were selected for sampling and were part of a larger study consisting of adult lower molars (Mathwich et al. Reference Mathwich, Pavão-Zuckerman and Ruff2019). Mission Guevavi was unique in its large sample size of teeth compared to other sites, and it offered the most individual samples. With that note, the sample was small. Because the samples came from a larger midden feature, the project sampled higher than the reported MNIs, which excluded teeth. Caprines had a higher MNI but a lower number of adult tooth samples due to the high frequency of deciduous teeth in the assemblage. The sampled specimens were selected for their completeness, tooth position, and age. The %C4 plants in the diet for cattle ranged from 75% to 104% of the animals’ diets, and δ18Obioapatite ranged from −4.36‰ to −0.44‰ ± 0.10 VPDB (Figure 2). The %C4 plants in the diet for caprine diets ranged from 21% to 52%, and the δ18Obioapatite ranged from 1.34‰ to 3.31‰ ± 0.10 VPDB. Mathwich and colleagues (Reference Mathwich, Pavão-Zuckerman and Ruff2019) and Grimstead and Pavao-Zuckerman (Reference Grimstead and Pavao-Zuckerman2016) found both caprine and cattle oxygen body-water values to be highly enriched compared to average precipitation. Independent sampling of modern water sources and leaf enrichment found that the likeliest explanation was evaporated, enriched water sources stemming from water storage. At Guevavi, the caprine water sources appeared to have been more enriched than cattle water sources. The Guevavi data are consistent with contemporary Pimería Alta sites, indicating that these proportions denote a broader dietary difference among cattle and caprines (Grimstead and Pavao-Zuckerman Reference Grimstead and Pavao-Zuckerman2016; Mathwich et al. Reference Mathwich, Pavão-Zuckerman and Ruff2019). Cattle ate primarily C4 plants, which reflects a dependence on semidesert grasslands—the largest concentrations of perennial C4 grasses in southern Arizona—particularly grama species (Bock and Bock Reference Bock and Bock1986). The high proportion of C4 plants in cattle diets is compatible