

Grazed grass is a low-cost feed for cattle and offers an economic advantage to regions capable of long grazing seasons and high productivity of DM per hectare( Reference O'Donovan, Lewis and O'Kiely 1 , Reference Ferris 2 ). For this reason, pasture-based systems predominate ruminant production in certain countries, such as Ireland, New Zealand and parts of the UK( Reference O'Donovan, Lewis and O'Kiely 1 – Reference Clark, Caradus and Monaghan 3 ), thus enhancing the competitiveness of dairy and beef industry, preserving rural landscapes and improving the image of milk and meat production for consumers( Reference Dijkstra, Kebreab and Bannink 4 ).
However, fresh grass-based diets provide lower amounts of glucogenic nutrients( Reference Dijkstra, Kebreab and Bannink 4 ), than diets rich in concentrate feeds, and energy supply is challenging in grazing systems, because of the high contribution of pasture in the diets. Efficient assessment of the feeding value of grass is crucial in order to avoid excessive over-supply that would increase production costs, or under-supply that would reduce productivity. Developing tools to predict metabolisable energy (ME) in fresh grass may greatly improve profitability of pasture-based systems, with a 180 000 MJ/ha of ME being recommended as the production target( Reference Clark, Caradus and Monaghan 3 ). Parameters such as digestible organic matter in DM (DOMD) and digestible energy (DE), predicted from contents of crude protein, neutral-detergent fibre (NDF), diethyl ether extract (EE), total digestible crude protein (tdCP), total digestible neutral-detergent fibre (tdNDF) and fatty acids of grass have been used to predict grass ME concentrations( Reference Dijkstra, Kebreab and Bannink 4 – 6 ). In the current UK feeding system( 6 ), DOMD is used to predict grass ME content, but the different proportions of digestible nutrients in DOMD are not accounted for( Reference Yan and Agnew 7 ). In addition, a number of limitations on the prediction of ME content of fresh grass raise concerns about the accuracy of ME predictions from grass nutrient contents and digestibility. The majority of the works on the prediction of ME content of grass (1) have been performed with diets based on conserved forages and concentrates, (2) they relied on prediction, instead of measurement, of methane emissions, and (3) they carried out the experiments with sheep instead of cattle( 6 , Reference Jarrige 8 ). Also, modern cows have higher production potential and maintenance requirements compared with those used to develop the existing energy systems( Reference Dijkstra, Kebreab and Bannink 4 , Reference Bruinenberg, Zom and Valk 9 ), and this may contribute further to the potential error when estimating grass ME contents. A previous work has reported discrepancies in current energy evaluations, when diets rich in good quality grass (>80 % of DM intake) were offered( Reference Bruinenberg, Zom and Valk 9 ). Consequently, the development of a quick and accurate assessment methodology for fresh grass ME contents, which would overcome these limitations and account for the various nutrients and their digestibility in fresh grass, seems to be a necessity for grazing cattle. The importance of using a greater range of grass nutrients in the evaluation of grass ME content has also been raised in previous reports( 6 ).
The objective of this study was therefore to develop predictions of grass energy contents, in particular ME, from grass nutrient contents and digestibility parameters for animals fed solely fresh-cut grass, which can be used for (1) the quick estimation of pasture ME contents under commercial conditions, e.g. using routinely available near-IR spectroscopy data and (2) the revision of existing feeding tables used in the development of grazing ruminant diets.
Experimental design and methods
The present study was performed under the regulations of the Department of Health, Social Services, and Public Safety of Northern Ireland, in line with the Animal (Scientific Procedures) Act 1986( 10 ). In total, 1392 records collected over three digestibility trials from consecutive grazing seasons and 3-d averaged data (n 464) were used in the dataset to develop prediction equations. Grass used was grown on ten swards in total representing different years (2007, 2008, 2009), harvest seasons (primary growth, first regrowth, second regrowth), maturity stages (3–9 weeks) and combination of ryegrass varieties (Aberstar, Aberzest, Fetione, Magella, Merbo, Merlinda and Spelga). In 2007, a first regrowth and a second regrowth swards were fertilised with 8 and 54 kg N/ha respectively. In 2008, two primary growth swards were fertilised with 45 and 70 kg N/ha, two first regrowth swards with 40 and 65 kg N/ha and two second regrowth swards with 35 and 60 kg N/ha respectively. In 2009, two primary growth swards were fertilised with 59 kg N/ha. Sward characteristics and management throughout the 3-year experiment have been described in detail by Stergiadis et al. ( Reference Stergiadis, Cheng and Allen 11 ). In total, eight groups of four cows (plus one substitute cow in 2008, n 33) were used to assess energy concentrations of fresh-cut grass. In 2007, two groups of four cows (n 8) assessed early and late maturity grass from a primary and a secondary growth swards, with the group assessing early maturity in the first regrowth sward being used to assess late maturity in the second regrowth sward and vice versa for the second group of cows. In 2008, four groups of four cows (plus one substitute cow, n 17) were used to assess early and late maturity swards during primary growth, first regrowth and second regrowth swards, with either medium (35–45 kg N/ha) or high (60–70 kg N/ha) N fertiliser input. Two groups were fed early maturity grass and two groups late maturity grass respectively throughout the experiment, while the other groups were swapping between early and late maturity grass over the three harvest dates. The two groups within the same maturity treatment received grass from medium or high fertiliser input, and they were swapped within this treatment between the three harvest dates. In 2009, two groups of four cows (n 8) assessed early and late maturity grass respectively, from a primary growth sward. For each group, daily recording of feed intake and total collection of faeces and urine were carried out for three consecutive weeks, following an adaptation period of solely fresh grass diets of at least two weeks. Detailed experimental design and sample collection and analytical procedures, were presented by Stergiadis et al. ( Reference Stergiadis, Cheng and Allen 11 , Reference Stergiadis, Allen and Chen 12 ). Details on animal data, grass chemical composition, nutrient digestibility and energy inputs/outputs and concentrations recorded over the 3 years of the present experiment are given in Table 1.
Table 1 Animal data, grass chemical composition (n 116), nutrient digestibility and energy intakes/outputs and concentrations (n 464) recorded over 3 years

Min, minimum value observed; Max, maximum value observed; GE, gross energy; DE, digestible energy; ME, metabolisable energy.
