



Introduction
Since the late 1960s, joint Mongolian-Russian and Mongolian-Polish paleontological expeditions have contributed greatly to the characterization of Cenozoic stratigraphy and paleontology in Mongolia (see review in Lopatin, Reference Lopatin2020). Detailed descriptions of stratigraphic and paleontological evidence from localities in the region of the Nemegt Basin, known as Naran Bulak (Fig. 1), and exposures farther west at Tsagan Khushu have led to the characterization of the Naran Bulak Formation in the region directly south of the Altan Mountains (Altan Uul). This formation is a terrestrial series of sandstones, clays, and carbonates that has been interpreted as having lacustrine and fluvial sediments (Dashzeveg, Reference Dashzeveg1988; Meng et al., Reference Meng, Zhai and Wyss1998).
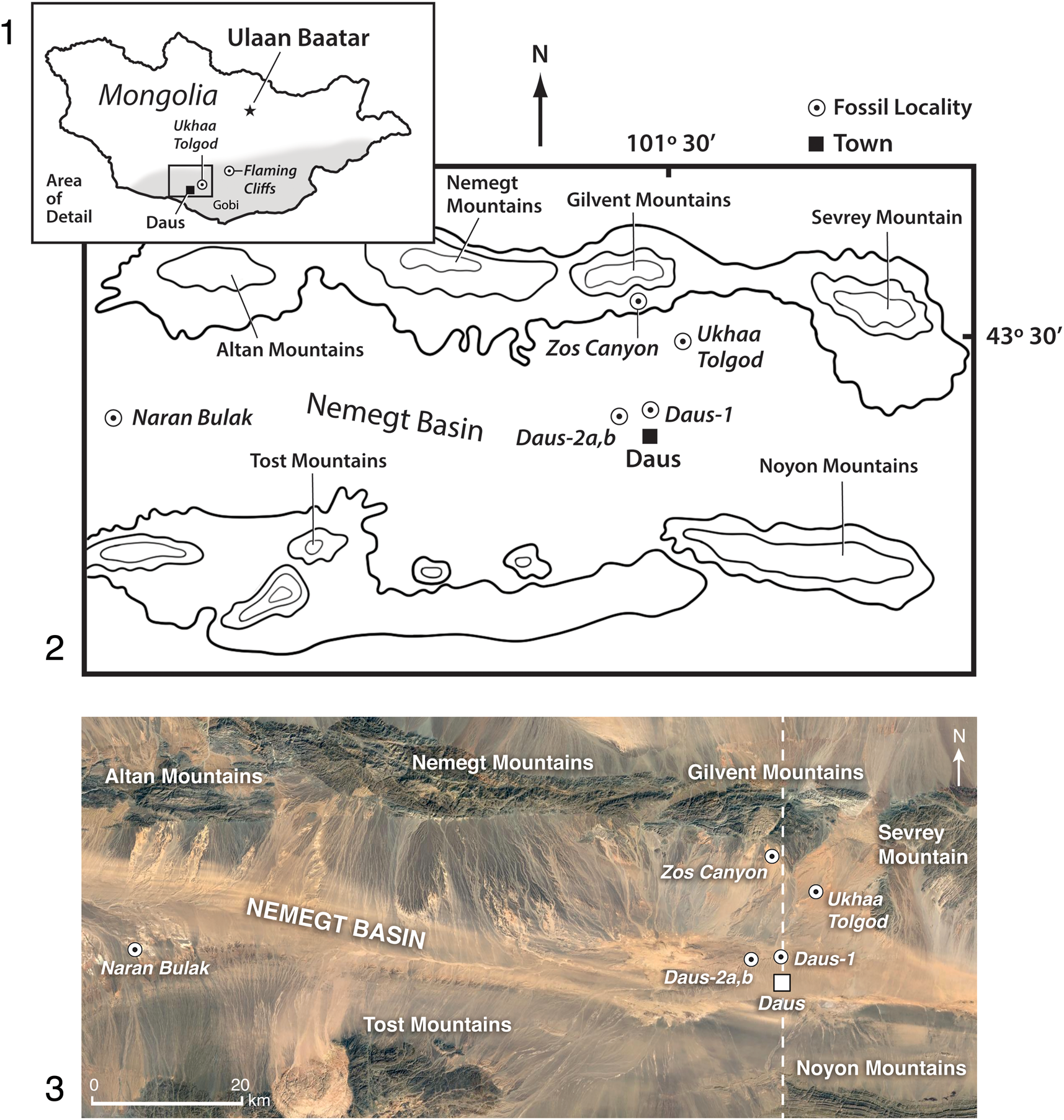
Figure 1. (1) Map of Mongolia and the Gobi Desert (gray shading) where the new fossils were found; boxed area is expanded in (2, 3). (2) Map of Nemegt Basin and adjacent mountains showing fossil localities (italicized text) relative to nearby towns; and (3) the same region shown as a Google Earth satellite image that reveals the relief of the localities and their south-trending exposure; dotted line is the transect shown in Figure 3. New Paleogene localities are Daus-1, Daus-2a, and Daus-2b, which are near the highly productive Late Cretaceous locality Ukhaa Tolgod. The locality Zos Canyon extends east from where it is marked in (2) and the locality Ukhaa Tolgod extends southwest. The type locality of the Paleogene Naran Bulak Formation is separated from the Daus localities by ~90 km. The Late Cretaceous locality Ukhaa Tolgod indicated is described elsewhere (Pol and Norell, Reference Pol and Norell2004; Dingus et al., Reference Dingus, Loope, Dashzeveg, Swisher, Minjin, Novacek and Norell2008).
Until now, the Naran Bulak Formation has been identified only in the western part of the Nemegt Basin (Dashzeveg, Reference Dashzeveg1988, fig. 1), however, we show that the central-eastern region of the Nemegt Basin also contains Naran Bulak Formation exposures. Our discovery occurred during extensive paleontological expeditionary work conducted at the highly fossiliferous locality Ukhaa Tolgod. Discovered in 1993 by a Mongolian Academy-American Museum Paleontological Expedition (MAE), this Upper Cretaceous Djadokhta Formation locality is noteworthy for its numerous, well-preserved dinosaur skeletons, embryos, and eggs, as well as fossil mammals, lizards, and other vertebrates (Norell et al., Reference Norell, Clark, Dashzeveg, Barsbold, Chiappe, Davidson, Mckenna and Novacek1994; Dashzeveg et al., Reference Dashzeveg, Novacek, Norell, Clark, Chiappe, Davidson, McKenna, Dingus, Swisher and Altangerel1995; Novacek et al., Reference Novacek, Rougier, Wible, McKenna, Dashzeveg and Horovitz1997; Clark et al., Reference Clark, Norell and Chiappe1999). Beginning in 1993, outcrops of stratigraphically higher rocks located 12 km to the southwest of the main section at Ukhaa Tolgod were first prospected (Fig. 1).
This more southerly unit initially appeared to be unfossiliferous until the 2014–2015 Mongolian Academy-American Museum of Natural History field seasons when the team discovered ostracodes and fragmentary teeth and bones of mammals. Subsequently, in 2017 and 2019, a concentration of mammalian fossils, including identifiable jaws and dentitions, were found in white beds at three localities, Daus-1, Daus-2a, and Daus-2b, located near the town of Daus in the Omnigov Imac province (Figs. 1–3). The sedimentary features, color, and stratification of the section at these new localities resemble those of the type section of the Naran Bulak Formation, and the specific fossil-yielding white beds we found correspond to the Naran Member of the Naran Bulak (Fig. 2; Dashzeveg, Reference Dashzeveg1988). Lopatin (Reference Lopatin2020) summarized the vertebrate fossils that have been recovered from the western exposures of the Naran Member (“White Beds”), including amiid fishes, crocodyliforms, aves, and mammals such as archaeolambdids, arctostylopids, multituberculates, mesonychians, coryphodontids, and the rodent Tribosphenomys, among other taxa.

Figure 2. (1) Naran Bulak Formation stratigraphy. Fossils described here come from the Naran Member (“White Beds”) of this formation. Three sections are summarized. The section described by Gradziński (Reference Gradziński1969) was later designated the type section by Dashzeveg and McKenna (Reference Dashzeveg and McKenna1977), who specified that although the Upper Red Beds are not found in the Gradziński (Reference Gradziński1969) section, they are to be included in the type Naran Bulak Formation. Dashzeveg (Reference Dashzeveg1982) provided names for the Naran Bulak Formation members. Discussions in these papers reveal disagreement over whether the upper part of the Naran Bulak Formation is Paleocene or Eocene. We treat the Zhigden, Naran, and Bumban beds as Paleogene for lack of additional temporal control. (2) Our ~30 m section at the Daus-1 locality representing all of the same Naran Bulak beds as the type section, except the lowest, conglomeratic “Pink Beds.” (3) Photograph of the Daus-1 locality with an explanatory schematic below indicating the Daus-1 locality within the Naran Member (“White Beds”) above an indurated sandstone that we interpret as a channel deposit. The part of section visible in the photograph is ~19 m high.
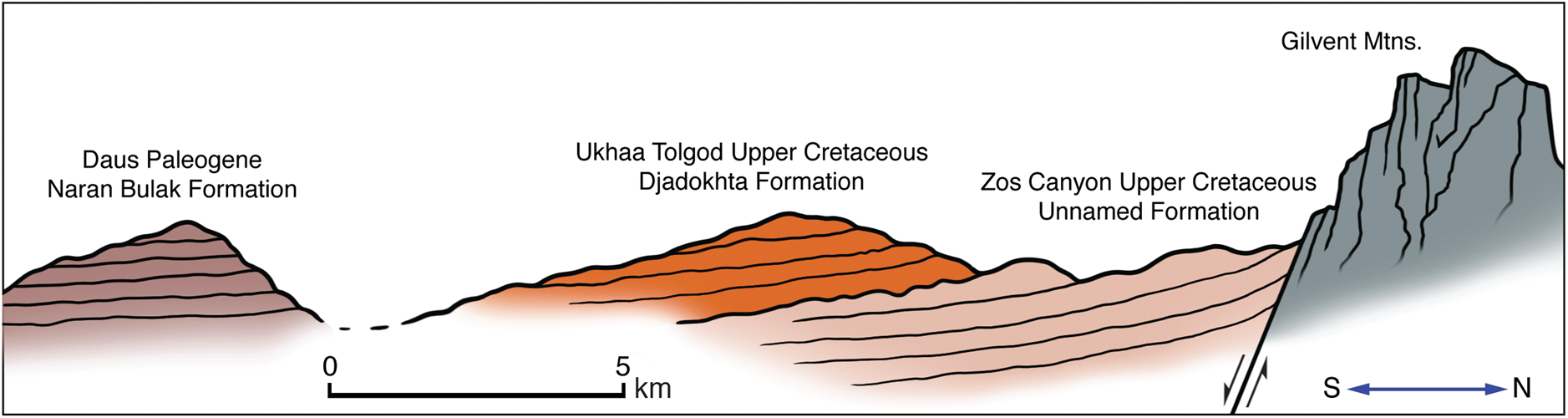
Figure 3. Schematic showing cross-section of the Naran Bulak Formation lying unconformably on the Upper Cretaceous Djadokhta Formation, which in turn overlies an older, unnamed Upper Cretaceous formation exposed in Zos Canyon (Pol and Norell, Reference Pol and Norell2004).
Due to a lack of marine and volcanic deposits in Central Asian regions, including the Nemegt Basin of the Gobi Desert, Mongolia (Dashzeveg, Reference Dashzeveg1988; Meng et al., Reference Meng, Zhai and Wyss1998; Ting, Reference Ting1998; Dingus et al., Reference Dingus, Loope, Dashzeveg, Swisher, Minjin, Novacek and Norell2008), the age estimate of the Naran Bulak Formation has been limited to biostratigraphy (Gradziński, Reference Gradziński1969; Szczechura, Reference Szczechura1971; Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977; Dashzeveg, Reference Dashzeveg1982, Reference Dashzeveg1988; Khand, Reference Khand1987; Beard, Reference Beard1998; Lucas, Reference Lucas, Aubry, Lucas and Berggren1998; Wang et al., Reference Wang, Meng, Beard, Li, Ni, Gebo, Bai, Jin and Li2010). The Naran Bulak Formation is thought to represent two key Asian Land Mammal Ages (ALMAs): the Gashatan and the Bumbanian, with the former historically considered to be of latest Paleocene age and the latter generally considered the earliest Eocene (Dashzeveg, Reference Dashzeveg1982, Reference Dashzeveg1988; Russell and Zhai, Reference Russell and Zhai1987; Ting, Reference Ting1998). More recent assessments, including those that tied biostratigraphy to magnetostratigraphy and chemostratigraphy, showed that while the Gashatan ALMA was entirely Paleocene, the Bumbanian ALMA may contain the Paleocene-Eocene boundary (Meng et al., Reference Meng, Zhai and Wyss1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002; Ting et al., Reference Ting, Bowen, Koch, Clyde, Wang, Wang and McKenna2003, fig. 4). However, Bowen et al. (Reference Bowen, Koch, Meng, Ye and Ting2005) refined this hypothesis with new data indicating that the Gashatan ALMA persisted close to the Paleocene-Eocene boundary.
Below we describe for the first time the general lithology and paleontology of three Naran Bulak Formation localities found >90 km east of the Naran Bulak Formation type section (Gradziński et al., Reference Gradziński, Kazmierczak and Lefeld1969; Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977; Dashzeveg, Reference Dashzeveg1988). The fossil occurrences at this site and the extensive lateral exposure of this unit indicate that there is a much more widespread distribution of Paleogene rocks in the Nemegt Basin south of the Altan, Nemegt, and Gilvent mountains than previously known (Figs. 1, 2).
Geological setting
Geological time unit
Early Paleogene (Fig. 2).
