



Introduction
The chicken (Gallus gallus domesticus) is the most widely distributed domestic animal on the planet (Nicol Reference Nicol2015). Transported around the world by people, the species is now established across a broad range of ecosystems and societies, providing humans with increasing quantities of both meat and eggs (Bennett et al. Reference Bennett2018). Given their modern ubiquity in the human food chain, it is easy to assume that chickens were domesticated primarily as a food source (e.g. Marino Reference Marino2017). There is, however, little evidence to support this hypothesis, and despite its global economic and cultural significance, the early history of the chicken is poorly understood.
Literature about the early history of chickens is largely speculative. For instance, West and Zhou (Reference West and Zhou1988) summarised—but did not challenge—claims regarding chicken domestication and diffusion. From their survey of the literature, they proposed that chickens were domesticated in Southeast Asia c. 6000 BC, before quickly becoming established in China and spreading rapidly into western Eurasia. West and Zhou (Reference West and Zhou1988) also suggested that chickens arrived in Eastern Europe by the Neolithic, before spreading throughout the Mediterranean during the Bronze Age and reaching temperate Europe in the Iron Age. Other studies (e.g. those cited by Ledogar et al. Reference Ledogar, Karsten and Sokhastskyi2019) have proposed that chickens were not only established in Eastern Europe by the Neolithic, but that the species may even have been native to the region (Boev Reference Boev1995). West and Zhou's (Reference West and Zhou1988) work continues to be cited frequently, although several recent studies have questioned the validity of the evidence it presents (e.g. Eda et al. Reference Eda2016; Peters et al. Reference Peters, Lebrasseur, Deng and Larson2016; Huang et al. Reference Huang2018; Peters et al. Reference Petersin press). Based on comprehensive zooarchaeological re-analyses and ecological modelling, these publications argue that chickens could not have been domesticated in the seventh millennium BC, and that the third millennium BC is more probable. Although these refined dates have important implications for diffusion models (Pitt et al. Reference Pitt, Gillingham, Maltby and Stewart2016), many publications (e.g. Bennett et al. Reference Bennett2018; Sykes Reference Sykes2018) continue to cite some of West and Zhou's (Reference West and Zhou1988) conclusions without questioning the underpinning archaeological data.
There are numerous reasons why these archaeological data should be questioned. Issues of taphonomy and recovery bias can lead to an under-representation of archaeological chicken bones, making it difficult to reconstruct ancient distributions (Serjeantson Reference Serjeantson2009; Dirrigl et al. Reference Dirrigl, Brush, Morales-Muñiz and Bartosiewicz2020). This is compounded by problems of identification. For example, re-analyses by Eda et al. (Reference Eda2016) and Peters et al. (Reference Peters, Lebrasseur, Deng and Larson2016) reveal that several bones originally identified as early chicken remains are actually from pheasants (Phasianus sp.). The most significant factor obfuscating the bio-cultural history of the chicken, however, is imprecise dating. Chicken bones are prone to stratigraphic movement via bioturbation or through building and agricultural activities. Flink et al. (Reference Flink2014) directly dated a chicken bone from an Iron Age context (280–15 BC) at Altenburg, Germany, and found it to be a recent intrusion (150±30 BP, cal AD 1667–1903, at 95.4% confidence). Similarly, Ledogar et al. (Reference Ledogar, Karsten, Madden and Schmidt2018) demonstrate that supposed Neolithic specimens from a Ukrainian cave were also intrusive. Such examples of direct dating are rare, yet they frequently highlight the fallibility of dating chicken remains using only stratigraphic contexts.
To test whether other early chicken bones are also intrusive, we directly radiocarbon-dated many of the earliest claimed specimens from Europe and north-west Africa. The results allow us to re-evaluate the arrival and spread of chickens across these regions and to discuss the shifting relationships between humans and chickens through time.
Materials and methods
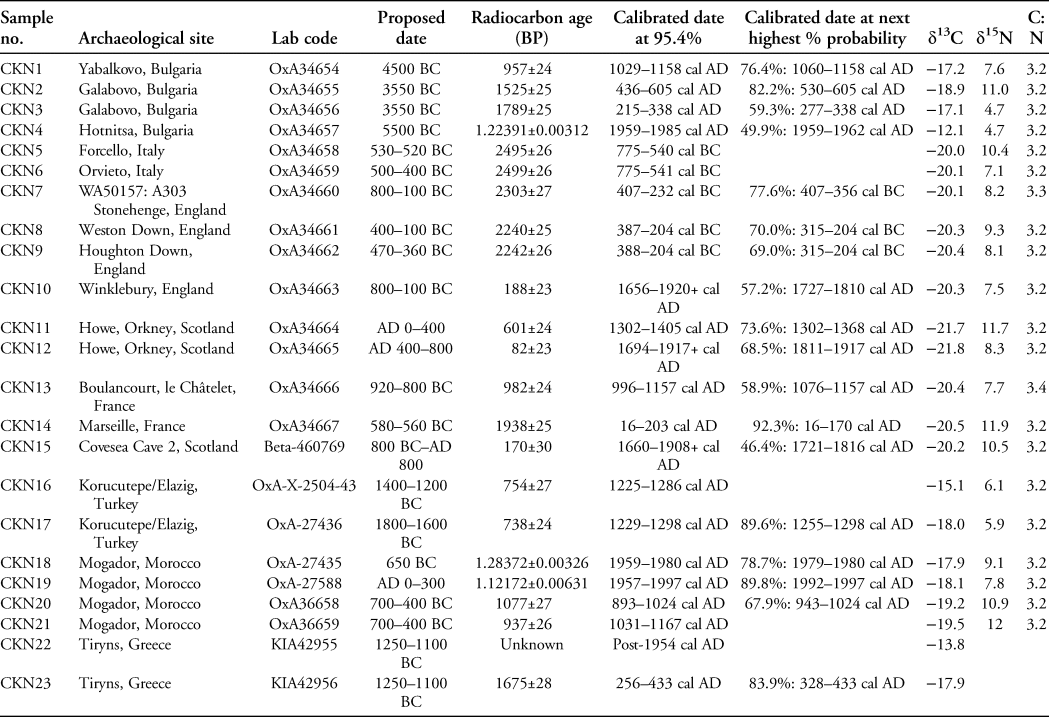
Twenty-three chicken bones were selected from 16 archaeological sites for direct radiocarbon dating (Figure 1; Table 1; for further details and references, see Table S1 in the online supplementary material (OSM)). The samples from Bulgaria were suggested to date to the Neolithic/Bronze Age, and those from Turkey and Greece were supposedly of Bronze Age date. For France, purportedly Bronze Age/Early Iron Age specimens were selected. We also targeted samples from Iron Age sites in Italy, Morocco and England, where claims have been made for the early presence of chickens. Lastly, we examined chicken bones from Iron Age sites in Scotland, although here the Iron Age extends much later, to AD 800.
