


In the last decade functional foods in human and animal nutrition have gained in importance. Within this field of research, scientific concepts underpinning directed modulation of the human gut microbiota towards a more beneficial composition have been developed(Reference Kolida, Saulnier and Gibson1, Reference Hamilton-Miller2). The prebiotic approach advocates targeting selected indigenous beneficial bacteria through non-viable food ingredients(Reference Gibson and Roberfroid3). The concept was recently updated by Gibson et al. (Reference Gibson, Probert and Van Loo4), and the weight of evidence for established and emerging prebiotics reviewed(Reference Roberfroid5). Much of the interest in the development of prebiotics aims at non-digestible oligosaccharides. These are short-chain carbohydrates that consist of two to twenty saccharide units. Examples include inulin-type fructans, galacto-oligosaccharides, isomalto-oligosaccharides, xylo-oligosaccharides, soya-oligosaccharides, gluco-oligosaccharides and lacto-sucrose(Reference Rastall and Maitin6, Reference Gibson, Berry Ottaway and Rastall7), although each of these varies in their prebiotic potential.
Inulin and fructo-oligosaccharides are plant β (2-1) fructans with a degree of polymerisation (DP) ranging from 2 to 60 or more for inulin, or from 2 to 10 for oligofructose(Reference Van Loo, Coussement and de Leenheer8).
There is a high degree of variability in DP distribution among products derived from different plant origins. Inulin derived from globe artichoke (Cynara scolymus) generally has the highest DP(Reference Praznik, Beck and Eigner9) whereas onion, garlic, Jerusalem artichoke (Heliantus tuberosus), leeks, asparagus and banana have lower DP(Reference Van Loo, Coussement and de Leenheer8). The inulin content of Jerusalem artichoke ranges from 17 to 20·5 % on a fresh weight basis. Praznik et al. (Reference Praznik, Beck and Nitsch10) found that 74 % of Jerusalem artichoke inulin had a DP < 19 while 20 % was between DP 19 and 40 and 6 % had a DP>40. The inulin content of chicory (Chichorium intybus), harvested from roots, ranged from 15·2 to 20·5 %, of which 55 % had a DP of 2 to 19, 28 % with DP between 19 and 40, and 17 % with DP>40(Reference Van Loo, Coussement and de Leenheer8). The globe artichoke root contains inulin in a range from 30·6 to 36·7 % on a DM basis(Reference Raccuia and Melilli11) with a maximum DP of about 200(Reference Hellwege, Czapla and Jahnke12).
DP distribution and molecular weight are influenced by the harvesting conditions and additionally the extraction process could influence the molecular weight distribution or composition of the inulin and fructo-oligosaccharides(Reference Frese, Suzuki and Chatterton13). DP distribution and molecular weight in turn influence the physico-chemical properties of inulins such as solubility, thickening power and resistance to crystal formation.
Research, to date, on prebiotic potential has been limited to inulin-type fructans derived from chicory roots, whereas inulin or fructo-oligosaccharides derived from other plant sources, such as globe artichoke, have received little attention. Only recently the interest in their tubers for food purposes has increased mainly due to their content of several potentially bioactive ingredients such as inulin and fructo-oligosaccharides, naturally occurring fructose, minerals, essential amino acids, vitamins and flavonoids(Reference Kleessen, Schwarz and Boehm14).
Very-long-chain inulin (VLCI) which is extracted from globe artichoke has an average DP of between 50 and 103 and is used for inhibiting the growth of ice crystals in frozen foodstuffs, preferably frozen confections. Its DP is considerably higher than that of the long-chain inulin Beneo®HP investigated so far. Beneo®HP is extracted from chicory roots by applying physical separation techniques to eliminate all oligomers resulting in an average DP higher than 23 but mainly lower than 60.
Inulin has thus far been reported to have a positive effect on the gut microbiota at doses ranging from 4 to 40 g/d(Reference Williams, Witherly and Buddington15–Reference Den Hond, Geypers and Ghoos21). Most physiological changes that inulin induces in the host are probably effected by bacterial metabolites. The most important metabolites are the SCFA acetate, propionate and butyrate(Reference Cummings, Macfarlane and Englyst22). SCFA acidify the colonic environment, which is beneficial for certain bacteria such as bifidobacteria and lactobacilli, and detrimental to the growth of potential pathogenic species(Reference Brandt23, Reference Blaut24). SCFA are rapidly absorbed from the colon and are then metabolised in various tissues: butyrate by the colonic epithelium, propionate and acetate partly by the liver and acetate partly by muscle and other peripheral tissues(Reference Gibson and Roberfroid3, Reference Blaut24).
In the present study, the potential of VLCI to beneficially modulate the gastrointestinal microbiota and their activities was determined against maltodextrin (placebo). The objective was to assess the capacity of VLCI compared with the placebo to selectively increase numbers of bifidobacteria and lactobacilli in faeces.