Stratigraphic information
The lithology of the three new localities (Daus-1, Daus-2a, Daus-2b) is geologically consistent with published accounts of the Naran Bulak Formation that crop out in the western Gobi Desert (Dashzeveg, Reference Dashzeveg1968; Gradziński et al., Reference Gradziński, Kazmierczak and Lefeld1969; Shuvalov et al., Reference Shuvalov, Reshetov and Barsbold1974; Shishkin, Reference Shishkin1975; Badamgarav and Reshetov, Reference Badamgarav and Reshetov1985; Bekhbat et al., Reference Bekhbat, Naranbaatar, Badarch and Davaa1999). Gradziński (Reference Gradziński1969) described the western Nemegt Basin Paleogene rocks at Naran Bulak, but did not explicitly name a type section. Subsequently, Dashzeveg and McKenna (Reference Dashzeveg and McKenna1977) recognized the section described by Gradziński (Reference Gradziński1969, fig. 12) as the Naran Bulak Formation type section, comprising, from lowest to highest, the conglomeratic “Pink Beds,” the “Lower Red Beds,” and the “White Beds.” Dashzeveg and McKenna (Reference Dashzeveg and McKenna1977) additionally recognized an upper unit, the “Upper Red Beds” of Gradziński et al. (Reference Gradziński, Kazmierczak and Lefeld1969), as the Bumbin-Nuru beds, which are referable to the type section of the Naran Bulak Formation even though such beds are not in the region of the type section (Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977, fig. 2). Dashzeveg (Reference Dashzeveg1988) later recognized the “Lower Red Beds” as the Zhigden Member, the “White Beds” as the Naran Member, and the “Upper Red Beds” as the Bumban Member, all of the Naran Bulak Formation. The unit below the Zhigden Member, the conglomeratic “Pink Beds” included in the Naran Bulak Formation by Dashzeveg and McKenna (Reference Dashzeveg and McKenna1977), was later recognized by Dashzeveg (Reference Dashzeveg1988, fig. 2) as a unit within the Cretaceous Nemegt Formation, a designation we follow here (Fig. 2). Dashzeveg (Reference Dashzeveg1982) also added, with reservation, an additional youngest member to the Naran Bulak Formation, the Aguyt, which has a poorly characterized fauna, and we do not discuss it further here.
According to Dashzeveg (Reference Dashzeveg1988, p. 473), the beds in the Naran Bulak area, where the type section is located, have 80 m of “alternating sandstones and clays, with intercalations of light grey carbonates; light grey and red colors predominate. The strata are horizontal and cross-bedding occurs” (see also Dashzeveg, Reference Dashzeveg1982). These Naran Bulak area rocks lie unconformably over the Cretaceous “Upper Nemegt Beds,” whose lithology was described extensively by Gradziński (Reference Gradziński1969). The lithological characterization of the type section of the Naran Bulak Formation is consistent with the stratigraphy at Daus-1, Daus-2a, and Daus-2b, although at Daus-1 the exposure is ~30 m (at Daus-2a and Daus-2b, the total section height has not yet been measured). At the Daus-1 locality there are lower red, middle white, and upper red beds, as have been described for the Naran Bulak Formation type section (Fig. 2; Gradziński et al., Reference Gradziński, Kazmierczak and Lefeld1969; Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977), that extend laterally for ~2 km. The lower red beds at Daus-1 consist primarily of coarse-grained sandstones and aleurite. The middle white beds consist of white to gray calcareous clay and equigranular sands. The upper red strata contain aleurite and inequigranular sands and sandstone. The Naran Member (“White Beds”), which yielded the new fossils, historically has been a very fossiliferous part of the section in the type region of Naran Bulak (Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977; Dashzeveg, Reference Dashzeveg1988; Lopatin, Reference Lopatin2020).
At the Daus localities, there is no evidence of exposures comparable to the Cretaceous Upper Nemegt Beds of Gradziński (Reference Gradziński1969) that unconformably underlie the type section of the Naran Bulak Formation. The nearest Cretaceous exposures lie ~1 km to north at Ukhaa Tolgod, a section described as the Upper Cretaceous Djadokhta Formation (Dingus et al., Reference Dingus, Loope, Dashzeveg, Swisher, Minjin, Novacek and Norell2008). The angle and direction of dip of the Ukhaa Tolgod section suggest that the Naran Bulak Formation at Daus rests unconformably on it (Fig. 3). The relationship of the Ukhaa Tolgod section to the younger Naran Bulak section at Daus, however, is not yet clear because the units are separated by a broad wash covered by surface pediment and vegetation.
Locality information
We present data on three new localities. The coordinates for locality Daus-1 are 43°27′20″N, 101°28′56″E in the central part of the greater Nemegt Basin, ~14.5 km south of the Gilvent Mountains of the Gurvan Tes soum (a small administrative unit under the Omnogov imac or “province”), Mongolia. Daus-1 is 10 km northwest of a salt factory at the town of Daus and 90 km east of the type Naran-Bulak section. The localities Daus-2a and Daus-2b, are located at 43°26′14″N, 101°26′17″E and 43°26′53″N; 101°26′41″E, respectively. Daus-2a and Daus-2b lie ~3 km and ~4 km, respectively, to west-southwest of Daus-1. The section at these localities is essentially the same as that described above for Daus-1, although the lower red beds assigned to the Zhigden Member at Daus-2a and Daus-2b are more poorly exposed, being confined to low hills and gullies at the base of the cliffs.
Biostratigraphy
As noted above, biostratigraphy based on fossil mammals has been the primary means of dating the Naran Bulak Formation. Fossil mammals also have been the foundation of more comprehensive efforts to establish Asian Land Mammal Ages (ALMAs; the origins of these terms are reviewed in Lucas, Reference Lucas, Aubry, Lucas and Berggren1998) to correlate terrestrial sediments within Asia and more broadly among Asian localities and those in Europe and North America. The two ALMAs most directly relevant to the new Daus localities are the Gashatan ALMA, in which the Zhigden and Naran members occur, and the Bumbanian ALMA, in which the Bumban Member occurs (Ting, Reference Ting1998).
A few key points from prior literature are most relevant to our work. The Zhigden and Naran members have been assigned to the Gashatan ALMA because they contain such taxa such Tribosphenomys, Arctostylops, Prodinoceras, and Archaeolambda planicanina Flerov, Reference Flerov1952. Of these taxa, only Arctostylops is found outside of Asia and, although the genus once served as an index taxon supporting the direct assignment of these beds to the Paleocene (Dashzeveg, Reference Dashzeveg1982, fig. 2, p. 278; Lucas, Reference Lucas, Aubry, Lucas and Berggren1998, fig. 21.22, p. 479), this role for the taxon has not withstood taxonomic reassessment. The Asian Arctostylops species, A. iturus Dashzeveg and Russell, Reference Dashzeveg and Russell1988, has been re-assigned to Palaeostylops iturus Matthew and Granger, Reference Matthew and Granger1925 (Cifelli et al., Reference Cifelli, Schaff and McKenna1989; Kondrashov and Lucas, Reference Kondrashov and Lucas2004), a species originally described by Matthew and Granger (Reference Matthew and Granger1925) from the Gashatan beds near Bayn Dzak (“the Flaming Cliffs,” see Fig. 1) and not known outside Asia. Archaeolambda planicanina Flerov, Reference Flerov1952, also may be found consistently in, and be a signature for, the Gashatan beds (Lopatin, Reference Lopatin2020), but it can only provide age data by proxy because it is not known from radiometrically dated rocks outside of Asia (note also that Dashzeveg [Reference Dashzeveg1988, fig. 4] reported its rare occurrence in Bumbanian beds, complicating matters further). The signature Gashatan taxon, Tribosphenomys, the first Asian appearance of the mammalian order Rodentia (Ting, Reference Ting1998, p. 131), is similarly constrained. Based on a combination of magnetostratigraphy, chronostratigraphy, and biostratigraphy, the Gashatan ALMA is currently considered Paleocene (Meng et al., Reference Meng, Zhai and Wyss1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002, Reference Bowen, Koch, Meng, Ye and Ting2005).
The Bumban Member has been assigned to the Bumbanian ALMA because it contains the taxa Orientolophus, Prodinoceras, Gomphos, Homogalax, and Hyopsodus, the latter two being truly Holarctic genera that appeared in the Eocene and are known from radiometrically dated sediments in North America (Woodburne and Swisher III, Reference Woodburne, Swisher, Berggren, Kent, Aubry and Hardenbol1995, fig. 1). Dashzeveg (Reference Dashzeveg1982, p. 277) mentioned the occurrence of Arctostylops in the Bumban Member of the Naran Bulak Formation of Mongolia, but this record has never been confirmed (Wang et al., Reference Wang, Meng, Ni and Beard2008). Arctostylops has not been reported in either the Gashatan or Bumbanian deposits in recent taxonomic compilations (Lopatin, Reference Lopatin2020). Ting (Reference Ting1998) argued that the appearance of the ceratomorph Orientolophus in the Bumbanian marked the first Asian appearance of the mammalian order Perissodactyla and was a useful biostratigraphic indicator. Bai et al. (Reference Bai, Wang, Li, Wang, Mao, Gong and Meng2018) more recently showed that as many as four distinct perissodactyl clades (equids, ceratomorphs, ancylopods, and brontotheres) appear to have been present in the early Eocene of Asia, but this discovery has yet to be used broadly for biostratigraphy. Gomphos, which is endemic to Asian rocks, also has been recognized as a signature for the Bumbanian ALMA. Knowledge of the actual age of Gomphos, however, currently derives from its association with Hyopsodus and Homogalax, a point noted by Dashzeveg (Reference Dashzeveg1988, p. 474, 477). The Bumbanian ALMA is considered primarily Eocene, but also may contain the Paleocene-Eocene boundary (Meng et al., Reference Meng, Zhai and Wyss1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002, Reference Bowen, Koch, Meng, Ye and Ting2005).
Finally, although microfossils are known from the Naran Bulak Formation, Dashzeveg and McKenna (Reference Dashzeveg and McKenna1977, p. 125–126) emphasized that such fossils have not been particularly useful index taxa because they are either too geographically localized or they lacked well-understood and restricted stratigraphic ranges. It is noteworthy, however, that the ostracode Limnocythere nemegtensis Szczechura, Reference Szczechura1971, which we report on here, recently has been recovered from Paleocene beds in China, strengthening its role as a signature taxon within Asia (Zhao, Reference Zhao1985; Wang et al., Reference Wang, Li, Bai, Jin, Mao and Meng2019). Later work showed that the ostracode fauna confirmed, to a certain extent, a late Paleocene age for the Zhigden and Naran members of the Naran-Bulak Formation and an early Eocene age for the the Bumban Member (Khand, Reference Khand1987).
Fossils discovered and depositional environment
Among the collected fossils are dentitions, skulls, and postcrania. Many of these elements are fragmentary and exhibit postmortem damage, thus we describe here those exhibiting diagnostic features. The gray-colored, sandy-clay composition of the Naran Bulak beds previously has been interpreted as lower alluvial and upper lacustrine horizons (Dashzeveg, Reference Dashzeveg1988).
Materials and methods
Taphonomy and preparation
The vertebrate and ostracode specimens were surface-collected from the white-gray calcareous clay and sand as in situ fossils, not as float, from the Naran Member (“White Beds” described above). The specimens, for the most part, were not found in lenses, as has been reported for more westerly localities of the Naran Bulak Formation (Dashzeveg, Reference Dashzeveg1988, fig. 2), although the fossils found at Daus-2b are possibly preserved in a thin, very restricted facies. Several fossil mammal specimens received manual preparation. The ostracode samples were immersed in water for 5–6 hours, then washed through sieves (500, 250, and 125μm). Ostracode specimens that were well preserved and particularly complete were selected for examination under a scanning electron microscope (2007 Zeiss EVO60 at the AMNH).
CT scanning
Two partial cranial specimens (Gomphos and the provisionally assigned arctostylopid) were CT scanned. Both specimens were scanned on a GE v|tome|x s240 system at the AMNH. For the Gomphos specimen, voltage was 170 kV, current was 160 μA, and exposure time was 750 ms. Voxel size was 0.0251 mm. Copper filter thickness was listed as “Unknown,” indicating that either no filter was used or the data were not entered. The parameters for the arctostylopid specimen were the same, except that the current was 170 μA, a copper filter of thickness 0.5 mm was used, and voxel size was 0.0289 mm. Segmentation and volume rendering on both scans were performed in VGSTUDIO Max 3.5.1.
Terminology
Terminology for Ostracoda follows that of Neustrueva et al. (Reference Neustrueva, Sinitsa, Khand and Melinikova2005). Terminology for mammalian teeth follows that of O'Leary et al. (Reference O'Leary, Bloch, Flynn, Gaudin and Giallombardo2013; i.e., that the primitive condition for the four premolar loci for species in the clade Placentalia is identified as p1/P1, p2/P2, p4/P4, and p5/P5 [traditional p1/P1, p2/P2, p3/P3, p4/P4]).
Repositories and institutional abbreviations
All newly described fossils are in the collections of the Paleontological Institute of the Mongolian Academy of Sciences, Ulaanbaatar, Mongolia. Casts of these specimens reside in the American Museum of Natural History, New York, New York, USA. Abbreviations are: American Museum of Natural History, New York, New York, USA (AMNH); Chernyshev's Central Museum of Geological Exploration Saint Petersburg, Russia (CCMGE); Inner Mongolian Museum, Hohhot, China (IMM); Institute of Vertebrate Paleontology and Paleoanthropology, Beijing, China (IVPP); Mongolian Academy-American Museum Paleontological Expedition (MAE); Mongolian Paleontological Center-Mammal Collection, Academy of Sciences of Mongolia, Ulaanbaatar, Mongolia (MPC-M); Museum of Comparative Zoology, Harvard University, Cambridge, Massachusetts, USA (MCZ); Museo de Historia Natural “Alcide d'Orbigny,” Cochabamba, Bolivia (MHNC); Paleontological Institute of the Russian Academy of Sciences (PIN); Paleontological Institute, Mongolian Academy of Sciences, Ulaanbaatar, Mongolia (PIN MAS, PSS-MAE); Paleozoological Institute of the Polish Academy of Sciences, Warsaw, Poland (Z. Pal.); Zoological Institute, Russian Academy of Sciences, Saint Petersburg (ZIN).
