

1. Introduction
Schizophrenia is the most disruptive of neuropsychiatric disorders, affecting approximately 1% of the world's population [Reference Saha, Chant, Welham and McGrath1]. The disease is characterized by profound and lasting impairments particularly in language, memory and cognition, as well as in the structure and function of brain regions sub-serving these domains [Reference Keshavan, Tandon, Boutros and Nasrallah2]. Influential studies have rightfully advocated for the study of higher order cognitive impairments in schizophrenia, considering these impairments as a central pathway for understanding core elements of the illness’ underlying pathophysiology [Reference Goldman-Rakic3]. Yet, a complimentary literature has come to emphasize that dysfunction in basic motor function and control represents a highly relevant physiological pathway in the illness [Reference Izawa, Asai and Imamizu4]. Dysfunction in motor behaviors has frequently been associated with prediction errors, for example, deficiencies that patients show in predicting the consequences of motor actions [Reference Synofzik, Thier, Leube, Schlotterbeck and Lindner5]. However, motor dysfunction in schizophrenia is even more direct, involving impairments in basic motor processing and control. That this is the case is supported by evidence that brain regions such as the dorsal anterior cingulate cortex, the basal ganglia and the cerebellum, each of which are associated with higher order functions such as memory and executive control and implicated in schizophrenia [6–Reference Woodcock, Wadehra and Diwadkar9] are also associated with basic motor function [Reference Witt, Laird and Meyerand10,Reference Middleton and Strick11].
Recent reviews have presciently acknowledged the singular and compelling role that motor abnormalities play in the illness. Hirjak and colleagues noted that these abnormalities cluster into at least three distinct classes: neurological soft signs, abnormal involuntary movements and catatonia [Reference Hirjak, Thomann, Kubera, Wolf, Sambataro and Wolf12], and argued for motivated imaging approaches to identify the correlates of these sub-types. Such approaches can parse apart the “motor endophenotype” in schizophrenia. In a separate review [Reference Walther13], Walther also focused on these classes of motor abnormalities, but emphasized focus on the cerebellar motor circuit, as a key neural pathway, impairments within which are relevant for many of the dimensional deficits in motor function in the illness. Here we provide a synthesis reaffirming the value of studying motor function and dysfunction in schizophrenia. Our approach summarizes some of the evidence indicating that motor dysfunction is at the heart of a core pathophysiological pathway in the illness [Reference Kasparek, Rehulova, Kerkovsky, Sprlakova, Mechl and Mikl14], yet our synthesis adds complementary elements not previously marshaled in support of developing an integrative framework. Thus, we build our rationale as follows: We first briefly summarize the current and historical context for motivating the relevance of motor dysfunction in schizophrenia. We follow with a pivot to basic movement physiology, reviewing the architecture of the primate motor system, one that is well-delineated by structural and electrophysiological studies, and confirmed with in vivo fMRI studies. Then we theoretically navigate the intersection between this organization of motor function, and schizophrenia-related pathophysiology that is observed in brain regions implicated in motor processing. We then transition to a discussion of the molecular correlates of schizophrenia, particularly in the context of the dopaminergic hypothesis that offers a manner of convergence with the motor system. Each of these principal sub-sections are summarized in accompanying schematic figures.
We reiterate that we provide more synthesis than review. Each of the sub-sections herein would demand more space for a comprehensive treatment [15–Reference Friston, Brown, Siemerkus and Stephan17], and as noted, separate reviews have effectively addressed some of these. Yet, in linking basic movement physiology, schizophrenia and neurotransmitters, we supplement previous advocacy for increased focus on the study of motor dysfunction and its pathophysiological bases in schizophrenia. The study of motor dysfunction in schizophrenia is in fact, not distinct from the study of higher order dysfunctions in the illness. Schizophrenia is a “thought disorder”, yet notably thinking has been conceptualized as an “active motor process” [Reference Graybiel18]; structures such as the basal ganglia that are considered cognitive pattern generators are also motor pattern generators [Reference Graybiel19], generating sequences of commands designed to sub serve future action. In this way, motor function is a foundation for higher-level cognitive function. Indeed, impairments in extended domains of the motor system, including oculomotor deficits, have been directly linked to the core pathophysiology of psychosis itself [Reference Thakkar, Diwadkar and Rolfs20]. The motor system itself provides a tractable framework for studying brain networks, as the simplest of tasks evoke sophisticated interactions between regions such as the anterior cingulate, supplementary motor area and primary motor cortex [21–Reference Diwadkar, Asemi, Burgess, Chowdury and Bressler23]. Moreover, these tasks are not characterized by many of the patient-control performance differences that confound the interpretation of in vivo imaging studies of higher order cognitive function [Reference Carter, Heckers, Nichols, Pine and Strother24]. A focus on motor function is also of significant clinical relevance. Schizophrenia patients with general motor abnormities have less favorable outcomes, and motor abnormalities in turn predict deficits in memory, executive functioning, and attention [Reference Bombin, Arango and Buchanan25], supporting the notion of thinking as an active motor process. Also, patients with motor dysfunction develop more severe side effects of antipsychotic medications [Reference Peralta and Cuesta26], and longitudinal studies indicate that Neurological Soft Signs (NSS) predict elevations in negative symptoms of schizophrenia [Reference Hirjak, Wolf, Wilder-Smith, Kubera and Thomann27].
