



Introduction
Anticosti Island (Québec, Canada) is well known for its stratigraphically thick and fossiliferous Ordovician/Silurian boundary section, with a diverse marine invertebrate fauna consisting of corals, brachiopods, mollusks, trilobites, and crinoids that have been the subject of taxonomic investigations for over a century (Logan, Reference Logan1863; Twenhofel, Reference Twenhofel1928; Bolton, Reference Bolton and Lesperance1981). Among these groups, the best documented have been the rhynchonelliform brachiopods, which have been described extensively in a series of monographs and papers (e.g., Jin, Reference Jin1989; Copper, Reference Copper1995; Dewing, Reference Dewing1999; Jin and Copper, Reference Jin and Copper2000; Jin and Zhan, Reference Jin and Zhan2008; Copper and Jin, Reference Copper and Jin2017). Turnovers within and among the orders of Rhynchonelliformea (Williams et al., Reference Williams, Carlson, Brunton, Holmer and Popov1996) have played a critical role in defining the terminal-Ordovician Hirnantian Stage, as well as the placement of the Ordovician/Silurian boundary on Anticosti Island (Bolton, Reference Bolton and Lesperance1981; Copper, Reference Copper2001; Copper et al., Reference Copper, Jin and Desrochers2013).
There are two major turnovers in brachiopod assemblages recorded throughout the Upper Ordovician on Anticosti Island. The first interval of brachiopod turnover is best observed along the western coast of Anticosti Island, where it is expressed as a gradual transition across the contact between the Vaureal and Ellis Bay formations. Notably, the lowermost member of the Ellis Bay Formation (sensu Copper et al., Reference Copper, Jin and Desrochers2013) is marked by the first occurrences of the spire-bearing Hindella Davidson, Reference Davidson1882 and Eospirigerina Boucot and Johnson, Reference Boucot and Johnson1967. Throughout the Ellis Bay Formation, there are no drastic turnovers in brachiopod assemblages, although Jin and Copper (Reference Jin and Copper2008) documented localized shifts in brachiopod associations. At the top of the Ellis Bay Formation, Jin and Zhan (Reference Jin and Zhan2008) recognized the type species of Hirnantia, H. sagittifera (M'Coy, Reference M'Coy1851), but extensive collections have demonstrated it is found only within the reefal Laframboise Member. Across the Ellis Bay/Becscie formational contact, a second major turnover in brachiopod assemblages is marked by the replacement of the typical Ellis Bay brachiopod assemblage by an assemblage dominated by smaller-shelled orthides, virginids, and athyrides (Jin et al., Reference Jin, Long and Copper1996; Li and Copper, Reference Li and Copper2006; Copper and Jin, Reference Copper and Jin2017). This turnover is observable in every exposure of the contact between the Ellis Bay and Becscie formations.
Recognizing these two major intervals of faunal turnover, Copper et al. (Reference Copper, Jin and Desrochers2013) interpreted the entire Ellis Bay Formation as Hirnantian in age, with each faunal transition representing a hypothesized pulse of the Late Ordovician mass extinction. In their framework, a Hirnantian age of the Ellis Bay Formation was supported by the first occurrences of the genera Hindella and Eospirigerina within the Fraise Member along the western coast of Anticosti Island, as well as the presence of Hirnantia in the uppermost Laframboise Member. According to Copper and Jin (Reference Copper and Jin2017), true Hindella species are confined largely to the Hirnantian, with only Hindella terebratulina (Wahlenberg, Reference Wahlenberg1818) from the limestone capping the Boda reef mounds possibly extending downward in the uppermost Katian (Sheehan, Reference Sheehan1977). In recent studies, however, the top of the Boda Limestone has been considered Hirnantian in age (e.g., Webby, Reference Webby2002; Suzuki et al., Reference Suzuki, Shiino and Bergström2009). This would imply Hindella is an “index” genus for the Hirnantian. However, Eospirigerina has been reported from notably older Late Ordovician rocks, such as the upper Dulankara Formation of Chu-Ili, Kazakhstan (Popov et al., Reference Popov, Nikitin and Sokiran1999). Furthermore, the brachiopod assemblages of the Ellis Bay Formation lack many taxa of the classic Hirnantia fauna that are characteristic of the so-called cool-water Kosov faunal province in marginal and peri-Gondwana, expanding from high southern latitudes (e.g., Colmenar et al., Reference Colmenar, Villas, Rasmussen, Hunter, Álvaro, Lefebvre, van Roy and Zamora2018) to tropical latitudes during the Hirnantian (for a summary, see Rong et al., Reference Rong, Chen and Harper2002; Huang et al., Reference Huang, Zhou, Harper, Zhan, Zhang, Chen and Rong2020a; Rong et al., Reference Rong, Harper, Huang, Li, Zhang and Chen2020a).
Here we report a new species of Hirnantia, H. notiskuani, from eastern exposures of the lowermost Ellis Bay Formation (sensu Copper et al., Reference Copper, Jin and Desrochers2013) on Anticosti Island. In many regards, this new species is similar to the morphologically plastic H. sagittifera except for the development of a uniplicate anterior commissure in both juvenile and mature specimens, a feature that has not been observed in the type species. Where it is most abundant, H. notiskuani is found in association with Nasutimena fluctuosa (Billings, Reference Billings1860), a brachiopod characteristic of Laurentian epicontinental seas during the Katian. Given the significance of Hirnantia as an indicator of the Hirnantian Stage in tropical basins, its recognition from the sub-Laframboise Ellis Bay Formation provides new biostratigraphic context for understanding patterns of faunal turnover within the Ellis Bay Formation in central and eastern areas of Anticosti Island, thus providing strong evidence for placing the lower Ellis Bay Formation within the Hirnantian Stage.