* Methane emissions were predicted from equations developed during the first year of the present study as in Yan & Mayne( Reference Yan and Mayne 13 ).
As a result of the contrasting sward characteristics and management, a large diversity of grass nutrient composition was observed; for example, grass N, water-soluble carbohydrates (WSC) and NDF contents ranged between 10·7–38·1, 70–272, and 413–601 g/kg DM, respectively. Similarly, gross energy (GE), DE and ME contents of grass ranged between 16·9–19·1, 11·8–16·7 and 9·0–14·1 MJ/kg DM, respectively. Concentrations of DOMD were between 0·636 and 0·851 kg/kg DM. Concentrations of DE, DOMD and DM, N, GE, NDF, acid-detergent fibre (ADF) and organic matter digestibilities were measured in metabolism units with non-pregnant non-lactating dairy cows fed at maintenance energy level. The ME concentration was estimated by extracting measured urine energy output and predicted methane energy output from measured DE intake. Methane emissions from all individual cows were also measured for two to three periods (1 d for adaptation and 2 d for measurements in each period) in the 2007 trials of the present study (n 40 data). The equations used to predict methane energy output from grass ADF, GE, DM, and EE contents (R 2 0·77) were previously presented by Yan & Mayne( Reference Yan and Mayne 13 ).
In order to assess feed quality of grass for immediate development of accurate diets that meet maintenance energy requirements, fresh grass DM concentration was estimated by microwaving at full power for 3–5 min. Grass ME was then predicted by near-IR spectroscopy( Reference Agnew, Park, Mayne, Rook and Penning 14 ), using a NIRS™ 5000/6500 Feed and Forage Analyser (FOSS), and ME concentration was estimated using the average value of samples collected daily over the previous two consecutive days. Maintenance ME requirements were estimated as described by Agnew et al. ( Reference Agnew, Yan, France and Thomas 15 ) (0·65 MJ/kg of metabolic body weight, kg0·75).
Statistical analysis
Analyses of data were carried out in GenStat( Reference International 16 ). The residual diagnostics were assessed using normality plots and were found to be acceptable. Regression equations were developed using residual maximum likelihood (REML)( Reference Robinson 17 , Reference Searle, Casella and McCulloch 18 ), so that the potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety could be accounted for. Linear regression relationships were established, where the responses were DE, ME and ratios of DE:GE or ME:GE; the fixed terms were (1) digestibilities of N (Nd), gross energy (GEd), NDF (NDFd), ADF (ADFd), DM (DMd) and organic matter (OMd) and DOMD, (2) grass contents of N, GE, WSC, EE, NDF, ADF and ash, and (3) tdCP and tdNDF; they were all developed either in univariate (equation 1) or multivariate (equation 2) linear models.
 $$ Y = a + bx , $$
$$ Y = a + bx , $$
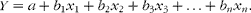 $$ Y = a + b _{1} x _{1} + b _{2} x _{2} + b _{3} x _{3} + \ldots + b _{ n } x _{ n }. $$
$$ Y = a + b _{1} x _{1} + b _{2} x _{2} + b _{3} x _{3} + \ldots + b _{ n } x _{ n }. $$
Total digestible amounts for each nutrient (kg/kg DM) were generated, separately for N and NDF, by multiplying grass nutrient content (kg/kg DM), measured by wet chemistry analysis, with nutrient digestibility (kg/kg), which represents the proportion of ingested nutrients not excreted in faeces, measured during the digestibility experiments. Candidate nested models of the random variation, with the same fixed-effect model, were compared using the deviance. The change in deviance between the nested random models was assessed using χ2 with the df given by the difference in df of the two models. The significance of the fixed terms was assessed using the Wald statistic. The squared correlation of the response and the fitted values (R 2) to represent the amount of variability explained was also derived.
An internal evaluation, using the methods of Yan et al. ( Reference Yan, Frost and Agnew 19 ) and Stergiadis et al. ( Reference Stergiadis, Allen and Chen 12 ) was performed to validate all prediction equations developed in the present study (Tables 2–5). The whole dataset (n 464) was divided into two sub-datasets of n 309 (two-thirds of the total data) and n 155 (one-third of the total data). The former dataset was used to develop equations (Table 6), applying models of the same random and fixed factors combinations to those produced from the whole dataset, while the latter was used to evaluate the precision of these new equations (Table 7). This was assessed by using the mean-square prediction error (MSPE) technique (equation 3).
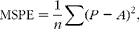 $$MSPE = \frac {1}{ n }\sum ( P - A )^{2}, $$
$$MSPE = \frac {1}{ n }\sum ( P - A )^{2}, $$
where P and A are the predicted and actual values, respectively, and n is the number of paired values for P and A compared. Mean prediction error (MPE) was used to describe the prediction accuracy (equation 4):
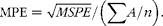 $$MPE = \sqrt { MSPE }/(\sum A / n ). $$
$$MPE = \sqrt { MSPE }/(\sum A / n ). $$
Table 2 Univariate linear prediction of energy contents and ratios in fresh grass (with non-pregnant non-lactating cows fed at maintenance energy level, n 464), using nutrient digestibility parameters

DE, grass digestible energy content; ME, grass metabolisable energy content; GE, grass gross energy content; R 2, pseudo-correlation coefficient; MPE, mean prediction error; DMd, DM digestibility; OMd, organic matter digestibility; DOMD, digestible organic matter in DM; GEd, gross energy digestibility.
* Units: MJ/kg DM for DE, ME and GE; kg/kg for DMd and OMd; kg/kg DM for DOMD; MJ/MJ for GEd. Values in subscript parentheses represent standard error of means. The effect of all explanatory variables was significant according to the Wald statistic (Fpr < 0·05). A combination of potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety was accounted for accordingly for each predicted variable.
† MPE derived from an internal validation with new equations, listed in Table 6, which were developed from two-thirds of the whole dataset and by using the exact model presented in the current table; the new equations were validated against the remaining one-third of the whole dataset.
Table 3 Multivariate linear prediction of energy contents and ratios in fresh grass (with non-pregnant non-lactating cows fed at maintenance energy level, n 464), using nutrient digestibility and grass chemical composition parameters

DE, grass digestible energy content; ME, grass metabolisable energy content; GE, grass gross energy content; R 2, pseudo correlation coefficient; MPE, mean prediction error; DMd, DM digestibility; N, grass nitrogen content; EE, grass diethyl ether extract content; ADF, grass acid-detergent fibre content; OMd, organic matter digestibility; WSC, grass water-soluble carbohydrate content; DOMD, digestible organic matter in DM; GEd, gross energy digestibility; NDF, grass neutral-detergent fibre content.