Here, we present the findings of a double-blind, placebo-controlled cross-over study with thirty-two free-living healthy subjects randomised into two groups and fed either VLCI (10 g/d) or maltodextrin (10 g/d) for 3 weeks. Changes within the gut microbiota and fermentation profiles were determined in stool samples before and after treatment and gastrointestinal symptoms and stool characteristics were recorded. In order to corroborate the in vivo results on gas production without the bias of volunteer perception, the in vitro gas production kinetics in five randomly selected volunteers at baseline and after the 3 weeks of inulin or maltodextrin intake was also examined.
Materials and methods
Molecular-weight analysis of very-long-chain inulin by gel permeation chromatography with light scattering and refractive index detection (GPC-RI-MALLS system)
VLCI (no. WO/2009/000535 – Use of very long chain inulin for inhibiting the growth of ice crystals in a frozen foodstuff, patent of Bayer Cropscience AG, Monheim, Germany) and Beneo®HP (Orafti, Tienen, Belgium) were dissolved in extra-pure water at a concentration of 0·5 % (w/v). The solutions were heated at 95°C for 30 min. Then, the polymers were analysed using the following devices: Alliance chromatography system (Waters Corp., Milford, MA, USA), DAWN-EOS light scattering detector (Wyatt Technology, Santa Barbara, CA, USA) with λ0 = 658 nm and sixteen detectors in the range of angles from 14·4 to 163·3°C, K5 flow cell. The polymers were fractionated on a pre-column and three columns having the separation ranges of 300–104, 5 × 104–2 × 106 and 106–108 (SUPREMA-Gel; PSS Polymer Standards Service GmbH, Mainz, Germany). For fractionation of the samples 100 μl of this solution were injected at a temperature of 30°C and a flow rate of 0·8 ml/min with 0·05 mm-NaNO3 as the eluent. The Astra V 5.1.8.0 program (Wyatt Technology) was used to analyse the molecular-weight distribution of the samples.
Polymer chain distribution analysis of inulin by high-performance anion exchange chromatography coupled with pulsed amperometric detection (known as HPAEC-PAD)
VLCI and Beneo®HP (Orafti) inulins (5 %, w/v) were solubilised completely in boiling water and filtered through a 0·22 μm filter. Filtrates with a concentration of 0·02 % inulin were used for analysis. To map VLCI profiles and to determine the DP, a Dionex series 500 ion chromatograph (DX 500; Dionex, Idstein, Germany) coupled with a pulsed amperometric detector (Carbo-Pac PA-100 column) was used. During the analysis, the carbohydrates were eluted with 150 mm-NaOH and 1 m-sodium acetate in 150 mm-NaOH solution being used as eluents A and B, according to the following time and composition program (eluents A and B were expressed as a percentage (v/v)): 0 to 5 min, 100; 5 to 40 min, 72 and 28; 40 to 70 min, 50 and 45; 70 to 75 min, 0 and 100; 75 to 80 min, 100; 80 to 85 min, 100 and 0; 85 to 90 min, 100. The flow rate was set at 1 ml/min. All mobile phases were kept under pressure by sparging with He in order to prevent the subsequent production of carbonate, which would act as a displacing ion and reduce retention times. Chromatographic data were collected and plotted using the Dionex 500 chromatography workstation.
Subjects
Thirty-two healthy volunteers (aged 20–42 years, mean age 25 years; eighteen females, fourteen males) were recruited from the Reading area. Written consent was obtained from all participants and the study protocol was approved by the Ethics and Research Committee of the University of Reading.
Inclusion criteria for participation in the study were: aged 18–50 years; BMI of 20–30 kg/m2; good general health. Volunteers with evidence of physical or mental disease, major surgery, history of drug abuse, severe allergy or a history of severe abnormal drug reaction and smokers were excluded. Volunteers were also excluded if pregnant, lactating or planning pregnancy. Intake of an experimental drug within 4 weeks before study, former participation in a prebiotic, probiotic or laxative trial within 3 months, or use of antibiotics within 6 months before the study, chronic constipation, diarrhoea or other chronic gastrointestinal complaint (for example, irritable bowel syndrome) were all exclusion criteria. Any intake of drugs active on gastrointestinal motility, antibiotic treatment or any class of laxative was not permitted. Any medication taken throughout the duration of the study was recorded in diaries. Volunteers were instructed not to alter their usual diet or fluid intake during the trial periods; however, they were asked to refrain from consuming prebiotics and probiotics including live yoghurts and fermented milk drinks.
Dietary intervention study
The dietary intervention study was performed in a double-blind, randomised, placebo-controlled cross-over manner. Thirty-two healthy volunteers were recruited onto the study, but one volunteer dropped out due to personal reasons. For a period of 14 d before the dietary intervention study, volunteers followed a restricted diet as described earlier. Thirty-one subjects were randomly allocated into one of two groups.