2. Motor dysfunction in schizophrenia: Background
Aberrant motor functioning in patients with schizophrenia has been noted since the earliest systematic clinical characterizations of the illness [Reference Walther and Strik28]. General movement disorders in schizophrenia were described as early as 1919, in the pre-neuroleptic era with Kraepelin describing schizophrenia patients with dementia praecox and paraphrenia as having “…spasmodic phenomena in the musculature of the face and of speech”, “involuntary choreic movements”, and “atheoide ataxia” [Reference Kraepelin and Shepeherd29]. More generally, studies have noted abnormal involuntary movements (dyskinesia), slowness in planning and execution of fine motor tasks (psychomotor slowing), problems with coordination and sensory-motor tasks Neurological Soft Signs (NSS), rigid posturing, grimacing, and mannerisms (catatonia), and Parkinsonism [Reference Morrens, Docx and Walther30]. With the advent of typical antipsychotics, motor dysfunction in schizophrenia has increasingly been associated with their extra-pyramidal side effects [Reference Whitty, Owoeye and Waddington31]. However, in addition to the early characterization of motor dysfunction, evidence suggests that antipsychotic medications may in fact serve to exacerbate the emergence of spontaneous motor disorders, rather than being the single underlying cause [Reference Peralta and Cuesta26,Reference Pappa and Dazzan32].
Estimates of the frequency of motor abnormalities in schizophrenia patients range between 50%–65% (as compared to ∼ 5% in healthy controls) [Reference Bombin, Arango and Buchanan25]. Retrospective studies have suggested that motor abnormalities significantly predate the onset of psychosis, with delayed motor development in childhood seen as a risk factor for future schizophrenia spectrum disorders [Reference Murray, Jones, Moilanen, Veijola, Miettunen and Cannon33]. These deficits also tend to aggregate in families of patients, suggesting that abnormal motor development is a vulnerability marker, and a fundamental correlate of psychosis [Reference Woods, Kinney and Yurgelun-Todd34]. The underlying neurobiology of motor behavior, particularly associated with the hand effectors is reasonably well established both in primate [Reference Armstrong35] and neuroimaging studies [Reference Witt, Laird and Meyerand10]. These detailed descriptions therefore provide a basis for the prospective assessment of the pathophysiology of brain networks in schizophrenia. In advocating this, we simply echo the “physiological approach” that has elucidated dysfunctional working memory in the illness [Reference Goldman-Rakic36,Reference Lewis37].
3. The architecture of the primate motor system: Motor regions and beyond
Studies of the primate motor system have provided some of the most carefully defined associations between a simple behavioral domain and mechanisms in the neural substrate. Motor behavior is subserved by highly coordinated brain sub-networks, principally involving the Primary Motor Cortex (M1), the Supplementary Motor Cortex (SMA), the Pre-Supplementary Motor Cortex (pre-SMA), the Dorsal Anterior Cingulate (dACC), the inferior parietal cortex, the basal ganglia, and the anterior cerebellum [Reference Dum, Levinthal and Strick38,Reference Dum and Strick39]. Notably, dysfunction in several of these brain regions has also been implicated in schizophrenia and higher-level cognitive dysfunction associated with the disease.
The primary motor cortex (M1; Brodmann Area 4) is responsible for eliciting localized movements in various parts of the body, with its specific areas topographically organized. The extent of cortical surface area responsible for the control of a body part varies in approximate proportion to the precision of movement it demonstrates [Reference Chouinard and Paus40]. BA 6, comprised of the premotor cortex and Supplementary Motor Area (SMA), is another important region implicated in motor control. The SMA is associated with the planning, initiation, and anticipation of specific movements [Reference Makoshi, Kroliczak and van Donkelaar41], and further divided into rostral pre-SMA and the caudal SMA that together form a dedicated subcortical-cortical temporal processing network [Reference Schwartze, Rothermich and Kotz42]. For example, the pre-SMA engages in the initiation and updating of non-automatized internally generated movements, as opposed to the SMA-proper, which plays a role in externally generated movements [Reference Nachev, Kennard and Husain43,Reference Picard and Strick44].
The dACC, also referred to as the mid-cingulate cortex, [Reference Hoffstaedter, Grefkes, Caspers, Roski, Palomero-Gallagher and Laird45] lies outside the core motor circuit, yet is generally regarded as a critical interface between cognition and motor control [Reference Asemi, Ramaseshan, Burgess, Diwadkar and Bressler22,Reference Paus46,Reference Kronhaus and Willshaw47]. Located on the medial surface of the cerebral hemispheres, the dACC has extensive structural and functional connections to the lateral prefrontal cortex, limbic structures, SMA, and the striatum [Reference Hutchison, Womelsdorf, Gati, Leung, Menon and Everling48,Reference Vogt, Pandya and Rosene49]. The structure's regulatory role in a hierarchical model of motor control, is sub-served through dense projections that transmit selective modulatory signals to the SMA and other structures [Reference Bates and Goldman-Rakic50,Reference Luppino, Matelli, Camarda and Rizzolatti51]. Recent fMRI studies have confirmed such a modulatory influence that allows maintenance of timing expectancy [Reference Asemi, Ramaseshan, Burgess, Diwadkar and Bressler22], and in general, the dACC is highly active during tasks requiring inhibitory or excitatory motor control [52–Reference Nakamura, Roesch and Olson54]. fMRI studies in humans and single-unit electrophysiological studies of primates have demonstrated functionally heterogeneous regions of the dACC that respond distinctly and disproportionately to reward-based decisions of motor selection [Reference Shima and Tanji55,Reference Bush, Vogt, Holmes, Dale, Greve and Jenike56]. This willed control of motor behavior is consistent with the idea of dACC involvement in suppressing externally triggered motor routines, as seen in lesion studies of the structure in primates.
