Refine listing
Actions for selected content:
Contents
Research Article
Aberrant ascus genotypes from crosses involving mutants at the g locus in Sordaria fimicola
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 229-250
-
- Article
-
- You have access
- Export citation
Population structure in artificial selection: studies with Drosophila melanogaster
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 113-126
-
- Article
-
- You have access
- Export citation
Cytogenetic and complementation analyses of recessive lethal mutations induced in the X chromosome of Drosophila by three alkylating agents
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 1-10
-
- Article
-
- You have access
- Export citation
A note on increasing the limit of selection through selection within families
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 127-135
-
- Article
-
- You have access
- Export citation
The role of latent viruses in subline differentiation in inbred strains of mice
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 11-17
-
- Article
-
- You have access
- Export citation
Genetic analysis of recombination at the g locus in Sordaria fimicola
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 251-279
-
- Article
-
- You have access
- Export citation
Uncorrelated genetic drift of gene frequencies and linkage disequilibrium in some models of linked overdominant polymorphisms
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 281-294
-
- Article
-
- You have access
- Export citation
Genetic regulation and pattern formation: A study of the yellow locus in Drosophila melanogaster
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 19-26
-
- Article
-
- You have access
- Export citation
Fitness of allozyme variants in Drosophila pseudoobscura: II. Selection at the Est-5, Odh and Mdh-2 loci*
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 137-149
-
- Article
-
- You have access
- Export citation
Bare-patches, a new sex-linked gene in the mouse, associated with a high production of XO females: II. Investigations into the nature and mechanism of the XO production
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 27-41
-
- Article
-
- You have access
- Export citation
Selection for growth-rate during asexual and sexual propagation in Phytophthora cactorum
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 295-309
-
- Article
-
- You have access
- Export citation
The effect of population subdivision on two loci without selection
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 151-162
-
- Article
-
- You have access
- Export citation
-
This paper is devoted to the study of the effects of population subdivision on the evolution of two linked loci. Two simple deterministic models of population subdivision without selection are investigated. One is a finite linear ‘stepping stone’ model and the other is a finite linear stepping stone chain of populations stretching between two large populations of constant genetic constitution. At equilibrium in the first model the gene frequencies in each population are equal and there is linkage equilibrium in each population. The rate of decay to zero of the linkage disequilibrium functions is the larger of (1 – c) and
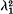 , where λ1 is the rate of convergence of the gene frequencies to equilibrium and c is the recombination frequency. In the second model at equilibrium there will be a linear cline in gene frequencies connecting the two large constant populations. This cline will be accompanied by a ‘cline’ of linkage disequilibria. The rate of convergence to this equilibrium cline is independent of the recombination frequency, and, in fact, the gene frequencies and the linkage disequilibria converge to equilibrium at the same rate.
, where λ1 is the rate of convergence of the gene frequencies to equilibrium and c is the recombination frequency. In the second model at equilibrium there will be a linear cline in gene frequencies connecting the two large constant populations. This cline will be accompanied by a ‘cline’ of linkage disequilibria. The rate of convergence to this equilibrium cline is independent of the recombination frequency, and, in fact, the gene frequencies and the linkage disequilibria converge to equilibrium at the same rate.
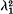 , where λ
, where λThe induction of ouabain-resistant mutants by N-methyl-N′-nitro-N-nitrosoguanidine in Chinese hamster cells
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 311-314
-
- Article
-
- You have access
- Export citation
Mitochondrial genetics, circular DNA and the mechanism of the petite mutation in yeast
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 43-57
-
- Article
-
- You have access
- Export citation
Genetic analysis of larval feeding behaviour in Drosophila melanogaster
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 163-173
-
- Article
-
- You have access
- Export citation
Methods for the isolation and analysis of plasmodial mutants in Physarum polycephalum
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 175-187
-
- Article
-
- You have access
- Export citation
Regulation of enzyme activities in Drosophila: I. The detection of regulatory loci by gene dosage responses
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 59-72
-
- Article
-
- You have access
- Export citation
Footpad bristles: a convenient metric character in mice
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 315-321
-
- Article
-
- You have access
- Export citation
The lactose system in Klebsiella aerogenes V9A: 4. A comparison of the lac operons of Klebsiella and Escherichia coli
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 323-331
-
- Article
-
- You have access
- Export citation
Genetic studies on male sterility of hybrids between laboratory and wild mice (Mus musculus L.)
-
- Published online by Cambridge University Press:
- 14 April 2009, pp. 189-206
-
- Article
-
- You have access
- Export citation