with the consumption of CAM plants such as cacti. However, studies on range behavior indicate that grasses are their preferred grazing food (Hanselka and Paschal Reference Hanselka and Paschal1990). CAM plant consumption may muddy the differences among taxa, but large distinctions in diets are still observable.
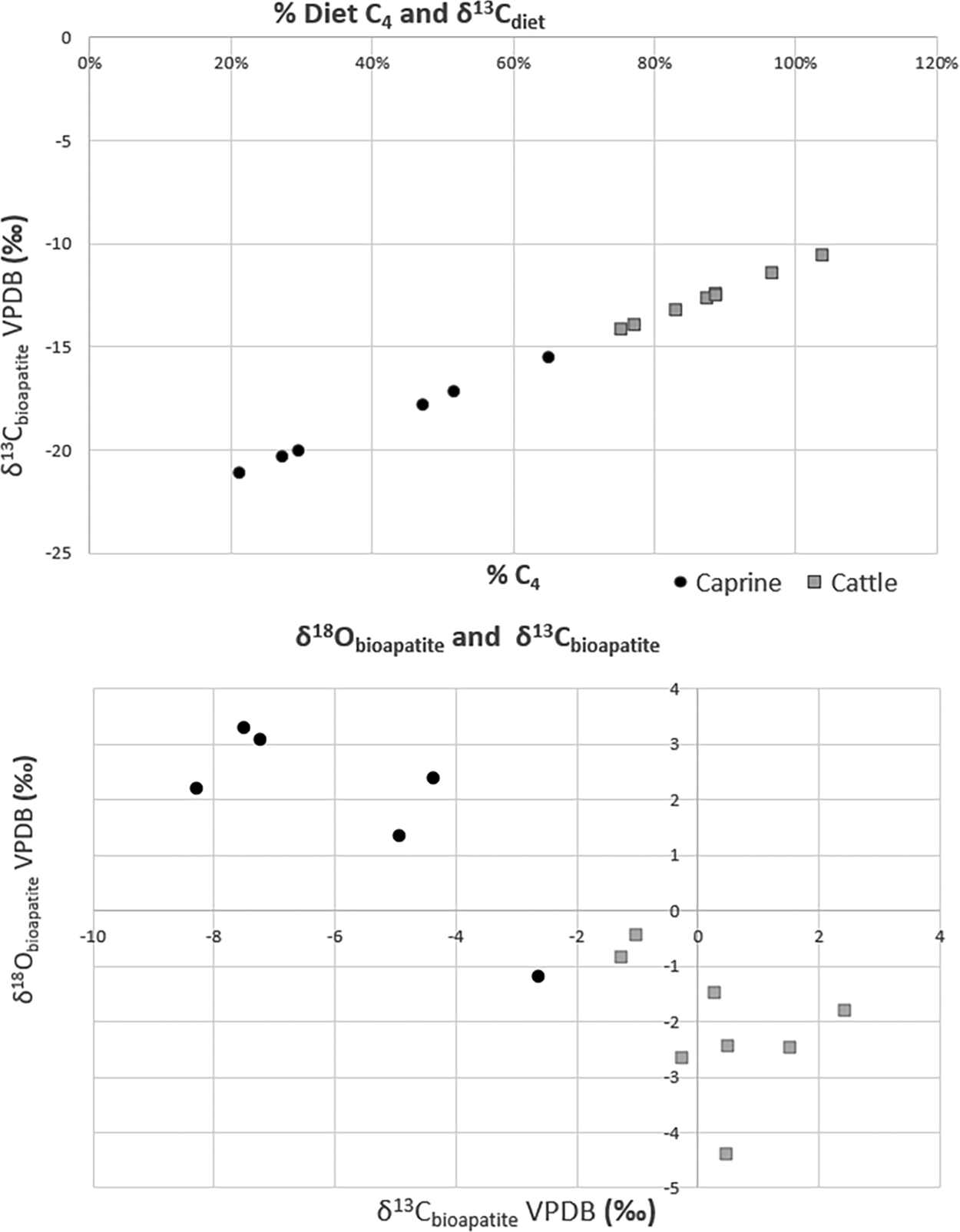
Figure 2. Scatter plot of δ13Cdiet and the percent of C4 in the animals’ diets, and δ13Cbioapatite and δ18Obioapatite from bulk samples of cattle and caprine teeth by site and taxa.
Age Estimation
In cattle, there are three main groups of epiphyseal fusion ages roughly within the following stages of the animal's life: early (7–20 months), middle (24–36 months), and late (42–48 months; Barone Reference Barone1976; Silver Reference Silver, Brothwell and Higgs1969). These broad categories capture a range of epiphyseal events, which vary individually due to nutrition, hormones, health, and geography. Modern epiphyseal fusion rates (Moran and O'Connor Reference Moran and O'Connor1994; Popkin et al. Reference Popkin, Baker, Worley, Payne and Hammon2012; Silver Reference Silver, Brothwell and Higgs1969) were used for age estimations of the domesticated ungulates.
Total MNI of cattle was at least five individuals, and there were at least eight caprines identified (Table 2). Of the cattle elements, 63% (17) of the late-fusing (36+ months) elements were fused, and 85% (17) of middle-fusing elements (24+ months) were fused. At least one younger individual was represented, but most of the individuals were full-grown adults, three to nine years old (Table 3). Among the caprines, 43% (25) of early fusing elements (3–16 months) were fused, but of the middle- and late-fusing elements (7–60 months), 79% (45) were unfused. Individuals ranged from three to four years old, with one older than four years. Sheep/goat were generally slaughtered at a younger age than cattle.
Table 2. Mission Guevavi, Feature 26: Summary of Domesticated Ungulates, 6.35 mm (¼″) Screened Materials.

Table 3. Mission Guevavi, Feature 26: Epiphyseal Fusion.