The selection of motor functions and higher-level cognitive decisions of voluntary movement are also influenced by the basal ganglia and its constituents including in particular the caudate nuclei and the putamen [Reference Avanzino, Pelosin, Vicario, Lagravinese, Abbruzzese and Martino57,Reference Morris, Kundu, Dowell, Mechelmans, Favre and Irvine58]. Furthermore, a series of parallel loops connect many cortical inputs to the basal ganglia and serve to control voluntary movements [Reference Nambu59]. These discrete loops also make reciprocal connections from the basal ganglia motor system that also includes the Globus Pallidus (GP), the substantia nigra and Sub-Thalamic Nuclei (STN) to cerebral cortical areas, including the dorsolateral prefrontal cortex, and the anterior cingulate [Reference Leisman, Braun-Benjamin and Melillo60]. The basal ganglia's components, together with the cerebral cortex and the ventrolateral nucleus of the thalamus, are implicated in movement control through a model [Reference Albin, Young and Penney61,Reference Alexander, Crutcher and DeLong62] in which the basal ganglia is controlled directly and indirectly by two discrete pathways. Both pathways are inhibitory, suppressing thalamic activity, and begin with cortical excitation of neurons located in the striatum. These motor pathways of the basal ganglia are of particular interest in schizophrenia because of their rich dopaminergic circuitry and the coinciding role of dopamine in reward, psychosis and movement [Reference Perez-Costas, Melendez-Ferro and Roberts63]. The implications for schizophrenia of this motor cortical-striatal motor pathway are self-evident both from the perspective of systems neuroscience, as well as dopaminergic models of the illness [Reference Howes and Murray64].
Dopamine produced and synthesized in dopaminergic neurons in the substantia nigra is released in the striatum, stimulating the direct pathway through D1 receptors. Dopamine also inhibits the indirect pathway by binding to D2 receptors of GABA neurons [Reference Brittain and Brown65]. This indirect pathway begins with cortical excitation of striatal neurons that excite populations of GABA neurons, distinct from those excited in the direct pathway. These project to neurons in the STN, which in turn project to the GP internal segment to inhibit motor related functions of the thalamus. The balance of the two conjugate pathways is thought to be key for normal motor function [Reference Harrington, Hening and Poizner66]. The role of the basal ganglia in movement balances muscle activation, helps in sequencing activation for fast movements, and selectively inhibits competing movements that could interrupt voluntary action [Reference Patel, Jankovic and Hallett67].
The cerebellum consists of two lateral hemispheres and a medial vermis [Reference Pollok, Butz, Gross, Sudmeyer, Timmermann and Schnitzler68], and these cerebellar regions have highly variegated and diverse roles in voluntary motor control, balance, coordination, and higher cognitive non-motor functions. The cerebellum can be further divided into ten mirrored lobules, which are somatotopically organized in a series of sensory-motor “homunculi”, arranged dorso-ventrally [Reference Batson, Petridou, Klomp, Frens and Neggers69]. Cerebellar lobules IV, V, and VIII are robustly activated during motor tasks [Reference Desmond, Gabrieli, Wagner, Ginier and Glover70], and cognitively demanding tasks elicit activation of lobules VI and VII [71–Reference Diwadkar, Meintjes, Goradia, Dodge, Warton and Molteno73]. Unsurprisingly, the structural architecture of the cerebellar lobules consists of motor and non-motor closed loop circuits to the thalamus other regions of the cerebral cortex [Reference Bernard, Seidler, Hassevoort, Benson, Welsh and Wiggins74]. Connections to the cortico-ponto-cerebellar system make the cerebellum a key control region for voluntary motor control, enhancing the structures’ relevance for schizophrenia [Reference Picazio and Koch75]. The main output center of the cerebellum is the dentate nucleus, which links cortical structures such as premotor, prefrontal, posterior parietal, and primary motor cortex mainly via the thalamus. The cortex sends projections back to the cerebellum for processing via the pons. These cortico-cerebellar loops link function of the cerebellum to the function of the cerebral cortex and vice versa [Reference Middleton and Strick76]. The cerebellum is thought to act as a timer, updating and predicting body dynamics for fast movements based on sensory feedback [Reference Salman and Tsai77], and this precision is assumed to have high temporal fidelity [Reference Ivry and Spencer78]. This model provides the rationale for cerebellar involvement in coordination, precision, and timing of movement, as well as sequential processing of spatial and temporal information [Reference Molinari, Chiricozzi, Clausi, Tedesco, De Lisa and Leggio79], and has been considered of fundamental importance in the context of schizophrenia [Reference Andreasen80,Reference Andreasen, Paradiso and O’Leary81].
Fig. 1 provides an overview of the general architecture of the motor system. It is notable that functional impairments and structural abnormalities in each of these regions have been associated with motor dysfunction in schizophrenia. In the subsequent section, we explore how and why the patterns of motor dysfunction in schizophrenia may emerge from the macroscopic network of brain regions depicted in Fig. 1.