* Units: MJ/kg DM for DE, ME and GE; kg/kg for DMd and OMd; kg/kg DM for N, EE, ADF, Ash, WSC, DOMD and NDF; MJ/MJ for GEd. Values in subscript parentheses represent standard error. The effect of all explanatory variables was significant according to the Wald statistic (Fpr < 0.05). A combination of potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety was accounted for accordingly for each predicted variable.
† MPE derived from an internal validation with new equations, listed in Table 6, which were developed from two-thirds of the whole dataset and by using the exact model presented in the current table; the new equations were validated against the remaining one-third of the whole dataset.
Table 4 Univariate and multivariate linear prediction of metabolisable energy (ME) content in fresh grass (with non-pregnant non-lactating cows fed at maintenance energy level, n 464), using digestible energy (DE) and grass chemical composition parameters

R 2, pseudo correlation coefficient; MPE, mean prediction error; N, grass nitrogen content; EE, grass diethyl ether extract content; WSC, grass water-soluble carbohydrate content; ADF, grass acid-detergent fibre content.
* Units: MJ/kg DM for ME and DE; kg/kg DM for N, EE, WSC and ADF. Values in subscript parentheses represent standard error. The effect of all explanatory variables was significant according to the Wald statistic (Fpr < 0.05). A combination of potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety was accounted for accordingly for each predicted variable.
† MPE derived from an internal validation with new equations, listed in Table 6, which were developed from two-thirds of the whole dataset and by using the exact model presented in the current table; the new equations were validated against the remaining one-third of the whole dataset.
Table 5 Multivariate linear prediction of energy contents in fresh grass (with non-pregnant non-lactating cows fed at maintenance energy level, n 464), using total digestible crude protein (tdCP), total digestible neutral-detergent fibre (tdNDF) and grass chemical composition parameters

DE, grass digestible energy content; ME, grass metabolisable energy content; R 2, pseudo correlation coefficient; MPE, mean prediction error; WSC, grass water-soluble carbohydrate content; EE, grass diethyl ether extract content.
* Units: MJ/kg DM for DE and ME; g/100g DM for tdCP and tdNDF; kg/kg DM for WSC and EE. Values in subscript parentheses represent standard error. The effect of all explanatory variables was significant according to the Wald statistic (Fpr < 0.05). A combination of potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety was accounted for accordingly for each predicted variable.
† MPE derived from an internal validation with new equations, listed in Table 6, which were developed from two-thirds of the whole dataset and by using the exact model presented in the current table; the new equations were validated against the remaining one-third of the whole dataset.
Table 6 Internal validation: univariate and multivariate linear prediction of energy contents and ratios in fresh grass (with non-pregnant non-lactating cows fed at maintenance energy level, n 464), from nutrient digestibility, grass chemical composition parameters, digestible energy (DE), total digestible crude protein (tdCP) and total digestible neutral-detergent fibre (tdNDF), using two-thirds of the whole dataset (n 309)

ME, grass metabolisable energy content; GE, grass gross energy content; R 2, pseudo correlation coefficient; MPE, mean prediction error; DMd, DM digestibility; OMd, organic matter digestibility; DOMD, digestible organic matter in DM; N, grass nitrogen content; EE, grass diethyl ether extract content; ADF, grass acid-detergent fibre content; WSC, grass water-soluble carbohydrate content; GEd, gross energy digestibility; NDF, grass neutral-detergent fibre content.
* Original equations are presented in Tables 2–5 and were developed by using the whole dataset.
† Units: MJ/kg DM for DE, ME and GE; kg/kg for DMd and OMd; kg/kg DM for DOMD, N, EE, ash, ADF, WSC and NDF; g/100g DM for tdCP and tdNDF; MJ/MJ for GEd. Values in subscript parentheses represent standard error. The effect of all explanatory variables was significant according to the Wald statistic (Fpr < 0.05), except for the effects of EE in equations P, S, AK and NDF in equation Q which only tended to be significant (0.05 < P< 0.10). A combination of potential random effects of cow, field, year, harvest date, fertiliser input, maturity stage and grass variety was accounted for accordingly for each predicted variable.
Table 7 Internal validation of equations developed from two-thirds of the whole dataset (n 309) using the remaining one-third of the whole dataset (n 155)

r 2, correlation between predicted and actual values; MPE, mean prediction error; R c, Lin's concordance correlation coefficient; Min, minimum value observed; Max, maximum value observed; DE, grass digestible energy content; ME, grass metabolisable energy content; GE, grass gross energy content.
* Equations are presented in Table 6.
† Units: MJ/kg DM for DE, ME and GE.
The same one-third of the present dataset was also used to validate equations of Givens et al. ( Reference Givens, Everington and Adamson 20 ), Terry et al. ( Reference Terry, Osbourn and Cammell 21 ), National Research Council (NRC)( 5 ) and Agricultural and Food Research Council( 6 ) for the prediction of ME concentrations, using digestibility and grass chemical composition parameters or DE (Table 8). In addition, Lin's concordance correlation coefficient was generated in order to quantify the agreement between in vivo measured ME and ME predicted by equations developed in the present (Table 7) or previously published( 5 , Reference Givens, Everington and Adamson 20 , Reference Terry, Osbourn and Cammell 21 ) (Table 8) studies, and part of this analysis is presented in Figs. 1–3.
Table 8 Validation of equations previously published for the prediction of metabolisable energy (ME) from grass chemical composition, digestible energy (DE) and digestible organic matter in DM (DOMD), using one-third of the whole dataset (n 155)

r 2, correlation between predicted and actual values; MPE, mean prediction error; R c, Lin's concordance correlation coefficient; Min, minimum value observed; Max, maximum value observed; CP, grass crude protein content; NDF, grass neutral-detergent fibre content; ADF, grass acid-detergent fibre content.