Systematic paleontology
Ostracoda Latreille, Reference Latreille1806
Podocopida Müller, Reference Müller1894
Cytheroidae Baird, Reference Baird1850
Limnocytheridae Klie, Reference Klie and Dahl1938
Genus Limnocythere Brady, Reference Brady1868
Type species
Limnocythere inopinata, Baird, Reference Baird1843.
Limnocythere nemegtensis Szczechura, Reference Szczechura1971
Figure 4.1
Type materials
Holotype: Z. Pal. No. MgO/25. Right valve. Paleogene, Naran Member (“White Beds”) of the Naran Bulak Formation, Gobi Desert, Mongolia.
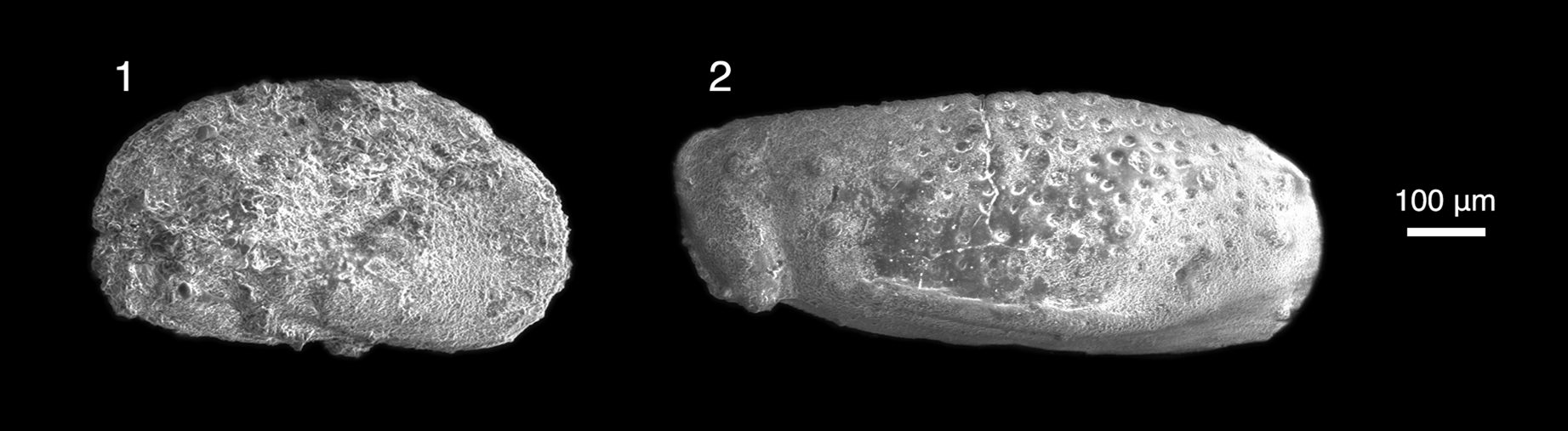
Figure 4. Freshwater Ostracoda from the locality Daus-1, collected in the Naran Member of the Naran Bulak Formation, western Gobi Desert, Mongolia. (1) Limnocythere nemegtensis Szczechura, Reference Szczechura1971 (N 5/69a PIN MAS). (2) Cyprideidae genus and species indet., (N 5/69b PIN MAS), left valve.
Diagnosis
“Carapace angularly-ovate in lateral outline, in front of the middle bearing a deep sulcus which is nodose anteriorly. Large, rib-like inflation occurs in anterior part of the carapace, extending adventrally below the sulcus turning upwards to disappear posteroventrally. Similar inflation runs below the posterodorsal margin. Valve surface strongly pitted” (Szczechura, Reference Szczechura1971, p. 92).
Occurrence
The type locality is in the Paleogene Naran Member (“White Beds”) of the Naran Bulak Formation, western Nemegt Basin, southwestern Mongolia at the localities Ulan Bulak, Tsagaan Khushu, and Naran Bulak (Szczechura, Reference Szczechura1971). The new specimen described here is also from the Naran Member, Naran Bulak Formation, but from the central-eastern Nemegt Basin locality Daus-1. The species has been reported from the late Paleocene Bayanulanian Asian Land Mammal Age of the Nomogen Formation, Erlian Basin, Nei Mongol, China (Wang et al., Reference Wang, Li, Bai, Jin, Mao and Meng2019) and from three additional Paleocene deposits: the Subeng section, Inner Mongolia (Nei Mongolia), China, and the Xhinzuang and Shashi formations of China (Van Itterbeeck et al., Reference Van Itterbeeck, Missiaen, Folie, Markevich, Van Damme, Gou and Smith2007).
Description
The specimen has the diagnostic features of the type as follows: carapace small, angularly ovate in lateral outline, dorsal margin almost straight, and ventral margin concave anteromedially and slightly rounded posteriorly. In the middle, at the dorsal margin, a transverse depression appears, extending to middle of carapace, and the depression expands to the dorsal margin. Conspicuous rib-like thickening extends along ventral margin of the carapace; and valve surface weakly pitted.
Materials
N 5/69a PIN MAS, one complete carapace. Collected in 2014.
Remarks
Brady (Reference Brady1868, p. 121) named the genus Limnocythere but did not provide a conventional species diagnosis writing only the species names “L. inopinata (Baird); monstrifica (Norman)” followed by notes on the morphology of their antennae. Meisch (Reference Meisch2000, p. 427) later specified that L. inopinata is the type species. Brady (Reference Brady1868) remarked that extant members of this genus were found in the clay of bottom substrates.
The Naran Member of the Naran Bulak Formation has yielded numerous ostracode fossils (Khand, Reference Khand1987). When describing Naran Bulak Formation ostracodes, Szczechura (Reference Szczechura1971, p. 87) wrote: “it is impossible to draw conclusions as to the age of the studied sediments on the basis of the ostracods.” Her basis for inferring a Paleocene age for Limnocythere nemegtensis was the context of geological work by Gradziński et al. (Reference Gradziński1969, p. 87), who stated that a Paleocene age could only be assigned “provisionally” to what is now considered the Naran Member of the Naran Bulak Formation. Szczechura (Reference Szczechura1971) wrote that the genus Limnocythere had been described as a Paleocene taxon citing “Morkhoven (Reference Van Morkhoven1963)” (apparently an erroneous citation of Van Morkhoven, 1962, because the titles are identical). We could not, however, corroborate this age in Van Morkhoven (Reference Van Morkhoven1963, v. II, p. 406–409) who described Limnocythere as Oligocene–Recent. Limnocythere nemegtensis appears to be endemic to Asia, known from both Mongolia and, more recently, from the Erlian Basin, Inner Mongolia, China, where the sediments yielding this ostracode have been dated using biostratigraphy, chronostratigraphy, and magnetostratigraphy as Paleocene Bayanulanian beds (Wang et al., Reference Wang, Li, Bai, Jin, Mao and Meng2019, p. 294–295). Limnocythere is a freshwater taxon (Van Morkhoven, Reference Van Morkhoven1963).
Cypridoidea Baird, Reference Baird1845
Cyprideidae Martin, Reference Martin1940
Genus and species indet.
Figure 4.2
Occurrence
Locality Daus-1, Paleogene, Naran Member (“White Beds”) of the Naran Bulak Formation, Gobi Desert, Mongolia.
Description
This taxon is known from a fragmentary valve that exhibits key morphological features distinguishing it from other taxa, including a rostrum-like process (beak adjacent to a groove and ridge) and a cyathus along the ventral margin of the left valve. The valve surface is strongly pitted.
Materials
N 5/69b PIN MAS left valve, fragment, lower part of right valve of ostracode. Collected in 2014.
Remarks
The morphological features noted above, as well as a distinct groove bordered by a ridge, and a cyathus are typical of the family Cyprideidae (Martin, Reference Martin1940, Reference Martin1958; Sames, Reference Sames2011). The family, particularly the genus Cypridea, is a common element in nonmarine late Mesozoic to early Cenozoic faunas nearly worldwide (Sames, Reference Sames2011). Despite having morphological features similar to those commonly described for Cypridea, N 5/69b PIN MAS is incomplete and damaged, making generic and specific assignment uncertain.
Mammalia Linnaeus, Reference Linnaeus1758
Eutheria Gill, Reference Gill1872
Placentalia Owen, Reference Owen1837
Pantodonta Cope, Reference Cope1873
Pantolambdoidea Cope, Reference Cope1883
Pantolambdodontidae Granger and Gregory, Reference Granger and Gregory1934
Archaeolambdidae Flerov, Reference Flerov1952
Genus Archaeolambda Flerov, Reference Flerov1952
Type species
Archaeolambda planicanina Flerov, Reference Flerov1952, p. 44, fig. 1, PIN 534-68, from “Paleocene of the Nemegt Basin, Gobi Desert,” Mongolia (Kielan-Jaworowska, Reference Kielan-Jaworowska1968, p. 135) and by original description.
Diagnosis
As given in Kielan-Jaworowska (Reference Kielan-Jaworowska1968, p. 135).
Archaeolambda cf. A. planicanina Flerov, Reference Flerov1952
Figure 5
Type materials
Holotype: PIN 534-68, a mandible with left canine–m3 and right p1–m3 from the Paleogene beds of the Nemegt Basin, Mongolia, by original description. Simons (Reference Simons1960) synonymized the genus Archaeolambda with the previously named Haplolambda; however, we follow Kielan-Jaworowska (Reference Kielan-Jaworowska1968, p. 133–134) who subsequently identified features distinguishing the genera.
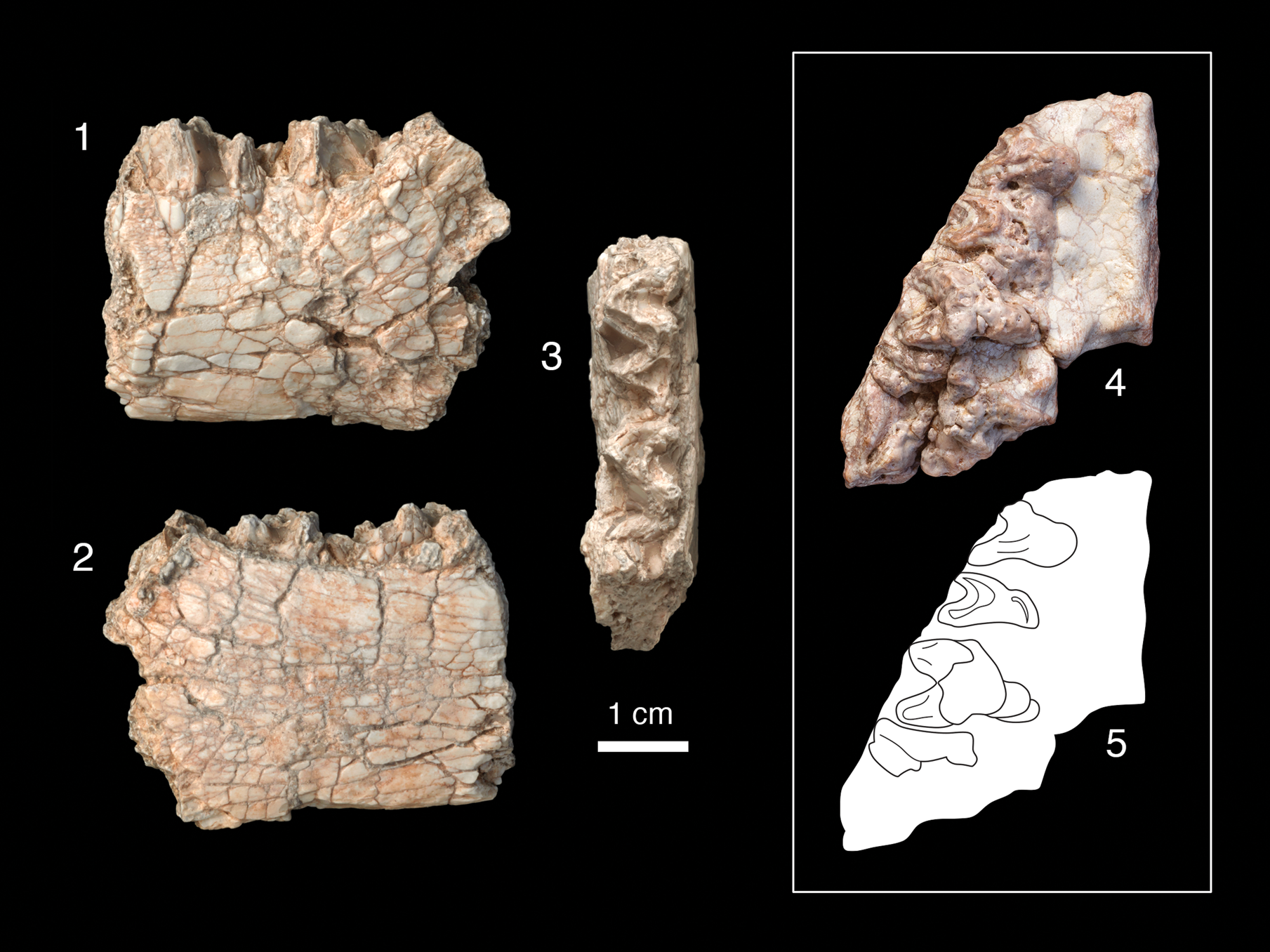
Figure 5. Dentitions of Archaeolambda cf. A. planicanina. Left dentary (PSS-MAE 662) with m2–m3 in (1) labial, (2) lingual, and (3) occlusal views; (4, 5) right maxilla (PSS-MAE 670) in occlusal view with drawing of the same showing P5–M2 and mesialmost labial root of M3.
Diagnosis
Specimens with the strongly crested, dilambdodont lower molars characteristic of Pantodonta, whereby the major cusps are connected by pronounced crests and trigonids are higher than talonids (Flerov, Reference Flerov1952; Simons, Reference Simons1960; Kielan-Jaworowska, Reference Kielan-Jaworowska1968). The lower dentition exhibits the diagnostic feature of the family Archaeolambdidae of “talonids in lower molars shorter and narrower than trigonids” (Kielan-Jaworowska, Reference Kielan-Jaworowska1968, p. 135). Additionally, the specimen exhibits the diagnostic genus-level features of low and narrow molar talonids (Simons, Reference Simons1960, p. 27).