Geologic setting
The exposed Upper Ordovician–lower Silurian stratigraphic succession of the Anticosti Basin is nearly a kilometer thick and divided into eight formations: the Upper Ordovician Vaureal and Ellis Bay formations and the lower Silurian Becscie (Copper and Jin, Reference Copper and Jin2014), Merrimack (Copper and Long, Reference Copper and Long1989), Gun River (Copper et al., Reference Copper, Long and Jin2012), Menier, Jupiter, and Chicotte formations (Copper and Jin, Reference Copper and Jin2015). In this paper, we focus on the stratigraphic interval spanning the uppermost Vaureal and lowermost Ellis Bay formations. These formations were deposited in a peripheral zone of the Appalachian foreland basin, the Anticosti Basin (Sanford, Reference Sanford, Wheeler, Scott and Aitken1993), which was situated along the southern margin of Laurentia, approximately 20° south of the equator (Torsvik and Cocks, Reference Torsvik and Cocks2016). Subsidence rates in the Anticosti Basin peaked during the Late Ordovician but decreased markedly across the Ordovician/Silurian boundary, beginning with the deposition of the uppermost Vaureal and Ellis Bay formations (Long, Reference Long2007; Lavoie, Reference Lavoie and Miall2008). Despite the influence of the Appalachian thrust sheet on the development of the Anticosti Basin, the stratigraphic succession on Anticosti Island was located on a promontory of the stable basement rock of the Canadian Shield beyond the Taconic deformation front and did not experience structural deformation associated with the Taconic Orogeny (Lavoie, Reference Lavoie and Miall2008; Pinet et al., Reference Pinet, Keating, Lavoie, Dietrich, Duchesne and Brake2012). This allowed for the development of a gently dipping homoclinal carbonate ramp, with deposition largely controlled by storm processes (Sami and Desrochers, Reference Sami and Desrochers1992; Long, Reference Long2007; Desrochers et al., Reference Desrochers, Farley, Achab, Asselin and Riva2010). The combination of high subsidence rates and gentle ramp gradient resulted in the deposition of a thick wedge of mixed carbonate and fine-grained siliciclastic sediments spanning the Ordovician/Silurian boundary.
Upper Ordovician and lower Silurian strata are exposed in an east–west trending belt spanning the entire length of Anticosti Island (~200 km), roughly oblique to the present southwestern dip of the ancient ramp (Desrochers and Gauthier, Reference Desrochers and Gauthier2009). Upper Ordovician strata can be divided into a western facies suite, comprising largely mixed carbonate and shale facies deposited within the offshore transition zone, and an eastern facies suite, composed of mixed carbonate–siliciclastic facies deposited in environments ranging from the foreshore to the offshore transition zone (Long and Copper, Reference Long and Copper1986; Copper et al., Reference Copper, Jin and Desrochers2013). While lithostratigraphic and sequence stratigraphic frameworks (e.g., Desrochers et al., Reference Desrochers, Farley, Achab, Asselin and Riva2010) have been proposed to correlate the western and eastern facies suites of the Upper Ordovician strata on Anticosti Island, in particular the members of the Ellis Bay Formation, rapid lateral facies shifts across the eastern and central parts of the basin continue to complicate regional stratigraphic correlation, as recognized in the original paper by Long and Copper (Reference Long and Copper1986) describing the members of the Ellis Bay Formation. We therefore tentatively use the nomenclature of Long and Copper (Reference Long and Copper1986) and Copper et al. (Reference Copper, Jin and Desrochers2013), pending further revision of correlation among eastern and central exposures of the Ellis Bay Formation.
Locality and stratigraphic information
Collections were made at three localities spanning the easternmost exposures of the lowermost Ellis Bay Formation (Fig. 1).

Figure 1. Map of the northeastern coast of Anticosti Island (Québec, Canada), marking the locations of Hirnantia-bearing exposures of the Prinsta Member (black circles). Inset map shows the location of Anticosti Island and the area detailed in the main map (Datum: WGS84).
Anse Mauvaise (Lousy Cove)
Samples were collected from the lower portion of the Prinsta Member at its type section at Anse Mauvaise (49.335°N, 61.887°W), where it overlies the cross-stratified sandstones of the uppermost Velleda Member of the Vaureal Formation (Fig. 2). Here the contact between the sandier Velleda Member and shalier Prinsta Member has been interpreted as a flooding surface coinciding with a regional rise in relative sea level (Desrochers et al., Reference Desrochers, Farley, Achab, Asselin and Riva2010). The Prinsta Member is expressed as nodular calcareous shales interbedded with very thin to thin-tabular grainstones and sandstones. The basal unit of the Prinsta Member contains half-meter-thick, reworked accumulations of aulacerids, stromatoporoids, and tabulate corals, referred to as “aulacerid biostromes” by Long and Copper (Reference Long and Copper1986).