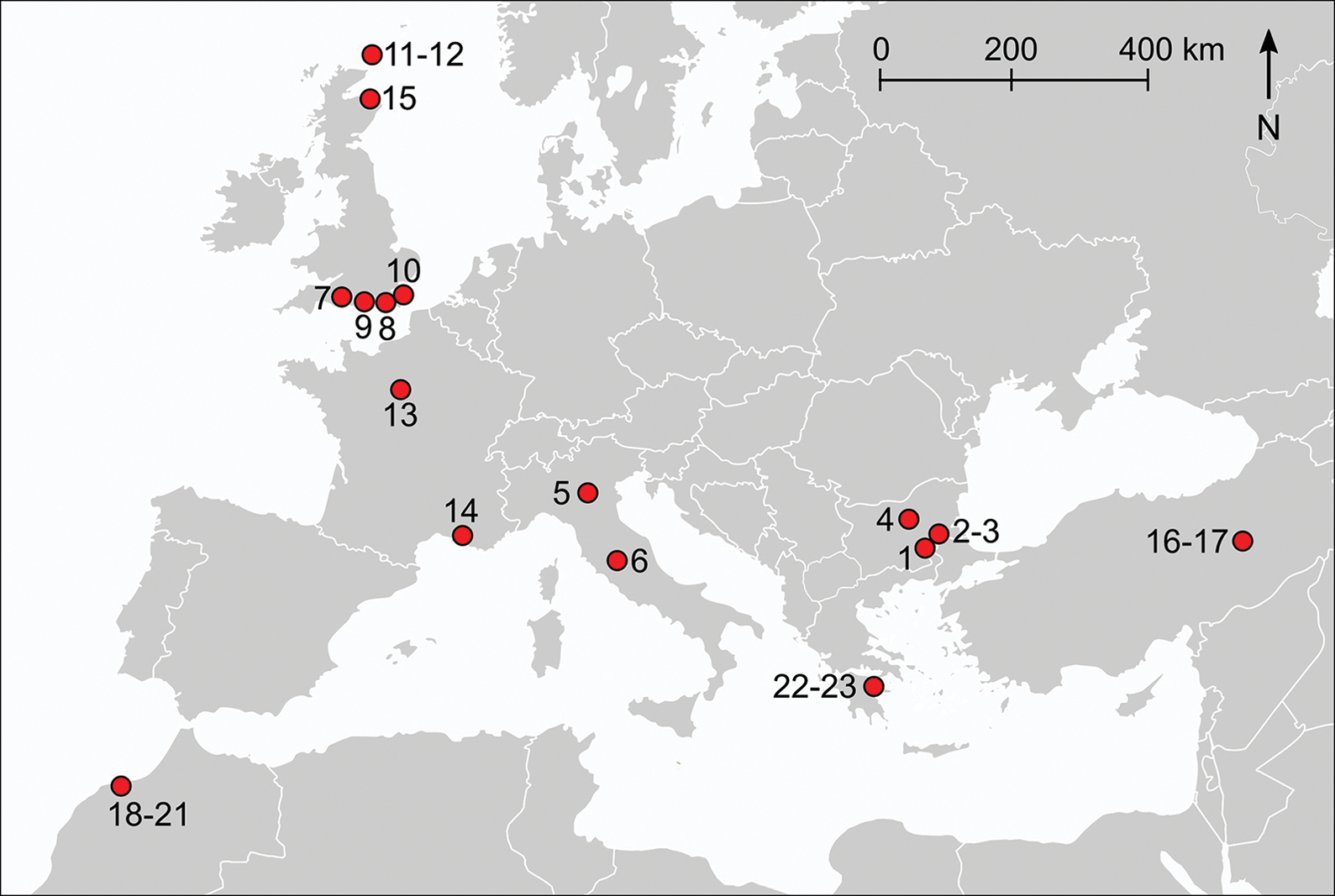
Figure 1. Map of sample locations by sample numbers CKN1–23 (see Table 1, Table S1 and OSM) (figure by S. Doherty).
Table 1. Site and location data for dated samples (see Figure 1; for sample references, see the OSM).