Occurrence
Holotype from the Paleogene beds of the Nemegt Basin, Mongolia (Flerov, Reference Flerov1952; Kielan-Jaworowska, Reference Kielan-Jaworowska1968; Li and Ting, Reference Li and Ting1983); new lower dentition from locality Daus-1 and the new upper dentition from Daus-2b, both from the Paleogene Naran Member (“White Beds”) of the Naran Bulak Formation, central-eastern Nemegt Basin, Mongolia.
Description
Both specimens, particularly the upper dentition (PSS-MAE 670), exhibit extensive postmortem damage. Although the lower dentition (PSS-MAE 662) appears slightly larger, the sizes of the two specimens are similar enough that without better material (and because much of the comparative material consists of isolated lower and upper dentitions), the new specimens are tentatively referred to the same taxon.
The lower dentition consists of a left m2 and m3, with the base of the mandibular ramus preserved laterally adjacent to the talonid of m3. The m2 is fully intact but the m3 preserves only the trigonid. The new lower dentition is slightly larger than the holotype, which is a lower dentition of Archaeolambda planicanina figured in Flerov (Reference Flerov1952, fig. 2) and examined on a cast of the same (AMNH 56621). Differences include that the length of the m2 of the new specimen, PSS-MAE 662, is closer in size to the length of the m3 of the holotype. The dentary under the trigonid of m2 is slightly deeper in the new specimen (holotype = 2.0 cm vs. PSS-MAE 662 = 2.6 cm). The talonid and trigonid of m2 are more open in the new specimen than they are in the cast of the holotype (AMNH 56621), the latter feature not being well represented, as illustrated in Flerov (Reference Flerov1952, fig. 2).
On the upper dentition, postmortem weathering has exposed the tooth roots and removed the cusps, which eliminated many details necessary for synapomorphy-based taxonomic assignments and for assessment of full tooth dimensions. We tentatively also refer this maxilla to Archaeolambda cf. A. planicanina based on the tooth outline shape and overall dimensions of the teeth. On what we recognize as M2, the tooth outline forms an acute triangle, suggesting the full tooth was limited to a paracone, metacone and protocone, also suggested by the pattern that the premolars and molars are each separated by a complementarily acute triangular space. The P5–M2 (traditional P4–M2) each had three roots (one lingual and two labial) and, judging by the position of the lingual root, the mesial-most two teeth are distinctly narrower in the labial-lingual dimension than either of the more distal teeth. This morphology and the overall tooth size have broad similarity to Archaeolambda planicanina (PIN 534-68, a P4-M1) figured in Flerov (Reference Flerov1952, fig. 1) and examined on cast AMNH 56618, which shares two loci (P5 [traditional P4] and M1) in common with the new specimen, given our hypothesis of homology. Additionally, pairs of exposed labial roots of the M1–M2 and a small part of the mesial-most root of M3 are visible and the lingual edge of the protocone of M2 is convex. The M2 of the new specimen forms a very acute angle lingually, more so than does the M2 of A. planicanina described by Kielan-Jaworowska (Reference Kielan-Jaworowska1968). In this feature, the new M2 also resembles the holotype of A. tabiensis Huang, Reference Huang1977 (IVPP 4333 and cast AMNH 126866). However, because of the poor preservation of the new specimen, no synapomorphic features can be identified with A. tabiensis and we do not refer it to that taxon here.
The morphology of the maxilla has also informed our tooth loci homologies. The well-preserved palatine process of the right maxilla has posterior and medial edges that are intact. The distal edge exhibits a bony protrusion, similar to that reported by Kielan-Jaworowska (Reference Kielan-Jaworowska1968, fig. 1) for Archaeolambda cf. A. planicanina (Z. Pal. No. MgM-I1/54) directly lingual to M2. Neither sutures nor greater and lesser palatine foramina are preserved on the oral surface of the palatine process, but two small foramina can be seen on the posterior end of the palate.
Materials
PSS-MAE 662, a poorly preserved left dentary with m2–m3, collected in 2017 from Daus-1, and PSS-MAE 670, a poorly preserved right maxilla with ?P5–M2 (traditional P4–M2) and the mesial-most labial root of M3, collected in 2019 from Daus-2b. These were compared with specimens AMNH 56618 and 56621, casts of the holotype, with an additional specimen of Archaeolambda planicanina (Flerov, Reference Flerov1952), with A. tabiensis, AMNH 126866 (cast of IVPP 4333), and with the literature (Flerov, Reference Flerov1952; Kielan-Jaworowska, Reference Kielan-Jaworowska1968).
Measurements
PSS-MAE 662, m2: maximum mesiodistal length (ML) = 16.5 mm; maximum labiolingual width (MW) = 11.7 mm; trigonid width = 11.7 mm; talonid width = 8.7 mm; m3: maximum length (ML) = 12.1 mm; maximum width (MW) = 9.3 mm. PSS-MAE 670, right P5: maximum length (ML) = 8.5 mm; maximum width (MW) = 10 mm (est.); right M2, maximum length (ML) = 10.8 mm, maximum width (MW) =12.8 mm.
Remarks
Mandibles from a Naran Bulak locality in the Nemegt Basin described by Flerov (Reference Flerov1952) were attributed to a new family, genus, and species: Archaeolambdidae and Archaeolambda planicanina. Simons (Reference Simons1960) collapsed this family into the Barylambdidae and the genus into Haplolambda. These synonymies were not, however, followed by Kielan-Jaworowska (Reference Kielan-Jaworowska1968) who presented additional maxillary material of Archaeolambda cf. A. planicanina and identified features of the maxillary dentition that supported retaining the taxonomy of Flerov (Reference Flerov1952). Because the material we present is inadequate to resolve taxonomic debates, we follow this more recent taxonomic interpretation.
Archaeolambda, which is not known outside Asia, has been extensively reported from the Naran Member (“White Beds”) of the Naran Bulak Formation as a taxon of the Gashatan or Gashatan-equivalent ALMA or older (Li and Ting, Reference Li and Ting1983; Dashzeveg, Reference Dashzeveg1988; Dashzeveg and Russell, Reference Dashzeveg and Russell1988; Lucas, Reference Lucas, Aubry, Lucas and Berggren1998; Ting, Reference Ting1998; Wang et al., Reference Wang, Hu, Zhou and Li1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002). It is important to note that Dashzeveg (Reference Dashzeveg1988) also reported Archaeolambda from the Bumban Member of the Naran Bulak Formation, thus suggesting that the taxon has an Eocene range in the Bumbanian ALMA, potentially complicating its stratigraphic usefulness. In a recent review, Lopatin (Reference Lopatin2020) distinguished A. planicanina as a Paleocene Gashatan taxon, versus A. bogdensis (Dashzveg, Reference Dashzeveg1980) as a middle Eocene Arshantan taxon, thus species-level discrimination may be necessary for Archaeolambda to be a useful index taxon (see also Dashzeveg, Reference Dashzeveg1982).
Genus Pantolambdodon Granger and Gregory, Reference Granger and Gregory1934
Type species
Pantolambdodon inermis Granger and Gregory, Reference Granger and Gregory1934.
cf. Pantolambdodon sp. indet.
Figure 6
Occurrence
The new specimen is from locality Daus-2b, Naran Member, Naran Bulak Formation and was collected in 2019. Previously described specimens come from the Bayan Ulan locality, Inner Mongolia, China (Paepen et al., Reference Paepen, Hong, Yan and Smith2021), and previously undescribed specimens come from the Ulan Shireh beds, Mongolia.

Figure 6. Tarsals of Pantolambdodon sp. (1–5) PSS-MAE 671, a newly discovered right astragalus from the Naran Member of the Naran Bulak Formation, Mongolia, and (6), AMNH 147008, a right astragalus and calcaneus (not associated) collected in 1925 from the Ulan Shireh beds, Mongolia. Photographs with corresponding drawings beneath. Views are (1) dorsal; (2) plantar; (3) medial; (4) distal; (5) proximal; and (6) medial view of astragalus and calcaneus in articulation. c, calcaneus; ca, calcaneal facet; cl, capitular lip; cu, cuboid facet; ect, ectal facet; f, fibular facet; h, head; mmf, facet for the medial malleolus of tibia; mtf, medial tibial facet; n, neck; nav, navicular facet; nh, facet for the navicular hook; st, sustentacular facet; sus, sustentaculum tali; and t, trochlea.
Description
PSS-MAE 671 is a weathered right astragalus with exposed trabecular structure and a damaged ectal facet. The trochlea is relatively broad mediolaterally, comparable in width to the astragalar neck, although the lateral edge of the trochlea is weathered, potentially distorting the comparison. The trochlea is also gently grooved with edges that are not strongly skewed; the medial edge is directed ~10° from a parasagittal plane. Medial and lateral edges converge somewhat distally. The articular surface of the trochlea does not extend onto the neck. The medial side of the trochlea is flattened for articulation of the medial (tibial) malleolus and there is no medial fossa. What remains of the fibular facet on the lateral side is also flat. The astragalar foramen is absent. In posterior view, the medially flaring nature of the ectal facet is apparent. The neck of the astragalus, which is relatively aligned with the trochlea, is short, comparable in length to the length of the trochlea.
The head of the astragalus extends medially from the neck and articulates with the navicular, cuboid, and a third bone described below. The navicular facet occupies approximately one half of the size of the head; the outline of the facet is broadly oval, with the long axis running from dorsal and lateral to plantar and medial. The surface of the navicular facet is convex dorsally, transitioning to concave plantad. A distinct lip marks the plantar extreme of the navicular facet. This distinctive structure we term here the capitular lip of the astragalus. The cuboid facet occupies approximately one quarter of the astragalar head. Despite damage, it is apparent that this facet was convex, relatively broad dorsally, and that it tapered to a point at the plantar extreme.
On the medial side of the astragalar head is an additional, relatively large facet occupying approximately one fourth of the head, which appears to have been for a bone in the tarsus known as the “navicular hook” or the “tibiale” (Matthew, Reference Matthew1937, p. 165, 179), or the “medial tarsal bone” (Hildebrand, Reference Hildebrand1978). Although this Daus astragalus is weathered, it is clear that the orientation of the facet for the navicular hook is almost perpendicular to the navicular facet. The facet for the navicular hook is convex and connects to the navicular facet by a smaller, flatter facet that is weathered.
On the plantar surface, the sustentactular facet is an elongate oval that does not contact the astragalar head. The ectal facet is gently concave and poorly preserved at its proximal and lateral extremes. Part of the cuboid facet extends onto the plantar surface where it likely contacted the calcaneus. Plantar view also reveals the facet for the navicular hook meeting the facet for the navicular on the astragalar head.
Materials
We made direct comparisons of PSS-MAE 671 to the following: AMNH 147008, a batch-catalogued sample of tarsals that are not associated, including eight astragali, attributed to ?Pantolambdodon from Eocene Irdin Manha beds of Inner Mongolia, China; AMNH 16663, Pantolambda bathmodon Cope, Reference Cope1882; AMNH 81715, a collection of multiple astragali (not associated) of the perissodactyl Lophialetes expeditus Matthew and Granger, Reference Matthew and Granger1925; and AMNH 5021, the mesonychian Mesonyx obtusidens Cope, Reference Cope1871. Additional comparisons were made in the literature to the astragalus Pantolambdodon sp. (IMM-1995-BAYU-070; Paepen et al., Reference Paepen, Hong, Yan and Smith2021) and to Alcidedorbignya inopinata (Baird, Reference Baird1843) (MHNC 8372; de Muizon et al., Reference de Muizon, Billet, Argot, Ladevèze and Goussard2015).
Measurements
Maximum length of the astragalus = 33.6 mm; maximum width of the trochlea = 20.3 mm; maximum width of the head = 20.9 mm. Overall maximum width inestimable due to postmortem damage.
Remarks
The new astragalus, PSS-MAE 671, compares closely with the astragalus recently described and figured as Pantolamdodon sp. from the late early-middle Eocene deposits of Bayan Ulan, Inner Mongolia, China (Paepen et al., Reference Paepen, Hong, Yan and Smith2021, fig. 4F:1–6). Importantly, those authors posited a faunal association between dental and tarsal specimens of Pantolambdodon sp. that assisted us in the identification of this isolated astragalus. Additionally, AMNH 147008, an unpublished, batch-catalogued collection of tarsals that includes eight astragali and a calcaneus collected by the 1925 Central Asiatic Expedition from the Eocene Irdin Manha beds of Inner Mongolia, China, and cataloged as ?Pantolambdodon (collection note, E. Manning) also appears to represent the same taxon. The bones cataloged under AMNH 147008 are much less weathered than PSS-MAE 671 and served as an important comparative basis for identifying articular facets on the new specimen.
The calcaneus of AMNH 147008, although not necessarily associated with any of the astragali batch catalogued under the same number, was placed in approximate articulation with one of them. This exercise revealed that the third facet on the astragalar head did not articulate with the calcaneus and was likely for an absent bone, the navicular hook. Such a bone, which was reported in Pantolambda bathmodon from Paleocene (Torrejonian) rocks (Matthew, Reference Matthew1937), was described as articulating with both the astragalus and the body of the navicular. This additional tarsal also has been described in rodents as a medial tarsal bone serving as the site of attachment for the tibialis posterior muscle (Hildebrand, Reference Hildebrand1978). A similar facet was figured in the Paleocene pantodont from Bolivia Alcidedorbignya inopinata and termed the supplementary astragalocalcaneal facet (de Muizon et al., Reference de Muizon, Billet, Argot, Ladevèze and Goussard2015, fig. 103; erroneously labeled as on the lateral side of the astragalus in fig. 102C.). However, as noted above in Pantolambdodon sp., it does not appear that this facet contacted the calcaneus. In both the Daus specimen PSS-MAE 671 and the Pantolambdodon specimens of AMNH 147008, the facet for the navicular hook has a small, distal extension that contacts the facet for the navicular on the head of the astragalus at the capitular lip.
