

Materials and methods
The description is based on a single, well-preserved, articulated, and almost complete specimen, including the distal-most caudal vertebrae (Fig. 1). It was collected from the Guanling Formation of Luoping County in Yunnan Province. It was prepared by, and is now housed in, the Institute of Vertebrate Palaeontology and Palaeoanthropology in Beijing.

Figure 1 Pectodens zhenyuensis n. gen. n. sp. (1) Photograph and (2) line drawing of the holotype IVPP V18578. Abbreviations: as=astragalus; ca=calcaneum; cav=caudal vertebra; cl=clavicle; co=coracoid; cv=cervical vertebra; dv=dorsal vertebra; fe=femur; fi=fibula; ga=gastralia; hu=humerus; hy=hyoid; il=ilium; is=ischium; mc=metacarpal; mt=metatarsal; pu=pubis; r=rib; ra=radius; rad=radiale; sc=scapula; t=tibia; ul=ulna; uln=ulnare.
Repository and institutional abbreviation
IVPP, Institute of Vertebrate Palaeontology and Palaeoanthropology, Beijing, China.
Systematic paleontology
Reptilia
Archosauromorpha von Huene, Reference Huene1946
?Protorosauria Huxley, Reference Huxley1871
Family incertae sedis
Genus Pectodens new genus
Type species
Pectodens zhenyuensis n. gen. n. sp. by monotypy.
Diagnosis
As for type species.
Etymology
From the Latin pecto meaning to comb and dens meaning teeth; in reference to the comb-like nature of the marginal dentition.
Occurrence
Luoping County of Yunnan Province, China; Member II of the Guanling Formation, Anisian, Middle Triassic.
Pectodens zhenyuensis new species
Holotype
IVPP V18578. Almost complete articulated skeleton.
Diagnosis
Small, gracile diapsid (about 38 cm long), with long tail and neck, ten premaxillary teeth, numerous elongate conical teeth forming comb-like marginal dentition, palatal teeth at least on the pterygoid present, temporal region short. At least ten, and probably eleven, or even twelve elongate cervical vertebrae bearing ribs with a short anteriorly projecting process and a long posterior shaft that extend parallel to the neck and bridge at least two intervertebral joints. Dorsals with prominent transverse processes that terminate in subcircular facets for the ribs. Between eleven and thirteen dorsal, two sacral, and 41 caudal vertebrae. The tibia as long, or just slightly longer than, the femur. An elongate metatarsal V that is not hooked.
Occurrence
Luoping County of Yunnan Province, China; Member II of the Guanling Formation, Anisian, Middle Triassic.
Description
Positioned on two blocks, the specimen is preserved with the right side of the skull and neck exposed, but most of the rest of the skeleton is preserved in ventral view. The skull has deflected backwards so that it underlies the proximal tail, pelvis and right leg. The right manus is also partly obscured by the anterior cervicals. The two blocks were separated along a relatively clean break, but unfortunately the break passes through the pelvic girdle and some details of the relationships of the individual pelvic elements have been lost. The left femur has also been lost together with a very small part of one of the cervical vertebrae. Because of the way the ribs are oriented, it has proved impossible to clear the matrix completely away from the dorsal vertebral series, particularly from the posterior-most dorsals.
The skull.—
The skull is well preserved (Fig. 2.1, 2.2) and has a relatively short temporal region. There are at least 10 tooth positions on the premaxilla and more than 24 on the maxilla. Of these, only two or three tooth positions have lost their teeth. These very characteristic teeth (Fig. 2.3) are elongate and narrow—the first tooth on the premaxilla is 1.0 mm long—and together provide a comb-like appearance to the jaws. The teeth have weakly expressed and rather broad longitudinal enameled ridges.

