


Introduction
The treatment of fishing in the afterlife amongst Pleistocene maritime cultures has received little attention in the archaeological literature. This is probably because burials of this age are rare in coastal contexts, with most Pleistocene coastal sites having been submerged by sea-level rise following the end of the Last Glacial phase. Here, we report on an assemblage of shell fish-hooks recovered from a burial in Tron Bon Lei rockshelter on Alor Island, Indonesia, dated to the Terminal Pleistocene. These are the oldest-known fish-hooks associated with mortuary practices from anywhere in the world, and perhaps indicate that fishing equipment was viewed as essential for the transition to the afterlife in the Wallacean Islands, where terrestrial fauna is depauperate, and where fish and other marine resources were the mainstay of the diet.
The majority of early prehistoric fish-hooks from around the world derive from habitation contexts. The oldest example comes from Japan, where a single-piece, rotating hook was recently found on Okinawa Island in a stratigraphic layer bracketed by AMS dates of c. 23000 cal BP and 20000 cal BP (Fujita et al. Reference Fujita, Yamasaki, Katagiri, Oshiro, Sano, Kurozumi, Sugawara, Kunikita, Matsuzaki, Kano, Okumura, Sone, Fujita, Kobayashi, Naruse, Kondo, Matsu'ura, Suwa and Kaifu2016). Single-piece, shell fish-hooks have been found in Timor-Leste, where one from Lene Hara Cave has been directly AMS dated to c. 11000 cal BP (O'Connor et al. Reference O'Connor, Ono and Clarkson2011). Another shell hook from Jerimalai Shelter in Timor-Leste is of a comparable age to the Okinawa find, in a deposit context dated to between c. 23000 and 16000 cal BP (O'Connor et al. Reference O'Connor, Ono and Clarkson2011). Single-piece hooks of bone and ivory appear in Europe in the Final Palaeolithic about 12000 years ago, becoming more common in the Mesolithic (Gramsch et al. Reference Gramsch, Beran, Hanik and Sommer2013). On Cedros Island, Mexico, hooks appear approximately 11000 years ago (Des Lauriers Reference Des Lauriers2010), and are present in coastal Chile by about 8000 cal BP (Llagostera Reference Llagostera1992). In coastal California, shell hooks were not used until the Late Holocene, but bone gorges appear by c. 10000 cal BP (Rick et al. Reference Rick, Vellanoweth, Erlandson and Kennett2002).
Prior to the finds described here, the earliest hooks with a mortuary association were the composite slate shaft pieces from the Ershi cemetery in Siberia, in a riverine environment and dated to the late Mesolithic c. 9000 cal BP (Basaliiski Reference Basaliiski, Weber, Katzenberg and Schurr2010). In a maritime context, the earliest burials with fish-hooks are from Oman, where rotating hooks made of pearl shell (Pinctada radiata) have been dated to c. 6000 cal BP from their depositional context (Santini Reference Santini1987).
A preliminary archaeological investigation conducted in June 2014 at Tron Bon Lei shelter (pit B), on the south coast of Alor (Figure 1), uncovered a partial human skeleton in a layer dating to the Terminal Pleistocene, containing five shell fish-hooks and a perforated bivalve (Figure 2). Direct dating of one of the hooks and radiocarbon dates on charcoal associated with the burial demonstrates that the body and grave goods were buried shortly after c. 12000 years cal BP (Table 1). Here we describe the shell fish-hooks, the context of the burial and the profoundly significant behavioural patterns that they represent within the context of Asia-Pacific prehistory and global technological adaptations more widely.
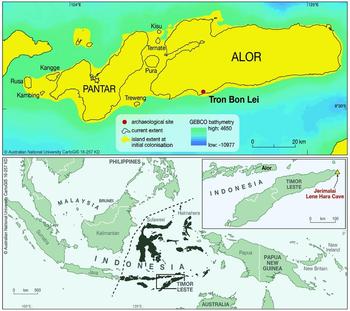
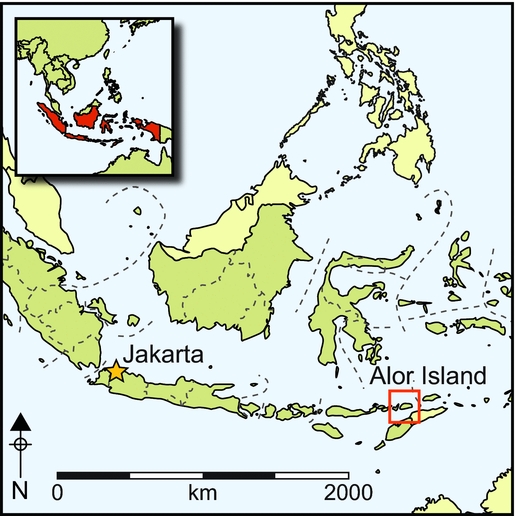
Figure 1. Map (upper) showing the location of Tron Bon Lei, the current extent of Alor, Pantar and other neighbouring islands, and the island as it would have been at 22000 cal BP. The lower map shows Wallace's Line (black dashed line) and the location of the Wallacean Islands in black; the inset shows Timor-Leste and the location of Jerimalai and Lene Hara Cave.

Figure 2. Tron Bon Lei cranium, photographed during excavation. Fish-hook A and perforated bivalve to the north of the mandible. (Photograph by Sofía Samper Carro.)
Archaeological context
Alor is part of the Lesser Sunda chain of islands in eastern Indonesia. Largely volcanic in origin, Alor has a mountainous interior, dropping steeply to a narrow coastal margin. The island has a land area of about 2100km2. As one of the islands of Wallacea, Alor has never been connected by a landbridge to Sunda (the enlarged southernmost extension of Eurasia), Sahul (Australia, New Guinea and the Aru Islands), or to the larger islands of Timor or Flores. During the lower sea stand of the Last Glacial Maximum about 21000 years ago, when sea levels were about 130m below those of the present, it merged with neighbouring Pantar, Pura, Kambing, Rusa, Ternate and Treweng Islands, forming a ‘mega island’ of about 3800km2 (based on an average uplift rate of approximately 0.5m/ka) (Figure 1, upper map). It is at about this time that the first evidence for occupation in Tron Bon Lei occurs. The Tron Bon Lei pit B shelter is one of several rockshelters sharing its name with a volcanic ridge above the village of Lerabain (Figure 3A–B). The pit B shelter is about 33m above sea level and 160m inland from the current seashore (Figure 4A–B).