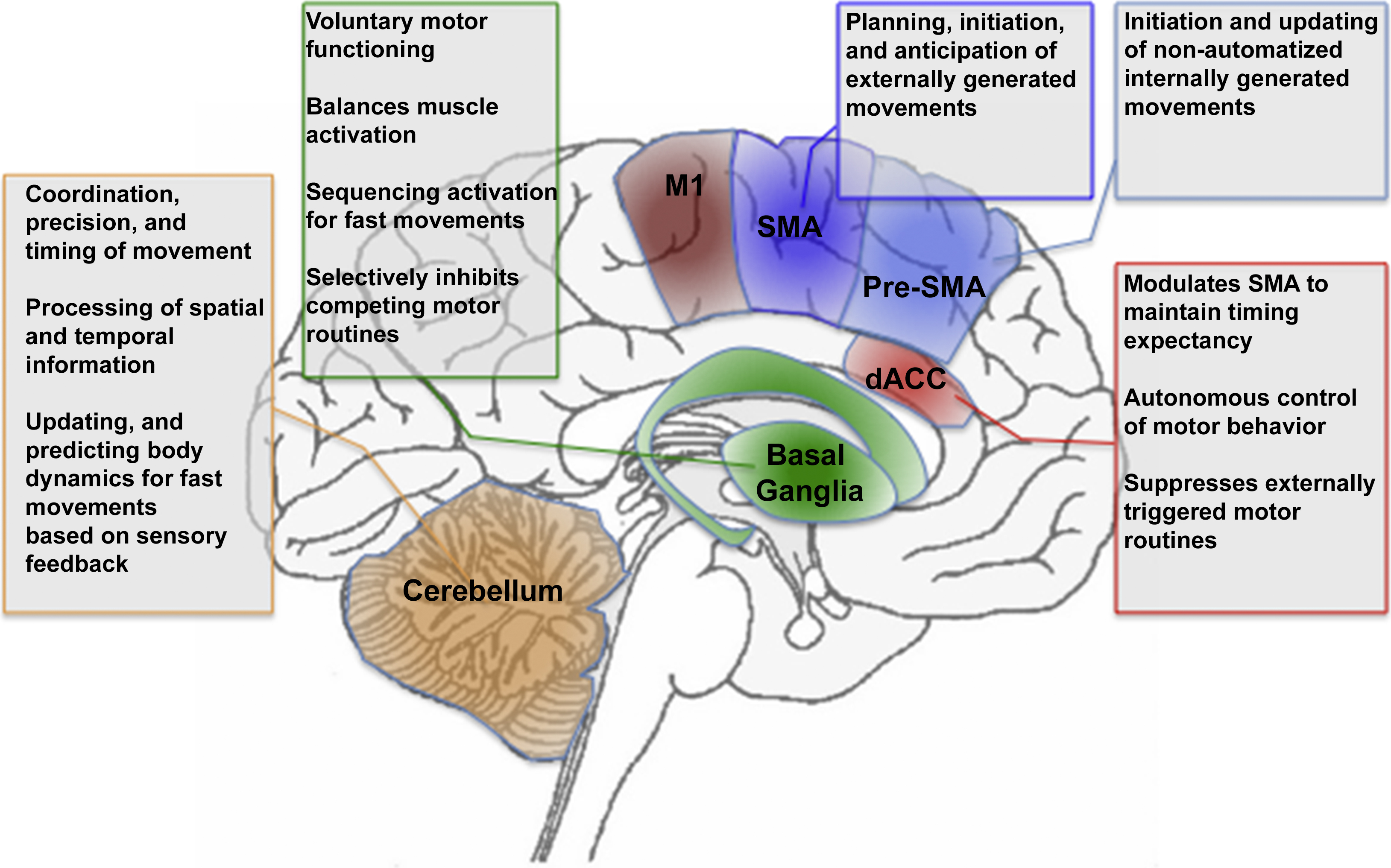
Fig. 1 The figure provides a schematic depiction of motor, cingulate and cerebellar regions and their role in aspects of motor behavior. Highlighted are M1: Primary Motor Cortex, SMA: Supplementary Motor Area, Pre-SMA: Pre-Supplementary Motor Area, dACC: Dorsal Anterior Cingulate Cortex, Basal Ganglia, and Cerebellum. The corresponding text boxes summarize known aspects of the relative specialization of function of the regions.
4. Dysfunction in regions of the core and extended motor network in schizophrenia
Several different abnormalities in structure, function, and connectivity of brain regions of the motor system may be involved in the emergence of motor dysfunction in schizophrenia. Here we provide a succinct summary of several representative findings of regional and network dysfunction of motor-function related brain regions.
4.1. M1 and the Supplementary Motor Cortices
Aberrant activity in M1 and the pre-motor cortex has been observed in schizophrenia patients during finger sequence learning and relatively simple sensorimotor detection tasks, though the precise nature of the aberration appears task specific. Kodama and colleagues trained patients and controls on a simple finger sequence learning task, demonstrating that compared to healthy controls, patients hyper-activated pre-motor cortex following sequence learning [Reference Kodama, Fukuzako, Fukuzako, Kiura, Nozoe and Hashiguchi82]. This pattern of hyper-activation is suggestive of impaired efficiency of motor control and learning mechanisms, and was conceptually replicated in a separate study by Minzenberg and colleagues [Reference Minzenberg, Yoon, Soosman and Carter83]. When patients were required to perform a basic sensorimotor detection task, they hyper-activated ipsi-lateral (to the response hand) M1 compared to healthy controls. However, aberrant hypo-activation of the motor cortices has also been documented using sequential and non-sequential finger tapping tasks that do not involve learning [84–Reference Muller, Roder, Schuierer and Klein86]. As is the case with other classes of tasks, such as working memory, the direction of the aberrance in schizophrenia is most likely yoked to the specific task demands and performance metrics of patients [Reference Manoach87,Reference Diwadkar, Pruitt, Zhang, Radwan, Keshavan and Murphy88]. Notably, impairments are observed not only in basic motor control when generating finger movements, but also during the application of motor force using the hand effectors. In a recent study, Martinelli and colleagues required participants to apply manual force to a lever that was commensurate with a visual cue signaling the degree of force that was to be applied [Reference Martinelli, Rigoli and Shergill89]. In healthy controls, the degree of applied force generated to this external cue was strongly correlated with fMRI-estimated responses in the primary somatosensory cortex, indicative of normal force signaling in motor regions. This coupling was absent in patients, suggesting that schizophrenia may alter mechanisms that encode and predict sensorimotor responses (the notion of impaired predictive coding is re-visited below). What appears relevant is that basic motor behaviors evoke impaired responses in the motor network. These effects are also generally associated with aberrant metrics of morphology.
Schizophrenia patients who display aberrant motor behavior also show aberrant morphometric characterization of several of the motor structures depicted in Fig. 1. In a study of first episode schizophrenia, reduced size of the pre-SMA was associated with impaired motor sequencing [Reference Exner, Weniger, Schmidt-Samoa and Irle90], an effect that has been replicated using the conjoint assessment of fMRI and voxel-based morphometric studies [Reference Singh, Goyal, Modi, Kumar, Singh and Bhatia91]. The changes in size have also been related to symptom dimensions and clinical scales. For example, volumetric reductions and fMRI measures of activation are negatively correlated with NSS scores in the SMA and M1 of recently diagnosed patients [92–Reference Venkatasubramanian, Jayakumar, Gangadhar and Keshavan94]. Alterations in the size of the SMA are also thought to represent a compensatory cortico-cortical pathway that may offset insufficient basal ganglia output [Reference Stegmayer, Horn, Federspiel, Razavi, Bracht and Laimbock95], and in catatonic patients with schizophrenia, increased severity of catatonia (based on the Bush Francis Catatonia Rating scale) predicts higher blood perfusion in the SMA [Reference Walther, Schappi, Federspiel, Bohlhalter, Wiest and Strik96]. This latter effect suggests that schizophrenia is associated with dysregulation of the SMA in the resting state.