Figure 2 Pectodens zhenyuensis n. gen. n. sp. (1) Photograph and (2) line drawing with the overlying postcranial elements removed. (3) Detail of the second premaxillary tooth. Abbreviations: a=angular; ar=articular; d=dentary; ec=ectopterygoid; f=frontal; j=jugal; l=lachrymal; mx=maxilla; n=nasal; pf=postfrontal; pm=premaxilla; po=postorbital; prf=prefrontal; pt=pterygoid; q=quadrate; sa=surangular; sq =squamosal.
An elongate medial process on each premaxilla separates the external nares and extends back to the nasal. The nasals appear to be broad and at least as long as, or longer than, the frontals. Paired frontals appear rather short although their anterior and posterior sutures are not clear. There is a prominent right prefrontal but determining details of the surrounding sutures is difficult. There appears to be a well-defined lacrimal (Fig. 2). The right squamosal is preserved, and its suture with the postorbital is clear (Fig. 2). The parietals appear to be completely covered by the right ilium (Fig. 2.1). The postorbital process of the jugal is thin and elongate extending posterodorsally to reach the postorbital and squamosal. There is no evidence for a posterior process on the jugal. Elements of the palate can be seen within the orbit; pterygoid teeth are small but clearly present. The full extent of these denticles is unclear, but ~15 teeth are exposed.
The orbit is relatively large, which may indicate immaturity of the individual. The skull is 25.7 mm long and the orbit, as preserved, is 10.5 mm long. However, the displacement of the prefrontal has caused a little distortion to the orbit so that in life it may have been a little shorter. The snout is ~11.7 mm long.
Two small rod-like elements with blunt ends lie in the vicinity of the anterior cervicals and the quadrate on the right side (Fig. 2). These almost certainly represent paired ceratobranchials.
The right mandibular ramus is incomplete, with the anterior part of the dentary missing, but as preserved it is 23.3 mm long. When complete it was probably ~25.0–26.0 mm long.
The vertebral column.—
The specimen is preserved essentially in complete articulation, but it is still a little difficult to be sure of the precise number of vertebrae in each specific region. In particular, there is a gap in the cervical series, while the pectoral girdle covers the anterior dorsals making it especially difficult to determine the boundary between the cervicals and dorsals. However, we consider it most likely that the new taxon had a total of between 66 and 68 vertebrae, with 11 or possibly even 12 cervicals, between 11 and 13 dorsals, two sacrals, and 41 caudals. The neurocentral sutures are closed.
The neck is approximately equal in length to the trunk (Tables 1, 2) and it seems likely that the skull is in natural articulation with the cervical series and has not disarticulated from the neck. The anterior part of the neck is obscured by matrix along with elements of the right manus (Fig. 1). The remains of eight cervical vertebrae are clearly visible and a bundle of four ribs positioned anterior to the first visible cervical vertebra (Fig. 3) indicates the occurrence of two more cervicals at the anterior end. Furthermore there would have been a much-reduced atlas giving a minimum cervical count of 11. However, in this region there may also be a sufficient gap to accommodate an additional vertebra.

Figure 3 Line drawing of the cervical series in the holotype of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578. Abbreviations: cl=clavicle; cv=cervical vertebra; hy=hyoid; r=rib; sc=scapula.
Table 1 Measurements (mm) of the individual cervical vertebrae in the holotype specimen of Pectodens zhenyuensis n. gen. n sp., IVPP V18578. Lengths are measured from the ends of the zygapophyses.

+ The ribs partly obscure the ventral edge.
* A fracture passes through the bone.
Table 2 Measurements (mm) of the individual dorsal vertebrae in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578. CL=centrum length; TPL=length of transverse process.