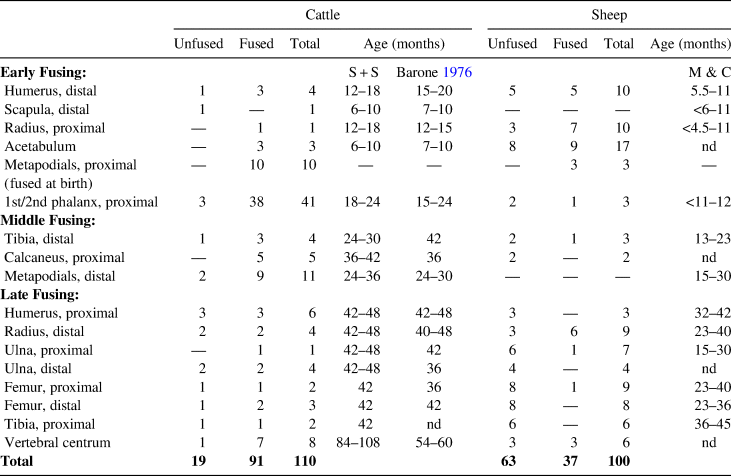
Sources: From Reitz and Wing (Reference Reitz and Wing2008:72) and derived from Schmid (Reference Schmid1972:75) and Silver (Reference Silver, Brothwell and Higgs1969) (S+S). Sheep fusion age estimates from Moran and O'Connor (Reference Moran and O'Connor1994) (M & C).
Note: Fusion and growth stages for both cattle and sheep come from studies cited above of growth rates of modern sheep and cattle populations.
A total of seven mandibles with teeth in situ were identified, but the sample size was not large enough to look at population-level tooth wear. Cattle mandibles with in situ teeth all possessed developed M3 teeth and showed a considerable degree of in-wear, putting age estimates beyond 36 months (Hillson Reference Hillson2005). Caprine age estimates from tooth eruption of four left mandibles ranged from one to two years old at death (Moran and O'Connor Reference Moran and O'Connor1994). Once erupted, the M3 becomes a less reliable indicator of age without population-level wear patterns, but generally, Guevavi cattle were older than three years at death. The caprines trended younger, with a single unerupted M3 present among the mandibles found at the site, and two well-worn dP4 yet to be shed.
Historical Sources
Livestock featured prominently in the earliest Spanish accounts of the Pimería Alta and were integral to the establishment of mission settlements, which served as a vanguard for later military and secular settlements. Mission founder Eusebio Kino recorded rough quantities of animals he brought with him to found new missions, but he wrote little about the management of the animals (KinoReference Kino and Bolton1919). Later inventories at Mission Guevavi between 1737 and 1767 illustrated the ontology of livestock from this period (Table 4). Initially, cattle numbers exceeded sheep, but later, sheep were most numerous. Categories of sheep and horses reflect discrimination of age and sex, but over the course of 30 years, no efforts were made to determine the general counts of age and sex in cattle herds.
Table 4. Livestock Counts from Mission Guevavi Entregas from 1737, 1761, and 1767 Inventory.

Notes: The above information is excerpted from Kessell (Reference Kessell1970:195–201) and was translated by Kessell from the documents housed in the W. B. Stephens Collection, University of Texas Library. The information from Estado del Rectorado de la Pimería (1767) was translated by the author with the assistance of the Office of Ethnographic Research, Arizona State Museum.
The surviving entregas, or reports, from Guevavi indicate that in a period of 17 years, the number of cattle more than tripled, and the number of sheep increased sharply (Table 4). The presence of other domesticate ungulates—such as mules, horses, and goats—was noted, but goats do not appear in the later entrega. Interestingly, chickens and pigs do not appear in this record, but they were found in the Feature 26 midden. Contemporary reports of chicken coops appear in other documents (Kessell Reference Kessell1970:88). Bladders of tallow and fat were reported in the entregas as well, indicating that some amount of grease extraction likely occurred at the site (Mathwich and Pavao-Zuckerman Reference Mathwich and Pavao-Zuckerman2018; Pavao-Zuckerman Reference Pavao-Zuckerman2011).