Table 1. AMS radiocarbon dates from Tron Bon Lei pit B, Alor. Dates calibrated using OxCal v.4.2.4 against IntCal13 and Marine13 (Bronk Ramsey Reference Bronk Ramsey2009; Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013).
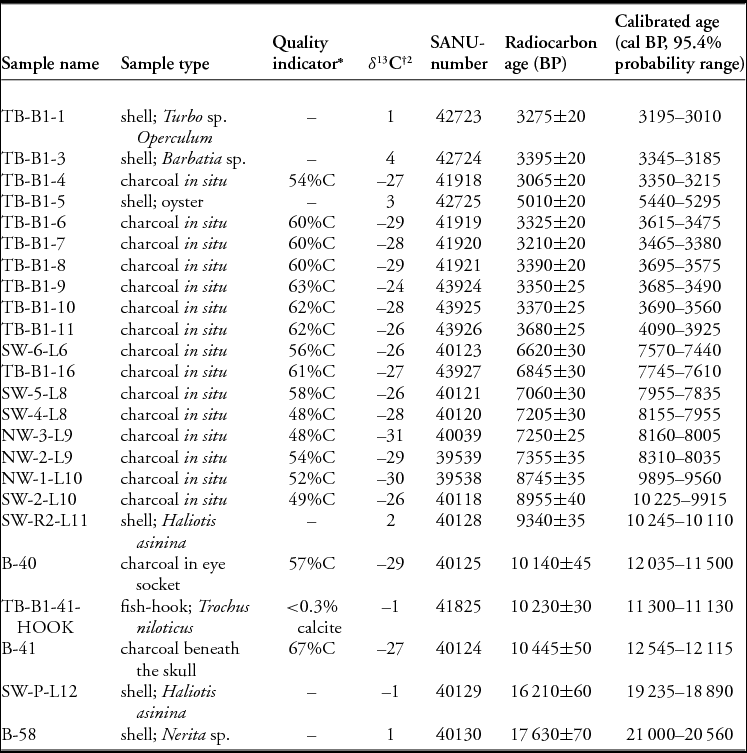
Radiocarbon dates associated with the burial. Charcoal dates have been calibrated against IntCal13, and the shell date against Marine13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013) in OxCal v.4.2 (Ramsey Reference Bronk Ramsey2009).
*Charcoal should contain more than approximately 50%C. %C was measured volumetrically during cryogenic collection and purification. If calcite was present in the aragonitic nacre from the shell hook, recrystallisation and thus contamination with groundwater carbonate may have occurred. %calcite was determined by X-ray diffraction.
†δ13C was measured by AMS and used in date calculation. It is not comparable to δ13C values measured by isotope-ratio mass spectrometry.

Figure 3. A) Northern view of the volcanic ridge where Tron Bon Lei is located. The white arrow indicates the general location of the rockshelters (photograph by permission of Putu Yuda); B) pit B looking east from the southern edge of the rockshelter (image from Samper Carro et al. Reference Samper Carro, O'Connor, Louys, Hawkins and Mahirta2016, reproduced with permission from Elsevier).
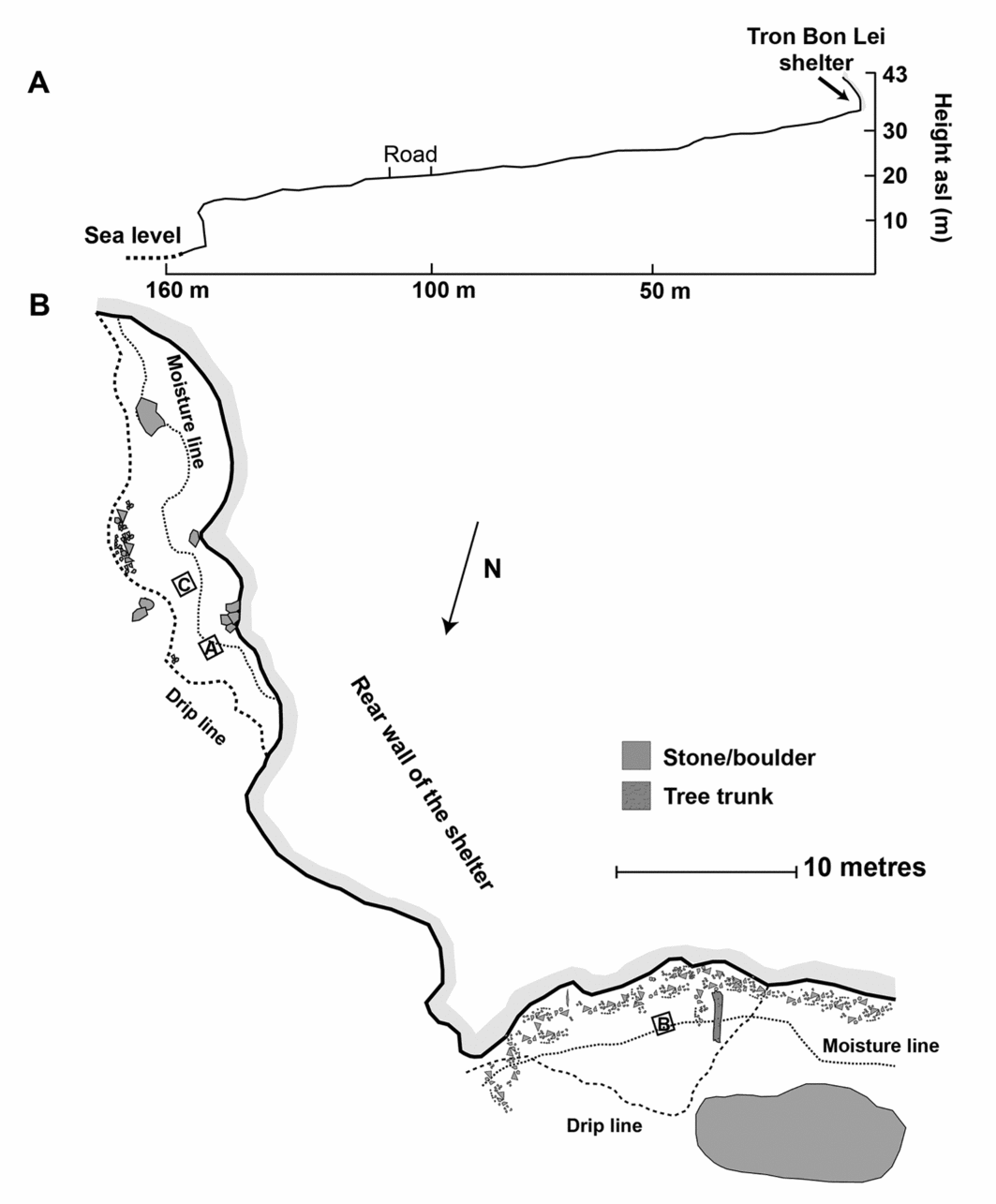
Figure 4. A) Transect showing the distance from sea level to Tron Bon Lei rockshelter; B) plan of Tron Bon Lei showing the location of test pit B (and also test pits A and C, not reported here); modified from Samper Carro et al. (Reference Samper Carro, O'Connor, Louys, Hawkins and Mahirta2016), reproduced with permission from Elsevier.
An exploratory 1m2 test pit (pit B) was placed inside of the dripline and excavated to an average depth of 3.2m, where bedrock was encountered (Figures 3B & 4B). Excavation employed arbitrary 50mm excavation units within stratigraphic layers. Thirteen stratigraphic layers were identified and have been described elsewhere (Figure 5) (Samper Carro et al. Reference Samper Carro, O'Connor, Louys, Hawkins and Mahirta2016). Radiocarbon dating suggests three main phases of activity at the site (Table 1): 1) Late