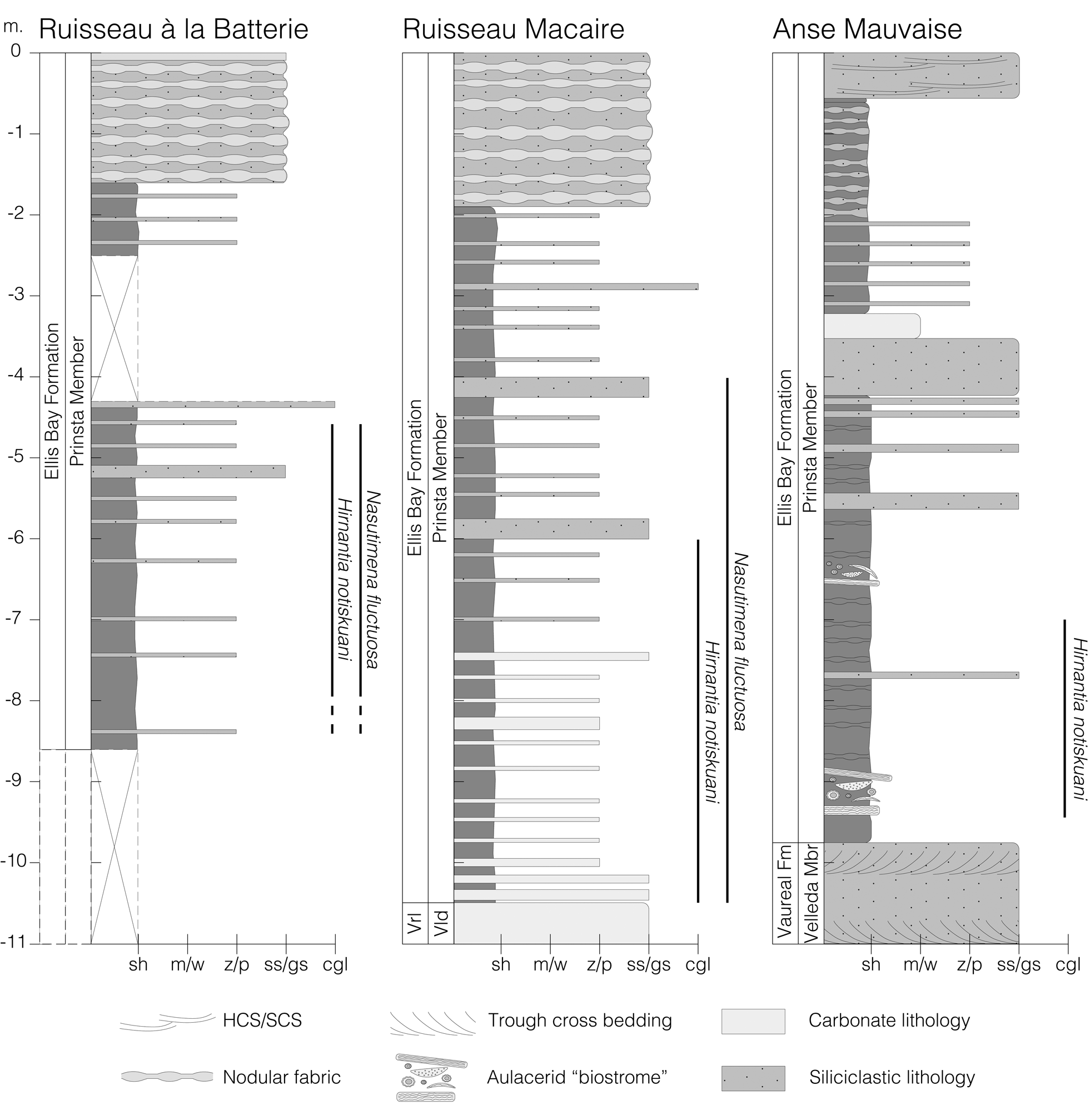
Figure 2. Stratigraphic columns illustrating the uppermost Velleda Member (Vaureal Formation) and Prinsta Member (Ellis Bay Formation) for each of the three localities in this study, as well as the stratigraphic range of Hirnantia notiskuani n. sp. and Nasutimena fluctuosa at each locality. The datum for each column is the base of the Lousy Cove Member of the Ellis Bay Formation (sensu Long and Copper, Reference Long and Copper1986). sh = shale; m/w = lime mud or wackestone; z/p = silt or packstone; ss/gs = sandstone or grainstone; cgl = conglomerate.
Ruisseau Macaire
Samples were collected from Ruisseau Macaire (49.381°N, 62.193°W) within an interval that is correlative with the type section of the Prinsta Member at Anse Mauvaise (Fig. 2). Here the base of the Prinsta Member is tentatively placed at the contact between recessive-weathering calcareous shales and the underlying weathering-resistant thin wavy-bedded bryozoan packstones to grainstones interbedded with laminar shale. Across this contact, the packstone to grainstone beds of the Prinsta Member gradually thin, while the relative proportion of calcareous shale increases, a trend that may reflect the rise in sea level observed across the Velleda/Prinsta contact at Anse Mauvaise. Throughout the measured section, the packstones to grainstones are gradually replaced by very thin to thin-tabular calcareous siltstone beds. The member is capped by nearly two meters of thin nodular very fine sandy grainstones with laminar shale interbeds, which is overlain by the argillaceous limestones of the Lousy Cove Member.
Ruisseau à la Batterie
Samples were collected from the uppermost Prinsta Member at Ruisseau à la Batterie (49.385°N, 62.237°W). Here the lower part of the Prinsta Member is not exposed due to damming of the river by beavers. The exposed part of the formation consists of calcareous shales interbedded with very thin to thin-tabular calcareous siltstone beds, which are overlain by nearly two meters of thin nodular very fine sandy grainstones with laminar shale interbeds (Fig. 2). This nodular interval is erosively overlain by the argillaceous limestones of the Lousy Cove Member, similar to the stratigraphic succession at Ruisseau Macaire.
Materials and methods
CT scanning
Specimens were scanned using a GE Phoenix Nanotom M micro-CT system in the Functional Anatomy and Vertebrate Evolution Laboratory, University of California, Berkeley, with an energy setting of 120 kV and current setting of 190 mA. The raw project images were reconstructed in the Phoenix Datos software at 0.126 microns per voxel and exported as an image stack in TIFF format.