Bone fusion from the Feature 26 midden and categories from the 1761 entrega suggest that sheep age was better monitored, with four subdivisions based on age and sex compared to a single category for cattle (Kessell Reference Kessell1970:195). In 1761, 75% of sheep were under a year old (Table 3). The entrega was written in May, after many sheep gave birth, and the fusion from the midden suggests that they were killed before they were 24 months old, which would likely correspond to the “year-old” lamb category in the 1761 entrega. Its author, Father Gerstner, may have sought to distinguish the lambs born in that spring from those born in the previous year. In contrast, age seems less important to categorizing cattle in both entregas. The 1767 “State of the Rectorate of the Pimería” (Estado del Rectorado de la Pimeria 1767) shows similar categorizations. The 1767 report was a formal inventory of mission holdings and wealth following the Jesuit expulsion to prepare for the new Franciscan administration, and it did not note the sex or age of cattle. Absence of evidence is not evidence of absence, but in consideration with the isotopic dietary information and the fusion ages, the absence of sex and age suggests that the missionaries and Indigenous herders at Mission Guevavi took a different approach toward cattle compared to other livestock. A variety of herd management practices is possible based on these age estimations, and when reviewed alongside other lines of evidence, they help to triangulate possible management strategies.
Discussion
In the following section, I will examine various possible interpretations and fit them within the broader context of Spanish colonial settlements in the Santa Cruz River Valley. The evidence presented here points to distinct taxa management strategies, but the different purposes these animals served within the Mission Guevavi went beyond the needs of cattle in contrast to sheep. Finally, I discuss how these interpretations intersect within contemporary perceptions of Indigenous ranching.
Distinct Roles of Livestock in Mission Economies
Management differences between sheep and cattle are evident in these data, but do these data rise to the level of semiferal ranching for either taxon? The isotopic ratios at Guevavi and other mission and fort sites throughout the region show differences between both water sources and plant diets among cattle and caprines. The biotic communities in the Santa Cruz River Valley are composed of lowland Sonoran and Chihuahuan desert scrub (mixed C4, C3, and CAM), riparian woodland galleries (C3 shrubs and trees), and semidesert grasslands (predominately C4; Biedenbender et al. Reference Biedenbender, McClaran, Quade and Weltz2004; Brown Reference Brown1994; Dimmitt Reference Dimmitt, Phillips and Comus2000). If cattle were semiferal and ranged more widely, it might follow that their C4 should be lower because they would have been semiferal and had more access to different vegetation communities. The higher C4 proportions may then be indicative of foddering of C4 crops such as maize, which is a known practice in Sonora. Range researchers’ observations of Arizona cattle note that although nutritionally flexible when necessary, cattle prefer grasses over shrubs whenever available (Sprinkle et al. Reference Sprinkle, Grumbles and Meen2002:2). Foddering from mission fields of wheat (C3) and maize (C4) is a possibility, but more equal C3:C4 proportions would be expected if that were the case. The high C4 cattle diet, however, is consistent with animal behavior of herds grazing on preferred grasses. Caprines had the higher quantities of C3 plants in their diets and consumed water from more evaporated, stored water sources. These C3 plants could be from either riparian vegetation or higher-elevation vegetation, but the highly enriched water sources imply that they were kept closer to riparian areas where herd managers created water storage features subject to evaporation and oxygen enrichment. The relationship of the animal's δ18Obody-water to leaf enrichment, seasonal precipitation, and water storage is discussed in detail in Mathwich et alia (Reference Mathwich, Pavão-Zuckerman and Ruff2019). For the purposes of this article, when comparing the isotopic data to other lines of evidence, the δ13Cdiet of cattle is most supportive of management away from occupation areas in river valleys. The results are consistent with semiferal cattle management and distinct from contemporary caprines.