Features in common among all of the specimens mentioned that are referred to Pantolambdodon sp. are similar overall size, relative shallowness of the trochlea, presence and proportions of cuboid and navicular facets on the astragalar head, presence of a facet for the navicular hook, and lack of an astragalar foramen. They also share a distinctive crest on the plantar aspect of the navicular facet on the astragalar head (visible in Paepen et al., Reference Paepen, Hong, Yan and Smith2021, fig. 4F6), which we call the capitular lip of the astragalus and hypothesize may be a synapomorphy of Pantolambdodon sp. and close relatives. The capitular lip is absent in other taxa that have both a cuboid and a navicular facet on the astragalar head such as Mesonyx obtusidens (AMNH 5021; see also O'Leary and Rose, Reference O'Leary and Rose1995) and is absent in other pantodonts such as Pantolambda bathmodon (AMNH 16663) and Alcidedorbignya inopinata (de Muizon et al., Reference de Muizon, Billet, Argot, Ladevèze and Goussard2015, fig. 102). There is, however, a capitular lip on the astragalus of perissodactyls such as Lophialetes expeditus (AMNH 81715) and Hyracotherium (Kitts, Reference Kitts1956). Pantodonta has been considered part of the clade Ferae (McKenna and Bell, Reference McKenna and Bell1997; Paepen et al., Reference Paepen, Hong, Yan and Smith2021), and thus not particularly closely related to Perissodactyla. Thus, this shared resemblance is likely to be a homoplasy.
A difference between PSS-MAE 671 and both the AMNH specimens and the Paepen et al. (Reference Paepen, Hong, Yan and Smith2021) specimen is that in PSS-MAE 671 the trochlea is more symmetrical. PSS-MAE 671 is also ~25% smaller than most of the other specimens (although one of the AMNH 147008 astragali is of comparable size), and appears to have had a slightly more elongate neck.
Pantolambdodon has been reported previously as an Eocene taxon from Arshatan and Irdinmanhan beds of Asia (Li et al., Reference Li, Wang and Fostowicz-Frelik2016; Paepen et al., Reference Paepen, Hong, Yan and Smith2021).
Glires Linnaeus, Reference Linnaeus1758
Mimotonidae Li, Reference Li1977
Gomphos Shevyreva, Chkhikvadze, and Zhegallo, Reference Shevyreva, Chkhikvadze and Zhegallo1975
Type species
Gomphos elkema Shevyreva, Chkhikvadze, and Zhegallo, Reference Shevyreva, Chkhikvadze and Zhegallo1975.
Other species
Gomphos shevyrevae Meng et al., Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009, and Gomphos ellae Kraatz, Badamgarav, and Bibi, Reference Kraatz, Badamgarav and Bibi2009.
Diagnosis
In part from Meng et al. (Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004): similar to other mimotonids but differing from all other Glires in having two lower incisors. Differing from Mimotona in having a well-separated paracone and metacone and a distinct mesostyle on the upper molars. Differing from Anatomylus in having a shallow horizontal ramus (Averianov, Reference Averianov1994). Differing from Mimolagus in that Gomphos is of smaller size and has taller cheek teeth crowns with sharper cusps and ridges (Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004).
Gomphos cf. G. elkema Shevyreva, Chkhikvadze, and Zhegallo, Reference Shevyreva, Chkhikvadze and Zhegallo1975
Figures 7, 8
Occurrence
The new specimen, collected in 2019, comes from locality Daus-2b, Naran Member (“White Beds”) of the Naran Bulak Formation, Paleogene of Mongolia. Gomphos elkema has been described from the Bumban Member of the Naran Bulak Formation in the region of Tsagan-Khushu, Nemegt Basin, Mongolia, beds that are considered lower Eocene (Shevyreva et al., Reference Shevyreva, Chkhikvadze and Zhegallo1975; Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004, p. 2), as well as the Gashato Formation, Ulan-Nur Basin of Mongolia (Dashzeveg, Reference Dashzeveg1988; Dashzeveg and Russell, Reference Dashzeveg and Russell1988; Asher et al., Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005). Gomphos elkema also has been reported from the Huheboerhe locality, Erlian Basin, Nei-Mongol (Inner Mongolia), where it has been used as an index taxon to identify the presence of Bumbanian-equivalent beds and the presence of a possible Paleocene-Eocene boundary section (Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004).

Figure 7. Skull of Gomphos cf. G. elkema (PSS-MAE 669). (1, 2) Left-lateral view and parasagittal CT scan slice showing left dI2 and I3 roots (dotted lines indicate distal ends of root; black line indicates maxillopalatine suture; note that the view of the dI2 root in 1 is shown from a slightly different position than it is in the CT slice in 2), and roots and crowns of P2, P4, P5, M1, M2, and M3; (3, 4) right-lateral view with the following teeth visible: dI2, I3, P2, P4, and P5; photograph and drawing; (5, 6) dorsal view, photograph and drawing; (7, 8) ventral view, photograph and drawing showing dI2 and I3 along with P2 (right side tooth; left side root), P4 and P5 and M1–M3. av, anteroventral zygomatic process of the maxilla; et; ethmoturbinal; fp sut; frontoparietal suture; fr, frontal; iof, infraorbital foramen; j, jugal; jm sut, jugomaxillary suture; mp sut, maxillopremaxillary suture; mx, maxilla; n, nasal; or, orbit; pa, piriform aperture; pmx, premaxilla; pp, posterior process of the premaxilla; sc, sagittal crest; sof, supraorbital foramina; ss, sagittal suture; and z, zygomatic process of the maxilla.
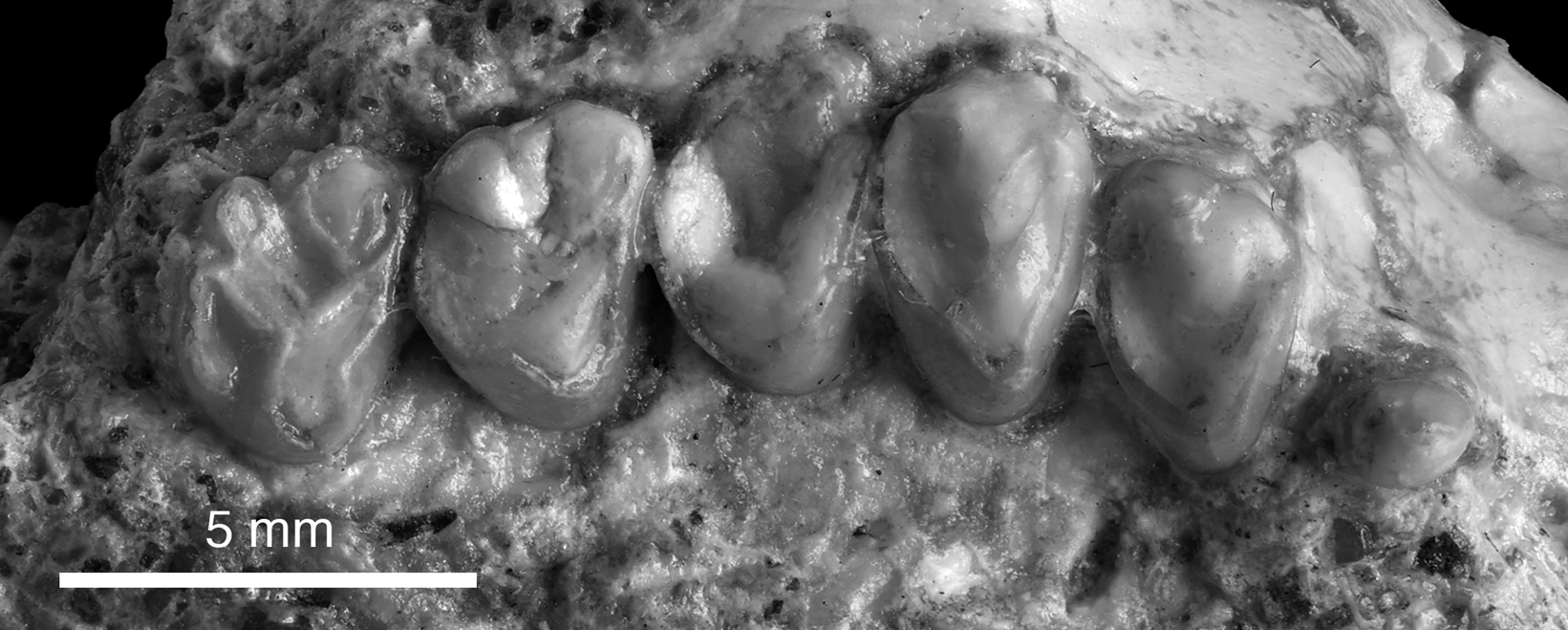
Figure 8. Right upper post-incisor dentition of Gomphos cf. G. elkema (PSS-MAE 669) P2, P4, and P5, M1–M3.
Description
PSS-MAE 669 preserves a nearly complete, somewhat transversely compressed rostrum and a partial portion of the orbit and skull roof. The premaxilla is extensive and encloses the elongated curved roots of the upper incisors (homologized here as dI2 and I3, see comments below). The maxillopremaxillary suture is clearly demarcated on the right lateral side (but not on the left side) of the rostrum. This suture is finely interdigitated, which is a feature described in the gliroid Rhombomylus as characteristic of more mature individuals (Meng et al., Reference Meng, Hu and Li2003). A conspicuous posterior process of the premaxilla (Meng et al., Reference Meng, Hu and Li2003) flanks the nasal on the roof of the rostrum. The posterior terminus of this process is obscured by damage. Asher et al. (Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005) specified a narrow anterior process of the frontal bone inserting between the premaxilla and the maxilla as a feature of Gomphos elkema, but this region is not preserved in the Daus specimen. The Daus specimen, however, does have a narrow anteriorly and broad posteriorly, V-shaped projection of the frontal bone between the nasals, which Asher et al. (Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005) described as shared with leporids.
On the palate, the maxillopremaxillary suture is located in the large diastema between the distal upper incisor and P2. This intersection is well mesial to P2. In CT scans the position of the maxillopremaxillary suture relative to the tooth roots can be observed. The premaxilla encases most of the roots of the upper incisors. The root of dI2, but not that of I3, extends for a short distance into the maxilla. On the palate in the midline, the maxillopremaxillary suture is obscured by matrix. A CT scan reveals symmetrical scrolls of bone inside the posterior nasal cavity; these are interpreted as ethmoturbinals.
The roof of the nasal cavity is partially intact, and the nasal bones are prominent elements of the facial skeleton. At the front of the snout the nasals are broken such that their lateral edges are poorly defined. Two oval openings, one in each nasal bone, are present near the midline. These openings may be foramina or products of breakage. The nasals project distinctly over the piriform aperture. The finely interdigitated frontonasal sutures indicate that the nasals extend posteriorly on the skull roof to the anterior margin of the orbit. The supraorbital foramina are minute, oval openings on the anterior aspect of the frontal, that are best preserved on the right side.
The maxilla in PSS-MAE 669 preserves the facial process, parts of the palatine, and parts of the zygomatic processes; the orbital process is obscured by matrix. The infraorbital foramen is visible on both sides, but is particularly clear on the right maxilla, where it opens above P2. A shallow trough extends forward several millimeters anteriorly from the foramen, but there are no other evident scars or muscular depressions on the face indicative of attachments for snout muscles. The zygomatic process of the maxilla is well developed and convex anteriorly. On the right side there is a partially preserved portion of the anteroventral zygomatic process of the maxilla as seen in Rhombomylus (Meng et al., Reference Meng, Hu and Li2003). A distinct jugomaxillary suture visible on the right side indicates that the jugal makes a substantial contribution to the anterior part of the zygoma. In ventral view, the root of the zygomatic process of the maxilla lies lateral to P5 and M1. The palatomaxillary suture is not visible and it is not clear whether it was absent or obscured by poor preservation. In PSS-MAE 669, a fragment of bone at the posterior aspect of the right orbit preserves the anterior part of the sagittal crest and the highly interdigitated suture between the right frontal and parietal. The sagittal crest is a low, blunt ridge.
Two enlarged upper incisors in the Daus specimen are preserved on the right side of the skull. Based on ontogenetic studies in rodents and lagomorphs (Luckett, Reference Luckett, Luckett and Hartenberger1985), the anterior incisor in Gomphos and other Glires has been homologized as dI2 (Li et al., Reference Li, Wilson, Dawson, Krishtalka and Genoways1987; Luckett and Hartenberger, Reference Luckett and Hartenberger1993; Meng et al., Reference Meng, Hu and Li2003). Asher et al. (Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005) observed that Gomphos resembles extant lagomorphs in having two pairs of upper incisors, whereas this more distal incisor is absent in many early Glires such as Rhombomylus (Meng et al., Reference Meng, Hu and Li2003) and rodents. The observation that the tooth at this locus undergoes replacement is the basis for its identification as I3 in adult lagomorph skulls (Luckett, Reference Luckett, Luckett and Hartenberger1985), a state shared with Gomphus (Asher et al., Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005, fig. S10). The left incisor crowns are not preserved. The left premaxilla has been sheared off laterally, exposing the pulp cavities of the broken left incisor roots. As a result, the pulp cavities are filled with matrix, which improved their contrast and visibility in the CT scan. As noted above, dI2 is a robust tooth with a root that arcs through the premaxilla into the anterior maxilla where it terminates in a crypt. The right dI2 crown is largely missing, apparently worn down through use in life. There is an indication of enamel on its labial and mesial surfaces. dI2 is oval in occlusal outline. I3 is much smaller than dI2 (~25% of the area of dI2). The extent of enamel on the surface of I3 is obscured.
In PSS-MAE 669, the post-incisor dentition shows considerable wear and damage, but the crown features of the cheek teeth are preserved well enough on the right side of the skull to show diagnostic cusps, connecting ridges, and wear facets. The right P2 has a very slightly bicuspid crown in lingual profile. A shallow lingual wear facet extends posteriorly from the apex of the crown. The tooth has a broadly circular outline in occlusal view.