Holocene (c. 4000–3000 cal BP); 2) Terminal Pleistocene to Early Holocene (c. 12000–7500 cal BP); 3) late Pleistocene (c. 21000–18000 cal BP). While there appears to be a chronological hiatus between each of these occupation phases, there is neither evidence of sterile deposits separating them, nor of sedimentary unconformities or lag deposits indicative of their erosion or removal.
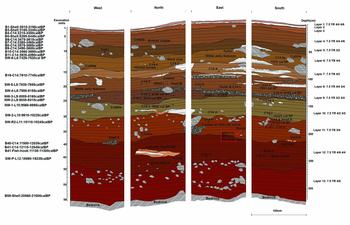
Figure 5. Stratigraphic drawing of the east section of the Tron Bon Lei pit B excavation, showing stratigraphic levels, excavation units, the location of samples taken for radiocarbon dates and the location of the burial and associated finds (red rectangle).
The Tron Bon Lei assemblage
The upper excavation units dating to the Late Holocene contained a small number of earthenware sherds. Shellfish, faunal remains and stone artefacts manufactured from basalt, obsidian and chert were found throughout the sequence, but were less abundant in the pottery-bearing layers. The faunal assemblage consists almost entirely of marine resources including fish, marine molluscs, urchins and crustacea and a small quantity of marine turtle bone. Pit B produced 28745 fish bones (number of identified specimens (NISP)), which accounted for 96.84 per cent of identified bone (Samper Carro et al. Reference Samper Carro, O'Connor, Louys, Hawkins and Mahirta2016). The non-fish bone comprises marine turtle (Cheloniidae), fruit bat (Pteropodidae), squamate lizard (Lacertilia), murid (Muridae), shrew (Soricidae), snake (Serpentes) and bird (Aves), with a total NISP of 939, but at 3.16 per cent of the assemblage, their contribution to subsistence was minimal, and many of the small murids, shrews and reptiles may actually be derived from the activities of roosting barn owls (Hawkins et al. Reference Hawkins, Samper Carro, Louys and O'Connor2017).
Analysis of the fish bones has shown that carnivorous fish taxa dominate in the layers containing the burial, and that an increase in smaller herbivorous/omnivorous reef fish occurred during the Mid Holocene as the sea level rose and the coral reef was established (Samper Carro et al. Reference Samper Carro, O'Connor, Louys, Hawkins and Mahirta2016; see too the online supplementary material (OSM)).