* Equations AM, AN, AO, AR, AS, AT were published by Givens et al. ( Reference Givens, Everington and Adamson 20 ), equations AP and AU by Terry et al. ( Reference Terry, Osbourn and Cammell 21 ), equation AQ by National Research Council( 5 ) and equation AV from Agricultural and Food Research Council( 6 ). Units: MJ/kg DM for ME and DE; kg/kg DM for CP, NDF, ADF and DOMD. In equation AQ, 1 Mcal = 4·184 MJ was used when revising the original model to represent MJ( 6 ). In equations AR and AS, DOMD was measured by NDF-cellulase and pepsin-cellulase, respectively, and in equations AT and AU, DOMD was measured by rumen fluid.

Fig. 1 Bland–Altman plots showing the agreement between in vivo measured metabolisable energy (ME) and residual (predicted minus actual) ME, with ME being predicted from equations published in other studies ((a) Givens et al. ( Reference Givens, Everington and Adamson 20 ); (b) Terry et al. ( Reference Terry, Osbourn and Cammell 21 ); (c) Agricultural and Food Research Council( 6 )) by using digestible organic matter in DM and crude protein as predictors (a and b) or digestible organic matter in DM as the sole predictor (c). (a) R c 0·603 (95 % CI 0·529, 0·668), (b) R c 0·709 (95 % CI 0·624, 0·778) and (c) R c 0·703 (95 % CI 0·626, 0·766), where R c is Lin's concordance correlation coefficient. DM represents DM content of fresh grass. Prediction equations are shown in Table 8 (equations AR, AU and AV).

Fig. 2 Bland–Altman plots showing the agreement between in vivo measured metabolisable energy (ME) and residual (predicted minus actual) ME, with ME being predicted from equations developed in the present study using two-thirds of the whole dataset, and by using nutrient digestibility and grass chemical composition parameters as predictors (predictors used: (a) organic matter digestibility; (b) gross energy digestibility; (c) organic matter digestibility and grass nitrogen content; and (d) gross energy digestibility and grass contents of nitrogen, gross energy, neutral-detergent fibre and acid-detergent fibre). (a) R c 0·848 (95 % CI 0·798, 0·887), (b) R c 0·864 (95 % CI 0·819, 0·899), (c) R c 0·813 (95 % CI 0·752, 0·760) and (d) R c 0·817 (95 % CI 0·757, 0·763), where R c is Lin's concordance correlation coefficient. DM represents DM content of fresh grass. Prediction equations are shown in Table 6 (equations K, M, O, Q).

Fig. 3 Bland–Altman plots showing the agreement between in vivo measured metabolisable energy (ME) and residual (predicted minus actual) ME, with ME being predicted from equations previously published ((a) Terry et al. ( Reference Terry, Osbourn and Cammell 21 ); (b) National Research Council( 5 )) or developed in the present study using two-thirds of the whole dataset (c and d), and by using digestible energy and grass chemical composition parameters as predictors (predictors used: (a) and (b) digestible energy; (c) digestible energy and grass contents of nitrogen and ether extract; (d) digestible energy and grass contents of water-soluble carbohydrates, ether extract and acid-detergent fibre). (a) R c 0·806 (95 % CI 0·745, 0·854), (b) R c 0·554 (95 % CI 0·477, 0·622), (c) R c 0·778 (95 % CI 0·710, 0·831) and (d) R c 0·810 (95 % CI 0·752, 0·855), R c represents Lin's concordance correlation coefficient. DM represents DM content of fresh grass. Prediction equations are shown in Table 6 (equations S and U) and Table 8 (equations AP and AQ).
Results
Prediction of digestible energy and metabolisable energy contents of grass using nutrients' digestibility as sole predictor
When the relations between energy and digestibility parameters were investigated using REML analysis, (1) contents of DE and ME and ratios of DE:GE and ME:GE in grass were positively related to DMd, OMd and DOMD and (2) there was a significant effect of DMd, OMd, DOMD and GEd according to the Wald statistic (Table 2; equations 1(a)–(c), 2(a)–(d), 3(a)–(c), and 4(a)–(d)). Relations of ME with nutrient digestibility parameters are graphically presented in online Supplementary Fig. S1.
For the prediction of DE or the DE:GE ratio, using OMd as sole explanatory variable showed the lowest MPE, using DOMD showed the highest MPE and using DMd showed intermediate values, although in case of the prediction of DE:GE the MPE for equations using DMd or DOMD was similar. Using GEd showed the lowest MPE, when DMd, OMd, DOMD or GEd was used as the sole explanatory variable for the prediction of ME and ME:GE ratio. Among the other three sole predictors, and similarly for the prediction of DE, OMd showed the lowest MPE, DOMD the highest MPE and DMd intermediate values.
Prediction of digestible energy and metabolisable energy contents of grass using nutrients' concentrations and digestibility
When the relations between energy parameters and nutrient concentrations and digestibilities were investigated, using REML analysis, the effects of (1) DMd, OMd, DOMD and GEd, and (2) grass N, EE, ADF, ash, WSC, GE and NDF contents on the prediction of energy parameters were significant, in various models, according to the Wald statistic (Table 3; equations 1(d)–(f), 2(e)–(h), 3(d)–(f), 4(e)–(h)). MPE was found generally reduced, when chemical composition variables of grass were added to prediction models of DE, ME, DE:GE and ME:GE that were developed by solely using DMd, OMd, DOMD or GEd as explanatory variable. The only exceptions were the higher MPEs for the predictions of ME when chemical composition parameters of grass were added to models using DMd, OMd or GEd as primary predictor.
MPE was shown as the lowest, when DE contents of grass were predicted from DMd, OMd or DOMD and chemical composition of grass, using a model with DOMD in which the Wald statistic identified as well N and EE contents of grass as significant explanatory variables. Using OMd as primary predictor, in conjunction with N, WSC, EE, and ash, showed lower MPE than using DMd as primary predictor with N, EE, ADF and ash.
MPE was shown as the lowest, when ME contents of grass were predicted from DMd, OMd, DOMD or GEd and grass chemical composition, using a model with GEd, in which the Wald statistic identified as well N, GE, NDF and ADF as significant predictors. However, this MPE was still higher when compared with the equation using GEd as the sole predictor. A slightly higher MPE was found when OMd was used as primary predictor, with only N being identified (by the Wald statistic) as significant additional variable. When DMd was used as primary predictor, N and ADF were also found as significant explanatory variables in the model, but using DOMD instead of DMd, was followed by a substitution of ADF with GE and EE in the model according to the Wald statistic, and the latter equation showed lower MPE.