Diffusion Tensor Imaging (DTI) studies of white matter connections of motor cortices have demonstrated alterations in the cortico-basal and cortico-cortical pathways. Specifically, in patients with motor abnormalities, there is an associated increase in motor activity between the pre-SMA to SMA-proper connection with less motor activity in the right pre-SMA to GP connection [Reference Bracht, Schnell, Federspiel, Razavi, Horn and Strik93]. These findings are similar to compensatory motor activity of decreased basal gangia output proposed in Parkinson's disease [Reference Grafton97]. This idea parallels DTI findings of abnormal white matter integrity found under the right SMA in schizophrenia patients with decreased resting state wrist movement [Reference Walther, Federspiel, Horn, Razavi, Wiest and Dierks98].
As previously noted, cognitive and motor control is partly mediated by the Dorsal Anterior Cingulate Cortex (dACC). Cingulate cortical neurons project to multiple areas of the motor and premotor cortex, suggesting that numerous parallel pathways exist whereby cingulate neurons can modulate motor output systems of the brain. Structural MRI studies suggest that reductions in gray matter volume in the dorsal and rostral ACC, frequently predate the onset of psychosis in high-risk individuals [Reference Fornito, Yücel, Dean, Wood and Pantelis99,Reference Calabrese, Wang, Harms, Ratnanather, Barch and Cloninger100]. In schizophrenia patients, decreased volume of the cingulate gyrus is significantly and selectively correlated with executive dysfunction [Reference Szeszko, Bilder, Lencz, Ashtari, Goldman and Reiter101]. Also, reduced synaptic density, deficits in inhibitory interneurons, and aberrant synaptic pathology have been found in the cytoarchitecture of cingulate cortex in patients [Reference Harrison102]. These aberrations complement fMRI studies of finger tapping, which demonstrate failed motor task-related activation of the ACC and disrupted connectivity to medial frontal cortex [Reference Honey, Pomarol-Clotet, Corlett, Honey, McKenna and Bullmore103], evidence of functional impairments in basic volition. Recent evidence sheds light on the structural correlations of avolition in the motor domain. Using estimates of non-Gaussian diffusion (based on Diffusion Kurtosis Imaging, (DKI)), Docx and colleagues examined the relationship between volitional motor activity (assessed with actigraphy) and white matter structure. In general, patients were less active than healthy controls, and showed greater diffusivity and lower fractional anisotropy on longitudinal white matter tracts [Reference Docx, Emsell, Van Hecke, De Bondt, Parizel and Sabbe104]. Moreover, in patients, mean kurtosis (a measure that better distinguishes between hindered versus unhindered diffusivity), was strongly associated with actigraph estimates of activity in patients. These estimates of motor activity related to effects in the brain's substrate also substantiate evidence of NSS deficits.
In schizophrenia, structural morphology of the basal ganglia, especially that of the caudate nucleus, putamen, and GP is atypical and associated with neurological soft signs [Reference Hirjak, Wolf, Wilder-Smith, Kubera and Thomann27], but the role that neuroleptics play appears to be quite heterogeneous. Grossly, the striatum is smaller than normal in antipsychotic-naive patients though upon antipsychotic treatment striatal and pallidial volumes enlarge [Reference Brandt and Bonelli105], which is a possible explanation for increased basal ganglia volumes in medicated schizophrenia patients [Reference Mamah, Wang, Barch, de Erausquin, Gado and Csernansky106]. In addition to size, alterations in shape of both the caudate and putamen have been found in schizophrenia patients as compared to healthy siblings [Reference Mamah, Wang, Barch, de Erausquin, Gado and Csernansky106]. Furthermore, morphological abnormalities of the putamen are observed in regions where prominent connections to non-motor cortical areas exist. Striatal abnormalities are exacerbated in the context of deficit syndrome schizophrenia, a sub-class of the diagnosis in which patients are characterized by pervasive negative symptoms, social withdrawal and increased anhedonia [Reference Amador, Kirkpatrick, Buchanan, Carpenter, Marcinko and Yale107]. Neurolepticnaïve deficit syndrome patients (relative to non-deficit syndrome patients) are characterized by increases in spontaneous movement disorders, consistent with the role of the basal ganglia in mediating both schizophrenia symptomatology and movement function [Reference Peralta, Moreno-Izco, Sanchez-Torres, Garcia de Jalon, Campos and Cuesta108]. Thus, impaired activity in the basal ganglia with hyperactivity in the primary, premotor, and somatosensory cortices, is thought to lead to cortical motor overflow and subsequent motor disturbances [Reference Minzenberg, Yoon, Soosman and Carter83].