The Tron Bon Lei burial and grave goods
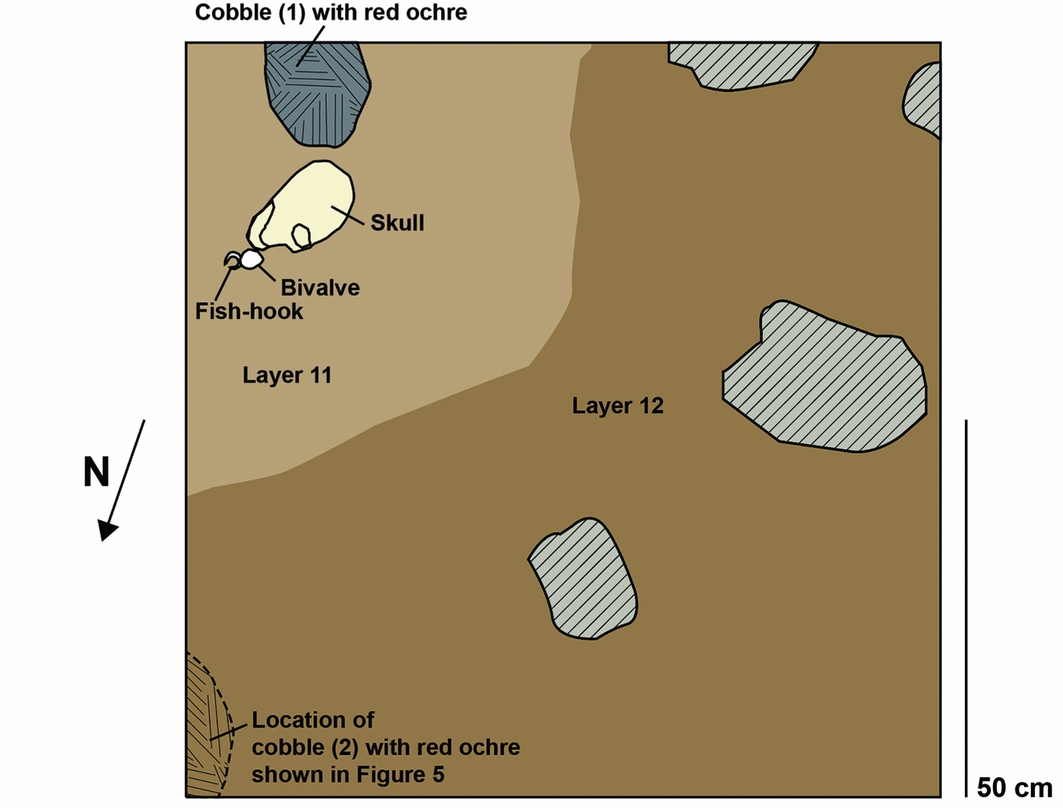
The burial is in the south-east area of pit B (Figures 5–6). The frontal part of the skull began to emerge in excavation unit 39 (around a depth of 2.1m) and continued into excavation unit 41 (reaching a depth of 2.26m in the south-east corner) (Figures 2, 5 & 6). The frontal bone and facial area of the skull were found facing north and resting on the left zygomatic. The cranium, mandible and maxilla, some fragments of cervical vertebrae, rib fragments, pieces of both scapulae and the diaphysis of a left humerus were also recovered. The remainder of the burial extends into the east wall of the pit. The recovered bones were in anatomical association with one another, but as most of the skeleton remains unexcavated, it is not possible to tell if it is a primary or secondary burial. Our focus here is on the burial context rather than the skeletal analyses, which will be presented elsewhere.

Figure 6. Plan view of the Tron Bon Lei cranium showing the position of fish-hook A, bivalve and cobbles with red ochre. Cobble 2 is higher in the profile than cobble 1 and is not in the plan; its location is, however, shown in Figure 6. The burial is in the lighter sediment layer 11. The darker sediment in the northern and western part of the pit is layer 12.
As we do not have the full skeleton, it is not possible to sex the individual with confidence. Despite this, the presence of a relatively gracile skull possessing fully erupted third molars with moderate wear, a weakly developed superciliary ridge, lack of gonial eversion and reduced nuchal lines and mastoid processes are suggestive of an adult female. The direction of error makes it less probable that females will be misclassified, particularly in individuals that are not of advanced age such as this one (Meindl et al. Reference Meindl, Lovejoy, Mensforth and Carlos1985). As the excavation could not be extended in the 2014 season, and there was a danger that the site would be disturbed by local villagers after we left, the decision was made to remove the skeletal remains in pit B and leave the unexcavated bones in situ for future excavation.
Stratigraphic context of the burial and grave goods
Based on depth below the surface, the burial and grave goods are interred in layer 12; they were, however, confined in a pocket of sediment of the same colour and texture as layer 11. It is thought that the burial was dug into the top of layer 12 from layer 11. Thus the burial is thought actually to belong to layer 11 (Figures 5–6). The burial, and the cultural material associated with it were excavated separately from the darker sediments in plan at the same depth (layer 12), and designated as excavation units 39, 40 and 41 in burial context layer 11 to differentiate them from the excavation units with the same number in layer 12. The body was interred fairly shallowly and associated with two flat cobbles that were coated in red ochre. One cobble was interred at the same level, and about 30mm from the back of the skull (Figure 6, cobble 1), while the other was in the north-east corner of the pit, approximately 100–150mm higher than the top of the skull (Figures 5–6, cobble 2). The radiocarbon dates support the assignment of the burial to layer 11, confirming a shallow grave cut.
AMS dating of the Tron Bon Lei burial context and grave goods
A %N test (Brock et al. Reference Brock, Higham and Ramsey2010, Reference Bearéz, Gay and Lunniss2012) of bone from the skull indicated that insufficient collagen was preserved for radiocarbon dating. Instead, samples of charcoal found near the burial and one of the associated fish-hooks (hook C) were dated. Charcoal dates have been calibrated against IntCal13, and the shell date against Marine13 (Reimer et al. Reference Reimer, Bard, Bayliss, Beck, Blackwell, Ramsey, Buck, Cheng, Edwards, Friedrich, Grootes, Guilderson, Haflidason, Hajdas, Hatté, Heaton, Hoffman, Hogg, Hughen, Kaiser, Kromer, Manning, Niu, Reimer, Richards, Scott, Southon, Staff, Turney and van der Plicht2013) in OxCal v.4.2 (Bronk Ramsey Reference Bronk Ramsey2009). A regional average ΔR of 32±41 14C years was applied to the shell date (see OSM).
The upper part of the layer that the hooks were buried within (layer 11) has an AMS radiocarbon age of c. 10000 cal BP (S-ANU-40128: 9340±35 BP, 9935–10245 cal BP at 95.4%) (Table 1). An AMS radiocarbon date of 11310–11075 cal BP (S-ANU-41825: 10230±30 BP) on hook C is consistent with a date on a fragment of charcoal from within the left eye socket of the individual of 12035–11500 cal BP (S-ANU-40125: 10140±45 BP), when taking into consideration that the charcoal age may be affected by an old wood effect (see OSM). Charcoal from directly beneath the skull in excavation unit 41 was dated to 10445±50 BP (S-ANU 40124), with a calibrated age of 12545–12115 cal BP (95.4%). The skeleton was therefore interred in the millennia after 12000 cal BP.