Radiocarbon dating was undertaken by three separate laboratories (Oxford Radiocarbon Accelerator Unit: n = 20; Kiel AMS: n = 2; and Beta Analytic: n = 1). Carbon (δ13C) and nitrogen (δ15N) isotope data derived from the dating process were incorporated into our wider project dataset. Prior to destruction, specimens were measured and examined for evidence of sex, age and butchery. Detailed analytical methods are provided in the OSM. These approaches help to ascertain a specimen's archaeological status. Bennett et al. (Reference Bennett2018) demonstrate that ancient and modern chickens can be differentiated morphologically, as modern chickens grow much faster, and their bones are larger in every dimension compared with ancient specimens. These differences are due to advances in poultry feeding and selective breeding that have resulted in significant genetic changes (Flink et al. Reference Flink2014; Loog et al. Reference Loog2017). Dietary differences can also be observed isotopically: modern chicken diets contain higher quantities of C4 plants—notably maize—and far lower levels of protein relative to their ancient counterparts (Bennett et al. Reference Bennett2018). Contextual information, along with data on whether specimens were recovered as isolated bones or complete skeletons, can indicate the risk of intrusion, while also revealing human attitudes towards chickens.
Results
Of the 23 dated chicken bones, only five were consistent with their reported stratigraphic phasing. The radiocarbon dates associated with the remaining 18 were more recent than their reported dates (Table 2; Figure 2).
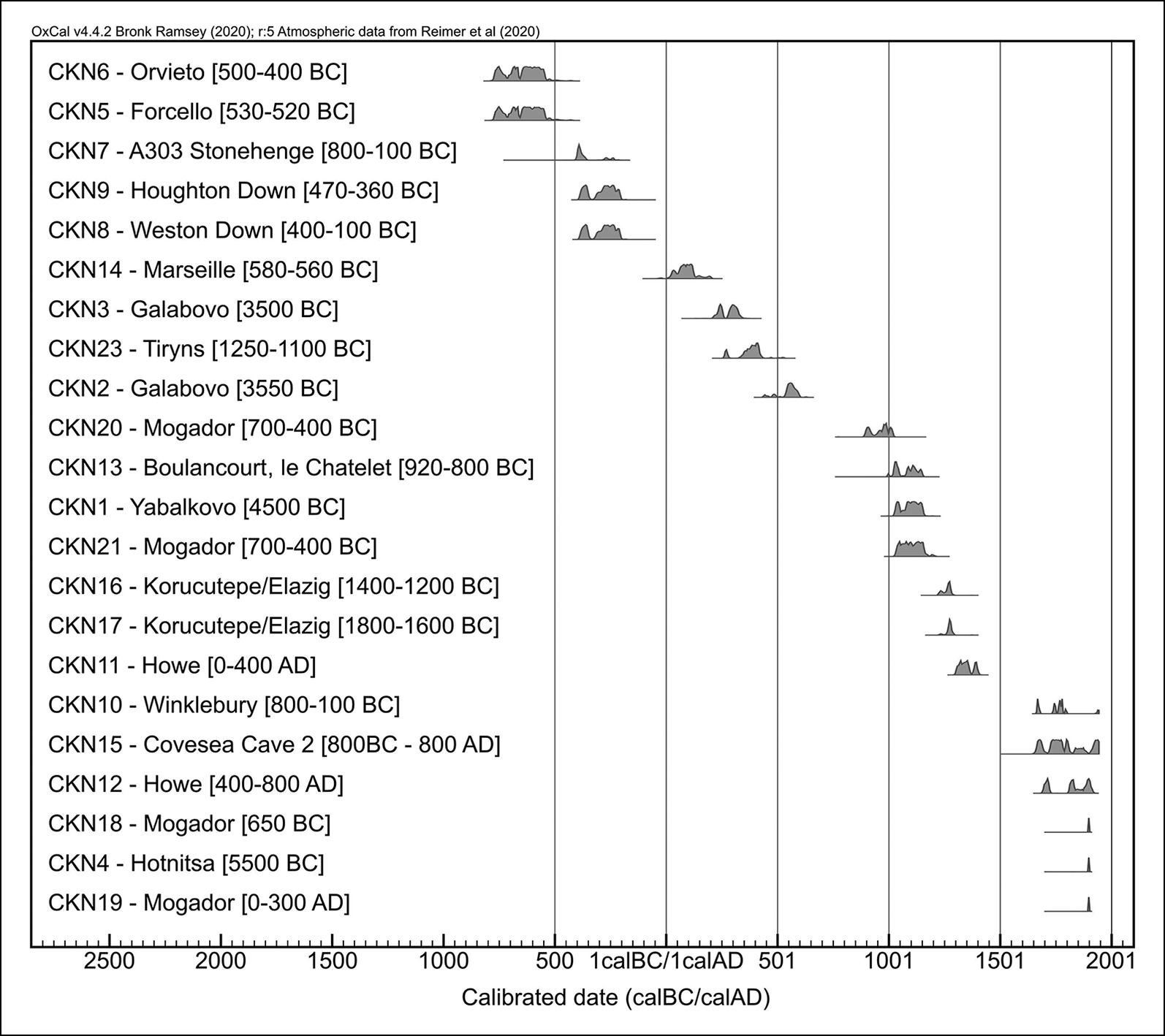
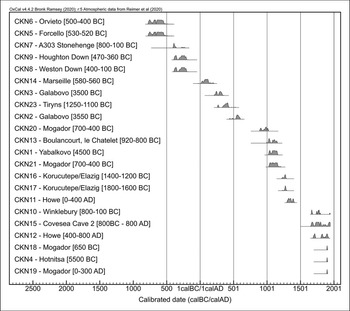
Figure 2. Calibrated radiocarbon results for each specimen, with stratigraphically proposed dates in brackets (for further information, see Tables 1 & 2, Table S1 and OSM). CKN22 (thought to date to 1250–1100 BC) was determined to be ‘post-1954’ and is not included (dates calibrated in OxCal v.4.4.2 using the IntCal20 atmospheric curve, except CKN4, CKN18 and CKN19, where the Bomb13NH1 curve was used; Bronk Ramsey Reference Bronk Ramsey2009; Reimer et al. Reference Reimer2020) (figure by J. Best and S. Doherty).