The first group (n 18) consumed VLCI (10 g/d; Bayer BioScience GmbH, Hermannswerder, Potsdam, Germany) for 3 weeks, and then after a 3-week washout period, they consumed the placebo (maltodextrin; Roquette, Lestrem, France) (10 g/d) for 3 weeks. The second group (n 13) received first the placebo (10 g/d) for 3 weeks, and then after a 3-week washout, they consumed VLCI (10 g/d) for another 3-week treatment period. Each product was given for 3 weeks, followed by 3-week washout periods during which no prebiotic was consumed. The composition of globe artichoke root (cv. Madrigal; Nunhems, Haelen, The Netherlands) derived VLCI was as follows: inulin, 97·5 %, moisture, 3·1 %, sugars (glucose, fructose, sucrose), 0·2 %; ash, 0·4 %.
All test products were packaged, labelled and randomised by Bayer BioScience GmbH (Hermannswerder, Potsdam, Germany) before the study, and the study was blinded to both the investigators in the University of Reading and the volunteers. Volunteers were asked to keep diaries throughout the study to record stool frequency, consistency (constipation, hard, formed, soft or diarrhoea), abdominal pain (none, mild, moderate or severe), intestinal bloating (none, mild, moderate or severe) and flatulence (none, mild, moderate or severe) on a daily basis. Any concomitant medication and adverse events were also recorded.
Faecal samples were collected from each volunteer at five different points, before and after each treatment: at baseline, before inulin intake (pre-VLCI), during VLCI intake, at washout 1 (post-VLCI), before maltodextrin (pre-maltodextrin), during maltodextrin intake, at washout 2 (post-maltodextrin).
Collection and stool sample preparation
Freshly voided faecal samples were stored in an anaerobic cabinet (10 % H2; 10 % CO2; 80 % N2) for no longer than 2 h before processing. Samples were diluted 1:10 (w/w) with PBS (0·1 m; pH 7·0) and mixed in a Stomacher 400 (Seward, Thetford, Norfolk, UK) for 2 min at normal speed.
Enumeration of faecal microbial populations by fluorescence in situ hybridisation
Fluorescence in situ hybridisation (FISH) was performed as described by Martín-Peláez et al. (Reference Martín-Peláez, Gibson and Martín-Orúe25). The composition of hybridisation and wash buffers depends on the rRNA-targeted oligonucleotide probe as reported in probeBase (http://www.microbial-ecology.net/probebase) and was used accordingly. Briefly, samples (375 μl) of 1;10 (w/w) stool sample suspension in PBS were fixed overnight at 4°C with 4 % (w/v) filtered paraformaldehyde (pH 7·2) in a ratio of 1:4 (v/v), washed twice with filtered PBS (0·2 μm pore size), re-suspended in 300 ml of a PBS–ethanol mixture (1:1, v/v) and then stored at − 20°C for up to 3 months. For the hybridisations, 20 μl of each sample were pipetted onto Teflon- and poly-l-lysine-coated, six-well (10 mm diameter each) slides (Tekdon Inc., Myakka City, FL, USA). To permeabilise the cells for use with probe Lab158, samples were treated with 50 μl of lysozyme (1 mg/ml in 100 mm-2-amino-2-hydroxymethyl-propane-1,3-diol-HCl, pH 8·0) at 37°C for 15 min before being washed (2–3 s) in water and were finally dehydrated in the ethanol series (50 %, 80 % and 96 % (v/v) ethanol, 3 min each). The probe (5 ng/μl probe in hybridisation buffer) was applied onto the surface of each well. Hybridisations were performed for 4 h in an ISO20 oven (Grant Boekel, Cambridge, Cambs, UK). For the washing step, slides were placed in 50 ml of wash buffer containing 20 μl of 4, 6-diamidino-2-phenylindole dihydrochloride (DAPI; 50 ng/μl; Sigma, St Louis, MO, USA) for 15 min. They were then washed (2–3 s) in ice-cold water and dried under a stream of compressed air. Then 5 μl of antifade reagent (polyvinyl alcohol mounting medium with DABCO™ antifading; Sigma) were added to each well and a coverslip was applied. Slides were stored in the dark at 4°C (for a maximum of 3 d) until cells were counted under a Nikon E400 Eclipse microscope (Nikon, Kingston upon Thames Surrey, UK). DAPI slides were visualised with the aid of a DM 400 filter and probe slides with the aid of a DM 575 filter. All probes were Cy3-labelled and synthesised by Sigma Aldrich. Table 1 gives the details of probes used in the present study(Reference Langendijk, Schut and Jansen26–Reference Walker, Duncan and Leitch33).
Table 1 Probes used for fluorescence in situ hybridisation analysis of bacterial populations in human faeces

ND, no information relating to these probes has been deposited in probeBase (http://www.microbial-ecology.net/probebase).