At Mission Guevavi, faunal evidence suggests that the mission ranchers used a mixed strategy with respect to caprines, encompassing wool, hide, and meat production. Global archaeological assemblages show an association of culling young male livestock with dairying practices, resulting in high proportions of neonate bone and older females (Greenfield and Arnold Reference Greenfield and Arnold2015; Helmer et al. Reference Helmer, Gourichon and Vila2007; Marciniak Reference Marciniak2011). Dairying may have been present at Guevavi, but it was likely minimal. The knowledge of cheese making and dairying was known in colonial Sonora, primarily with cattle and goats (Segesser and Classen Reference Segesser and Classen2012:78). Dairy is not a strong explanation for the younger caprine ages because the individuals at Guevavi died after being weaned. At Guevavi, the young sheep were killed within their first two years of life, long after being weaned two months after birth. Wool was another labor-intensive animal product produced at missions. Sheep reach breeding age at around 5–12 months, and the relatively young age at death is more consistent with meat production than dairying or wool production strategies. These ages, however, could still encompass a first shearing, which—depending on the breed—happens about once a year (Opatik Reference Opatik2008). Tooth and fusion ages were consistent with at least one shearing before death and the animals being at full adult weight. Although there were no elements diagnostic to sex among these specimens, it is likely that these younger individuals were male because most sheepherders prefer to keep higher ratios of ewes to rams (Keeling Reference Keeling2001:231).
In contrast, the older fusions in cattle suggest looser monitoring typical of semiferal strategies. Free-range, semiferal cattle require less monitoring, but scattered grazing makes it harder to find and harvest the animals at maturity. Cattle herd composition during the mission period was likely a mix of ages, and the fusion data point to mature, full-weight adults chosen for culling and processing. These behaviors are consistent with both semiferal management and a meat and growth-focused approach toward mission cattle herds.
The missionaries’ goal was to grow herds for sale at secular settlements and create starter herds for other missions, and keeping cattle alive longer increased breeding opportunities. Historical documents from the region show consistent growth of cattle populations throughout the eighteenth century (Jackson Reference Jackson1982; Sayre Reference Sayre1999). Herd growth accelerated in later years under Franciscan management, as financial need grew following the Spanish government's curtailment of mission support (Mathwich and Pavao-Zuckerman Reference Mathwich and Pavao-Zuckerman2018). Recording individual ages for large herds required effort and labor, especially in herds of several thousand spread out across desert grasslands. If the herd sizes appeared to be growing with little intervention, there was equally little reason for close monitoring.
Colonial sheep production has been well documented (Tarcan Reference Tarcan2005; Trigg Reference Trigg2004), but its role in the Pimería Alta has yet to be fully explored, despite documentary and archaeological evidence that substantial amounts of Tohono O'odham labor went into it. Although sheepherding has fallen out of favor in modern Arizona and Sonora, it was important for decades, and it had its origins in the colonial period. Sheep herds grew large at Guevavi, surpassing cattle in later periods, and they were also numerous at the neighboring presidio, Tubac (Hewitt Reference Hewitt, Shenk, Teague and Hewitt1975). Other researchers, however, suggest that the increase in sheep in later periods was a response to raiding pressures (Merrill Reference Merrill, Pease and Taylor1994; Pavão-Zuckerman and Martínez-Ramírez Reference Pavão-Zuckerman and Martínez-Ramírez2019). Raiding during the mid-1700s devastated mission residents and led to shortages of livestock throughout the region and to the eventual abandonment of Mission Guevavi. In 1775, the mission holdings and its residents were moved and consolidated with Mission San Cayetano de Tumacácori (Kessell Reference Kessell1970). Historical and archaeological evidence shows a consistent presence of sheep in nearly equal ratios to cattle (Mathwich and Pavao-Zuckerman Reference Mathwich and Pavao-Zuckerman2018). Sheepherding primarily as a mitigation strategy for raiding, however, downplays the importance of wool production in the Pimería Alta economy and for reproducing Spanish culture in the Sonoran Desert. Wool in the Spanish borderlands was an important commodity as a trade item and as clothing. Semiferal management of cattle would complement a more labor- and resource-intensive strategy toward sheep, and the two strategies could be adapted in response to raiding pressure.