SEM imaging
Before imaging, specimens were sputter coated in a layer of osmium (~14 nm thick) using a Filgen OPC 80T. Specimens were imaged using a Zeiss 1540XB field emission scanning electron microscope in the Western University Nanofabrication Facility.
Repositories and institutional abbreviations
Specimens figured and otherwise referenced in this study are deposited in the following institutions: American Museum of Natural History (AMNH-FI), New York, United States of America; the Nanjing Institute of Geology and Palaeontology (NIGP), Nanjing, China; and the Geological Survey of Canada (GSC), Ottawa, Canada.
Systematic paleontology
Order Orthida Schuchert and Cooper, Reference Schuchert and Cooper1932
Superfamily Enteletoidea Waagen, Reference Waagen1884
Family Draboviidae Havlíček, Reference Havlíček1950
Genus Hirnantia Lamont, Reference Lamont1935
Type species
Orthis sagittifera M'Coy, Reference M'Coy1851, Foel-y-Ddinas Mudstone Formation (Hirnantian), Wales, UK.
Hirnantia notiskuani new species
Figures 3–8
- Reference Jin and Zhan2008
Hirnantia sp. nov. A, Jin and Zhan, p. 53, pl. 35, figs. 1–16.
Types
Holotype, AMNH-FI 137221 (Fig. 3.1–3.7); Prinsta Member (Hirnantian), Ellis Bay Formation, Ruisseau Macaire locality, eastern Anticosti Island, Québec, Canada. Figured paratypes (all from the same locality and stratigraphic interval), AMNH-FI 137220 (Fig. 4.1–4.8), AMNH-FI 137223 (Fig. 5.1–5.5), AMNH-FI 137224 (Fig. 5.6–5.14), and one dorsal valve AMNH-FI 137222 (Figs. 3.8–3.9, 4.9).
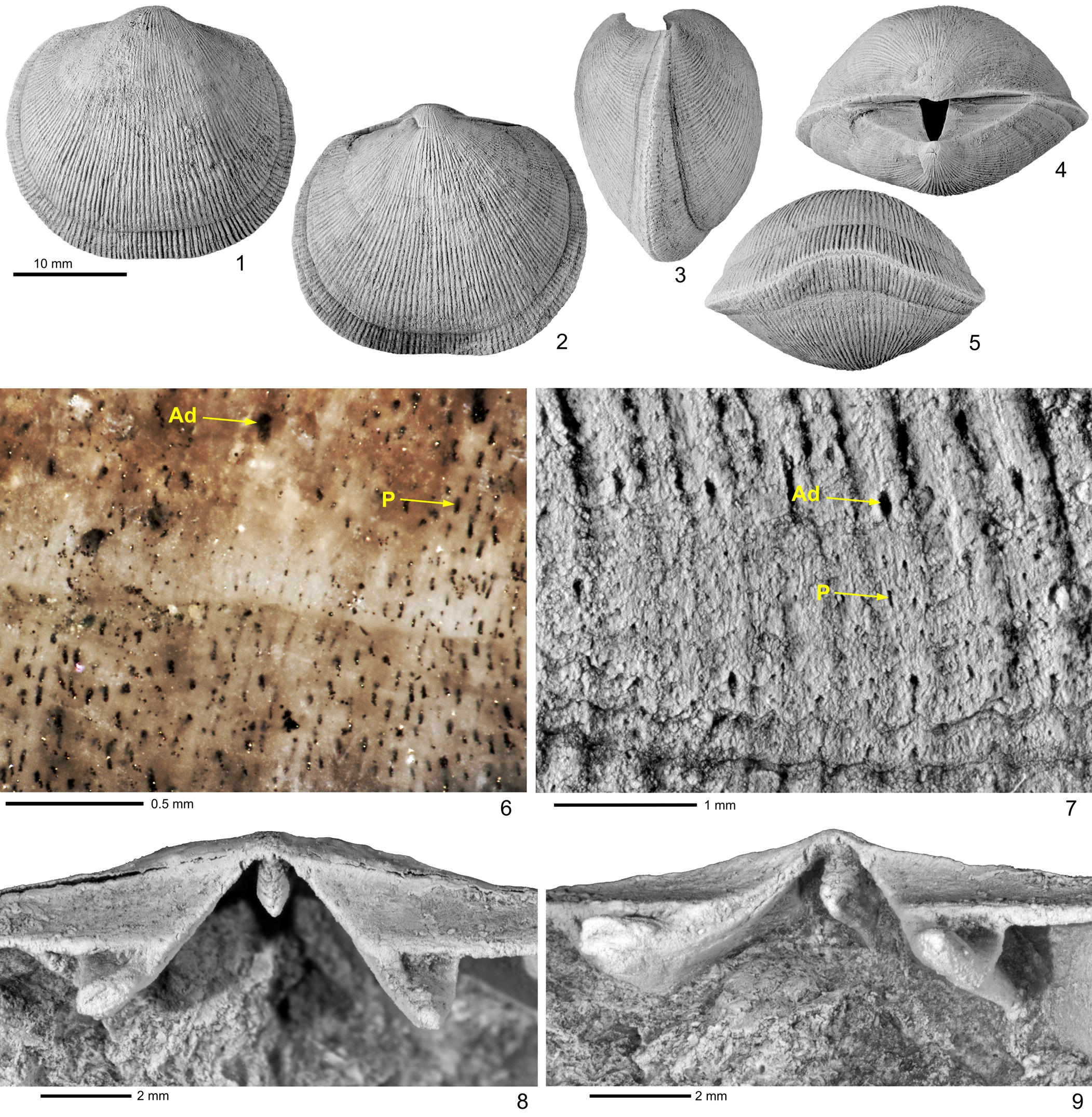
Figure 3. Hirnantia notiskuani n. sp. from the Prinsta Member of the Ellis Bay Formation, Hirnantian, at Ruisseau Macaire, eastern Anticosti Island. (1–7) Holotype AMNH-FI 137221: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of a strongly dorsibiconvex shell, demonstrating the well-developed uniplicate anterior that distinguishes this species; (6, 7) local enlargement showing aditicules and densely spaced punctae that characterize the shell microstructure. (8, 9) Paratype AMNH-FI 137222: enlargement of the dorsal interior of a uniplicate shell illustrating the cardinalia of H. notiskuani, with a high shaft and relatively small, bilobed, crenulated myophore; (8) perpendicular view; (9) oblique view. Ad = aditicule; P = punctae.