† Probe designation according to probeBase.
‡ These probes were used together in equimolar concentrations (both at 50 ng/μl). Formamide (20 %) was included in the hybridisation buffer.
Faecal SCFA analysis
Samples (1 ml) of 1:10 (w/w) stool sample suspension in PBS were dispensed into 1·5 ml tubes and centrifuged at 13 000 g for 5 min to pellet bacteria and other solids. Supernatant fractions were filtered using 0·2 μm polycarbonate syringe filters (Whatman International Ltd, Maidstone, Kent, UK) and injected (200 μl) into an HPLC system (Merck, Whitehouse Station, NJ, USA) equipped with refractive index (RI) detection. The column used was an ion-exclusion REZEX ROA organic acid column (Phenomenex, Inc., Macclesfield, Cheshire, UK) maintained at 85°C. Sulfuric acid in HPLC-grade water (0·0025 mm) was used as the eluent and the flow rate was maintained at 0·5 ml/min(Reference Martín-Peláez, Gibson and Martín-Orúe25). The carbonic acids in the samples were quantified through calibration curves of lactic, acetic, propionic and butyric in concentrations ranging between 2·5 and 100 mm. Peaks were integrated using Atlas Lab managing software (Thermo Lab Systems, Mainz, Germany).
In vitro rate of gas production
A pressure technique(Reference Mauricio, Mould and Dhanoa34) was used to monitor the cumulative and rate of gas production in five randomly selected volunteers participating in the feeding study at baseline, and upon completion of treatments (VLCI and maltodextrin).
Pre-reduced gut model culture medium (80 ml)(Reference Macfarlane, Macfarlane and Gibson35) was dispensed into clean serum vials (125 ml working volume; Belco Glass Inc., Vineland, NJ, USA) in an anaerobic cabinet (10 % H2, 10 % CO2, 80 % N2) and sealed with a gas-impermeable butyl rubber septum and aluminium cap. Vials were autoclaved and placed overnight in a 37°C incubator. The culture medium consisted of the following (g/l) dissolved in distilled water: starch (potato), 5·0; pectin (citrus), 2·0; guar gum, 1·0; mucin (porcine gastric type III), 4·0; xylan (oatspelt), 2·0; arabinogalactan (larch wood), 2·0; inulin (Beneo®ST; Orafti), 1·0; casein (BDH Ltd, Poole, Dorset, UK), 3·0; peptone water (Oxoid, Basingstoke, Hants, UK), 5·0; tryptone (Oxoid), 5·0; bile salts no. 3 (Oxoid), 0·4; yeast extract (Oxoid), 4·5; FeSO4·7H2O, 0·005; NaCl (Fisher Scientific, Springfield, NJ, USA), 4·5; KCl (VWR International, Lutterworth, Leics, UK), 4·5; KH2PO4, 0·5; MgSO4·7H2O, 1·25; CaCl2·6H2O, 0·15; NaHCO3, 1·5; cysteine HCl, 0·8; hemin, 0·05; Tween 80, 1·0 ml; vitamin K1, 10 μl. Unless otherwise stated, all chemicals were obtained from Sigma-Aldrich (Poole, Dorset, UK).
Each vial containing a volume of 90 ml was inoculated with 10 ml of 1:10 diluted faecal slurry, prepared as above, via a sterile syringe, giving a final volume of 100 ml.
The gas production potential of the faecal flora after treatment was evaluated by recording the head space pressure (pounds per square inch; psi) of each vial. Seven replicate vials were prepared for each donor. Vials were incubated at 37°C and were continuously shaken. Pressure readings were obtained every 3 h for a 27 h fermentation period by piercing the rubber caps with a U200/66 needle adaptor connected to a pressure transducer (type 2200BGF150WD3DA; Keller Ltd, Dorchester, Dorset, UK) with a T443A digital panel meter (Bailey and Makey Ltd, Birmingham, UK). Pressure readings (psi) were converted into gas volume (ml) using an established linear regression of pressure recorded in the same vials with known air volumes at the incubation temperature. Gas production data were fitted to the model proposed by France et al. (Reference France, Dijkstra and Dhanoa36) using the program CurveExpert (version 1.37; Daniel G Hyams, Hixson, TN, USA) to provide parameters describing cumulative gas release in terms of potential gas production (A; ml), lag time (T; h), time to half-asymptote (T/2; h) and fractional rate of gas production at T/2 (μ; per h).
Statistical analysis
A general linear model was used to test for time-dependent effects (Genstat® for Windows® 8th edition; VSN International, Hemel Hempstead, Herts, UK). When there was no significant effect for ‘sequence’ all time points were included and subsequent statistical analyses were performed using one-way ANOVA, including post hoc tests appropriate for the individual datasets (Tukey's post test with significance set at P < 0·05). We used GraphPad Prism (version 5·0; Graph-Pad Software, Inc., San Diego, CA, USA) for FISH, SCFA and gas production analysis.