Table 2. Sample details and results for the new series of AMS dates.
Radiocarbon dates for the chicken bones derived from Neolithic/Bronze Age Hotnitsa (Bulgaria: CKN4), Bronze Age Tiryns (Greece: CKN22) and two specimens from Iron Age/Roman Mogador (Morocco: CKN18 and CKN19) were modern (post-1950s). These recent dates also explain the morphological and isotopic results from the same bones, which more closely resemble those of modern broilers (commercially raised meat birds) than ancient chickens (Figures 3 & 4).
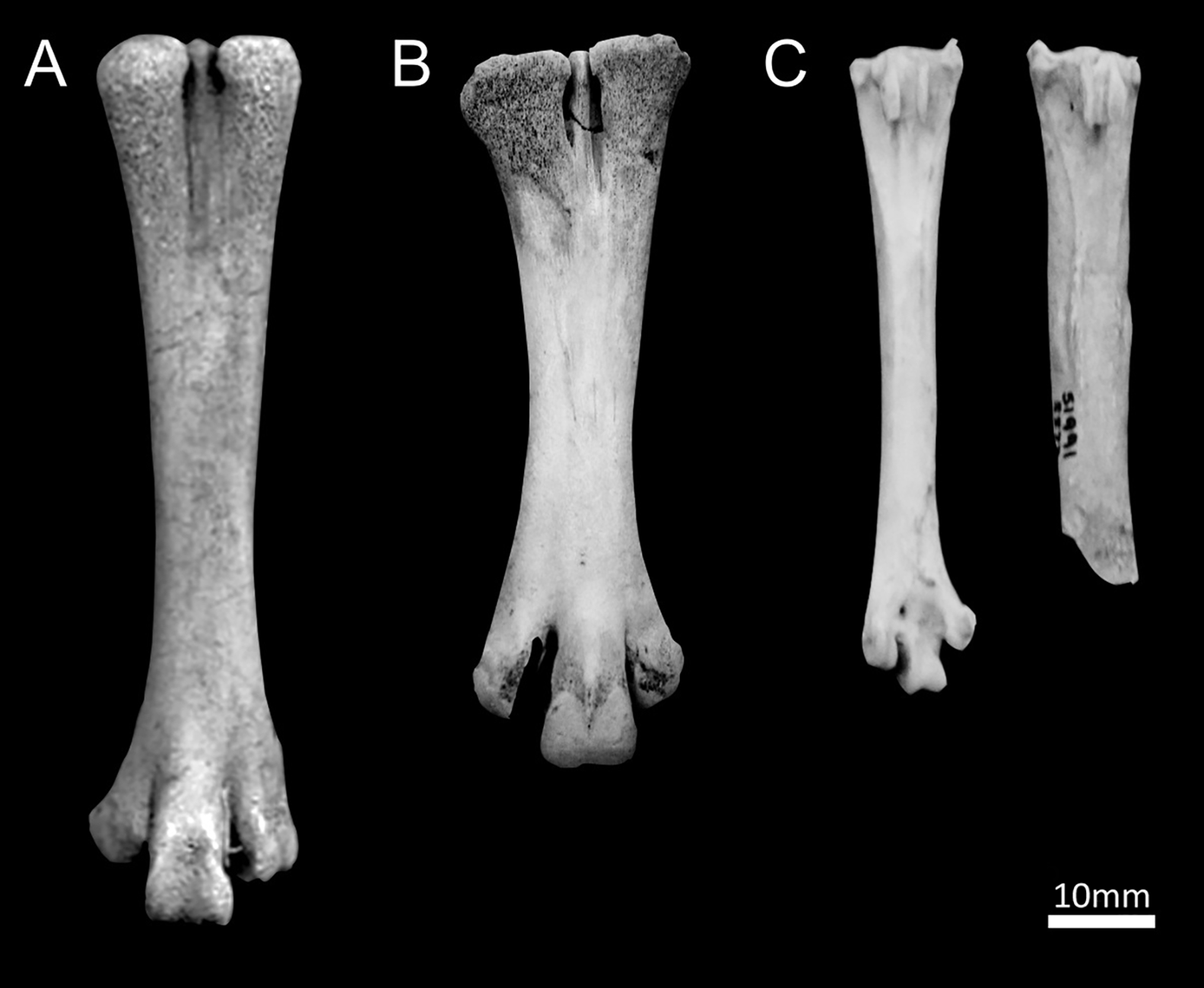
Figure 3. Comparison of chicken tarsometatarsi from: A) Mogador (Becker Reference Becker, von Carnap-Bornheim and Kroll2013); B) modern broiler; C) Iron Age Weston Down (photograph by J. Best), showing healthy bone on the left and fractured bone on the right (CKN8) (graphic by S. Doherty).