MPE was shown as the lowest, when grass DE:GE ratio was predicted from DMd, OMd or DOMD and grass chemical composition, using a model with OMd as primary predictor; the Wald statistic identified NDF as significant additional predictor in the same model. When DMd or DOMD was used as primary predictor, MPE was only slightly lower for equations using DOMD; the latter model included N, NDF and ADF, while the former had only ADF, as additional explanatory variable(s) according to the Wald statistic.
Using a model with GEd showed the lowest MPE, when grass ME:GE contents were predicted from DMd, OMd, DOMD or GEd and grass chemical composition; N and GE were identified as significant explanatory variables in the same model by the Wald statistic. Among the other three equations, when OMd was used as primary predictor, the Wald statistic identified the same additional explanatory variables (N, GE) as significant; MPE was found lower when compared with equations using DMd or DOMD. According to the Wald statistic, using DOMD as primary predictor showed slightly higher MPE than using DMd, and the optimum model included the combination of N, GE and EE instead of N, GE and ADF.
Prediction of metabolisable energy contents of grass using digestible energy and nutrient concentrations
According to the Wald statistic, when the prediction of grass ME content from grass DE contents and chemical composition was developed using REML analysis, the effects of grass DE, N, EE, WSC and ADF were found significant in various models (Table 4, equations 2(i)–(m)). These equations showed similar range of MPE with prediction models for ME using nutrient digestibility and contents, although the lowest MPE observed in this set of equations (0·047, equation 2(l)) was not as low as the lowest observed when nutrient digestibility was used (0·043, equation 2(d)). When using the present set of data, it was observed that the addition of grass chemical composition parameters to prediction models for ME from DE improved MPE, but only when grass N contents were not among the explanatory variables, although the latter was also identified as a significant predictor according to the Wald statistic.
When using all available chemical composition parameters in the analyses in a model with DE as the primary predictor, the Wald statistic identified N and EE contents of grass as additional significant explanatory variables. However, this model showed slightly increased MPE, when compared to the model using DE as sole predictor and it also showed an increased MPE when further excluding EE from the multivariate model (thus leaving only DE and N as predictors). When using all available chemical composition parameters (excepting N) in the analyses, in a model with DE as the primary predictor, the Wald statistic identified WSC, EE and ADF contents of grass as additional significant explanatory variables. This model showed the lowest MPE among all models that included DE as primary predictor. However, it also showed an MPE value similar to that of the model using DE as the sole predictor, by further excluding EE from the multivariate model (thus leaving only DE, WSC and ADF as predictors).
Prediction of digestible energy and metabolisable energy contents of grass using total digestible nutrients and nutrient concentrations
When DE and ME concentrations of grass were predicted from tdCP, tdNDF and WSC and EE contents of grass, using REML analysis (Table 5, equations 1(g)–(i), 2(n), 2(o)), the effect of all explanatory variables was significant, except for the effect of EE in the prediction of ME. Predictions of DE and ME from this set of variables showed the highest MPE compared to those of all other models developed in the present study. For the prediction of DE and ME, stepwise addition of WSC and EE (only in the case of DE) showed decreased MPE, in a model which already includes tdCP and tdNDF as explanatory variables.
Internal validation of prediction equations
Thirty-eight new predictions, one for each equation developed in the present study by using the whole dataset, were developed from two-thirds of data by using the same random and fixed model and they are presented in Table 6 (equations A–AL). These new equations were validated using the remaining one-third of the dataset, and the outcome is presented in Table 7.
Mean predicted values were very close to the actual data for all the predicted energy concentration and ratio parameters. MPE (1) were lower for the predictions of DE:GE (0·009–0·029, average = 0·022), (2) showed intermediate values for the predictions of DE (0·021–0·080, average = 0·041), and (3) were higher for the predictions of ME (0·043–0·087, average = 0·056) and ME:GE (0·047–0·063, average = 0·056). Mean differences between the predicted and actual values represented less than 1·8, 3·1, 0·2, and 0·9 % for DE, ME, DE:GE and ME:GE, respectively, with some models showing a difference as low as 0·01, 0·13, 0·03 and 0·14 % of the actual values, respectively. Similarly, the proportions of standard errors in relation to the actual values for DE, ME, DE:GE and ME:GE were less than 5·4, 5·0, 1·9 and 3·9 %, respectively, with equations showing proportions as low as 1·4, 2·7, 0·7 and 3·2 % of the actual values, respectively.
Equations previously published for the prediction of ME concentrations( 5 , 6 , Reference Givens, Everington and Adamson 20 , Reference Terry, Osbourn and Cammell 21 ) from DE or nutrient digestibility and contents were also validated against the same one-third of dataset, and the results are presented in Table 8 (equations AM–AV). The MPE of these equations were found to be higher, when compared with those of the equations developed in the present study. The use of DE as the sole predictor, as shown by Terry et al. ( Reference Terry, Osbourn and Cammell 21 ), showed lower MPE when compared with that of DOMD as the sole predictor( 6 ), or in conjunction with crude protein content of grass( Reference Givens, Everington and Adamson 20 , Reference Terry, Osbourn and Cammell 21 ).
In order to assess agreement between predicted and actual values in the equations developed in the present or previously published( 5 , 6 , Reference Givens, Everington and Adamson 20 , Reference Terry, Osbourn and Cammell 21 ) studies, an agreement analysis using Lin's concordance correlation coefficient was performed, and the results are presented in Tables 7 and 8 respectively. Bland–Altman plots were generated to graphically represent the agreement between in vivo measured ME values and the difference between predicted and actual values for the equations: (1) that they showed the lowest MPE among the ones using nutrient digestibility and chemical composition of grass (Figs. 1 and 2) or using DE and chemical composition of grass (Fig. 3), and (2) that they are being used in the existing energy feeding systems in the UK( 6 ) (Fig. 1) and the US( 5 ) (Fig. 2). Lin's concordance correlation coefficients and 95 % CI are also shown in the same graphs, and they all recorded some agreement between predicted and actual values. These plots showed an under-prediction of ME when actual values were over 11·8–12·3 MJ/kg DM. The scatter of differences between predicted and actual values was smaller for the equations developed in the present analysis, by using nutrient contents and digestibility parameters as predictors, when compared with previous( Reference Givens, Everington and Adamson 20 , Reference Terry, Osbourn and Cammell 21 ) studies across the range of the in vivo measured ME values. When DE was used to predict ME, the scatter was only slightly improved in the equations developed in the present study when compared to the scatter of those published by Terry et al. ( Reference Terry, Osbourn and Cammell 21 ); also it indicated a lower average difference between predicted and actual values of the former. However, the recommended equation from the NRC( 5 ) over-predicted ME throughout the range of in vivo ME investigated.