The occurrence and intensity of gastrointestinal symptoms were scored in the run-in period during the intervention period (3 weeks). Volunteers were requested to rank their wellbeing with respect to gastrointestinal complaints, such as abdominal pain, intestinal bloating, flatulence, nausea, borborygmi, diarrhoea and constipation. The respective ranks given were: 0 = none; 1 = mild; 2 = moderate; 3 = severe. Moreover, stool frequency and consistency were also monitored and ranked as follows: 0 = watery/diarrhoeal; 1 = soft; 2 = normal; 3 = hard. Statistical analyses were also conducted using GraphPad Prism (version 5; Graph-Pad Software, Inc.).
The average cumulative gas production in vitro was fitted iteratively to the model(Reference France, Dijkstra and Dhanoa36), using the program CurveExpert (version 1.37; Daniel G Hyams).
Results
Molecular-weight analysis of very-long-chain inulin by gel permeation chromatography with light scattering and refractive index detection (GPC-RI-MALLS system)
GPC-RI-MALLS chromatograms of VLCI and commercially available chicory root inulin (Beneo®HP) are shown in Fig. 1. It is obvious that VLCI has a markedly higher molecular-weight distribution compared with Beneo®HP. The average DP of VLCI was>55 as compared with>23 in the case of Beneo®HP.

Fig. 1 Gel permeation chromatography with light scattering and refractive index detection (GPC-RI-MALLS system; Mainz, Germany). (—), Beneo®HP (Orafti, Tienen, Belgium); (- - -), globe artichoke (Cynara scolymus) inulin (very-long-chain inulin).
Polymer chain distribution analysis of inulin by high-performance anion exchange chromatography coupled with pulsed amperometric detection (known as HPAEC-PAD)
The inulin profiles of chicory (Beneo®HP) and globe artichoke tubers (VLCI) were separated into defined DP ranges. As shown in Fig. 2, chicory inulin (Fig. 2(a)) contains a much higher amount of shorter molecules as compared with globe artichoke inulin (Fig. 2(b)) and the range of high-DP species detected in the VLCI profile clearly exceeds that of chicory.

Fig. 2 High-performance anion-exchange chromatography with pulsed amperometric detection (DX 500 Dionex, Idstein, Germany) of (a) Beneo®HP (Orafti, Tienen, Belgium) and (b) globe artichoke (Cynara scolymus) inulin (very-long-chain inulin).
Faecal microflora
The main objective of the study was to assess the impact of consumption of inulin extracted from globe artichoke on the intestinal microbiota. Fourteen 16S rRNA-based FISH probes were used to target predominant groups or species of human faecal bacteria (Table 1) covering 70–80·6 % of total bacteria detected in stool samples. Bacterial numbers at the end of each treatment period – either 10 g VLCI/d or 10 g maltodextrin/d – were compared with samples obtained before treatment (pre-) and after washout (post-) of the respective treatment. Statistical significance of the results was evaluated using one-way ANOVA with Tukey's post test (95 % CI). Average faecal bacterial numbers for thirty-one healthy volunteers over the trial period as determined by FISH and expressed as log10 cells/g faeces (n 31) are shown in Table 2.
Table 2 Average faecal bacterial numbers for thirty-one healthy volunteers over the trial period as determined by fluorescence in situ hybridisation (FISH), expressed as log10 cells/g faeces
(Mean values and standard deviations)

VLCI, very-long-chain inulin.
* Mean value was significantly different from that for pre-VLCI (P < 0·05; Tukey's post test).
† Mean value was significantly different from that for VLCI (P < 0·05; Tukey's post test).
‡ Mean value was significantly different from that for post-VLCI (P < 0·05; Tukey's post test).
§ Mean value was significantly different from that for pre-maltodextrin (P < 0·05; Tukey's post test).
∥ Mean value was significantly different from that for maltodextrin (P < 0·05; Tukey's post test).
Total bacterial levels (detected by Eub I-II-III) remained unaffected by intervention. A significant increase in Bifidobacterium numbers was observed during inulin intake compared with samples taken immediately before inulin intervention (2·82-fold increase; P < 0·05). There was also a significant difference in bacterial population levels in faeces when comparing inulin and maltodextrin treatment samples. Numbers of bifidobacteria were significantly higher following ingestion of inulin compared with the maltodextrin treatment period (2·75-fold increase; P < 0·001).
Numbers of faecal lactobacilli–enterococci increased significantly with ingestion of VLCI compared with pre-inulin levels (2·42-fold increase; P < 0·01) and the increase was even more pronounced when compared with maltodextrin (5·88-fold increase; P < 0·05) in thirty-one volunteers. During the ingestion of maltodextrin, the Lactobacillus–Enterococcus group significantly decreased with respect to pre-maltodextrin (2·64-fold decrease; P < 0·001).