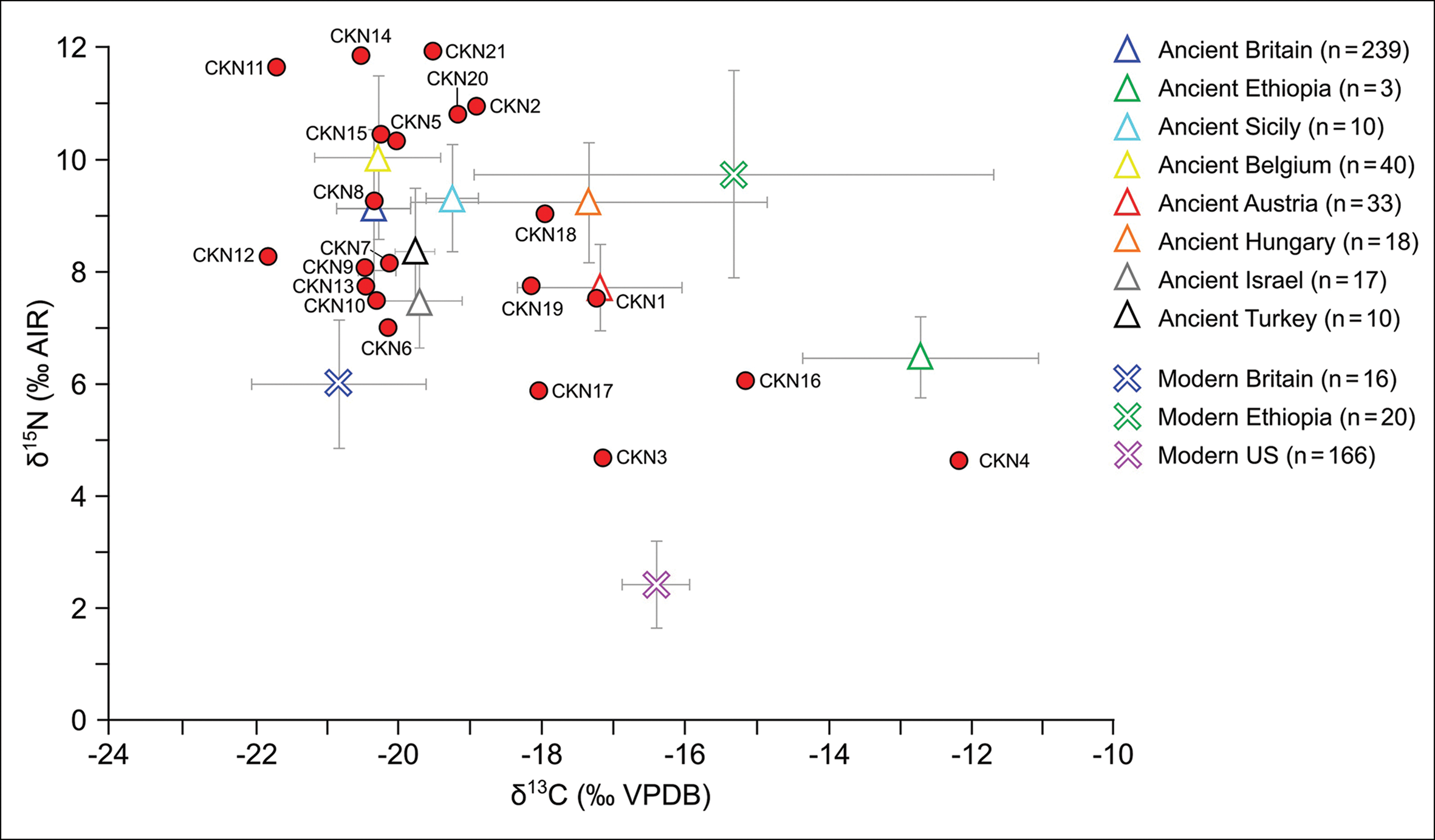
Figure 4: Isotope values for the dated specimens (see Table 2) against broader isotope dataset for ancient and modern chickens (figure by H. Miller and S. Doherty).
Differences between ancient and modern poultry production are exemplified by the chickens from Mogador. The two modern individuals (CKN18 and CKN19) had more positive δ13C values and more negative δ15N values compared with the two medieval-dated specimens (CKN20 and CKN21), whose isotope values plot within the distribution of other ancient chicken remains (Figure 4).
Other intrusive chicken bones include specimen CKN23 from Tiryns (Greece), which is approximately 1300 years younger than its Bronze Age context. The two specimens from Galabovo (Bulgaria) are over 3500 years younger than originally claimed (CKN3: cal AD 215–338 and CKN2: cal AD 436–605), and specimen CKN1 from Yabalkovo (Bulgaria) is around 5000 years younger, dating to the eleventh to twelfth centuries AD.
Both proposed Late Bronze Age specimens from Korucutepe in Turkey (CKN16 and CKN17) date to the thirteenth century AD, and CKN13 from Boulancourt, le Châtelet (France) is also medieval, rather than Bronze Age. Specimen CKN14 from Marseille (France), which was thought to be from a secure Iron Age context, is re-dated to the Roman period. At Covesea Cave 2 (Scotland), specimen CKN15 dates to the seventeenth to twentieth centuries AD, rather than Iron Age (800 BC–AD 800). Finally, the two chicken bones selected from Howe, Orkney, date to the fourteenth to fifteenth centuries AD (CKN11) and the seventeenth to twentieth centuries AD (CKN12), rather than their AD 200–800 context.