The Tron Bon Lei fish-hooks and bivalve
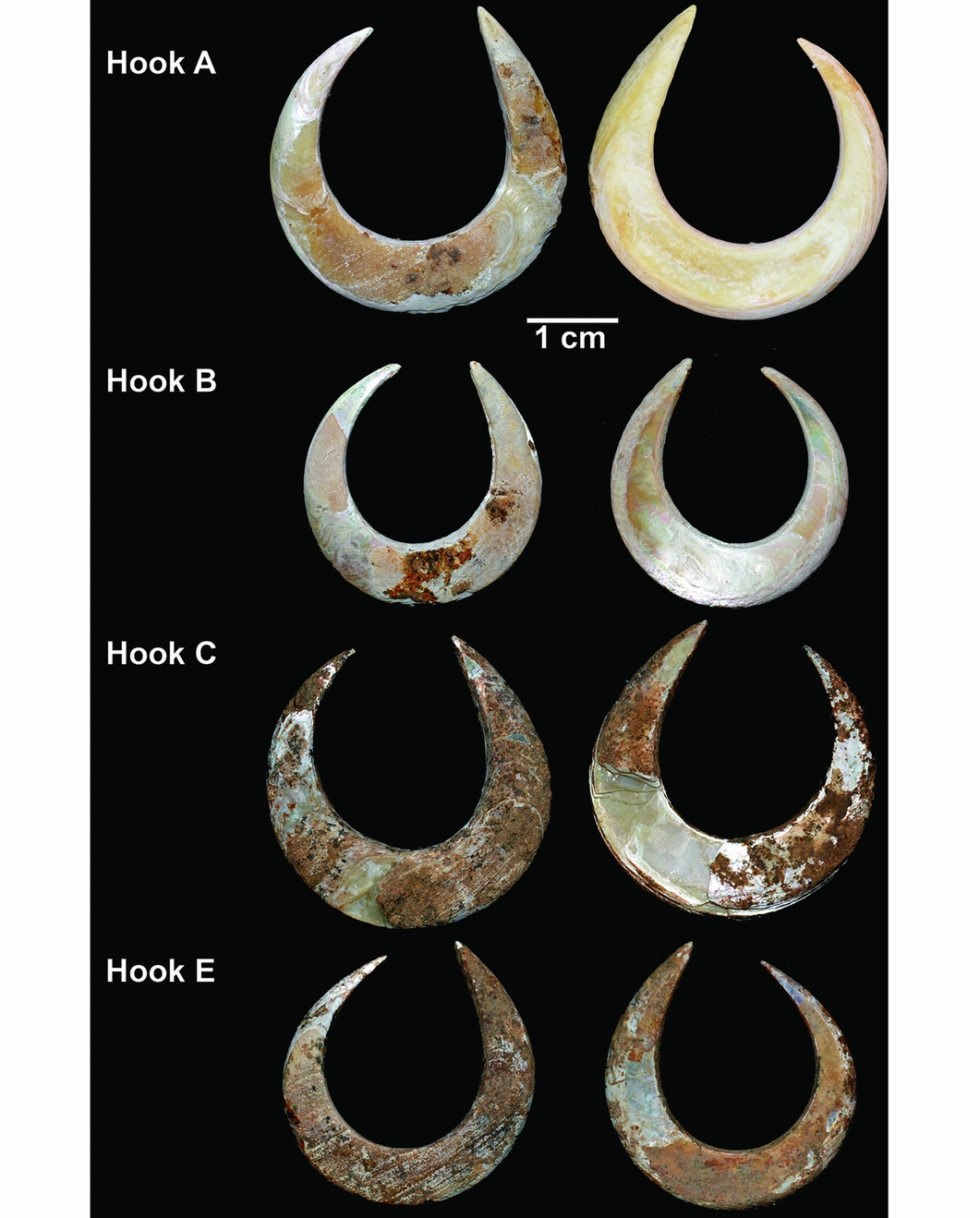
An intact fish-hook (hook A) and a perforated bivalve were found directly beneath the chin (Figures 2 & 6). Three other fish-hooks were found around the mandible (hooks B, C and E) (Figure 7). A partial hook (D) was found under the mandible (Figure 8). All were within excavation unit 41 (Figure 5).