Figure 4. Hirnantia notiskuani n. sp. from the Prinsta Member of the Ellis Bay Formation, Hirnantian, at Ruisseau Macaire, eastern Anticosti Island. (1–8) Paratype AMNH-FI 137220: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of a juvenile individual with a rectimarginate anterior commissure; (6–8) local enlargement of (6) shell posterior, depicting transverse striae on ventral interarea, (7) interior of dorsal umbo as in (6) but rotated, revealing cardinal process, and (8) aditicules. (9) Paratype AMNH-FI 137222: oblique view of dorsal interior of a uniplicate shell.

Figure 5. Hirnantia notiskuani n. sp. from the Prinsta Member of the Ellis Bay Formation, Hirnantian, at Ruisseau Macaire, eastern Anticosti Island. (1–5) Paratype AMNH-FI 137223: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of a juvenile individual with a broadly uniplicate margin. (6–14) Paratype AMNH-FI 137224: (6) dorsal, (7) ventral, (8) lateral, (9) posterior, and (10) anterior views of a moderately convex, broadly uniplicate shell; (11, 12) SEM images of shell surface showing aditicules (often filled with pyrite, see (13)) and densely spaced punctae; (13) enlargement of shell, photographed while immersed in alcohol, to highlight punctae and aditicules, both filled by pyrite; (14) enlargement of anterior commissure showing densely spaced aditicules along the anterior margin of the shell: note that aditicules can be variably arranged in single and double columns.
Diagnosis
Dorsibiconvex shells of Hirnantia with a uniplicate anterior commissure, best developed at gerontic stage. Predominantly one column of aditicules per rib. Cardinal process consisting of relatively narrow, bilobed myophore with chevron-like crenulations and high, blade-like shaft merging anteriorly with a low median ridge. Brachiophore plates sitting directly on valve floor, diverging anteriorly from each other at ~80°.
Occurrence
All specimens are from the Prinsta Member of the Ellis Bay Formation (Late Ordovician) at Ruisseau Macaire, Ruisseau à la Batterie, and Anse Mauvaise sections along the northeastern coast of Anticosti Island, Québec, Canada.
Description
Shell medium to large for draboviids, subcircular to transversely ellipsoid in outline, biconvex to dorsibiconvex in profile; maximum width located around mid-length of shell. Hinge line straight, with rounded cardinal extremities, reaching 50–60% of maximum shell width. Anterior commissure rectimarginate in some small and medium-sized specimens (Fig. 4.5), often gently uniplicate, particularly in large, gerontic (strongly convex) specimens (Figs. 3.5, 5.10). Shell surface multicostellate, with costellae increasing in number by both intercalation and bifurcation on both valves (Fig. 4.8). Fine capillae present between some costellae. Growth lines usually better developed in anterior parts of shell. Predominantly single column of aditicules per rib, sporadically interspersed with two columns of aditicules, densely spaced at shell margin (Fig. 5.11–5.14). Punctae fine, densely spaced, oriented at fairly low angles to the shell surface (Figs. 4.8, 5.13).
Ventral exterior
Umbo weakly convex with suberect to slightly incurved beak. Interarea relatively high, apsacline curved in longitudinal profile, bearing transverse striae (Fig. 3.3, 3.4). Delthyrium open (Fig. 3.4). Gentle sulcus developed in anterior part, particularly of large specimens (Fig. 3.5).
Dorsal exterior
Valve moderately to strongly but uniformly convex, resulting in central part of valve having greatest convexity and depth, particularly in mature specimens. Umbo more strongly convex than ventral umbo, with ventrally directed beak sometimes extending slightly beyond commissural plane (Figs. 3.3, 5.3). Interarea lower than that of ventral valve, very slightly anacline to orthocline, with slightly curved longitudinal profile. Notothyrium open (Fig 4.7). Gentle fold developed anteriorly in large specimens to accommodate anterior sulcus of opposite valve (Fig. 3.5).
Ventral interior
Dental plates present, weakly divergent anteriorly, extending anteriorly into low lateral bounding ridges of muscle field (Fig. 6.5–6.12). Pedicle callist present in apical area of umbonal cavity, represented by thin, transverse plate located just below delthyrial opening (Fig. 7).

Figure 6. Serial cross sections of Hirnantia notiskuani n. sp. (AMNH-FI 137220), created using micro-CT scanning, progressively moving from the posterior to the anterior part of the shell. Lighter gray areas indicate shell calcite, while darker gray areas indicate infilling matrix; note the low contrast between the shell and enclosed carbonate-rich matrix. White splotches and lines are likely pyritized grains; the smearing effect above the specimen is caused by the positioning of the electron source. CT images reveal several internal morphological features: (1–4) a high, thin, plate-like shaft of the cardinal process, (4–12) which extends anteriorly as a low, rounded median ridge; (5–9) weakly divergent dental plates, (10–12) extending as low lateral bounding ridges of the ventral muscle field.