Articulated skeletons are generally acknowledged to be reliable indicators of a secure archaeological context (Baker & Worley Reference Baker and Worley2019: 18). Despite this, the purportedly Iron Age chicken skeleton from Winklebury (CKN10, England) returned a post-medieval/modern date. The skeletons from Weston Down (CKN8) and Houghton Down (CKN9) are, however, consistent with their fourth/third century BC contexts. The isolated chicken bone from the Stonehenge Road Improvement (CKN7) appears to be slightly earlier than the Weston Down and Houghton Down examples, extending into the late fifth century BC.
The earliest radiocarbon dates returned in this programme of dating are from two Italian sites, Forcello (CKN5) and Orvieto (CKN6), both of which are consistent with their respective sixth and fifth century BC contexts. Their broadest radiocarbon date ranges extend into the mid-eighth century BC, but this probably reflects the Hallstatt plateau—a flat area of the calibration curve that reduces the precision of determinations during this period. The specimen from Forcello (CKN5), again, derives from an articulated skeleton.
Discussion and conclusion
Our programme of radiocarbon dating redefines the established chronology for the arrival and dispersal of chickens across Europe and north-west Africa. Specifically, we found no evidence for chickens in Europe before the first millennium BC, nor do our results support claims of an autochthonous Holocene population of junglefowl in Eastern Europe. Instead, our results suggest that all claims for the presence of pre-Iron Age European chickens should be rejected unless supported by direct radiocarbon dating of the bones themselves.
A revised spatio-temporal pattern of the spread of chickens
Our results undermine claims of a seventh-century BC presence of chickens in north-west Africa (Mogador, Morocco) but specimens from this site did date to the ninth to twelfth centuries AD. This is consistent with current models suggesting that, following their ninth to sixth-century BC introduction in the Horn of Africa (Woldekiros & D'Andrea Reference Woldekiros and D'Andrea2016: 334), chickens spread across the continent slowly. Mwacharo et al. (Reference Mwacharo, Bjørnstad, Han and Hanotte2013) argue that chickens were not established in the north-west until the medieval period, whereas Oueslati et al. (Reference Oueslati2020) propose a first-century BC arrival, but neither of these studies are based on directly dated specimens.
Our results support the accepted chronology that chickens were present in Italy by the eighth century BC (Trentacoste Reference Trentacoste2020; Corbino et al. Reference Corbino, Minniti, De Grossi Mazzorin and Albarellain press). This is a similar timeframe to their arrival on the Balearic Islands, where a chicken bone has been directly dated to the eighth to sixth century BC (Ramis et al. Reference Ramis, Anglada, Ferrer, Plantalamor and Van Strydonck2017). It seems likely that chickens were transported throughout the Mediterranean along routes ecologically suited to these thermophilic birds (Pitt et al. Reference Pitt, Gillingham, Maltby and Stewart2016), probably via early Greek, Etruscan and Phoenician maritime trade (Peters et al. Reference Petersin press).
The human-assisted movement of chickens into Central and northern Europe occurred over the following centuries. A directly dated chicken skeleton from the Czech Republic (Kyselý Reference Kyselý2010), along with zooarchaeological and iconographic data from Bulgaria, indicate their arrival in Central Europe in the sixth to fifth centuries BC (Boev Reference Boev1995). Chickens were also introduced into France and southern Britain by the sixth to fifth centuries BC (Kitch Reference Kitch2006; Peters et al. Reference Petersin press).
It took almost 1000 years longer for chickens to become established in the colder climates of Scotland, Ireland, Scandinavia and Iceland (Best Reference Best2014; Best & Mulville Reference Best and Mulville2014; Sykes Reference Sykes2018; Walker & Meijer Reference Walker and Meijer2020). Our direct dates also support the suggestion that chickens were not introduced to the Scottish Isles until the Norse arrivals, from c. AD 800 (Best Reference Best2014).
The movement of chicken bones through archaeological stratigraphies
With one exception, all of the specimens that did not match their contextually assigned dates were isolated bones. This result highlights the ease with which chicken bones (and other faunal remains) can migrate through archaeological contexts or be misassigned, thus necessitating confirmation of date by direct methods. Direct dating is also desirable for articulated skeletons, since the purportedly Iron Age example from Winklebury (England) proved to be a modern intrusion. However, direct dates from the other articulated remains examined here corroborate their stratigraphic phasing, showing them to be among the earliest regional specimens in our European dataset.
Many of the earliest dated chicken remains reported are from complete or nearly complete skeletons. This may be because they have been preferentially targeted for radiocarbon dating programmes (Baker & Worley Reference Baker and Worley2019: 18). A more intriguing possibility, however, is that the deposition of complete chickens reflects how the species was perceived and treated by humans during the earliest stages of their human-mediated dispersal.
The dynamics of human-chicken relationships
Globally, the first convincing evidence for close relationships between humans and chickens comes from complete skeletons placed alongside Bronze Age human burials in Thailand (e.g. Ban Non Wat, c. 800 BC) and China (Dasikongcun royal cemetery, 1320–1046 BC) (Peters et al. Reference Petersin press). The same applies to Italy, where the earliest identified chicken is from a tenth to ninth-century BC tomb (Corbino et al. Reference Corbino, Minniti, De Grossi Mazzorin and Albarellain press), with other possible eighth-century examples (see OSM) (De Grossi Mazzorin & Minniti Reference De Grossi Mazzorin and Minniti2019; Trentacoste Reference Trentacoste2020), although, crucially, none of these have been directly dated. It is possible that this pattern could be the product of research bias resulting from the preferential excavation of funerary contexts. Earlier evidence for chickens may be awaiting discovery on other site types.