The role cattle played in the broader colonial economy has been well documented. Tallow and hide production from cattle have been identified archaeologically at Mission San Agustin 97 km (60 miles) north of Guevavi and at Mission Cocóspera 72 km (45 miles) to the south, and it is likely that hide and tallow production for mining was incorporated into Guevavi's economy (Pavao-Zuckerman Reference Pavao-Zuckerman2011). It is not clear, however, whether the wool was for local consumption or regional trade. The Parral silver-mining district in southern Chihuahua was known to import wool from New Mexico, but trade often did not cross the Sierra Madre Occidental (West Reference West1949). Further historical and archaeological research will be required to establish the markets of Mission Guevavi's sheep products.
Semiferal management had the potential to reduce local resource conflicts among farmers, ranchers, and raiders. Cattle possess a much higher biomass than sheep and require more water and range (Gillespie Reference Gillespie2004), increasing their potential for conflict with Tohono O'odham farming and water requirements. Sheep herds grew large in the later mission period, and it is not clear from historical evidence or the archaeological data if herd managers shifted them to grasslands when the herd sizes became larger. Although the sample size was quite small, the isotopic data point to dietary strategies that differed with respect to sheep and cattle, suggesting that grasslands were the preferred grazing locations for cattle. Allowing these animals to range farther from riparian areas and irrigated farms and into semidesert grasslands decreased resource conflicts over water and grazing, with agriculture in arable river valleys. Raiders on horseback frequently stole presidio and mission herds, and by keeping less labor-intensive livestock away from permanent settlements, raider violence around larger villages might have been reduced. There were many good reasons to let cattle wander and keep them in pastures farther from fields and homes.
Past Labor, Contemporary Marginalization
Enormous amounts of Tohono O'odham time, energy, and skill went into the care of mission herds and the production of animal products. Isotopic ratios, age at death, and historical sources suggest that this labor was part of a mixed strategy built around the behaviors and needs of introduced livestock. Missionaries and Spanish colonists used Native labor under coercive and colonial systems, yet these animals fed mission inhabitants and their family members and Native raiding groups, as well as Spanish colonists and soldiers. Other documentary evidence shows continued historical collection of wild foods such as cactus fruits and mesquite in addition to irrigated agriculture of Native and introduced cultigens (Fontana and Schaefer Reference Fontana and Schaefer1989; Mathwich Reference Mathwich2018; Rea Reference Rea1997; Weber et al. Reference Weber, Ariffin, Nabhan, Idouraine and Kohlhepp1996). This mixed pastoral strategy and the semiferal management of cattle should be understood within the broader context of seasonal subsistence patterns and demands on Tohono O'odham time. A low investment toward cattle may have enabled mission inhabitants to set aside more time for social activities, farming, and gathering. Alternatively, the distance of mission cattle ranches from the primary mission settlement and fields minimized conflicts with the large bovids’ water needs and Tohono O'odham agriculture. Beyond economic considerations, cattle and horses became an important part of twentieth-century Tohono O'odham culture, and herd management was successfully practiced for decades alongside farming until Tohono O'odham groups were pushed from their ancestral water sources (Bahr et al. Reference Bahr, Gregorio, Lopez and Alvarez1974; Ezell Reference Ezell1974; Fontana and Schaefer Reference Fontana and Schaefer1989). The zooarchaeological evidence at Guevavi points to the pragmatism and strategy that mission inhabitants employed when balancing out raiding pressures and colonial demands on their time, lands, and water.