Figure 7. Serial cross sections of the ventral umbo of Hirnantia notiskuani n. sp. (AMNH-FI 137220) created using micro-CT scanning. Images are taken at a slightly oblique angle to avoid imaging artifacts created by pyrite within the enclosed matrix. Coloration as seen in Figure 6. (1–6) CT images of the ventral umbo reveal two separate chambers that are separated by a thin pedicle callist, which starts near the apex of the umbo and parts anteriorly. (7–12) Images as in (1–6), but with each chamber shaded.
Dorsal interior
Brachiophores moderately robust, wedge shaped, supported by high, straight, brachiophore plates that diverge from each other anterolaterally at ~80° (Fig. 3.8, 3.9). Sockets small, triangular, bounded anteromedially by brachiophores, open anterolaterally, with socket floor raised above the valve floor, supported by fulcral plates that are directly attached to the brachiophore plates (Fig. 3.9). Brachiophore plates sitting directly on valve floor, without forming delthyrial platform (Fig. 6.4–6.9). Cardinal process consisting of high, thin, plate-like shaft (Fig. 6.1–6.4) that extends anteriorly as low, rounded median ridge (Fig. 6.4–6.12); myophore delicate, with bilobate, crenulated top (Figs. 3.8, 4.7). Muscle scars poorly preserved in dorsal valves available for study.
Etymology
After the traditional Innu name for Anticosti, Notiskuan, which translates from the Innu-aimun language to “where bears are hunted.”
Materials
Conjoined shells (22) isolated from matrix plus seven embedded in slab; ventral valve (one); dorsal valves (two).
Remarks
Hirnantia notiskuani n. sp. is similar to the type species with regard to several external and internal characters. Externally, the gradual change in convexity throughout ontogeny, from a biconvex to dorsibiconvex profile, is consistent with the description of the type species by Temple (Reference Temple1965). Other external characteristics, including a punctate shell, fine multicostellate ornamentation, and an apsacline, slightly curved ventral interarea, are consistent with the description of the type species from several regions (e.g., Melou, Reference Melou1987; Villas et al., Reference Villas, Lorenzo and Gutiérrez-Marco1999; Hints et al., Reference Hints, Pärnaste and Gailite2012; Rong et al., Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b). Internally, the high, divergent brachiophore plates braced by fulcral plates, bilobed myophore, and high shaft of the cardinal process, which extends anteriorly as a broad median ridge, are also common features shared with the type species of Hirnantia (Amsden, Reference Amsden1974; Lespérance and Sheehan, Reference Lespérance and Sheehan1976; Melou, Reference Melou1987; Villas et al., Reference Villas, Lorenzo and Gutiérrez-Marco1999; Hints et al., Reference Hints, Pärnaste and Gailite2012; Benedetto, Reference Benedetto2013). The configuration (size and orientation) of the ventral interarea and the pedicle callist in the new species resemble most closely those of H. sagittifera from the Hirnantian Kuanyinchiao beds at the Hirnantian type section in Hubei, China (Fig. 9), although the pedicle callist was not reported specifically in previous studies of H. sagittifera from South China (e.g., Rong, Reference Rong1984; Rong et al., Reference Rong, Chen and Harper2002; Huang et al., Reference Huang, Zhou, Harper, Zhan, Zhang, Chen and Rong2020a, Reference Huang, Rong, Harper and Zhoub). A similar pedicle callist was described, but not illustrated, by Havlíček (Reference Havlíček1977, p. 267) for H. sagittifera from the Kosov (Hirnantian) strata of Bohemia. While Havlíček (Reference Havlíček1977) described several other species of Hirnantia from Katian (pre-Kosov) strata of Bohemia, most of those forms do not have the fine multicostellae or the subquadrate-outlined dorsal muscle field that are characteristics of the type species. The generic validity of the Bohemian species, therefore, needs further detailed investigations, especially as some of Havlíček's species occur in Katian strata of Anti-Atlas, Morocco (Mergl, Reference Mergl1983; Colmenar et al., Reference Colmenar, Villas, Rasmussen, Hunter, Álvaro, Lefebvre, van Roy and Zamora2018).

Figure 9. Silicon rubber cast of a ventral internal mold of Hirnantia sagittifera, specimen NIGP180325, Kuanyinchiao beds, GSSP section for the base of the Hirnantian Stage, Wangjiawan, Yichang district, Hubei, China. (1–4) Various enlarged views of the ventral interior; note the presence of pedicle callist, a thin transverse plate situated in the apical area, characteristic of the type species.
In contrast to the type species of Hirnantia, which has a rectimarginate to weakly unisulcate anterior commissure (Harper, Reference Harper and Kaesler2000), H. notiskuani is characterized by a uniplicate anterior commissure that is best developed in large and strongly convex (presumably gerontic) forms. However, this feature can be highly variable in both large forms (pronounced anterior plication to broad, shallow plication) and small to medium forms (broad, shallow plication to rectimarginate commissure; Fig. 8). A similar variation in anterior commissure has been observed in the orthoid genus Plaesiomys Hall and Clarke, Reference Hall and Clarke1892 from the same stratigraphic interval of the Ellis Bay Formation (Jin and Zhan, Reference Jin and Zhan2008). Plaesiomys anticostiensis (Shaler, Reference Shaler1865), for example, has a gently uniplicate anterior margin, whereas the type species, P. subquadrata (Hall, Reference Hall1847), has a rectimarginate anterior. Despite the great degree of morphological variation in both internal and external structures of the type species of Hirnantia (Temple, Reference Temple1965; Rong, Reference Rong1984; Villas et al., Reference Villas, Lorenzo and Gutiérrez-Marco1999; Hints et al., Reference Hints, Pärnaste and Gailite2012; Huang et al., Reference Huang, Rong, Harper and Zhou2020b; Rong et al., Reference Rong, Aung, Zhan, Huang, Harper, Chen, Zhou and Zhang2020b), H. notiskuani is the only species with a uniplicate anterior commissure. Juvenile specimens of H. notiskuani that lack a uniplicate anterior commissure can be distinguished externally from H. noixella Amsden, Reference Amsden1974 by their more evenly biconvex profile, in contrast to the ventribiconvex shell of H. noixella.