Discussion
Compared with other works that developed prediction equations for grass ME using nutrient digestibility and laboratory measurements, the present study differed in some exclusive features. Firstly, the grass fed varied considerably in nutrient composition, as a result of growing in highly variant swards in terms of harvest date (primary growth, first regrowth, second regrowth), maturity (3 to 9 weeks), fertiliser input (8–70 kg N/ha) and grass variety (six different seed mixtures). Secondly, the number of samples (n 464, as a result of 3-d averaged 1392 records) was over 2·5 times higher than that in similar studies( Reference Givens, Everington and Adamson 20 ), with records being evenly collected over the different treatments, which were considered the main sources of the variation in grass quality. Thirdly, energy losses in methane production were systematically measured in calorimetric chambers in the first year and methane energy data were calculated from equations developed from part of the same dataset( Reference Yan and Mayne 13 ) used to develop the prediction equations in the present study, instead of using published equations from independent datasets. Last but not least, the present study was performed with cattle fed fresh grass to meet maintenance energy requirements, instead of sheep or cows fed at higher feeding levels; the latter are the most commonly used ones in ruminant metabolism research for the development of energy prediction equations and values of feeding tables( 5 , 6 , Reference Jarrige 8 , Reference Givens, Everington and Adamson 20 ). Compared with sheep, the digestibility in cattle is lower when offered diets that have a DMd above 660 g/kg, whereas the reverse is true for diets with a DMd lower than 660 g/kg and this may underestimate energy values for forages in the latter case( 22 ). Similar findings were reported by Yan et al. ( Reference Yan, Agnew and Gordon 23 ), based on direct comparisons of sheep and cattle fed silage-based diets, but little difference was observed when ensiled forage was offered at maintenance level.
The equations developed in the present study may be used in a wide range of pasture-based systems, where pure ryegrass swards are used, because they have been developed with animals fed fresh-cut grass of a highly diverse quality. These equations may not be efficient in predicting energy concentrations of swards of different botanical composition, because other forage plants were not assessed. Also, small differences regarding energy expenditure for foraging and the number and duration of meals may occur between zero-grazing and grazing in situ animals, which would probably slightly influence energy utilisation. These differences are difficult to be taken into consideration in metabolism experiments, because an accurate total 24 h collection of faeces and urine at pasture is not possible in practice. However, existing feeding systems recommend various correction factors for grazing cattle given that all equations used in animal rationing have been developed with housed livestock.
Prediction of digestible energy and metabolisable energy contents of grass using digestibility of nutrients as sole predictor
The positive relation between DMd, OMd and DOMD with DE and ME contents of grass was not surprising, because higher digestibility of these nutrient groups would decrease energy losses in faeces( Reference Waghorn and Clark 24 ). The most accurate prediction of ME in the present study was achieved when GEd was used as the sole predictor, and this may be due to the fact that energy losses in faeces, which define GEd, represented over 0·605 of total energy losses. An improved precision of the prediction of ME by using predictors which account for the proportional energy loss in faeces has previously been recommended by Givens et al. ( Reference Givens, Everington and Adamson 20 ) as well. However, GEd is not as extensively available as DMd, OMd or DOMD in practice, and equations based on the last three of these nutrient digestibility parameters may be necessary under commercial conditions. Among these three potential predictors, OMd was the most accurate for predicting DE and ME, based on the lower MPE observed in the equations using OMd as sole predictor, when compared with equations using DMd or DOMD. This may be explained by the inclusion of ash, which contains no energy, but is present in grass DM content, which influences DMd and DOMD values. However, MPE values were also relatively low when using either DMd or DOMD and, practically, these explanatory variables may be used for the prediction of ME content of fresh grass when details of GEd or OMd are not available. The importance of using digestibility parameters for the prediction of DE and ME is noteworthy because equations based only on laboratory measurements of grass derived relatively higher MPE in a previous study, either using equations developed from the same data as in the present study (MPE = 0·091) or those published by other authors (MPE = 0·094)( Reference Stergiadis, Allen and Chen 12 ); the same was observed to be the case in the present work when previously published equations( Reference Givens, Everington and Adamson 20 ) were validated.
It is however notable that using NDFd as sole predictor also showed an MPE similar to that of using DMd (0·050; online Supplementary Fig. S1). In a recent study in Ireland, NDF, ADF and CP contents in four commonly used varieties of Lolium perenne L. were measured at an average of 0·386, 0·247 and 0·233 of DM respectively during the vegetative growth stage( Reference Wims, McEvoy and Delaby 25 ). In the present study, the respective contents were found to be 0·503, 0·267 and 0·141 of DM, because a wider range of grass quality was investigated. Therefore, NDF accounts for a larger proportion of the DM contents than ADF or N, and the regression of NDFd digestibility against ME contents of grass tended to be similar to that of DMd.
Prediction of digestible energy and metabolisable energy contents of grass using nutrients' concentrations and digestibility
The addition of nutrient contents of grass as predictors in the equations based on nutrient digestibility parameters improved MPE, and seemed necessary in order to optimise the prediction of DE contents of grass in practice. N, GE, EE and ADF of grass were identified as significant predictors by the Wald statistic in various equations. Interestingly, when chemical composition of grass is not available, the use of equations based on OMd may be recommended. However, when nutrient contents of grass are known, an equation using DOMD as primary predictor would show lower MPE and require less additional explanatory variables (only N and EE) than an equation using OMd (which requires N, EE, WSC and ash). Therefore, decisions on the optimum models to predict DE in practice should be made based on what analyses are available to the researchers, nutritionists, analysts or producers.