Many of the alterations of motor symptoms described in schizophrenia, such as NSS, abnormal eye movements, and disequilibrium, indicate dysfunctions in the cerebellum. Abnormal cerebellar structure and connectivity with the motor cortex has been implicated in motor and timing dysfunction in the illness [Reference Andreasen and Pierson109,Reference Yang, He, Zhong and Multimodal110]. Transcranial magnetic stimulation directed at the cerebellum has shown increased excitability of the motor cortex in motor dysfunctions [Reference Fitzgerald, Brown, Daskalakis and Kulkarni111]. These findings suggest a decrease in cortico-cerebellar inhibition of motor cortex, which is important in the coordination of motor sequencing. Investigations of schizophrenia patients with motor sequencing abnormalities revealed lower functional connectivity between the cerebellum and motor cortex during the resting state [Reference Kasparek, Prikryl, Rehulova, Marecek, Mikl and Prikrylova112], a suggestive marker for structural dysconnectivity. Interestingly, schizophrenia patients with marked motor sequencing impairment had more psychotic episodes than those without movement sequencing disturbances [Reference Kasparek, Rehulova, Kerkovsky, Sprlakova, Mechl and Mikl14], a finding that suggests that consecutive psychotic outbreaks play a role in progressive worsening of cortico-cerebellar connectivity. NSS, like impaired movement sequencing, seem to be a result of impaired coordination between these cortico-cerebellar areas involved in the motor system [Reference Kasparek, Rehulova, Kerkovsky, Sprlakova, Mechl and Mikl14]. From a neuropathological perspective, cerebellar purkinje cells, responsible for modulating input from the cerebellum to the cortex, have decreased size and linear density in schizophrenia [Reference Tran, Smutzer, Doty and Arnold113]. Purkinje cells and their extensive connectivity of climbing fibers in the inferior olive mediate cerebellar coordination of cognitive activity [Reference Popa, Hewitt and Ebner114] and the many cortical and sensory inputs received permit the purkinje cells to detect error and selectively shuttle information back to the cerebral cortex through inhibitory output of deep nuclei. Thus, cerebellar organization influences the fidelity of perceptions, detects errors, and rapidly modulates coordination. Schizophrenia may impair the proper modulation and coordination of multiple signals causing a misinterpretation of erroneous sensory associations usually suppressed by the cerebellum, resulting in the frequently noted evidence of abnormal predictive coding in the illness [Reference Friston, Brown, Siemerkus and Stephan17].
The preceding discussion is summarized in Fig. 2, wherein the text boxes that in Fig. 1 represented the putative functions of each region, are replaced with a summary of regional deficits observed in schizophrenia. Fig. 2 presents a systems level perspective on deficits in the motor system in schizophrenia, yet effects at this level can also be associated with evidence of functional alterations at the molecular scale in schizophrenia-relevant neurotransmitter systems. In the following section, we explore the synthesis between dopamine and dysfunctional motor control in schizophrenia.
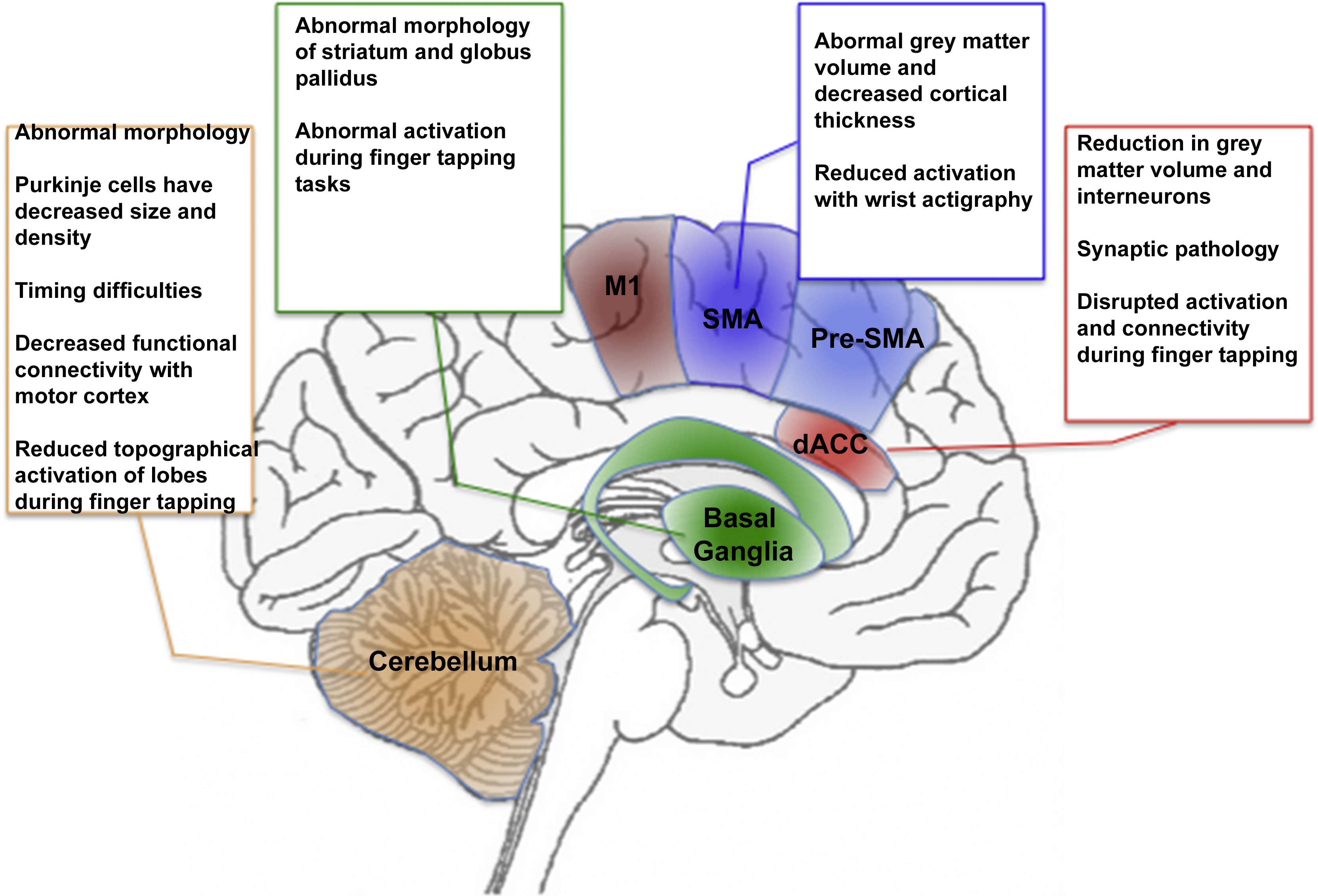
Fig. 2 Fig. 1 is adapted to summarize some of the known findings in schizophrenia. As seen in the text boxes (and more extensively discussed in the text), regions central to motor behavior also are characterized by widespread and multifaceted abnormalities in schizophrenia. These include abnormal structure (at multiple scales) and aberrant responses during motor tasks. The color and naming schemes are maintained from Fig. 1.