Figure 8. Shell dimensions of Hirnantia notiskuani n. sp. from the Prinsta Member of the Ellis Bay Formation at Ruisseau à la Batterie, Ruisseau Macaire, and Anse Mauvaise (Lousy Cove). Note that although there is variability in the anterior commissure, both small- and large-sized shells can possess a uniplicate commissure that is characteristic of the species.
Order Strophomenida Öpik, Reference Öpik1934
Superfamily Strophomenoidea King, Reference King1846
Family Strophomenidae King, Reference King1846
Genus Nasutimena Jin and Zhan, Reference Jin and Zhan2001
Type species
Strophomena fluctuosa Billings, Reference Billings1860, Vaureal Formation (Richmondian), Anticosti Island, Canada.
Nasutimena fluctuosa Billings, Reference Billings1860
Figure 10
- Reference Billings1860
Strophomena fluctuosa Billings, p. 57, fig. 6.
- Reference Billings1862
Strophomena fluctuosa; Billings, p. 123, figs. 102a, b.
- Reference Logan1863
Strophomena fluctuosa; Logan, p. 221, fig. 207A.
- Reference Hall and Clarke1892
Strophomena fluctuosa; Hall and Clarke, p. 251, pl. llA, figs. 4, 5.
- Reference Foerste1924
Strophomena fluctuosa; Foerste, p. 119, pl. 12, figs. 8a, b.
- Reference Twenhofel1928
Strophomena fluctuosa; Twenhofel, p. 193, pl. 22, figs. 3–5.
- Reference Troedsson1928
Strophomena fluctuosa; Troedsson, p. 93, pl. 22, fig. 13.
- Reference Macomber1970
Strophomena fluctuosa; Macomber, p. 440, pl. 78, figs. 16–24.
- Reference Bolton1972
Strophomena fluctuosa; Bolton, p. 22, pl. 1, fig. 9.
- Reference Bolton and Lesperance1981
Strophomena fluctuosa; Bolton, p. 50, pl. 3, fig. 4.
- Reference Dewing1995
Luhaia fluctuosa (Billings); Dewing, p. 139, pl. 10, figs. 2, 4–8, pl. 11, figs. 1–13.
- Reference Jin, Caldwell and Norford1997
Strophomena fluctuosa; Jin et al., p. 26, pl. 5, figs. 1–16, pl. 6, figs. 1–7.
- Reference Dewing1999
Gunnarella fluctuosa (Billings); Dewing, p. 20, pl. 6, figs. 2, 4–8, pl. 7, figs. 1–13.
- Reference Jin and Zhan2001
Nasutimena fluctuosa (Billings); Jin and Zhan, p. 31, pl. 6, figs. 9–19, pl. 7, figs. 5–12, pl. 20, figs. 1–7, pl. 21, figs. 5, 6, figs. 16, 17.

Figure 10. Nasutimena fluctuosa from the Prinsta Member of the Ellis Bay Formation, Hirnantian, at Ruisseau Macaire, eastern Anticosti Island. AMNH-FI 137239: (1) dorsal, (2) ventral, (3) lateral, (4) posterior, and (5) anterior views of a well-preserved shell, with enlargement of (6) the ventral posterior, displaying rugae, characteristic of the species, and (7) trail with well-developed parvicostellate ribbing.
Holotype
The original holotype of Strophomena fluctuosa collected by Billings (Reference Billings1860) from the Vaureal Formation (per Jin and Zhan, Reference Jin and Zhan2001) was misplaced before 1945 (Wilson, Reference Wilson1945, p. 142). However, a published hypotype (GSC 2017) from the Vaureal Formation, Carlton Point, Anticosti, was illustrated by Twenhofel (Reference Twenhofel1928, pl. 22, fig. 5) and subsequently by Jin et al. (Reference Jin, Caldwell and Norford1997, pl. 5, figs. 1–4).
Occurrence
All specimens are from the Prinsta Member of the Ellis Bay Formation (Late Ordovician) at Ruisseau Macaire and Ruisseau à la Batterie on eastern Anticosti Island, Québec, Canada.
Description
Shells medium sized, subtriangular in outline; resupinate in lateral profile with sharp ventrally directed geniculation (110–120°); trail high, with anteromedial dorsal fold and ventral sulcus (Fig. 10.1–10.5). Hinge line long, strophic, with rounded to alate cardinal extremities, commonly maximum width of specimen. Ribbing well defined, parvicostellate (Fig. 10.7). Concentric rugae variably developed on disc as either continuous (uninterrupted by costae) or discontinuous (interrupted by costae) wrinkles (Fig. 10.6). Concentric growth lines fine, strongly expressed on the trail. Pseudopunctae fine, densely spaced.