* Presumed to be completely covered by elements of the pectoral girdle
** Posterior end of the centrum covered by matrix and gastralia
Many of the cervical ribs cross at least two (and possibly even three) intervertebral joints. As a consequence, at any given point there are at least six ribs forming a bundle alongside the ventrolateral aspect of the cervical vertebral column. The rib on the first preserved cervical (which has the anterior end covered by matrix) extends across two intervertebral joints. In the mid-region of the neck the anterior process of each rib is very prominent and extends anteriorly to partially overlap the preceding vertebra. This can be clearly seen in the third preserved vertebra. The neural spine remains low until vertebra eight. Adjacent to this vertebra there are eight rib shafts running parallel to each other, suggesting that the ribs from at least vertebra five extend back this far (three intervertebral joints). In the following vertebrae, there is a more pronounced, but still very elongate spine. The ribs on the third to last cervical extend across just one intervertebral joint. The ribs on the last two cervicals are short and extend barely beyond the length of the centrum.
There are 11 definitive dorsals, and possibly as many as 13. The facet for the rib in the first dorsal is pronounced, anteroposteriorly elongate (almost elliptical) and extends dorsally from a position low down on the centrum onto the neural arch. The articulation for the rib is broadest at the most postero-dorsal part of the process. The process is not as laterally elongate as in more-posterior dorsals. The rib associated with this vertebra is much longer than the preceding cervical rib, but it does have a small anterior “spur” on it. We therefore regard this as the first dorsal.
Vertebra 15, and possibly also 13 and 14 (but they are covered), has a very pronounced transverse process (probably at least 3 mm long) (Fig. 1). It is narrow with a ventral ridge and depressions anteriorly and posteriorly where it attaches to the neural arch. There are no clear rib heads exposed, apart from the 12th one and possibly one lying just under the transverse process on 16. Therefore it cannot be said with certainty what form these took, but judging from the rather circular to ovoid ends to the transverse processes it can be assumed that they were holocephalous ribs with oval heads, which is consistent with the presumed head on rib 16. The mid-dorsal vertebral centra are somewhat elongate, at least they are similar in length to the most posterior cervical vertebrae. Again it would seem that the transverse processes on all the dorsals are long and slender as in vertebra 15.
Vertebra 23 has what appears to be a small anterior-projecting transverse process (parapophysis? – fused transverse process and rib), but it is incomplete distally and it is probably a broken rib (Fig. 1). Vertebra 24 exhibits the anterior part of the centrum, but the rest is lost in the fracture between the two main blocks. Nevertheless, there are two rib shafts running into that fracture and these were most probably associated with vertebra 24. A rib adjacent to vertebra 22 is clearly flexed.
The sacrals are difficult to determine as the fracture runs through this part of the specimen. However, there would seem to be sufficient space to accommodate two sacrals, assuming that vertebra 27 is indeed the first caudal and that its transverse process did not contact the pelvis.
The anterior caudals are mostly preserved in ventral view and it is therefore not possible to make any comment on the nature of the neural spine. From caudal 10 onwards they have twisted over onto their side, and it is clear that the neural spine was anteroposteriorly elongate and low (Fig. 1). Anterior caudals bear long transverse processes (Table 3) and the facets for the chevron bones first appear on vertebra 29. The first chevron lies alongside vertebra 30 and is on its side so that the broad distal end is clearly exhibited. For the next four vertebrae, the chevrons have flipped and are visible more or less end on, but on vertebra 34 the chevron is again in fully articulated position and the distal end is seen to have an even more expanded form. On the next vertebra the chevron loses that shape and the anterior lamina/projection disappears altogether. It is also at this point that the transverse processes are lost. In some respects then, the caudal region is similar to Tanystropheus, as figured by Wild (Reference Wild1973, p. 67).
Table 3 Measurements (mm) of the individual caudal vertebrae in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578. CL=centrum length; TPL=length of transverse process.

The pectoral girdle and forelimb.—
The scapula appears dorsoventrally low and similar in overall form to the coracoid. One scapula seems to be lying on top of the other (Fig. 1). Both coracoids are displayed and the coracoid foramen is clear on the left side. The right coracoid is a little fragmented around the edges. Either one of the clavicles, or possibly a small exposure of the interclavicle, extends across the last cervical and towards the scapula. It is quite compressed, but nothing more can be said about this element.
The humeri are long slender elements but with prominent deltopectoral crests contributing to markedly expanded proximal heads. The distal ends of both humeri appear to be very narrow and expand little beyond the diameter of the shaft; however, it is impossible to determine whether they were expanded in ventral aspect. The humeri are somewhat longer than the epipodials, which are also relatively slender elements (Table 4). The ulna has a slightly more robust proximal head than the radius and it is also marginally longer.
Table 4 Measurements (mm) of the individual forelimb elements in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578. L=Length; PW=width of proximal head.