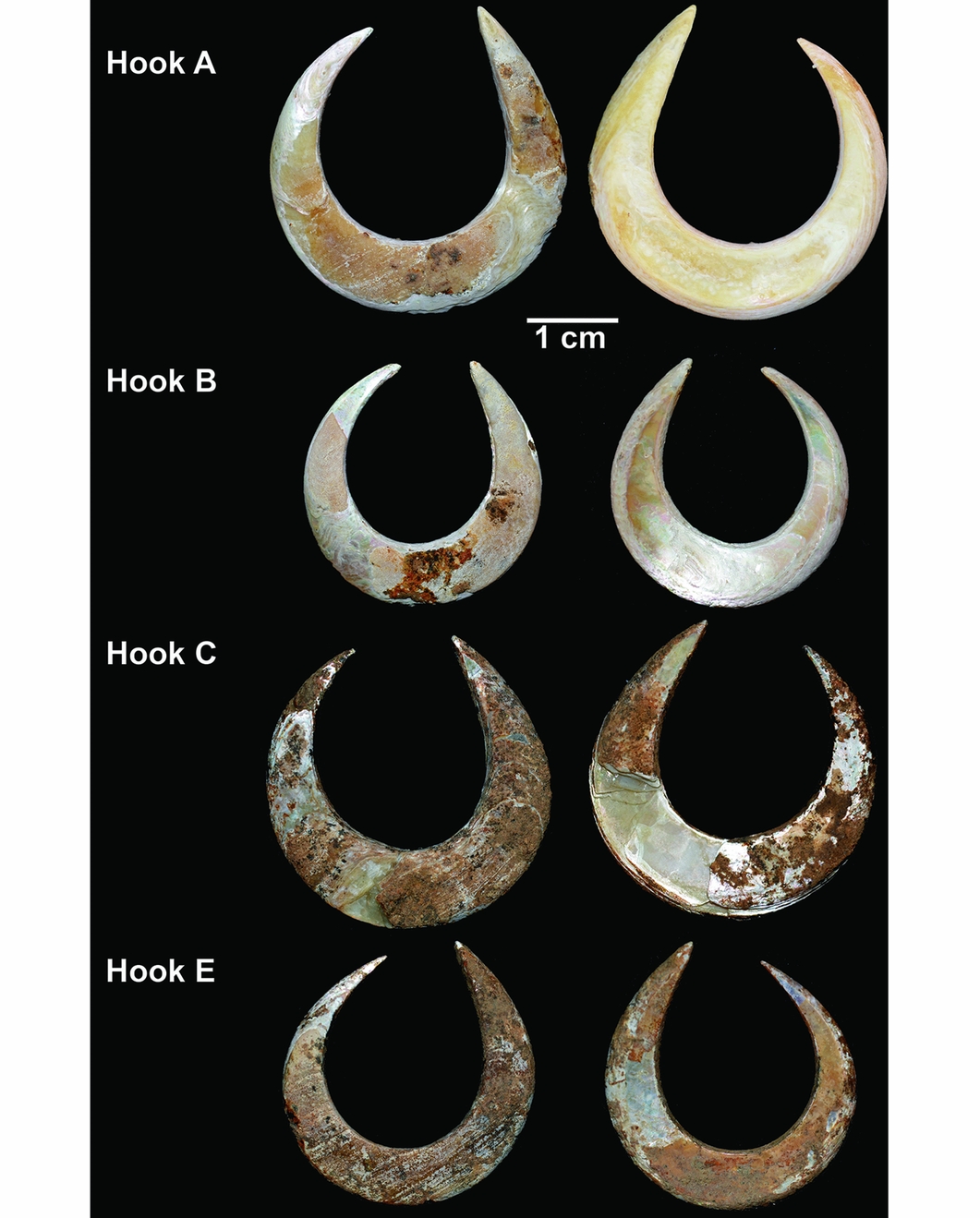
Figure 7. Circular rotating fish-hooks (A, B, C and E) found with the burial. (Photograph by Sofía Samper Carro.)
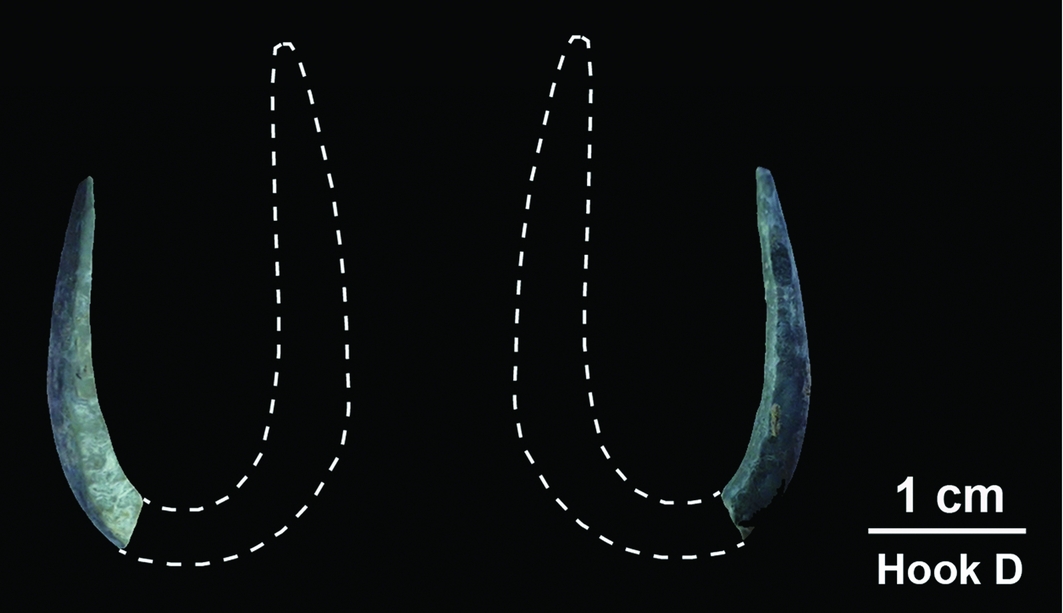
Figure 8. Tron Bon Lei fish-hook D. The dotted line shows the probable shape of the shaft.
Single-piece fish-hooks are commonly divided into two main types: J-shaped hooks in which the point and shank are more or less parallel, and circular rotating hooks, in which the point curves inwards towards the shank, leaving only a narrow opening (gape) between the point end and the shank (Kirch Reference Kirch1985; Allen Reference Allen1996). With the exception of the hook fragment (hook D; Figure 8), all of the fish-hooks (A, B, C and E) are rotating hooks (Figure 7). The hooks are described below and the measurements are provided in Table 2 (for the location of the measurements, see Figure 9).
Table 2. Measurements taken on fish-hooks.