To understand how human-chicken relationships evolved from the point of introduction, and as their populations increased, it is necessary to focus on the evidence from regions for which there is an extensive (zoo)archaeological record that covers a variety of site types. For northern Europe, and in particular Britain, there is a sufficient body of securely dated evidence to propose a model for how attitudes to chickens changed through time (Figure 5A–E). In many areas, chickens appear initially not in human burials, but as individually buried skeletons. In addition to those dated from Weston Down (CKN8) and Houghton Down (CKN9) in Britain, articulated chicken remains have been recovered from Iron Age sites across Europe (e.g. Peters et al. Reference Petersin press). For the Czech Republic, Kyselý (Reference Kyselý2010) reported an adult cockerel skeleton from Rubín that is radiocarbon dated to 2380±30 BP (542–393 cal BC, at 93.9% confidence). It is possible that isolated bones of early date could also have been buried as complete animals but became disarticulated as a result of taphonomic processes.
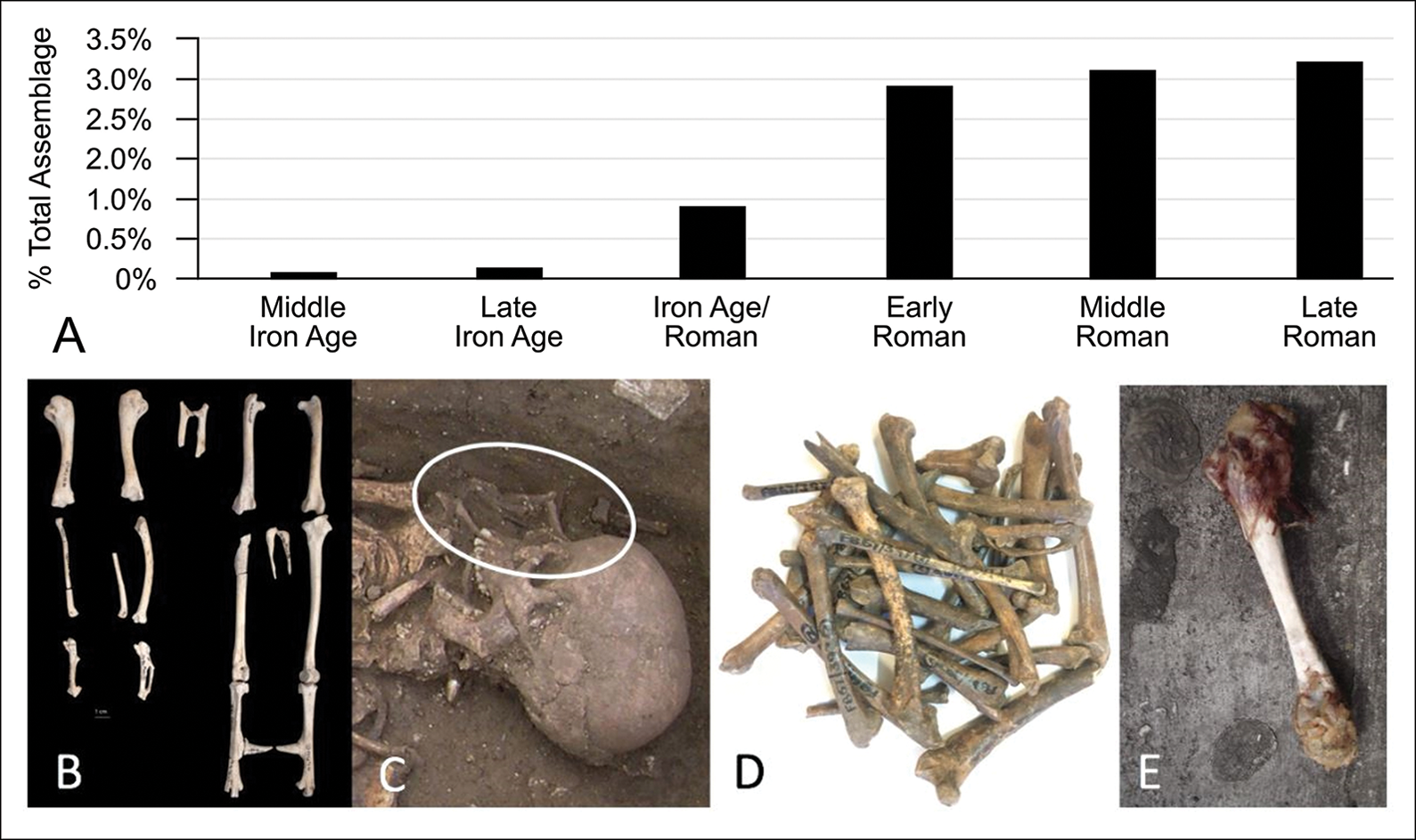
Figure 5. The association between (A) zooarchaeological representation (Skelton Reference Skelton2019) and depositional context. Earliest chickens were rare and often individually buried, as at (B) Middle Iron Age Houghton Down (CKN9). As chicken populations increased, they moved from being incorporated into human-chicken co-burial, as at (C) Roman Broughton (Yorkshire), to foodstuff, as at (D) Fishbourne Roman Palace. Today, chickens are so omnipresent that they are simply litter on the streets (E) (figure by S. Doherty).