P4 (P3 of Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004) is oval in outline in occlusal view. Its lingual portion shows extensive wear and the protocone is preserved as a flattened ridge with continuous, well-worn mesial and distal lophs that extend labially. On the labial margin of the crown, a prominent, conical centrolabial cusp lies mesial and slightly lingual to an additional, less robust, distolabial cusp. A shallow triangular basin lies between the centrolabial cusp and the protocone. Deep within this basin is a low ridge connected to the mesial loph, and the apex of this ridge forms a weak conule. The hypocone and lingual and labial cingula are absent.
P5 (P4 of Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004) is oval in occlusal outline, but more transversely elongate than is P4. The apex of the protocone shows a distinct circular wear facet from which mesial and distal lophs extend. The centrolabial cusp is robust and a distolabial cusp is hardly discernable. A well-excavated, arcuate basin extends from the distolabial corner of the crown to the apex of the protocone; the basin is not evident on the anterior side of the tooth. As in P4, P5 lacks a hypocone and lingual and labial cingula.
The upper molars are more rectangular and less labiolingually transverse in occlusal view than P4 and P5. The right M1 is considerably worn, but its lingual profile suggests the presence of a hypocone in addition to a protocone. A prominent, swollen mesial loph extends from the protocone to the mesiolingual base of what was likely the paracone; only the cusp is broken. The posterior protocone loph is more crest-like and extends to the distolingual base of the metacone. The metacone is not as prominent as the paracone, but both cusps are considerably worn. The metacone is slightly more lingual than the paracone, giving the labial margin of the crown an oblique outline that trends mesiolabial to distolingual. The most distinctive feature of M1 is a broad and deeply excavated trigon basin rimmed by the surrounding cusps and lophs. A metaconule is either absent or obscured by wear. The stylar region of the tooth is poorly preserved.
M2 is similar to M1 in its occlusal outline and many other features. M2 also differs from M1 in lacking a deeply excavated trigon basin. M2 has a distinctly robust paracone and metacone. The presence of a mesostyle is ambiguous on M1 due to wear, however, the mesostyle is clearly present on M2. A narrow trough extends from the labial margin of the crown between the paracone and mesostyle to the center of the shallow trigon basin.
M3 shows well-defined wear facets at the apices of the paracone and metacone that extend lingually to become confluent with the wear facets of the protocone and its lophs. A distinctive feature of M3 is a small, but well-defined hypocone separated from the distal face of the protocone by a narrow trough. The worn apex of the hypocone is confluent with a postcingulum that extends to a crest descending from the metacone. Accordingly, the hypoconal postcingulum is clearly distinct from the higher distal loph of the protocone.
Materials
PSS-MAE 669, an anterior skull preserving right and left maxillae, nasals, left anterior jugal, anterior infraorbital foramina, base of crowns of right dI2, I3, anterior roots of left dI2–I3, right P2, alveolus of left P2, right and left P4–5, M1–3. Comparisons were made with published illustrations as follows: upper teeth of Gomphos elkema, P3–M1 (IVPP-V13509.5), P5 (traditional P4)–M1 (IVPP-VI13509.6), P5 (traditional P4)–M2 (IVPP VI1509.4) from Meng et al. (Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004, figs. 3, 4); upper cheek teeth of Gomphos shevyrevae, V14669, V14671.1–V14671.4 from Meng et al. (Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009, fig. 2); left upper dentition of Mimotona wana Li, Reference Li1977, with P4 (traditional P3)–M3, V4324 in Li (Reference Li1977, fig. 2, pl. II 1a, b); upper teeth of Gomphos ellae, maxilla with M1–2 MPC 30/1 in Kraatz et al. (Reference Kraatz, Badamgarav and Bibi2009, fig. 4); and upper teeth of Anatomylus rozhdestvenskii Averianov, 1974, labial fragment of right M1 (ZIN 9161) in Averianov (Reference Averianov1994, fig. 4 K, L) (the right M1 is denoted as a left M1 in the text [Averianov, Reference Averianov1994, p. 404], but appears to be a right M1 in fig. 4] and right M3 (ZIN 9162, Averianov, Reference Averianov1994, p. 404; not illustrated).
Measurements
PSS-MAE 669, all measurements of right dentition. P4 (P3 of Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004), maximum mesiodistal length (ML) = 2.5 mm, maximum labiolingual width (MW) = 4.2 mm; P5 (P4 of Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004), ML = 2.7 mm, MW (est.) = 5.0 mm; M1, ML = 2.9 mm, MW = 4.8 mm; M2, ML = 2.8 mm, MW = 4.2 mm; M3, ML = 3.2 mm, MW = 3.6 mm.
Remarks
PSS-MAE 669 from Daus-2b closely resembles specimens of Gomphos elkema, a taxon named by Shevyreva et al. (Reference Shevyreva, Chkhikvadze and Zhegallo1975) and described extensively based on new material by Meng et al. (Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004). The species is known to occur in the upper beds of the “Nomogen Formation,” the Bumban Member of the Naran Bulak Formation, and members II and III of the Gashato Formation (Dashzeveg, Reference Dashzeveg1988; Dashzeveg and Russell, Reference Dashzeveg and Russell1988). Faunas from the Bumban Member generally have been recognized as early Eocene (Dashzeveg, Reference Dashzeveg1982, Reference Dashzeveg1988; Dashzeveg and Russell, Reference Dashzeveg and Russell1988; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002; Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004), although some authors have considered them to be late Paleocene (Beard, Reference Beard1998). Some features of the skull, dentition, and postcranial skeleton of G. elkema were briefly described by Asher et al. (Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005), who also published a phylogenetic analysis of the taxon and other selected Glires, non-gliroid eutherians, and marsupials. A few features of the skull are described here for the first time.
Other species in the genus also have been assigned to the Eocene. A new species, Gomphos shevyrevae, was named and described from the middle Eocene lower beds of the Irdin Manha Formation in the Erlian Basin (Meng et al., Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009). A third species, G. ellae, was described from the middle Eocene Kholboldchi Formation at Tsagan Khutal, Mongolia (Kraatz et al., Reference Kraatz, Badamgarav and Bibi2009). Gomphos ellae has been recognized as one of the youngest known mimotonids and the youngest species of Gomphos (Meng et al., Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009). In addition, a mimotonid genus, Anatomylus, was described from the locality Andarak 2 in the lower part of the Alay beds, Kyrgyzstan, and proposed to have close similarities to Gomphos (Averianov, Reference Averianov1994). The Alay beds are considered early or middle Eocene (Averianov, Reference Averianov1994).
Comparisons between the Daus specimen, PSS-MAE 669, and the above listed taxa, underscore its close similarity and favored assignment to Gomphos elkema. Gomphos shevyrevae has upper molars with more inflated and posteriorly expanded hypocones and lingual cingula. The widths of the cheek teeth of PSS-MAE 669 are more closely similar to those of G. elkema than to G. shevyrevae (compare measurements herein with Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004, table 1; Meng et al., Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009, table 1). Gomphos shevyrevae was diagnosed as having more robust teeth than G. elkema, but the measurements of the upper teeth of the latter reported by Meng et al. (Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004) are actually greater than those given for G. shevyrevae (Meng et al., Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009).
The third species of Gomphos, G. ellae, is represented by a single specimen, a lower jaw with a nearly complete dentition and an associated maxillary fragment with a heavily worn M1 and M2 (Kraatz et al., Reference Kraatz, Badamgarav and Bibi2009). Gomphos ellae was only distinguished diagnostically from G. elkema by its deeper ramus, and distinguished from all other mimotonids, including other species of Gomphos, in having a proportionately longer diastema. Unlike the Daus specimen and G. elkema, the M1 and M2 referred to G. ellae do not show evidence of a well-developed hypocone, but upper molars in G. ellae are so heavily worn that this distinction is tentative.
Anatomylus is another mimotonid taxon primarily distinguished by its short, robust, and deep jaw (see diagnosis above and Averianov, Reference Averianov1994). Only an incomplete M1 and a badly worn M3 in A. rozhdestvenskii can be compared with the Daus specimen. In contrast to the latter and Gomphos elkema, the M1 mesostyle in A. rozhdestvenskii is minute and the large postcingulum of M3 occupies almost half the crown.
Some gliroids (e.g., Rhombomylus) have well-documented skull morphology (Meng et al., Reference Meng, Hu and Li2003). Notwithstanding the brief description of Gomphos in Asher et al. (Reference Asher, Meng, Wible, McKenna, Rougier, Dashzeveg and Novacek2005), there has not been a detailed monographic treatment of the cranial osteology of Gomphos. One emerging difference between Gomphos and Rhombomylus that invites further examination is the morphology of the nasal bones, which may be pierced by foramina in Gomphos but are not in Rhombomylus (Meng et al., Reference Meng, Hu and Li2003, fig. 23).
All previously described taxa of Gomphos are thus from Bumbanian-aged faunas generally assigned to the early Eocene or from younger faunas in Mongolia and the Erlian Basin of China that are assigned to the middle Eocene (Shevyreva et al., Reference Shevyreva, Chkhikvadze and Zhegallo1975; Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004, Reference Meng, Wyss, Hu, Wang, Bowen and Koch2005, Reference Meng, Kraatz, Wang, Ni, Gebo and Beard2009; Kraatz et al., Reference Kraatz, Badamgarav and Bibi2009). The only mimotonid of the earlier Gashatan ALMA is the genus Mimotona (Li, Reference Li1977; Dashzeveg and Russell, Reference Dashzeveg and Russell1988). The Daus specimen, however, is clearly referable to Gomphos, and not Mimotona, and more specifically to G. elkema, in having less-transverse cheek teeth, a clearly separated paracone and metacone, and a distinct mesostyle on the upper molars.
Arctostylopida Cifelli, Schaff, and McKenna, Reference Cifelli, Schaff and McKenna1989
Arctostylopidae Schlosser, Reference Schlosser, Broili, Schlosser and von Zittel1923 Genus and species indet.
Figures 9–12
Occurrence
Locality Daus-2b, Naran Member (“White Beds”) of the Naran Bulak Formation, Mongolia. Collected in 2019.

Figure 9. Skull of an arctostylopid, genus and species indet (PSS-MAE 673) in lateral view. (1) Digital photograph; (2) CT rendering; and (3) composite drawing. fr, frontal; frn sut, frontonasal suture (nasals not preserved); iof, infraorbital foramen; or, orbit; pa, piriform aperture (opening preserved in part); pmx, premaxilla, and tooth loci (I3, C, P1–P5, M1–M3) are indicated. In the CT rendering, roots of some adjacent premolars and molars overlap in lateral view. Note the absence of a diastema. Striped areas are not preserved.

Figure 10. Skull of an arctostylopid, genus and species indet (PSS-MAE 673) (1–3) in dorsal views: digital photograph, CT rendering, and composite drawing; and (4–6) in ventral views: digital photograph, CT rendering, and composite drawing; tooth loci (I3, C, P1, P2, P4, P5, M1–M3) indicated on (6). bpc, basipharyngeal canal; ch, choanae; fr, frontal; frn sut, frontonasal suture; if, incisive foramen or foramina (may be one or two foramina); or, orbit; pa, piriform aperture (nasal bones that would have been at its superior aspect are not preserved); pal, hard palate; poc, postorbital constriction; ppt, postpalatine torus; pt, pterygoid process.

Figure 11. CT renderings and drawings of the skull of an arctostylopid genus and species indet (PSS-MAE 673) in (1, 2) anterior view and (3, 4) posterior view; tooth loci (I3, P5, M2, M3, M3 root) indicated on (4). bpc, basipharyngeal canal; bs, basisphenoid; ec, endocranial cavity; fr, frontal; frn sut, frontonasal suture; mx, maxilla; or, orbit (dimensions of orbit are approximated); pa, piriform aperture; pal, hard palate; pt, pterygoid process; ss, sphenoid sinus.
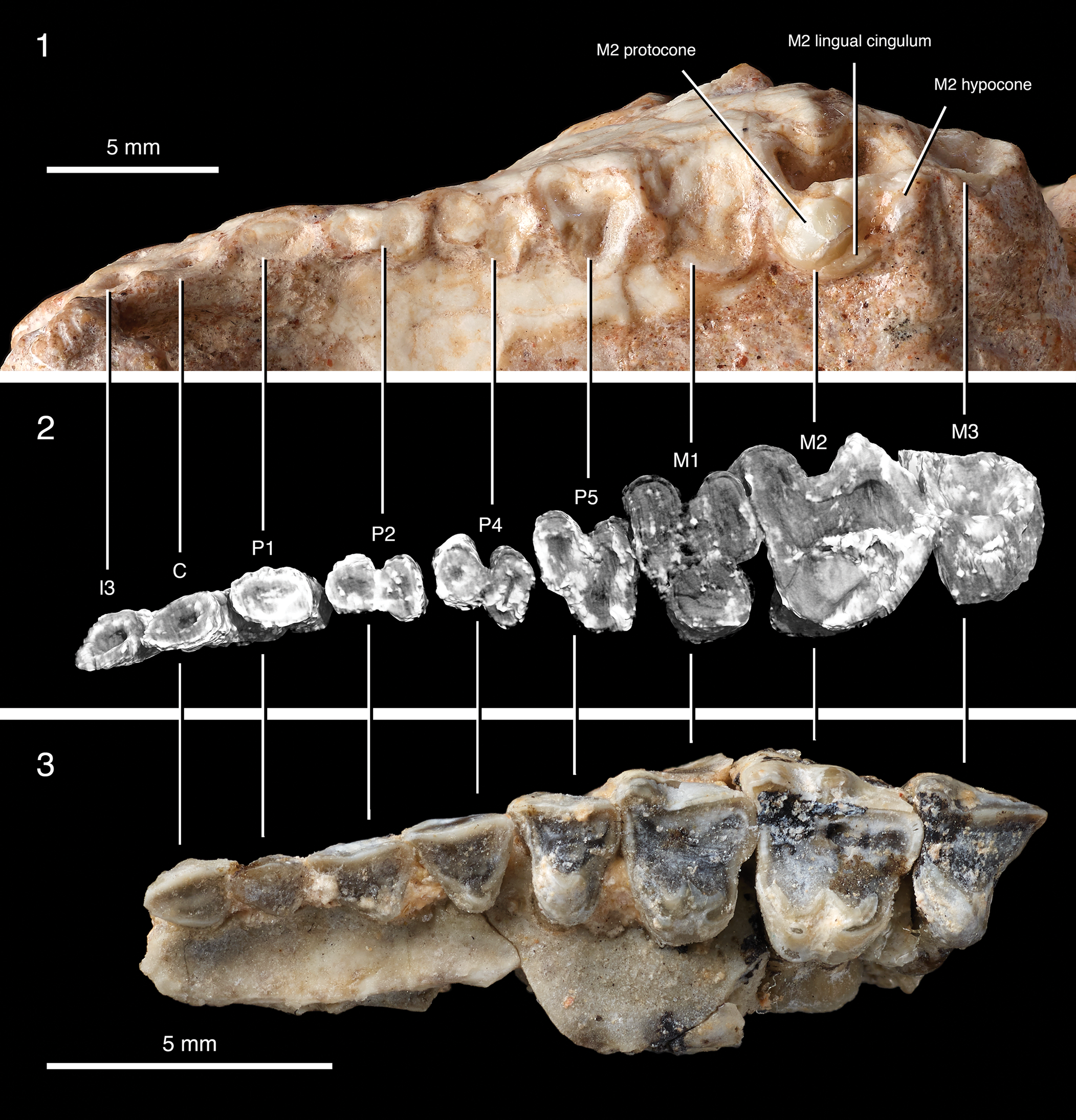
Figure 12. Left upper dentition of an arctostylopid, genus and species indet (PSS-MAE 673) with tooth loci (I3, C, P1–P5, M1–M3) indicated. (1) Digital photograph, (2) CT rendering, and (3) comparison with the upper dentition of Palaeostylops iturus, AMNH 21743. No tooth crown is fully preserved in PSS-MAE 673.