In contrast with DE contents of grass, the MPE for the prediction of ME was slightly increased by adding nutrient content parameters as predictors to equations already using nutrient digestibility parameters. However, the average difference between predicted and actual values was reduced by 40 and 24 % when chemical composition of grass parameters were used as additional predictors in equations using GEd and OMd respectively, followed by a respective increase in the explained variation. This may imply that although the addition of chemical composition of grass parameters may be preferable in order to account for the type of nutrient within the digestible organic matter that provides the energy, the less complicated equations developed in the present study by using OMd or GEd as sole predictor may also offer a reliable option for the prediction of ME of grass for grazing cattle.
Higher N contents in feeds have previously been associated with an increase in DE and ME concentrations( 22 ). Positive and negative correlations of N and ADF contents of grass, respectively, with energy contents of grass were also seen in the present dataset as previously reported( Reference Stergiadis, Allen and Chen 12 ). Notably, N content of grass was identified by the Wald statistic as a significant explanatory variable in all prediction equations, thus emphasising its importance to the energy evaluation of fresh-cut grass in order to account for the possible losses associated with its metabolism and excretion in grazing cattle( Reference Bruinenberg, Zom and Valk 9 ). The negative intercepts in the prediction equations of grass N content were not associated to a negative relationship with ME, because the correlation coefficient of their univariate linear regression was positive( Reference Stergiadis, Allen and Chen 12 ). This finding was more likely a result of confounding effects when digestibility parameters were used as primary predictors in the same equation. Also, in the the present study, animals were fed iso-energetic diets regardless of N concentrations in grass. This may have led to N underfeeding or overfeeding in cases of poor quality or high-protein grass respectively. Feeding N below requirements might interact with the other explanatory variables in the same equations, because it is associated with reductions of DMd and OMd( Reference Fanchone, Noziere and Portelli 26 ). Likewise, cases of N overfeeding might increase energy losses for the hepatic synthesis of urea (which may account for up to 4 % of ME), excretion to kidneys, N turnover and ammonia removal process( Reference Bruinenberg, Zom and Valk 9 , Reference Waghorn and Clark 24 ). The latter may in particular occur in the absence of concentrate feeds, because inadequate readily available energy in rumen may decrease utilisation of high amounts of rapidly degradable N by rumen microbial population( Reference Hoekstra, Schulte and Struik 27 ). This would not affect DE evaluation (a positive intercept with grass N contents was found), but would reduce ME contents because the excess of N intake is excreted in urine rather than in faeces( Reference Stergiadis, Cheng and Allen 11 ).
According to the Wald statistic, the finding of the present study that EE contents of grass strongly affected energy contents is in line with Givens et al. ( Reference Givens, Everington and Adamson 20 ), with slopes being higher than those for other nutrients (except N), indicating a higher effect on a per weight basis; the latter possibly shows limited practical importance due to the low concentrations and variation of EE in grass compared with those of other nutrients.
The other main explanatory variables were GE, NDF and ADF, as identified by the Wald statistic during fixed model development for the predictions of DE and ME. Content of GE of grass was used by Givens et al. ( Reference Givens, Everington and Adamson 20 ) while fibre contents are used in the French and Australian energy systems in the forms of crude fibre and ADF or modified ADF respectively( Reference Jarrige 8 , 22 ). NDF and ADF contents of grass showed a negative relation with DE, ME and ME:GE concentrations, possibly as a result of increased energy excretion in faeces with higher ADF intake( Reference Givens, Everington and Adamson 20 ).
The ratio of ME:GE represents the efficiency of energy utilisation in the animal, and its practical implications may directly influence the UK( 6 ), the French( Reference Jarrige 8 ), the Australian( 22 ) and the Dutch( Reference Van Es 28 ) systems. In these energy evaluation systems, the ratio ME:GE is used to predict the partial efficiencies of milk and meat production and, in turn, the net energy value of feeds, from prediction equations using ME as the primary explanatory variable. Equations developed in the present study showed that the prediction error may be lowest when grass N and GE contents are used in conjunction with GEd (optimum model) or OMd (to be recommended when GEd is not available). When details of GEd or OMd are not known, DMd or DOMD may be used as primary predictors but this would reduce prediction accuracy and would also require the addition of ADF or EE, respectively, in the models.
Bland–Altman plots investigating the agreement between predicted values, from equations using nutrient contents and digestibility parameters as predictors, and the difference between predicted and in vivo measured ME, showed that predicted values were generally lower than actual values when ME contents of grass were relatively high. This pattern appeared in the predictions based on the equation of Givens et al. ( Reference Givens, Everington and Adamson 20 ) for ME values over 11·8 MJ/kg DM, while in the equation of Terry et al. ( Reference Terry, Osbourn and Cammell 21 ), this appeared at slightly higher values (over 12·8 MJ/kg DM). Interestingly, the equations currently used in the UK feeding system( 6 ) over-predicted ME when in vivo measured ME was lower than 11·3 MJ/kg DM, but under-predicted ME when in vivo measured ME was over 12·8 MJ/kg DM. This may be a result of the fact that the existing equation does not take into consideration the nutrient composition of DOMD. Poor quality pasture (with less N and EE and more WSC and fibre) may contain less ME per kg of DOMD than high quality pasture (with more N and EE and less fibre), because energy content of N and EE is higher, per weight, than that of WSC and fibre. Therefore, by multiplying solely DOMD with a single factor to predict ME may over-predict ME in poor quality grass, and under-predict ME in good quality grass; the latter highlights the necessity of including chemical composition of grass parameters in the predictions of ME of grass for grazing cattle. Equations developed in the present study also generally under-predicted ME when actual values were higher than 12·8 MJ/kg DM. However, the difference between predicted and actual values was smaller, thus offering support to the observation that the new equations may decrease the possible error in prediction within this range. This was also found applicable for the whole range of ME values, as implied by the lower MPE of the equations developed in the present study.
The findings of the present study are in accordance with previous reports and recommendations that the feeding value of grass can be more accurately predicted when parameters of nutrient content and digestibility are used in combination( 22 , Reference Waghorn and Clark 24 ). The same has also been recommended for the prediction of ME contents in feeds in experiments with silage-based diets( Reference Yan and Agnew 7 ).