Notably, none of these skeletons show evidence of butchery or human consumption; they are also often older animals. The long spurs on the Houghton Down cockerel (Figure 5B), for example, suggest it was over two years old (Doherty et al. Reference Doherty2021). Similarly, the hen from Weston Down (CKN8) was a mature individual with a well-healed leg fracture (Figure 3C), which could suggest evidence of human care.
Rather than being considered as a source of food, these early arrivals to northern Europe were more likely regarded as exotica, especially given their limited population size at the time (Figure 5A). The idea that chickens were too rare or too important to be slaughtered for meat is consistent with Caesar's De Bello Gallico (5.12; Edwards Reference Edwards1989) that states: “The Britons consider it contrary to divine law to eat the hare, the chicken, or the goose”. Helms (Reference Helms1993) suggested that, in many cultures, animals and things derived from the outer realms are often attributed with cosmological powers. Given the exotic nature of chickens at the time of introduction, this could explain their depiction on Late Iron Age coins—themselves artefacts of power—recovered from southern Britain and northern France (Feider et al. Reference Feider, Hambleton, Maltby, Selsvold and Webb2020).
During the Late Iron Age and Early Roman period in Britain (approximately 50BC–AD 100), there is an observable shift not only in the frequency of chicken remains within zooarchaeological assemblages (Figure 5A), but also towards human-chicken co-burials (Figure 5C)—a phenomenon seen in other areas of northern Europe at this time (Lauwerier Reference Lauwerier1993; Sykes Reference Sykes2012; Kunst & Doneus Reference Kunst and Doneus2013). Our survey of British co-burials indicates that these funerary rites were often strongly gendered: males were buried with cockerels and females with hens (as at Broughton, Yorkshire; Figure 5B). Chickens may have been included within human graves as psychopomps, whose role it was to lead human souls to the afterlife. Such a role would have befitted their association with Mercury (the Roman god of communication and travel), to whom large quantities of cockerels were sacrificed at the Temple of Uley, Gloucestershire (Woodward & Leach Reference Woodward and Leach1993). On other occasions, the presence of chickens in graves clearly represents a food offering, a practice that became more common in Britain through the Roman period (White Reference White, Croxford, Ray, Roth and White2007).
The expansion of the Roman Empire helped to popularise chickens and eggs as foodstuffs (e.g. Peters Reference Peters1998; Maltby et al. Reference Maltby2018). In Britain, the earliest evidence for high levels of chicken consumption comes from the ‘Romanised’ site of Fishbourne Palace, where Allen (Reference Allen2011) has demonstrated that exceptional numbers of chickens were eaten as early as the first century AD (indicated by two direct dates: 1970±30 BP and 1920±30 BP). Here, chicken bones comprised eight per cent of the total assemblage, far higher than on other British Iron Age/Roman sites (Figure 5A). Elsewhere in Britain, chickens were not regularly consumed until the third century AD, again, primarily on highly Romanised urban and military sites (Maltby et al. Reference Maltby2018).
This evidence suggests that, in Britain, 700–800 years elapsed between the initial introduction of the chicken as an exotic (whose flesh was apparently prohibited for consumption) and the acceptance of these animals as a source of dietary protein. An equivalent time-lag also appears to apply in Italy, where chickens were sporadically represented in tombs and cult places for the first few centuries after their arrival in the tenth/ninth to sixth centuries BC. By the sixth to fifth centuries BC, chickens had become more abundant and were occasionally eaten on settlement sites (e.g. at Forcello, Bagnolo San Vito, where chicken bones show butchery marks), and they only became a more frequent dietary component from the fourth century BC (De Grossi Mazzorin & Minniti Reference De Grossi Mazzorin and Minniti2019; Trentacoste Reference Trentacoste2020). Similarly, in the Levant, although chickens were present in the ninth/eighth century BC (Peters et al. Reference Petersin press), it was not until the fourth/third century BC that they became a commonly exploited source of meat (Perry-Gal et al. Reference Perry-Gal, Erlich, Gilboa and Bar-Oz2015).
The trend toward perceiving chickens solely as dietary protein has accelerated to the present day. Whereas the earliest chickens in Europe were rare and lived beyond maturity, today this is often inverted. Of the >70 billion chickens now on the planet, most are commercially raised broilers that grow exceptionally quickly during their short lives (the average slaughter age is 42 days; European Food Safety Authority Panel on Animal Health & Welfare 2010). They are seldom buried as individuals or with people, and instead are often disposed of as fast-food refuse, littered in the street (Figure 5E). Although recent changes in chicken size, shape, genetics and diet allow for a more robust assessment of their intrusive status (Figures 3 & 4) in archaeological contexts, these characteristics are also an eloquent expression of how dramatically human-chicken relationships have changed over the last three millennia.
Acknowledgements
We thank the two anonymous reviewers, plus Umberto Albarella, David Allen, Chiara Corbino, Raffaele de Marinis, Gail Drinkall, David George, Catriona Gibson, Julie Hamilton, Lorraine Mepham, Jacqueline Pitt, Marta Rapi and The Friends of Active Archaeology (notably Melanie Dalton and David Snowden) for their contributions.
Funding statement
This article was supported by the AHRC (AH/L006979/1 and AH/N004558/1), by the NERC Radiocarbon Facility (NF/2015/2/5) and by the Chair of Palaeoanatomy, LMU Munich.
Supplementary material
To view supplementary material for this article, please visit https://doi.org/10.15184/aqy.2021.90








 Open access
Open access