Description
The anterior cranium of PSS-MAE 673 shows surface bone only on the left side. In lateral view, the skull has a relatively short rostrum and a tall skull roof above the orbit. The face, particularly the anterior rostrum and the piriform aperture, has been elucidated with CT scans. The nasal bones are not preserved, but the position of what was the frontonasal suture appears largely intact. The anterior ends of the frontal bones protruded in the midline with the left and right sides meeting at a delicate point. The superior aspect of the piriform aperture is slightly recessed. The infraorbital foramen can be identified on the left side of the skull. Maxillopremaxillary sutures are not visible. In ventral view, the palate contains a large anterior opening, but it is not clear whether this was two separate incisive foramina or a single large opening. CT images show that choanae are preserved and the posterior edge of the palate is heart-shaped (posteriorly concave with a midline point), and that the posterior edges of the palatine were marked by a thickened rim known as a palatine torus. Interesting but more rarely described features of fossil mammal skulls, such as the sphenoid sinus, the basisphenoid, and a pterygoid process, also can be discerned in cross-sectional CT images of PSS-MAE 673.
The dentition is best preserved on the left side of the skull, but the right side was elucidated using a CT scan. The teeth are highly damaged and may also have been worn. Additionally, postmortem damage has resulted in a skewed relationship between the left and right sides of the dentition such that homologous alveoli are not aligned mediolaterally. Nonetheless either alveoli, roots, or partial teeth at each locus are preserved on each side.
In general, the specimen preserves tooth loci as roots at the junction of the crown. There is some distortion of the skull due to damage, and the right dentition is shifted slightly mesial to the left dentition. For reasons given below, we provisionally identify the most mesial alveoli on the right and left side as I3. Just distal to I3 are alveoli that we identify as those for the upper canines. The left I3 and C appear buccolingually compressed. Both have very worn crowns and there is no diastema between them. The CT scan shows that the pulp cavities of these teeth are exposed laterally, so the apparent buccolingual compression can be explained by breakage or erosion. What appears to be a root for the right I3 is rounder than the locus for I3 on the left. Additionally, the right root for I3 has an intact exterior surface that is surrounded by alveolar bone. Therefore, the right side is likely more informative about the incisor morphology.
There is no diastema between the canine and the succeeding postcanine tooth. Distal to the canine, a tooth root as it meets the crown base exists for what we hypothesize to be the P1, which has a rounded base and is not markedly large relative to the adjacent teeth. Both C and P1 are single-rooted. Despite this uncertainty in the identity of the most mesial alveoli, the homologies of the more distal teeth are less ambiguous, and we identify these as P2, P4 (traditional P3), P5 (traditional P4), M1, M2, M3. P2 is two-rooted, P4 has two distinct roots and an incipient third root, and P5–M3 are three-rooted. P2 is mesiodistally elongate and transversely narrow. P4 and P5 are preserved only as the bases of the tooth crowns, which are roughly triangular in occlusal outline. M1 and M2 are quadrate in occlusal outline. Only the left M2 preserves some features of the molar crown, a lingual moiety with a prominent conical protocone. Distal and slightly labial to the protocone is a distinct sulcus bordered distally by a slight rise that appears to be part of a hypocone. Although the distolingual corner of the crown is heavily worn and damaged, it is extensive, suggesting that the hypocone was well developed. A distinct lingual cingulum extends distally from the base of the protocone. The surface of the cingulum is marked by a wear facet and crenulations that may be two or three tiny cuspules. M3 is only preserved as a deeply worn base of the crowns and roots; the CT scan shows evidence of a highly worn protocone. The outlines of the premolars and molars as indicated by the roots show labial indentations, but the labial crowns are not preserved, and it cannot be determined whether an ectoflexus, or other aspects of this region of the crowns, was present or absent.
Materials
PSS-MAE 673, partial skull with worn and damaged crowns and tooth roots with partial crown of left M2. Compared directly with the following specimens and literature: Palaeostylops iturus, paratype AMNH 20415, left maxilla with partial C, P1, P2, P4, P5, M1–3 described by Matthew et al. (Reference Matthew, Granger and Simpson1929, fig. 2); AMNH 22143, left maxilla with I1–3, C, P1, P2, P4, P5, M1–3 described by Matthew et al. (Reference Matthew, Granger and Simpson1929, fig. 13); AMNH 21743, left maxilla and upper cheek teeth, numerous specimens from the PIN collections described and figured by Kondrashov and Lucas (Reference Kondrashov and Lucas2004, fig. 2); a large sample of Palaeostylops iturus in AMNH collections from the Gashato beds, Mongolia; Palaeostylops macrodon (Matthew, Granger, and Simpson, Reference Matthew, Granger and Simpson1929), AMNH paratype 22144, left maxilla with P2, P4, P5, M1–3, right maxilla with M1–3 described by Matthew et al. (Reference Matthew, Granger and Simpson1929, figs. 11, 12); a large sample of P. iturus and P. macrodon from the Gashat locality in the MPC-M collections figured, measured, and statistically analyzed by Missiaen et al. (Reference Missiaen, Escarguel, Hartenberger and Smith2012, fig. 2, table 1); Arctostylops steini Matthew, Reference Matthew1915, AMNH cast of MCZ 20004, anterior skull and associated mandible with nearly complete dentition described and figured by Cifelli et al. (Reference Cifelli, Schaff and McKenna1989, fig. 2); Allostylops periconatus Zheng, Reference Zheng1979, AMNH cast of IVPP 5043, type, anterior skull with well-preserved P2, P4, P5, M1–3 described by Zheng (Reference Zheng1979) from the Chijiang Basin, China; Asiostylops spanios Zheng, Reference Zheng1979, AMNH cast of IVPP 5042, holotype, skull and mandibles with premolars and molars described by Zheng (Reference Zheng1979); Anatolostylops dubius Zhai, Reference Zhai1978, AMNH cast of IVPP 4357, holotype, left M2–3 described by Zhai (Reference Zhai1978, fig. 8); Anatolostylops zhaii Wang et al., Reference Wang, Meng, Ni and Beard2008, IVPP 14657, holotype, and figured by Wang et al. (Reference Wang, Meng, Ni and Beard2008); Enantiostylops (Sinostylops) progressus (Tang and Yan, Reference Tang and Yan1976), AMNH cast of IVPP 4264, right maxillary fragment with M1–3, described by Tang and Yan (Reference Tang and Yan1976) as Sinostylops progressus, erected as Enantiostylops progressus by Averianov (Reference Averianov2020, appendix 1); Kazachostylops occidentalis Nessov, Reference Nessov1987, CCMGE 12/12455, right maxillary fragment with P4–5, M1–2 described and figured by Averianov (Reference Averianov2020, fig. 2); Migrostylops rosella Tong and Wang, Reference Tong and Wang2006, IVPP V10734: left I1–M3 described by Tong and Wang (Reference Tong and Wang2006, fig. 7.4; see also Missiaen et al., Reference Missiaen, Escarguel, Hartenberger and Smith2012, fig. 7.4).
Comparisons were also made with Hyopsodus orientalis Dashzeveg, Reference Dashzeveg1977, described by Dashzeveg (Reference Dashzeveg1977, figs. 3–8) from the base of the Bumban Member of the Naran Bulak Formation at the Tsagan Khushu locality, Mongolia, and various upper dentitions of Hyposodus from the early Tertiary of North America, including Hyopsodus paulus Leidy, Reference Leidy1870, AMNH 10974, anterior skull with right cheek tooth dentition, and Hyopsodus despiciens Matthew, Reference Matthew1909, AMNH 11877, holotype, skull and jaws with nearly complete dentition described by Matthew (Reference Matthew1909, figs. 103–105).
Comparisons were also made with various early perissodactyls: casts of Rhodopagus pygmaeus Radinsky, Reference Radinsky1965 (AMNH 21554); figures and descriptions of Rhodopagus (Radinsky, Reference Radinsky1965), Pataecops minutissimus (Reshetov, Reference Reshetov1979) (Averianov and Godinot, Reference Averianov and Godinot2005), Yimengia (Wang, Reference Wang1988; Bai et al., Reference Bai, Meng, Zhang, Gong and Wang2020; Paepen et al., Reference Paepen, Hong, Yan and Smith2021), Pataecops parvus Radinsky, Reference Radinsky1965, and Minchenolestes erlianensis Wang et al., Reference Wang, Meng, Jin, Beard, Bai, Li, Ni, Li and Gebo2011.
Measurements
All measurements of left upper tooth loci. We note that, with the exception of M2, the teeth are preserved only at the junction of the root and crown; thus, these dimensions are approximate. P2: maximum mesiodistal length (ML) = 3.25 mm, maximum labiolingual width (MW) = 1.5 mm; P4 (traditional P3), ML = 3.10 mm, MW = 2.25 mm; P5 (traditional P4) ML = 3.2 mm, MW = 4.1 mm; M1, ML = 3.5 mm, MW = 4.5 mm; M2, ML = 5 mm, MW = 5 mm; M3, ML = 3.5 mm, MW = 4.25 mm.
Remarks
This poorly preserved, enigmatic specimen from Daus-2b is so lacking in well- preserved dental features that its definite identification and allocation are precluded. Yet we argue here, based on comparisons of various taxa known from the Asian Paleocene and Eocene, that it has features most like those of arctostylopids. As in the diagnosis for Arctostylopidae (Cifelli et al., Reference Cifelli, Schaff and McKenna1989, p. 5), the Daus specimen lacks any evidence of a diastema, a feature typical of early perissodactyls compared here, such as the ceratomorphs that are common to Eocene faunas in Mongolia and Asia. Another potential similarity to arctostylopids relates to the number of teeth. In arctostylopids the dentition is complete or nearly complete, with the number of upper incisors varying from three (e.g., Palaeostylops iturus, see Matthew et al., Reference Matthew, Granger and Simpson1929, p. 12, fig. 13; Migostylops rosella Tong and Wang, Reference Tong and Wang2006) to two (e.g., Arctostylops steini, see Cifelli et al., Reference Cifelli, Schaff and McKenna1989, p. 8). Counting forward from M3 in the Daus specimen indicates the presence of a complete series between P1 and M3 and suggests that the three most-mesial alveoli are those for I3, C, and P1, in alignment with the condition in A. steini. There is enough damage at the anterior end of the rostrum to suggest that at least an I2 may have been lost in preservation. Also, in correspondence with the diagnosis of arctostylopids (Cifelli et al., Reference Cifelli, Schaff and McKenna1989), the alveoli in the Daus specimen indicate that the upper canine is subequal in dimensions to the I3 and P1.
Unfortunately, the presence of the most striking feature of the arctostylopid upper dentition—the salient, straight ectoloph on the upper molars—cannot be ascertained in the Daus specimen because all the crowns of the latter, with the exception of the lingual moiety of M2, are either heavily worn to their roots or destroyed. Nevertheless, a number of features of the dentition and skull bear strong resemblance to various arctostylopids. Based on the dimensions and outlines of the base of the crowns and the roots preserved, the M2 is notably larger and more quadrate than M1, a feature very characteristic of Palaeostylops (Matthew and Granger, Reference Matthew and Granger1925; Matthew et al., Reference Matthew, Granger and Simpson1929; Kondrashov and Lucas, Reference Kondrashov and Lucas2004), Arctostylops (Matthew, Reference Matthew1915; Cifelli et al., Reference Cifelli, Schaff and McKenna1989), Anatolostylops (Wang et al., Reference Wang, Meng, Ni and Beard2008), and other taxa. This feature strongly differs from that seen early condylarths, including Hyopsodus orientalis from the Bumbanian of Mongolia (Dashzeveg, Reference Dashzeveg1977), as well as early Asian perissodactyls (Radinsky, Reference Radinsky1965; Wang, Reference Wang1988; Averianov and Godinot, Reference Averianov and Godinot2005; Wang et al., Reference Wang, Meng, Jin, Beard, Bai, Li, Ni, Li and Gebo2011; Bai et al., Reference Bai, Meng, Zhang, Gong and Wang2020; Paepen et al., Reference Paepen, Hong, Yan and Smith2021) where the M1 and M2 do not differ so greatly in basal dimensions. Although M3 is badly damaged in the Daus specimen, it does appear notably smaller in overall dimensions than M2, a feature also characteristic of Palaeostylops, Arctostylops, and Anatostylops (Cifelli et al., Reference Cifelli, Schaff and McKenna1989, fig. 8). Other features of PSS-MAE 673 that are found in arctosylopids, including Palaeostylops iturus, are a P1 that is single-rooted and laterally compressed, P4, P5 with three roots, P4 base triangular in shape, M2 with a well-developed sulcus distal to the protocone, and a lingual cingulum with two or three small cuspules (see Kondrashov and Lucas, Reference Kondrashov and Lucas2004, p. 197–199). The expanded distolingual corner of the crown in M2 also suggests that a well-developed but heavily worn hypocone is present. Comparison with a cast of Arctostylops steini (MCZ 20004) shows that this species has a heart-shaped posterior margin of the palate with a well-developed palatine torus that closely resembles the condition in the Daus specimen.