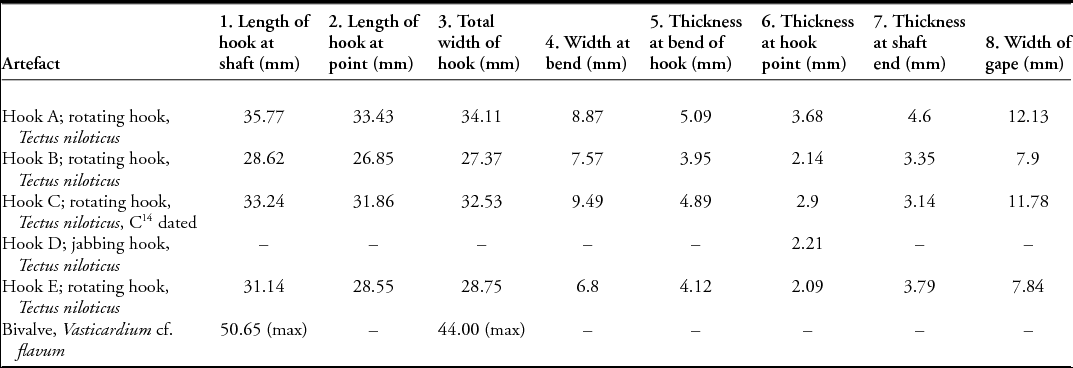
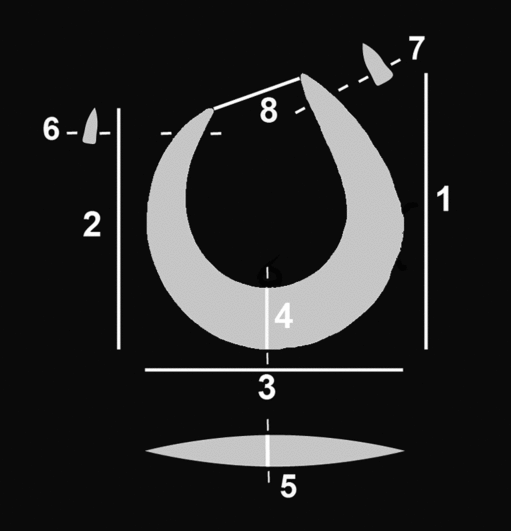
Figure 9. Schematic diagram showing the location of measurements taken on hooks in Table 2.
The rotating hooks are made on Tectus niloticus shell and are almost circular with the shaft only slightly longer than the point, and with a narrow gape. All are thickest and widest at the bend, midway between the shank and the hook, to provide mechanical strength (see OSM). Hook E retains evidence of striae that are probably the result of attempts to remove the periostracum (outer layer) to expose the nacreous surface beneath (Figure 7), which may have helped attract fish (Fujita Reference Fujita2014: 129). Hook C was fragmented during excavation and a small fragment was used for AMS dating. Hook D is thinner and finer than the rotating hooks, and appears to be from a J-shaped hook that, if complete, would have had a straight shaft (Figure 8).
The bivalve, Vasticardium cf. flavum, was found next to hook A (Figure 2). It is a left valve and is perforated in the umbo region (Figure 10). Although cracked through the perforation, there is evidence of smoothing and polish, which may result from threading. There is also evidence of extreme wear and rounding on the ventral margin of the valve consistent with use for scraping, and there are traces of a red colourant on the surface.
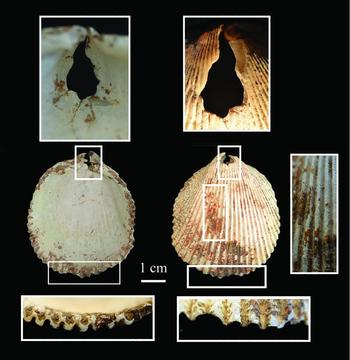
Figure 10. Bivalve Vasticardium cf. flavum (L) showing detail of the perforation at the umbo (above) and wear and rounding on the ventral margin.
Mechanical and functional attributes of fish-hooks with reference to the Tron Bon Lei assemblage
Rotating hooks and J-shaped hooks have distinct mechanical properties and function very differently (Reinman Reference Reinman1967; Allen Reference Allen1996). Nordhoff (Reference Nordhoff1930: 156) describes the differences in the mechanical action and operation of the two types in Polynesia. With a J-shaped hook, the line must be pulled sharply when the fish bites to secure the catch, whereas with a rotating hook, the inward curved point and shank rotate in the prey's mouth when the bait is taken, and the fish essentially hooks itself as it swims away. The utility of rotating hooks vs J-shaped hooks will depend primarily on marine conditions and the desired prey. The rotating hook is regarded as the optimal choice in deep waters or where strong currents or wind prevent tension being maintained on the line (Reinman Reference Reinman1967; Johannes Reference Johannes1981; Allen Reference Allen1996). Rotating hooks are also useful in shallow water when large fish are the target, and where coral heads can potentially snag a jabbing hook (Johannes Reference Johannes1981).
Some earlier writers suggested that the circular rotating hooks would not have been functional, as the gape (the distance between the hook point and the shaft) was so narrow (Bowers Reference Bowers1883). Near identical hooks have, however, been recorded ethnographically in Australia (Attenbrow Reference Attenbrow, Hutching, Lunney and Hochuli2010), Near and Remote Oceania (e.g. Anell Reference Anell1955; Reinman Reference Reinman1967; Johannes Reference Johannes1981; Allen Reference Allen1996) and California (Hoover Reference Hoover1973; Greenwood Reference Greenwood and Heizer1978). In Australia, such hooks were made and used exclusively by women. A wealth of ethnography records their primary function as being for fishing (Attenbrow Reference Attenbrow, Hutching, Lunney and Hochuli2010). Spare hooks were occasionally worn around the neck and could be used to replace those lost or broken during fishing (Figure S2).
None of the Alor hooks are barbed, and none have a notched shank for securing the line (Figure 7). Rotating hooks found in prehistoric contexts elsewhere in the world, along with ethnographic accounts of hook use, show that while some hooks have knobs or notches to prevent slippage of the line, many do not (see OSM, Figures S2 (central panel) & S3). Anell (Reference Anell1955) illustrates and discusses rotating hooks from throughout Oceania where the shaft has no modification to attach a line (Figure S3). Instead, the line was secured by lashing it along the length of the shank, and in some instances partway around the bend. This also appears to be the case with some of the archaeological rotating hooks from Espíritu Santo, Mexico, dating to about 8500 cal BP, and from the southern California coast (3000 cal BP), which also come to a point at the end of the shank akin to the Alor hooks (Hoover Reference Hoover1973; Rick et al. Reference Rick, Vellanoweth, Erlandson and Kennett2002).