No significant changes were observed between pre-treatment and post-treatment samples for bifidobacteria as well as for lactobacilli counts.
A significant increase in the Atopobium group was observed (2·83-fold increase; P < 0·05) whereas numbers of the Bacteroides–Prevotella group were reduced (1·77-fold decrease; P < 0·05) after consumption of VLCI compared with maltodextrin (Table 2).
No differences were noted in the levels of Escherichia coli and the Eubacterium rectale–Clostridium coccoides group enumerated by the Erec482 probe. The Ehal1469 probe specific for subgroups within the Erec482 (Clostridium cluster XIVa) group did not show significant differences between treatment and placebo. Similarly, the butyrate-producing E. rectale–Roseburia group, also a component of cluster XIVa, was present at no more than 10 % of the total E. rectale–C. coccoides group at all time points analysed, when enumerated by Rrec584.
Cluster IX representatives (detected by Prop853) and Ruminococcus numbers (Clostridium cluster IV, detected by the Rfla729/Rbro730 probes) did not exhibit any statistically significant changes upon ingestion either of the placebo or VLCI (P>0·05). The Clit135 probe, specific for some members of the C. lituseburense group (C. lituseburense group – cluster XI) and C. histolyticum accounted for no more than 1 % of the total faecal bacteria population.
Specific members of Clostridium cluster XVI (detected by the Ecyl387 probe) did not exhibit changes throughout the duration of the intervention study (P>0·05).
SCFA concentrations
Table 3 shows the concentrations of faecal SCFA. No significant changes in faecal concentrations of acetic, propionic and butyric acids were observed over the course of the trial, neither between pre-inulin and inulin and maltodextrin and pre-maltodextrin nor when inulin or maltodextrin were compared.
Table 3 SCFA concentrations measured by HPLC present in faecal samples collected from thirty-one volunteers over the course of the trial
(Mean values and standard deviations)
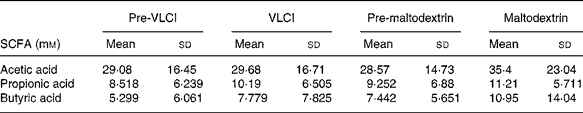
VLCI, very-long-chain inulin.
Analysis of bowel habit and gastrointestinal symptoms
Table 4 summarises data on bowel habit and gastrointestinal symptoms, as recorded by the volunteers during intake of VLCI or maltodextrin. Stool frequency did not differ during inulin ingestion compared with placebo. Stool consistency, qualitatively graded by volunteers as hard, formed, or soft, varied greatly between individuals. No diarrhoea was reported by any subjects, while there was a significant increase in bloating upon ingestion of VLCI (P < 0·05). However, on average, even the increased bloating was still rated as mild. There were no significant differences between treatments with regard to flatulence and abdominal pain. None of the gastrointestinal symptoms was severe enough for volunteers to discontinue the study.
Table 4 Summary of bowel habit and gastrointestinal symptom data for thirty volunteers over the course of the trial†
(Mean values and standard deviations)
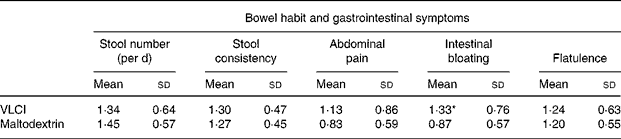
VLCI, very-long-chain inulin.
* Mean value was significantly different from that for maltodextrin (P < 0·05; paired t test, two-tailed).
† Analysis was performed by allocating numbers to defined conditions. Stool consistency: hard = 0; formed = 1; soft = 2. Abdominal pain, bloating and flatulence: none = 0; mild = 1; moderate = 2; severe = 3.
Gas production kinetics
Fig. 3 shows the cumulative gas production at 3 h intervals for a 27 h fermentation period, in five randomly selected volunteers participating in the feeding study upon completion of treatments with VLCI and maltodextrin. In general, VLCI showed higher gas volumes than maltodextrin from 5 h onwards. There was a significant increase in the rate of gas production pattern during the first 5 h of in vitro fermentation, in particular with faecal samples after the intake of VLCI compared with the maltodextrin (Fig. 3).

Fig. 3 Cumulative gas production over 27 h with maltodextrin (a) and very-long-chain inulin (b). Values are means of five volunteers (seven replicates for each volunteer on each respective treatment). (![]() ), Volunteer 1; (
), Volunteer 1; (![]() ), volunteer 2; (
), volunteer 2; (![]() ), volunteer 3; (
), volunteer 3; (![]() ), volunteer 4; (
), volunteer 4; (![]() ), volunteer 5.
), volunteer 5.