Semiferal animal husbandry is currently out of favor in the United States, although it was once widespread (Jordan-Bychkov Reference Jordan-Bychkov1993). An increasingly urbanized American West meant that the open, unfenced land—typical of the time when livestock were introduced in North America—became more and more enclosed (Charnley et al. Reference Charnley, Sheridan and Nabhan2014). In the nineteenth century, Native American groups were pushed off their land for commercial ranching (Specht Reference Specht2019). Semiferal practices in the Southwest have not disappeared, but they have become integrated into contemporary industrial food systems. In southern Arizona, the reorganization of animal production for distant markets took the form of commercial cattle ranching and became entangled in a suite of settler-colonialist strategies to “Americanize” the Tohono O'odham in the twentieth century. As archaeologists gain better knowledge of Spanish and Native American range strategies, there needs to be serious consideration of how past animal husbandry is characterized within the historical marginalization of Native American ranching. Herding and ranching are significant cultural and economic activities for Native Americans across the western United States and have been since domestic livestock entered Indigenous landscapes, especially in the Southwest (Campbell Reference Campbell2021; Kozak and Lopez Reference Kozak and Lopez1999; Mitchell Reference Mitchell2015; Nabhan et al. Reference Nabhan, Hodgson and Fellows1989; Weisiger Reference Weisiger2011).
In historical narratives, livestock are viewed primarily as a European introduction, and historians of ranching in the United States frequently discount colonial animal husbandry as negligible to the immense Anglo cattle drives that followed (Christiansen Reference Christiansen1988; Curtin et al. Reference Curtin, Sayre and Lane2002; Sayre Reference Sayre1999). This is a jarring contrast to the historical records, archaeology, and oral histories that demonstrate the breadth and longevity of Indigenous animal husbandry. As archaeologists investigate the dynamics of colonial societies, they must be attentive to political and economic ramifications of disconnecting contemporary Native American ranching from animal-husbandry practices that appeared post-contact. Zooarchaeology has the potential to contribute physical evidence of the longevity and capability of Indigenous herders who adapted to the new resource that livestock provided within the constraints of colonial and Indigenous resource use.
Conclusion
This article examined the creation of range practices at Spanish colonial Mission Los Santos Ángeles de Guevavi, near what is today Nogales, Arizona. Using multiple lines of evidence, this article proposed a set of indicators to identify semiferal ranching in both the archaeological and historical records. Isotopic evidence from caprine and cattle teeth showed that semidesert grasslands were grazed by both cattle and sheep, but cattle likely grazed them most frequently. Epiphyseal fusion data and historical analyses suggest that cattle ages were loosely monitored compared to those of sheep. Cattle were culled at older ages than optimal for meat and grease extraction, suggestive of a low-investment management strategy. Less direct labor investment in cattle may have helped Tohono O'odham maintain social, agricultural, and gathering practices, which persisted into the historical period. Finally, semiferal cattle ranching was widespread in historical North America, but it has since become marginalized in contemporary food production. Mainstream US culture often disconnects and minimizes Native contributions to ranching cultures in the western United States, but zooarchaeology investigations into Indigenous animal husbandry in North America have the opportunity to reveal physical evidence of the practice's time depth and the expertise of Indigenous ranchers.
Acknowledgments
This work was funded by Arizona State Museum (ASM) and the Research Corporation for Scientific Advancement Cottrell Scholars program. I thank ASM, University of Arizona School of Anthropology, and Tumacácori National Historic Park for use of their collections. I am grateful to Barnet Pavao-Zuckerman and J. Homer Thiel for their work and guidance at Mission Guevavi. I acknowledge the many University of Arizona students who participated in the field school and cataloged the materials from Mission Guevavi. I am indebted to Katie MacFarland and to my pandemic Zoom writing group.
Data Availability Statement
The author confirms that the data supporting the findings of this study are available within the article and its supplementary material.
Supplemental Material
For supplemental material accompanying this article, visit https://doi.org/10.1017/aaq.2022.46.
Supplemental Text 1. Excavation and contextual information on the Guevavi midden feature.
Supplemental Table 1. Bulk isotopic assays from Mission Guevavi livestock teeth.
Competing Interests
The author confirms that there are no relevant financial or non-financial competing interests to report.









 Open access
Open access