The left manus only preserves two ossifications (presumably the ulnare and radiale), although there is a large gap between these ossifications and the metacarpals, and the metacarpals are in complete articulation with the phalangeal elements (Fig. 1). This suggests that there were other elements that had not ossified—a feature that is consistent with the specimen being a juvenile.
Metacarpals and phalanges are relatively slender elements with quite marked articular expansions. The metacarpals (Table 5) and phalanges of the left manus are in complete articulation. Nevertheless, in digit V, the complete phalangeal formula is difficult to determine and it is unclear whether it has three or four phalanges.
Table 5 Measurements (mm) of the individual metacarpals in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp. IVPP V18578.
The right manus overlies the anterior cervicals and is partly obscured by the posterior part of the skull so that it provides even less clarity. The manual phalangeal formula is therefore 2:3:4:5:3 or 4.
The pelvic girdle and hindlimb.—
The pelvis is difficult to interpret. Parts of all three elements that are visible on both sides do seem to be compressed together so that the ventral edges of both pubes appear to be exposed (Fig. 1). The fracture unfortunately passes directly through the puboischiadic plate. There is no clear indication of a thyroid fenestra, and while there is a slight emargination of the anterior edge of the ischium it appears to represent a broken edge.
The femur is a relatively straight and slender element with little expansion of the proximal and distal heads. The tibiae and fibulae are also very long, slender elements that are just a little longer than the femur (Table 6). The tibia is the more robust element of the two epipodials.
Table 6 Measurements (mm) of the individual right hindlimb elements in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578. L=Length; PW=width of proximal head; DW=width of distal head.