Both treatment groups appeared to generate gas at 3 h after inoculation of faecal slurry and up to 27 h (Fig. 3). The cumulative gas production after maltodextrin treatment was significantly lower up to 27 h fermentation as compared with VLCI. Apart from volunteer 4, samples of the other volunteers generated similar levels of cumulative gas production during the 24 h fermentation period. Volunteer 4 had markedly higher rates of gas production compared with the other donors.
The gas production pattern over the 27 h of incubation and the results of the cumulative gas production are presented in Table 5. Samples after VLCI intervention generated higher gas volume production (A) than after maltodextrin intervention with lower lag time and fractional degradation rate at 12 and 24 h. These data are in accordance with bloating ratings recorded in the volunteers' diaries during the course of the trial.
Table 5 Gas production kinetics obtained by the in vitro fermentation of very-long-chain inulin (VLCI) and maltodextrin*

a,b,c,d Values within a column with unlike superscript letters were significantly different (P < 0·05).
* Parameters obtained from the adjustment of gas production by France et al. (Reference France, Dijkstra and Dhanoa36). Potential gas production (A; ml); lag time (T; h), time to half-asymptote (T/2; h) and fractional rate of gas production at T/2 (μ; per h).
Discussion
The bifidogenic nature of chicory inulin is very well established(Reference Gibson and Roberfroid3, Reference Kleessen, Sykura and Zunft18, Reference Bouhnik, Flourié and Ouarne37–Reference Takahashi44). Positive shifts in the composition of microflora have been shown in vivo in human studies at doses ranging from 4 to 40 g/d(Reference Williams, Witherly and Buddington15–Reference Den Hond, Geypers and Ghoos21). While the majority of studies to date have investigated the effect of chicory inulin on the human gut microbiota using culture-based methodologies(Reference Gibson and Roberfroid3, Reference Kleessen, Sykura and Zunft18, Reference Bouhnik, Vahedi and Achour19, Reference Ramirez-Farias, Slezak and Fuller45, Reference Brighenti, Casiraghi and Canzi46), more recent studies have applied molecular methods such as FISH and real-time PCR for the bacterial analyses(Reference Kleessen, Schwarz and Boehm14, Reference Yap, Mohamed and Husni47–Reference Kolida, Meyer and Gibson55).
However, the in vivo effect of globe artichoke-derived inulin in human subjects has not been studied to date. It is clear that globe artichoke inulin differs in DP distribution as well as in average DP(Reference Van Loo, Coussement and de Leenheer8). Recent in vitro studies reported that DP may be a factor affecting not only the prebiotic efficacy of a substrate, but also its persistence in the colon and the manifestation of undesirable gastrointestinal side effects(Reference Van De Wiele, Boon and Possemiers56, Reference Hughes, Shewry and Li57). The inulin with the highest average DP to be studied in vivo in human subjects so far has been Beneo®HP(Reference Kleessen, Sykura and Zunft18, Reference Tuohy, Kolida and Lustenberger54).
Using direct molecular-based enumeration of faecal bacteria, we have shown that VLCI derived from globe artichoke selectively stimulates the levels of faecal bifidobacteria. This is concordant with other in vivo intervention studies with fructans which often reported increases between 0·5–1·0 log10 bifidobacteria counts(Reference Kolida and Gibson58). However, the magnitude of increase seems to depend on the baseline values, and higher increases are often observed in volunteers with low initial bifidobacteria numbers(Reference Kolida, Meyer and Gibson55). While the bifidogenic potential is well proven, the effect of consumption of inulin on other bacterial groups is less well established. A decrease in Clostridia was mainly reported by culture-based bacteria analysis(Reference Gibson and Roberfroid3, Reference Yap, Mohamed and Husni47); however, a recent study which used FISH also found decreased bacteria numbers for the C. histolyticum group and C. lituseburense group after inulin ingestion(Reference Kleessen, Schwarz and Boehm14). In the same study(Reference Kleessen, Schwarz and Boehm14) there was also a decrease in Bacteroides numbers after inulin consumption. We also found that Bacteroides numbers were significantly lower after VLCI than after maltodextrin intervention. Additionally, we observed a significant increase in lactobacilli–enterococci after VLCI treatment. While there was a small increase after inulin consumption in one human intervention trial(Reference Ramnani, Gaudier and Bingham59) and some indication from in vitro fermentation studies with inulins and fructo-oligosaccharides(Reference Probert, Apajalahti and Rautonen60, Reference Gomez, Tuohy and Gibson61) that the lactobacilli–enterococci group could be influenced by inulin, this is the first time that such a high increase in this bacterial group has been reported for inulin in a human intervention study. The only other bacterial group in the present study being stimulated after VLCI ingestion was Atopobium. The role of Atopobium in the human colon is not clear. However, they are commonly isolated from healthy human faeces and are present at 1·5–3 % of the total faecal populations(Reference Leitch McWilliam, Walker and Duncan62). It is not clear from the results of the present study whether Atopobium can ferment VLCI or if the increase is due to cross-feeding between different bacterial groups. It is noteworthy that the increase in bifidobacteria and lactobacilli–enterococci in the present study were paralleled by a decrease in the levels of potentially harmful bacteria such as Bacteroides–Prevotella. As total bacterial levels remained unchanged throughout the study, changes upon VLCI ingestion appear to be inter-population only.