Obviously, the preservation in PSS-MAE 673 does not allow very resolved identification, but the features of the lingual moiety of M2, the only partially preserved aspect of the crowns of the teeth, can be compared with those of described arctostylopids. The lingual cingulum and the suggestion of a well-developed hypocone compare closely with the condition in Palaeostylops iturus (Matthew and Granger, Reference Matthew and Granger1925; Kondrashov and Lucas, Reference Kondrashov and Lucas2004, fig. 2) and P. macrodon (Matthew et al., Reference Matthew, Granger and Simpson1929, fig. 12). It is of interest that Cifelli et al. (Reference Cifelli, Schaff and McKenna1989) elevated the latter species to a new genus, Gashatostylops macrodon, which is diagnosed by the presence of the relatively larger upper and lower second molars, larger incisors, cuspules on the lingual cingula, absence of a sulcus between the protocone and hypocone, two rather than three incisors, and a laterally constricted snout. Based on a large sample of Palaestylops iturus from the Gashatan horizon at Tsagan Khushu in the PIN collections, Kondrashov and Lucas (Reference Kondrashov and Lucas2004) found that nearly all the distinctive characters of Gashatostylops were either within the range of variation for specimens of P. iturus or could not be identified with certainty (e.g., the number of incisors) given the incomplete preservation in the cited specimens. In agreement with Kondrashov and Lucas (Reference Kondrashov and Lucas2004) and Missiaen et al. (Reference Missiaen, Escarguel, Hartenberger and Smith2012), we recognize the original designation of Matthew et al. (Reference Matthew, Granger and Simpson1929) of P. macrodon.
As in Palaeostylops, the lingual region of M2 in Arctostylops shows a well-developed hypocone, but the lingual cingulum extends without interruption from the mesial face of the protocone to the distal face of the hypocone (Cifelli et al., Reference Cifelli, Schaff and McKenna1989, figs. 2, 8). In contrast to these genera, Asiostylops spanios Zheng, Reference Zheng1979, from the late Paleocene of China (Li and Ting, Reference Li and Ting1983), either lacks a hypocone, or the hypocone is represented by a small cusp at the lingual terminus of a broad postcingulum. Kazachostylops occidentalis from the late Paleocene of Kazakhstan also lacks a hypocone on the upper molars, but has both pre- and postcingula that are non-continuous (Nessov, Reference Nessov1987; Averianov, Reference Averianov2020). Anatostylops dubius and A. zhaii from the early Eocene of China are distinctive in having a protocone and hypocone placed well labially and forming a continuous ridge separated by a deep fossette from the ectoloph (Zhai, Reference Zhai1978; Wang et al., Reference Wang, Meng, Ni and Beard2008). Allostylops periconatus Zheng, Reference Zheng1979, from the late Paleocene of China (Li and Ting, Reference Li and Ting1983) is distinguished in having a prominent pericone mesiolingual to the protocone on the upper molars. Enantiostylops (Sinostylops) progressus (Tang and Yan, Reference Tang and Yan1976) was distinguished as a new genus by (Averianov, Reference Averianov2020, appendix 1) in having a strong internal invagination on the parastyle of M1 and M2. Migrostylops rosella (Tong and Wang, Reference Tong and Wang2006) from the early Eocene Wutu Formation of China has a continuous lingual cingulum extending from the mesial face of the protocone to the distal face of the hypocone, a relatively weak, more labially positioned hypocone, and a relatively larger M3. Bothriostylops notios Zheng and Huang, Reference Zheng and Huang1986, from the late Paleocene of China is distinguished only by the lower dentition. Thus, the limited morphology preserved in the M2 of PSS-MAE 673 most closely resembles that of Palaeostylops. Averianov (Reference Averianov2020) provided a revised phylogeny for the Arctostylopidae that grouped Palaeostylops, Arctostylops, Anatostylops, and Migrostylops in a clade (Arctostylopinae) that excludes the other arctostylopids compared above.
As noted, the tooth measurements of PSS-MAE 673 are only approximate, but the teeth are clearly larger than in Palaeostylops iturus, comparable in size to Palaeostylops macrodon, and smaller than in Arctostylopus steini and Anatostylops zhaii. Based on a sample of 21 specimens, Missiaen et al. (Reference Missiaen, Escarguel, Hartenberger and Smith2012, table 1) calculated a mean value of 5.01 mm for maximum length of M2 in P. macrodon. The subequal length and width estimated for M2 in the Daus specimen (5 × 5 mm) is similar to, but slightly greater than, these dimensions in Kazachostylops occidentalis (4.6 × 4.5 mm in Averianov, Reference Averianov2020, table 1), but only a better-preserved specimen than PSS-MAE 673 will provide more meaningful size comparisons.
In summary, the incomplete features preserved in PSS-MAE 673 only allow a provisional identification of this new fossil as an indeterminate genus and species of arctostylopid. The specimen, as noted above, does show similarities to Palaeostylops iturus, a taxon that is abundantly represented in Gashatan faunas of Mongolia (see compilations in Kondrashov and Lucas, Reference Kondrashov and Lucas2004; Missiaen et al., Reference Missiaen, Escarguel, Hartenberger and Smith2012). The Daus specimen may belong to a taxon larger than Palaeostylops iturus, but similar with respect to features of the lingual area of M2, including the well-developed crenulated cingulum.
Arctostylops was once thought to be an index taxon for the Asian Paleocene based on the presence of this genus also in the Paleocene of North America. As noted above, however, the Asian Arctostylops species, A. iturus (see Dashzeveg and Russell, Reference Dashzeveg and Russell1988), has been re-assigned to Palaeostylops iturus (Cifelli et al., Reference Cifelli, Schaff and McKenna1989; Kondrashov and Lucas, Reference Kondrashov and Lucas2004), a species restricted to Asia, thus diminishing the role of this taxon as a global index fossil. The abundance of Palaeostylops in the Gashatan faunas may be biostratigraphically informative, at least within Asia, although arctostylopids are also represented in Bumbanian-aged faunas in China (Wang et al., Reference Wang, Meng, Ni and Beard2008). The more recent higher taxonomic assignment of arctostylopids to Gliriformes (Missiaen et al., Reference Missiaen, Smith, Guo, Bloch and Gingerich2006) would obviate the long-held notion that the family belongs to the Notoungulata and part of a taxon of remarkable geographic range extending through Eurasia, North America, and South America. The taxonomic assignment by Missiaen et al. (Reference Missiaen, Escarguel, Hartenberger and Smith2012), however, has not been entirely accepted. For example, Averianov (Reference Averianov2020) recognized arctostylopids as belonging to the order Arctostylopida, which was erected by Cifelli et al. (Reference Cifelli, Schaff and McKenna1989), an assignment we adopt here, pending greater resolution of the matter.
Discussion
We describe the lithology and paleontology of three new Naran Bulak Formation localities situated >90 km east of the type section (Gradziński et al., Reference Gradziński, Kazmierczak and Lefeld1969; Dashzeveg and McKenna, Reference Dashzeveg and McKenna1977; Dashzeveg, Reference Dashzeveg1988). The new site has yielded both invertebrates and fossil mammals, including some of the first reported skull material of two taxa: the gliroid Gomphos and a partial skull with a worn and damaged dentition that we provisionally identify as an indeterminate genus and species of an arctostylopid.
The Nemegt Basin is significant for being, along with the Lingcha Formation, Hengyang Basin, Hunan, China, one of the few localities in Asia that has continuous Paleocene–Eocene sedimentary rocks (Ting et al., Reference Ting, Bowen, Koch, Clyde, Wang, Wang and McKenna2003, p. 522). This feature, along with the superposition of the new localities on the highly fossiliferous Djadokhta Formation Cretaceous rocks at Ukhaa Tolgod (Dingus et al., Reference Dingus, Loope, Dashzeveg, Swisher, Minjin, Novacek and Norell2008), makes additional exploration of these new deposits important for their potential to improve stratigraphy across the Cretaceous-Paleogene and Paleocene-Eocene boundaries in Asia.
Biostratigraphy has been the primary method of dating the Naran Bulak Formation because the formation lacks beds amenable to radiometric dating. Significant prior work exists naming Asian Land Mammal Ages (ALMAs) and fossils that characterize them. Noteworthy regarding the data in this paper is that the fossils collected in situ from the new Daus localities exhibit a mixed temporal signature when compared to prior findings. Lithologically, the localities are in the Naran Member (or ‘White Beds’) of the Naran Bulak Formation, which has been associated repeatedly with the Gashatan ALMA and is equivalent to the Nungshanian (Li and Ting, Reference Li and Ting1983), and assigned a Paleocene age (Kielan-Jaworowska and Dovchin, Reference Kielan-Jaworowska and Dovchin1968; Gradziński, Reference Gradziński1969; Szczechura, Reference Szczechura1971; Shuvalov et al., Reference Shuvalov, Reshetov and Barsbold1974; Shishkin, Reference Shishkin1975; Dashzeveg, Reference Dashzeveg1982, Reference Dashzeveg1988; Beard, Reference Beard1998; Meng et al., Reference Meng, Zhai and Wyss1998; Ting, Reference Ting1998; Wang et al., Reference Wang, Hu, Zhou and Li1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002, Reference Bowen, Koch, Meng, Ye and Ting2005; Ting et al., Reference Ting, Bowen, Koch, Clyde, Wang, Wang and McKenna2003; Clyde et al., Reference Clyde, Tong, Snell, Bowen, Ting, Koch, Li, Wang and Meng2008; Lopatin, Reference Lopatin2020). The presence of Archaeolambda, known previously from Gashatan (and equivalent Nungshanian) or older rocks (Li and Ting, Reference Li and Ting1983; Ting, Reference Ting1998; Wang et al., Reference Wang, Hu, Zhou and Li1998; Bowen et al., Reference Bowen, Clyde, Koch, Ting, Alroy, Tsubamoto, Wang and Wang2002), suggests a Paleocene age for the new localities consistent with current hypotheses about the age of Naran Member beds. However, we note that Dashzeveg (Reference Dashzeveg1988, figs. 2, 4) had at one time proposed an Eocene age for Archaeolambda and that Lopatin (Reference Lopatin2020) underscored species-level differences for this taxon between Paleocene and Eocene beds. The presence of an arctostylopid with some dental similarities with the abundantly represented Gashatan genus Palaeostylops potentially also suggests a Paleocene age for the Daus fauna. However, this Paleocene temporal signature is contradicted by the presence of a taxon that has been recognized as a signature for the younger Bumban Member, the Bumbanian ALMA, and the early Eocene: Gomphos elkema (Dashzeveg, Reference Dashzeveg1982, Reference Dashzeveg1988; Dashzeveg and Russell, Reference Dashzeveg and Russell1988; Meng et al., Reference Meng, Bowen, Ye, Koch, Ting, Li and Jin2004). Additionally, Pantolamdodon sp., recorded as an astragalus in the Daus beds, is known elsewhere in Asia from the late early-middle Eocene (Paepen et al., Reference Paepen, Hong, Yan and Smith2021). Finally, the presence of the ostracode Limnocythere nemegtensis also recently has been associated with Gashatan beds outside of Mongolia and considered Paleocene (Wang et al., Reference Wang, Li, Bai, Jin, Mao and Meng2019). However, this endemic Asian taxon has not yet met the criteria to be an index fossil of broad use. Thus, a mixed Paleocene and Eocene signal emerges from the new fossils recovered at this Naran Member locality. Alternative hypotheses have been proposed and may warrant further consideration, including that the Gashatan ALMA (Nungshanian) spans the Paleocene-Eocene boundary (Lucas, Reference Lucas, Aubry, Lucas and Berggren1998) or that the Bumbanian faunas are late Paleocene (Beard, Reference Beard1998).
Two diminutive skulls are among the remains found in this first exploration of the Daus localities. Like the Cretaceous Djadokhta Formation, the Naran Bulak beds at Daus appear, at least preliminarily, to have a tendency to preserve small and delicate mammal skulls, a taphonomic signature that invites further exploration. Finally, more extensive study of the fluvio-lacustrine paleoecology suggested by these new localities, such has been done for similar deposits described elsewhere on the Mongolian plateau in Inner Mongolia, People's Republic of China (Van Itterbeeck et al., Reference Van Itterbeeck, Missiaen, Folie, Markevich, Van Damme, Gou and Smith2007), will be necessary to better understand the paleoenvironment in which these early mammals existed.
Acknowledgments
We are extremely grateful to the following people for expedition collaboration and research assistance: S. Goldberg, C. Merrill, and R. O'Leary. For helpful discussion on the manuscript we thank L. Antonietto, J. Meng, L.T. Holbrook, K.D. Rose, and S.P. Zack. This field research was accomplished with funding from the Margaret and Will Hearst Paleontological Research Fund and the Frick Laboratory Endowment at the AMNH. We thank the National Science Foundation for supporting this research through award DBI 0619559 for the AMNH SEM.
Declaration of competing interests
The authors declare none.
Data availability statement
Additional data is stored in the public database MorphoBank at project P4217: http://dx.doi.org/10.7934/P4217.













 Open access
Open access