There are only two clear ossifications preserved in both tarsi, which are assumed to represent the proximal tarsals. In the right tarsus there is an elongate fragment of bone in the approximate position of distal tarsal four, but it almost certainly represents part of the underlying caudal vertebra rather than any tarsal element. There is, nevertheless, perhaps the merest hint of a gap between the tarsals and metatarsals. However, on the left side there is not even the slightest gap in the tarsus that might have accommodated the distal tarsals. Instead the proximal tarsals appear to be in direct articulation with metatarsals III–V. The presumed astragalus and calcaneum are circular elements with no distinguishing features—there is no indication of a perforating foramen, for example. However they are in close articulation with each other in both ankles.
The fifth metatarsal is completely straight just like the other metatarsals, and is the third longest metatarsal after metatarsals III and IV, with metatarsal I being the shortest (Fig. 1; Table 7).
Table 7 Measurements (mm) of the individual metatarsals in the holotype specimen of Pectodens zhenyuensis n. gen. n. sp., IVPP V18578.
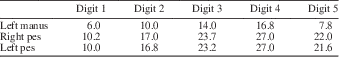
The phalanges (Table 8) are all clearly preserved in complete articulation in both feet with a pedal phalangeal formula of 2:3:4:5:4. Apart from the unguals, each phalanx has a “waisted” appearance with expanded proximal and distal ends.
Table 8 Lengths of each digit (mm), including the metacarpals and metatarsals, in each foot of the holotype specimen of Pectodens zhenyuensis n. gen. n sp., IVPP V18578.
Etymology
In honor of Zhenyu Li, who contributed greatly to the collection of the specimen from the field.
Remarks
The cervicals closely resemble those of Tanystropheus with an extremely low neural spine (Fig. 3). The centra of cervicals six, seven, and eight are eroded, presumably from poor preservation of the bone close to the surface, although it conceivably hints at hollow centra.
The number of dorsal vertebrae is unclear. The inferred 12th vertebra is considered to be the first dorsal and is mostly exposed, although the posterior end is covered by the coracoids (Fig. 1). If, as seems likely, the skeleton is still in complete articulation, then there is one dorsal vertebra entirely hidden. Furthermore, it is also difficult to determine whether the anterior edge of a centrum positioned adjacent to the crack running through the puboischiadic plate represents the last dorsal or the first sacral. On balance, it would appear most likely that this is the last dorsal rather than the first sacral and that the first sacral is completely obscured.
Unlike Tanystropheus, there is no evidence of dichocephalous ribs in the anterior-most dorsals. Unfortunately there are no dorsal vertebrae showing the nature of the neural spine and whether it was relatively tall and elongate as in tanystropheids generally.
The shape of the scapula is apparently not dissimilar to the condition seen in Macrocnemus or Tanystropheus (e.g., Peyer, Reference Peyer1937; Wild, Reference Wild1973; Nosotti, Reference Nosotti2007).
On first inspection it appears that there are only two phalanges in digit V of the left manus, but a combination of a potentially very short second phalanx together with a thin calcite vein running through the more distally preserved part of the digit, introduces considerable uncertainty. Firstly, there is a distinct swelling on the section of bone distal to the fifth metacarpal. This is best explained by assuming that the swelling represents an articulation, and that the bone actually represents two phalanges partially overlapping each other, with the second one being relatively short in comparison with the other digits. In this case, the apparent ungual phalanx would be unusually long by comparison with the unguals of the other four digits. It is therefore possible that the narrow calcite vein actually obscures an articulation between separate phalanges. If both of these interpretations are correct, then Pectodens possessed four phalanges on the fifth digit. An alternative view is that there is a matrix-filled break in the second phalanx (just adjacent to the calcite vein), which gives the impression of an articulation. In this case then there are just three phalanges—a condition that is widespread within protorosaurs.
Somewhat unusually, metatarsal V is long and slender and exhibits no indication of an offset head (or “hooked” shape) as seen in Tanystropheus (e.g., Wild, Reference Wild1973; Nosotti, Reference Nosotti2007).
Conclusions
A new, small terrestrial tetrapod is described from the Middle Triassic of Yunnan, China. Pectodens zhenyuensis n. gen. n. sp. bears very characteristic elongate teeth forming a comb-like marginal dentition. The elongate cervicals of Pectodens zhenyuensis n. gen. n. sp. with low neural spines together with the morphology of the cervical ribs are features consistent with protorosaurs, such as Macrocnemus. However, the imperforate puboischiadic plate, simple rounded proximal tarsals, and a straight 5th metatarsal are primitive characteristics. A key protorosaurian character is the long neck with elongated cervical ribs that typically extend across intervertebral articulations. It was mostly on the basis of these characters that Dinocephalosaurus from the Middle Triassic of China was referred to the protorosaurs (Li, Reference Li2003). Another long-necked form, Fuyuansaurus, also exhibits certain affinities with protorosaurs, in particular the tanystropheids (Fraser et al., Reference Fraser, Li and Rieppel2013). Yet both taxa also display characters that are inconsistent with at least the tanystropheids. Unlike tanystropheids, but in common with Protorosaurus (personal observation, N.C. Fraser, Reference Fraser, Li and Rieppel2013), both lack a thyroid fenestra in the pelvis.
The Middle Triassic vertebrate faunas of southern China are largely dominated by marine reptiles and fishes, but occasional terrestrial components (e.g., Macrocnemus fuyuanensis) are recovered from localities in the Zhuganpo Member of the Falang Formation (Li et al., Reference Li, Zhao and Wang2007; Jiang et al. Reference Jiang, Rieppel, Fraser, Motani, Hao, Tintori, Sun and Sun2011). Likewise, Pectodens zhenyuensis n. gen. n. sp. exhibits no adaptations for an aquatic lifestyle; instead the long, slender limbs with pronounced articular ends, and elongate digits together with the claw-like distal phalanges speak to an entirely terrestrial existence. No fully terrestrial vertebrates have been documented previously from the Panxian-Luoping Fauna, although the archosaur Qianosuchus mixtus exhibits a combination of terrestrial and aquatic characteristics (Li et al., Reference Li, Wu, Cheng, Sato and Wang2006). Pectodens is therefore the first fully terrestrial reptile reported from the Guanling Formation. The occurrence of terrestrial reptiles such as Macrocnemus and Pectodens are indicative of the proximity of the ancient coastline at the localities where they occur.
Acknowledgments
This work was supported from a special grant for fossil excavation and preparation by the Chinese Academy of Sciences. We especially thank Z. Li for his assistance in collecting the new specimen. We are grateful to H. Zhou for preparation of the specimen, W. Gao for photography, and W. Yu for the drawings. NCF acknowledges the European Union SYNTHESYS fund for travel support. He also thanks U. Göhlich (Natural History Museum Vienna), D. Schwarz-Wings (Museum für Naturkunde Berlin), and M. Farrell and K. Hussey (Hunterian Museum, Royal College of Surgeons England) for access to Protorosaurus material in their care. Anonymous reviewers provided helpful comments on an earlier version of the manuscript, which was handled by editors B. Hunda and D. Varricchio.