No differences were observed in faecal SCFA throughout the study. This is not surprising, as over 95 % of SCFA produced in the human large intestine are thought to be rapidly absorbed within the colon, and only a small proportion of all SCFA produced in the large bowel is excreted in the faeces(Reference Salminen, Bouley and Boutron-Ruault63, Reference Coussement64).
Several studies have shown that fermentation of inulin-type fructans may stimulate bowel movements and may normalise stool frequency(Reference Kleessen, Sykura and Zunft18–Reference Bouhnik, Vahedi and Achour19, Reference Den Hond, Geypers and Ghoos21). Increased bloating and mild flatulence upon inulin-type fructan ingestion is a well-known and accepted side effect(Reference Bouhnik, Vahedi and Achour19, Reference Kruse, Kleessen and Blaut20, Reference Coussement64–Reference Van den Heuvel, Schaafsma and Muys67).
Analysis of the gastrointestinal symptom volunteer diaries indicates a significant increase in intestinal bloating during VLCI intake in healthy volunteers. The formation of H2, which is a metabolic endproduct of bacterial fermentation in the colon, is probably the major cause of this symptom(Reference Kleessen, Schwarz and Boehm14).
However, it has to be emphasised that bifidobacteria, the numbers of which increased significantly with inulin, are not gas producers. Several studies have reported increased gas production upon the ingestion of inulin, which is not accompanied by a concomitant increase in gas-producing bacterial groups and the authors have commonly attributed this effect to consumer perception(Reference Tuohy, Kolida and Lustenberger54, Reference Kolida, Meyer and Gibson55). Therefore the gas-generating potential of samples obtained from a group of randomly selected volunteers was also followed in vitro, at baseline and at the end of each treatment.
The in vitro observations confirmed the in vivo reports, as after ingestion of VLCI the rate of gas production in faeces increased and led to higher cumulative gas levels compared with the maltodextrin faeces. VLCI selectively increased the faecal levels of bifidobacteria and lactobacilli–enterococci in vivo, whilst Clostridia, which are prolific gas producers in faeces, remained stable. However, increased gas production may be due to increased or altered metabolic activity of these bacteria facilitated probably by a higher availability of potential substrates such as acetate. Bacterial enumeration cannot account for possible increases in metabolic activity. Furthermore, there may be shifts at species levels promoting the growth of strains that possess higher gas-producing capacities. Further clarification of the relationship between specific intestinal bacteria and gas production as a result of colonic fermentation is required. In general, both maltodextrin and VLCI treatments were well accepted by the human volunteers and all subjects completed the study without severe side effects being noted.
The prebiotic ability of a VLCI may have further implications in vivo. While short-chain fructans are more rapidly fermented(Reference Gomez, Tuohy and Gibson61), higher molecular weight and low solubility may improve persistence in the gut(Reference Van De Wiele, Boon and Possemiers56, Reference Roland, Nugon-Baudon and Andrieux68). Thus, due to its physico-chemical characteristics – high molecular weight and low solubility – VLCI is expected to prolong saccharolytic fermentation into distal colonic areas and in vivo animal studies are under way to investigate the effect of VLCI on the site of fermentation and the selectivity compared with Beneo®HP.
In conclusion, the present study has demonstrated the prebiotic efficacy of VLCI from globe artichoke, which combined significant increases in the levels of both bifidobacteria and lactobacilli in a selective manner with little adverse gastrointestinal effects at 10 g/d.
Acknowledgements
We would like to extend our gratitude to the volunteers who participated in the present study. The study was financially supported by Bayer Group.
A. C. was the principal investigator for conducting the experiment and writing the manuscript. A. K. contributed to data interpretation, manuscript preparation and the literature search. S. K. contributed to the analysis of in vitro rate of gas production. G. R. G. supervised the study design along with drafting the manuscript. E. G. contributed to the analysis of VLCI by the GPC-RI MALLS and HPAEC-PAD methods. V. L. was the overall coordinator of the VLCI project. M. B. was responsible for the preparation of the VLCI material (harvest, isolation, purification, quality check). C. F. was involved in the functionality testings of VLCI. All other authors provided suggestions, alterations to the manuscript and comments for the analyses.
E. G., C. F., M. B. and V. L. were Bayer Group (Bayer BioScience GmbH and Bayer Cropscience AG) employees at the time of the study who functioned as coordinators of this sponsored project. There were no conflicts of interest that affected the design, the execution of the study protocol, analyses or interpretation of the data. None of the other authors has any conflicts of interest to declare.