5. Dopamine and motor dysfunction in schizophrenia
In 1966 van Rossum proposed an early version of the “dopamine hypothesis”, suggesting that the illness may be related to the pathophysiological overproduction of dopamine or overstimulation of dopamine receptors [Reference Madras115]. Evidence in the 1950s associating Parkinson's disease with a deficiency of dopamine, laid the framework for altered dopamine transmission as the cause of extrapyramidal disturbances seen with antipsychotics, and later, the underlying pathology in schizophrenia itself [Reference Baumeister and Francis116]. By 1975, the dopamine hypothesis was substantiated by direct binding assays of antipsychotics to dopamine receptors [Reference Seeman and Lee117].
Dopaminergic projections most associated with schizophrenia involve the mesocortical system [Reference Fallon, Opole and Potkin118]. This system arises in the Ventral Tegmental Area (VTA) and projects to multiple cortical areas including the prefrontal cortex, anterior cingulate cortex, and areas of the temporal lobes. Alterations in dopamine signaling of this projection, as well as the mesolimbic and nigrostriatal projections are likely to be involved in schizophrenia. Higher dopamine levels are found in the substantia nigra and the striatum of the basal ganglia, with lower levels found in the cortex of schizophrenia patients [Reference Meador-Woodruff, Haroutunian, Powchik, Davidson, Davis and Watson119]. In neuroleptic-naive patients, an overactive striatal dopaminergic system is thought to result from changes in dopamine synthesis, storage, and release [Reference Perez-Costas, Melendez-Ferro and Roberts63], and indeed, molecular imaging studies found increased pre-synaptic dopamine synthesis and storage in the striatum of schizophrenia patients [Reference McGowan, Lawrence, Sales, Quested and Grasby120]. This elevated striatal dopamine function has been strongly associated with motor dysfunction and psychotic symptoms [Reference Howes and Kapur121].
Dopamine receptor abnormalities, especially D1 and D2, likely underlie some of the behavioral and abnormal motor symptoms seen in schizophrenia [Reference Laruelle122]. In the nigrostriatal projection, dopamine is produced and synthesized in the dopaminergic neurons in the substantia nigra, in turn stimulating movement via the direct pathway through D1 receptors. Dopamine inhibits the indirect pathway, suppressing movement, by binding to the D2 receptors that are on the indirect pathway's GABA neurons. In the striatum, excessive stimulation of D2 receptors has been found in patients diagnosed early in the disease course [Reference Cazorla, de Carvalho, Chohan, Shegda, Chuhma and Rayport123]. This hyper-stimulation is thought to result from highly sensitive D2 receptors, and it has been shown that schizophrenia patients who are exposed to dopamine-related drugs such amphetamine or methylphenidate often present with exacerbated psychotic symptoms [Reference Seeman and Seeman124].
In addition to hypersensitivity, abnormal density of dopamine receptors may also impact motor areas implicated in schizophrenia. A decreased density of D2 receptors is found in the ACC, with higher density of D2 receptors found in the striatum [Reference Tune, Wong, Pearlson, Strauss, Young and Shaya125,Reference Suhara, Okubo, Yasuno, Sudo, Inoue and Ichimiya126]. In patients on antipsychotic treatment regimens, density of D2 receptors in the striatum is subsequently reduced, and is associated with an abnormal reduction in the speed of finger tapping [Reference Yang, Chiu, Chen, Chen, Yeh and Lee127]. In a study where mice had up-regulated D2 receptors, increased inhibition of the indirect pathway was found, suggesting that the increased D2 receptor expression and increased striatal excitability could lead to reversible changes that would alter the way in which the direct pathway is activated in schizophrenia [Reference Cazorla, de Carvalho, Chohan, Shegda, Chuhma and Rayport123]. This disruption of balance of the pallidal pathways, mediated by dopamine signaling, may play a role in the development of dysfunctional motor control. The link between putative pharmacologic mechanisms and treatment response, has led to the notion of dopamine (particularly striatal) sensitivity as a “final common pathway” in schizophrenia [Reference Howes and Murray64], though given the complex nature of dopamine signaling, it is unlikely that a single hypothesis is responsible for all the manifestations of the illness [Reference Fallon, Opole and Potkin118]. Fig. 3 (consistent with the format of previous figures), depicts the implicated dopamine pathways with a summary of pertinent findings in schizophrenia.