Prediction of metabolisable energy contents of grass using digestible energy and nutrient concentrations
Similar to NRC( 5 ) recommendations, predictions of ME contents using DE as sole predictor were developed in the present study while regressions using DE in conjunction with chemical contents of grass were also investigated. Although using DE as the sole predictor showed relatively low MPE, the inclusion of WSC, EE and ADF contents of grass as predictors may be recommended for further improvement of prediction accuracy in practice. However, if EE is not available in practice, adding just WSC and ADF as predictors may not improve prediction accuracy, but, in the present study, it still reduced standard error and the difference between predicted and actual values by 13 and 32 % respectively, explaining the variation further when compared with the equation using DE as the sole predictor. The positive association between WSC, EE and ADF with ME can be explained by (1) the fact that these nutrients would be the main source of energy for N utilisation by rumen microbes and (2) the negative relation of grass N with WSC and fibre contents( Reference Hoekstra, Schulte and Struik 27 , Reference Tas, Elgersma, Dijkstra and Tamminga 29 ); these two features would decrease energy losses related to excess N metabolism and excretion( Reference Bruinenberg, Zom and Valk 9 ).
The significant effect of grass N on the prediction of ME may be explained by the fact that the difference between DE and ME is highly dependent on energy excretion in urine, and consequently on N intakes and outputs( Reference Waghorn and Clark 24 ). Although this was observed in the present study and the Wald statistic too identified grass N content as significant predictor of ME when DE was used as primary predictor, the addition of N and EE increased MPE when compared with the equation using DE as the sole predictor; but it also explained more variation and decreased the average difference between predicted and actual ME by 44 %. Therefore, using grass N content as an additional predictor may be recommended when WSC and ADF are not available in practice.
Bland–Altman plots were drawn to assess the agreement between predicted values, from equations using DE and nutrient contents as predictors, and the difference between predicted and in vivo measured ME; they showed, similar to equations using nutrient contents and digestibility, that ME was under-predicted for ME values over 12·8 MJ/kg DM in all equations, either in previously published studies( Reference Terry, Osbourn and Cammell 21 ) or developed in the present study. However, the smaller MPE of the equation developed in the present study implied that using (1) the updated equations when DE is the sole predictor and (2) WSC, EE and ADF as additional predictors when available in practice, may improve ME prediction accuracy. Interestingly, the current equation presented by NRC( 5 ) showed an over-prediction of grass ME contents throughout the whole ME range investigated in the present study, implying a possible under-supply of ME by using NRC's equation for grazing cattle.
Prediction of digestible energy and metabolisable energy contents of grass using total digestible nutrients and nutrient concentrations
For the prediction of DE and ME contents of grass, NRC( 5 ) recommends equations using chemical composition of grass as explanatory variables (NDF, crude protein, EE, ash, and lignin), in conjunction with truly digestible nutrients (non structural fibre carbohydrates, crude protein and NDF). A couple of limitations may occur in their use for pasture-based systems, because these equations were developed with animals fed on conserved forage and concentrates at three times maintenance level. Similar explanatory variables were used to predict ME contents in forages in the Dutch system( Reference Van Es 28 ), with non-lactating cows fed at 0·5–1·7 times maintenance level. Based on the principles of these two systems, and in an attempt to simplify the existing relations to meet the UK routine laboratory analyses, equations in the present study were developed by using tdCP and tdNDF as primary predictors, by also adding contents of EE and WSC of grass. Indeed, these parameters showed a significant effect on the prediction of DE and ME, and MPE was decreased when EE and WSC were added in the equations, although the effect of EE contents on the prediction of ME was not significant according to the Wald statistic. However, NDFd was used to calculate tdNDF; MPE was relatively low (0·050; online Supplementary Fig. S1), when NDFd was used as the sole predictor in cases where details of GEd, OMd, DMd or DOMD were not used. Therefore, using NDFd seems to be an easier, and possibly more accurate approach, than using digestible nutrients and chemical contents of grass to predict ME. This set of equations, among all the equations developed in the present study or previously published studies, produced the highest MPE during the internal validation, and may preferably be avoided for grazing cattle.
Conclusions
The equations developed in the present study may be applicable to a range of grazing conditions, where pure ryegrass pasture is used, because they have been developed with animals fed fresh-cut grass of a highly diverse quality, and may be used accordingly for different sets of available analyses. The present study highlighted that the use of a combination of chemical composition of grass and nutrient digestibility parameters may improve the accuracy of the prediction of energy concentrations in fresh grass, mainly because the former variables account for the type of nutrient, within the organic matter, that provides the energy. However, the developed equations using GEd or OMd may also be a reliable alternative to predict ME, as indicated by their low prediction errors, and may be recommended instead of equations using DOMD. Similarly, when ME was predicted by using DE as the main predictor, combinations of energy-supplying nutrient contents (EE, WSC and ADF, or N and EE) may be used to improve prediction on a commercial scale. Using DE as the sole predictor for ME contents of grass was a relatively reliable technique and the updated equations formulated in the present study may reduce prediction error, in the case of grazing cattle, when compared with existing equations using solely DE. As implied by the lower prediction errors of the equations developed in the present study, when compared with existing equations used in the US and the UK feeding systems, prediction of DE and ME may be improved by their implementation in animal rationing and routine grass chemical analysis. An under-prediction of ME for high quality grass (in vivo measured ME ≥ 12·8 MJ/kg DM) was common for all equations across the studies although this error was slightly reduced in the equations developed in the present study, compared with existing predictions, as illustrated in Bland–Altman plots.
Supplementary material
To view supplementary material for this article, please visit http://dx.doi.org/10.1017/S0007114515000896
Acknowledgements
The authors would like to thank colleagues at the Agri-Food and Biosciences Institute (Hillsborough, the UK) for assistance in the collection of experimental data used in the present study.
The authors gratefully acknowledge financial support from the Department of Agriculture, Food and the Marine of the Republic of Ireland under a Research Stimulus Fund Program (11/S/105) and the Department of Agriculture and Rural Development of Northern Ireland. The authors declare no conflicts of interest.
S. S. developed the datasets, performed the statistical analyses, interpreted the data and wrote the manuscript. M. A. advised on and performed the statistical analyses, wrote parts of the ‘Materials and methods’ section and contributed to the critical review of the ‘Materials and methods’ and ‘Results’ sections. X. C. and D. W. collected the experimental data. T. Y. managed the research project, designed and supervised the study and the writing up process, and contributed to the critical review of the manuscript. All authors contributed and approved the final version of the manuscript.