The Tectus niloticus shell of which the Alor hooks are made is highly lustrous when the outer periostracum layer is removed. This quality apparently enhanced hook performance (Fujita Reference Fujita2014), and in western Melanesia, single-piece Tectus hooks were sometimes used without bait, their lustre being sufficient to attract the fish (Anell Reference Anell1955).
Discussion
Fossil remains of modern humans dating prior to the Holocene are rare east of the Wallace Line in Island Southeast Asia, and burials of this age containing grave goods have not previously been reported (Piper Reference Piper, Oxenham and Buckley2016: 33–34). Burials dating to between c. 12000 and 9000 cal BP found in the West Mouth of Niah Cave on Sarawak indicate complex treatment of the bodies of the deceased, but grave goods have only been found with one flexed inhumation (burial 27), which had a rhinoceros radius placed as a ‘pillow’ (Piper Reference Piper, Oxenham and Buckley2016: 33–34). The only other examples of burials with grave goods are a flexed burial from Hang Cho Cave in northern Vietnam, dated to c. 11000 cal BP and apparently associated with plant processing and other stone tools, and an Early Holocene burial at Song Terus in Java, which contained a monkey skull (Trachypithecus auratus) (Piper Reference Piper, Oxenham and Buckley2016: 33–34). The fish-hooks placed as grave goods in the Alor burial are thus unique within Mainland and Island Southeast Asia.
Circular rotating hooks are found in archaeological contexts in Australia (Attenbrow Reference Attenbrow, Hutching, Lunney and Hochuli2010), throughout Oceania (Anell Reference Anell1955; Reinman Reference Reinman1967; Kirch Reference Kirch1985; Allen Reference Allen1996; Burley & Shutler Reference Burley, Shutler, Anderson, Green and Leach2007; Szabó Reference Szabó, Anderson, Green and Leach2007), in Japan (Fujita et al. Reference Fujita, Yamasaki, Katagiri, Oshiro, Sano, Kurozumi, Sugawara, Kunikita, Matsuzaki, Kano, Okumura, Sone, Fujita, Kobayashi, Naruse, Kondo, Matsu'ura, Suwa and Kaifu2016), Arabia (Santini Reference Santini1987), along the California coast and islands (Rick et al. Reference Rick, Vellanoweth, Erlandson and Kennett2002), in the islands of Baja California, Mexico (Fujita Reference Fujita2014), Chile (Llagostera Reference Llagostera1992) and Ecuador (Meggers & Evans Reference Meggers and Evans1962; Bearéz et al. Reference Bearéz, Gay and Lunniss2012) (see OSM, Figure S3). Santini (Reference Santini1987) comments on how similar the rotating hooks from burial RH-10 in the Qurum area of Oman are to those in the Indo-Pacific region. The rotating hook made from Tectus niloticus found on Okinawa Island dating to between c. 23000 and 20000 cal BP is also nearly identical to those in the Tron Bon Lei burial.
The technological parallels between the circular rotating hooks on the American coast and those in Polynesia have been argued to result from diffusion as a result of transpacific voyaging and culture contact (Heyerdahl Reference Heyerdahl1952), migration or diffusion from Japan to Oceania (Anell Reference Anell1955; Reinman Reference Reinman1967), and even inadvertent transfer by migrating tuna (Landberg Reference Landberg1966). The absence of such hooks in Indonesia led Heyerdahl (Reference Heyerdahl1952) to speculate that they had their origin on the coast of South America and then spread to Polynesia. The appearance of almost identical hooks in Japan, Indonesia and Timor-Leste, dated to the Terminal Pleistocene, and their absence in the Philippines, Borneo, Sarawak and other Indonesian islands at this early date makes cultural diffusion an unlikely explanation for their appearance in the archaeological record in widely spaced regions of the world. Rather, rotating hooks can be seen as an example of convergent cultural evolution in technology, in which this particular morphology arose independently in different areas as a response to particular functional requirements and under similar ecological conditions.
Conclusion
In both life and death, the Pleistocene inhabitants of Alor Island were intrinsically connected to the sea. The association of the fish-hooks with a burial denotes the cosmological status of fishing in this island environment, probably because of the role that fishing played during daily life for the inhabitants of an island that largely lacked other sources of protein. If the burial is confirmed as that of a female, the inclusion of fish-hooks as grave goods may indicate that women were responsible for hook-and-line fishing in Alor, as they were in Australia.
The Alor hooks, which bear an uncanny resemblance to the rotating hooks used in Japan, Australia, Arabia, California, Chile, Mexico and Oceania, provide a salutary warning against reading cultural diffusion as the cause of such similarities. Convergent technological evolution, whereby the same form of artefact develops independently in multiple, widely spaced locations because it is the most fitting form to meet the functional requirements of particular ecological conditions, is clearly possible. We argue that it is the most probable explanation for the distribution of circular rotating hooks in these widely spaced locales across the globe. To our knowledge, the fish-hooks from Alor represent the earliest excavated evidence for a direct link between cosmology and fishing. These finds, in conjunction with rare representations of fish found in the late Pleistocene rock art of France and Egypt (Roussot Reference Roussot1984; Huyge et al. Reference Huyge, Vandenberghe, De Dapper, Mees, Claes and Darnell2011), and fish-hooks in burials dated from the Early Holocene in Siberia and Oman (Santini Reference Santini1987; Basaliiski Reference Basaliiski, Weber, Katzenberg and Schurr2010), speak to the cosmological significance of fishing to late Palaeolithic societies throughout the world, perhaps to a larger extent than has previously been recognised.
Acknowledgements
An ARC Laureate to O'Connor (FL120100156) funded the research, which was conducted under an Indonesian Foreign Research Permit (O'Connor 1304/FRP/SM/V/2014). We would like to thank the community of Lerabain, the students from Universitas Gadjah Mada, Yogyakarta, and Putu Yuda from Balar Arkeologi Bali.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2017.186












 Open access
Open access