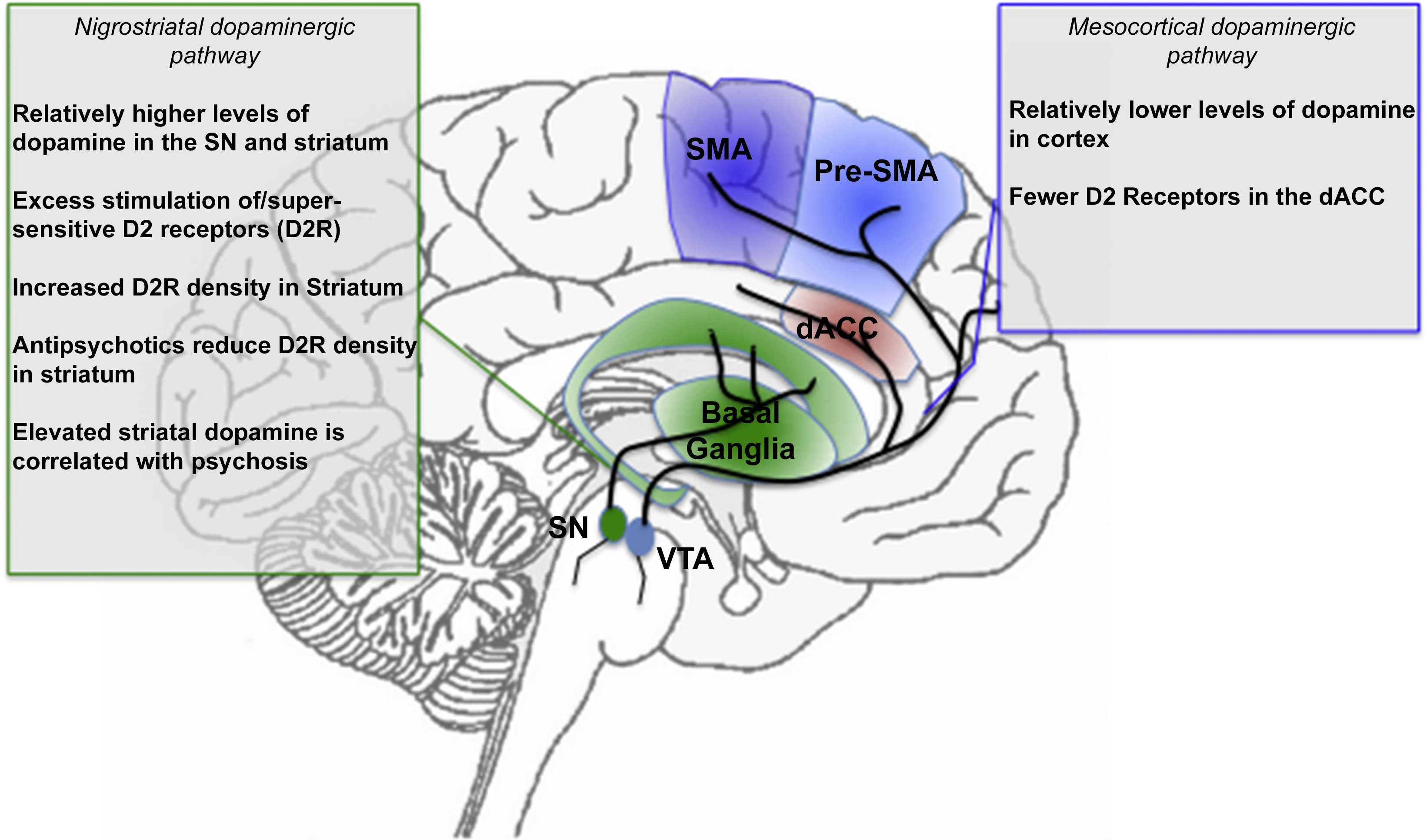
Fig. 3 A summary of dopamine projections and implications for schizophrenia. The nigrostriatal dopaminergic projection system originates from the Substantia Nigra (SN), whereas the mesocortical dopaminergic projection system originates from the Ventral Tegmental Area (VTA). These systems project to the basal ganglia, and the motor and cingulate regions respectively (see marked projections). The text boxes summarize evidence of dysfunction in schizophrenia. The color and naming schemes are consistent with previous figures.
6. Implications and Conclusions
Schizophrenia is a “multi-level” disorder with aberrations noted at micro-, meso- and macroscopic scales in the neuronal substrate. Yet, its origins are obscure and its pathophysiologic bases can only be reached through a process of “inference” from brain signals [Reference Silverstein, Bressler and Diwadkar128]. The organization of motor function, certainly at the macroscopic scale is remarkably well understood (Fig. 1), and as our synthesis suggests, aspects of motor function, intersect with questions of interest in the pathophysiology of schizophrenia (Figs. 2-3).
Individuals with schizophrenia are characterized by a diverse set of motor symptoms, among them involuntary, uncoordinated, and inhibited movement. As noted, dyskinesia is frequently medication induced, but motor symptoms were noted long before the discovery of first-generation anti-psychotics, and frequently predate the onset of the illness. Given that motor symptoms are at the core of schizophrenia-related pathophysiology and provide a relatively well-understood mapping to brain systems, the motivation to study the neurobiological bases of motor dysfunction in schizophrenia is obvious. These motivations are not isolated. There is a vivid history of studying the oculomotor system in the context of schizophrenia and psychosis, in part because of the ready translational framework, and because motor control (or lack thereof) within the eye movement system can provide fundamental insights into the bases of psychosis [Reference Thakkar, Diwadkar and Rolfs20]. Thus, a framework involving motor dysfunction measured by neuroimaging of both neurotransmitter systems and brain activity may reconcile and delineate non-homogeneous findings and help in categorizing endophenotypes of the disease. Moreover, the pathophysiology of brain regions known to be involved in motor function may help inform understanding of disturbed cognitive functions in schizophrenia, given that thinking may be an active motor process. With no single underlying etiology to connect these dysfunctions to clinical features, an exploratory approach involving task-underpinned neuroimaging will be essential. Basic motor control and behavior are linked with a rich set of cortical and sub-cortical brain regions [Reference Witt, Laird and Meyerand10], an aspect that is appropriately recognized in the Research Domain Criteria constructs matrix [Reference Morris and Cuthbert129]. Whether the explication of domain related effects will ultimately inform and transform the diagnostic concept of schizophrenia, is itself an open question [Reference Morris, Vaidyanathan and Cuthbert130,Reference Sprooten, Rasgon, Goodman, Carlin, Leibu and Lee131], yet one worthy of investigation.
Disclosure of interest
The authors declare that they have no competing interest.
Acknowledgements
Preparation of this work was supported by a Career Development Chair from Wayne State University, the Charles H. Gershenson Distinguished Faculty Fellowship from Wayne State University, the Lyckaki-Young Fund from the State of Michigan, the Prechter Family Bipolar Foundation, the Children's Hospital of Michigan Foundation, the Children's Research Center of Michigan, the Cohen Neuroscience Endowment, and the National Institute of Mental Health under Grant No MH 59299, MH111177.



Comments
No Comments have been published for this article.