Ventral exterior
Valve weakly convex in umbonal area, becoming flat and concave anteriorly. Low planar interarea 2–3 mm high, weakly striated, slightly apsacline; delthyrium covered by thick, strongly convex pseudodeltidium (Fig. 10.4). Trail with anteriomedial fold to accommodate fold of dorsal valve.
Dorsal exterior
Valve flat to weakly concave in umbonal area, becoming convex anteriorly. but with distinctive cleft running down middle of disk (Fig. 10.1). Planar interarea approximately 1 mm high and anacline; notothyrium covered by thick, strongly convex chilidium. Protruding fold developed on trail medioanteriorly (Fig. 10.1, 10.5).
Ventral interior
Teeth robust and triangular, strongly divergent; dental plates extending anteriorly to form well-developed ridges that laterally bound deeply impressed subcircular to diamond-shaped muscle field, but often with 2–3 mm gap anterior of muscle field. Adductor scars lanceolate, divided by median ridge that tapers anteriorly, terminates just beyond anterior margin of muscle field. Diductor scars anteriorly divergent, enclosing adductor scars laterally. Peripheral rim well developed, starting laterally of muscle-bounding ridges and extending to line of geniculation where it curves anteriorly and continues along margin of the disc. Trail marked by well-developed radial thickenings of the shell, forming distinct ridges on interior and exterior of trail.
Dorsal interior
No dorsal interiors were recovered during collection.
Materials
Conjoined valves (6); ventral valves (5; often broken along anterior margin).
Remarks
These specimens are virtually identical to those described by Dewing (Reference Dewing1999) from the Vaureal Formation (Katian) on Anticosti Island and are assigned to the species on the basis of their distinctive subtriangular outline marked by an anteromedial fold, sharp ventrally directed geniculation, and variably developed rugae covering the disc (Jin and Zhan, Reference Jin and Zhan2001). However, in contrast to the description of Dewing (Reference Dewing1999), the teeth of the specimens described from the Ellis Bay Formation are more robust, a trait observed in specimens from southern Manitoba (Jin and Zhan, Reference Jin and Zhan2001). Specimens from the Ellis Bay Formation also have a prominent cleft running down the midline of the dorsal disk, a trait that has not been described in other specimens of this species.
Discussion
A Hirnantian age for the Ellis Bay Formation
The newly described occurrences of Hirnantia notiskuani in the sub-Laframboise Ellis Bay Formation provide a critical biostratigraphic constraint for understanding the Upper Ordovician succession on Anticosti Island. By expanding the stratigraphic range of the genus Hirnantia to the base of the Ellis Bay Formation, H. notiskuani provides compelling evidence for a Hirnantian age of the Ellis Bay Formation. A member of the cool-water “Kosov faunal province” in marginal and peri-Gondwana, Hirnantia was a cool-water, high-latitude genus that did not begin to expand its range to tropics until the onset of global cooling at the start of the Hirnantian Stage (Sheehan, Reference Sheehan2001; Rong et al., Reference Rong, Chen and Harper2002; Huang et al., Reference Huang, Zhou, Harper, Zhan, Zhang, Chen and Rong2020a). While several species of Hirnantia have been reported in Sandbian and Katian strata of high-latitude sites such as Bohemia (Havlíček, Reference Havlíček1977), these are represented mostly by relatively small, coarsely ribbed shells, unlike the typical shells of Hirnantia. So far, the type species and the new species from Anticosti Island, characterized by large, strongly dorsibiconvex shells with fine multicostellae, appear to be confined predominantly to the Hirnantian. Furthermore, in the rare instances where Hirnantia has been identified in lower-latitude, pre-Hirnantian strata, such as at the top of the Wufen Formation in Southwest China, the genus is found over a restricted stratigraphic interval only a few centimeters below an abrupt shift in facies and fossil assemblages that marks the base of the Hirnantian (Rong et al., Reference Rong, Chen and Harper2002). On Anticosti Island, the first occurrences of Hirnantia, represented by H. notiskuani, are found above major shifts in the stratigraphic and fossil records that mark the base of the Ellis Bay Formation. The differences in regional context between these patterns of first occurrences of Hirnantia provide greater confidence that the occurrences of H. notiskuani at the base of the Prinsta Member signify a Hirnantian age for the Ellis Bay Formation on Anticosti Island. These results are consistent with a growing number of palynological and chemostratigraphic studies that, in contrast to earlier studies (e.g., Brenchley et al., Reference Brenchley, Marshall, Carden, Robertson, Long, Meidla, Hints and Anderson1994; Bergström et al., Reference Bergström, Saltzman and Schmittz2006), suggest that the entire Ellis Bay Formation falls within the Hirnantian Stage (Delabroye et al., Reference Delabroye, Munnecke, Vecoli, Copper, Tribovillard, Joachimski, Desrochers and Servais2011; Achab et al., Reference Achab, Asselin, Desrochers and Riva2013; Mauviel and Desrochers, Reference Mauviel and Desrochers2016; Mauviel et al., Reference Mauviel, Sinnesael and Desrochers2020).
A diverse Hirnantia fauna
The new occurrences of H. notiskuani from the basal Ellis Bay Formation provide new context for assessing the biostratigraphic significance of the brachiopod assemblages throughout the Hirnantian on Anticosti Island. In contrast to other occurrences of Hirnantia, H. notiskuani does not frequently occur in direct association with other common members of the Hirnantia fauna (Jin and Zhan, Reference Jin and Zhan2008). Long and Copper (Reference Long and Copper1986) reported Hindella from the lower Prinsta Member at Anse Mauvaise, while subsequent collection by Jin and Zhan (Reference Jin and Zhan2008) yielded specimens of Eospirigerina. Common members of the Hirnantia fauna, including brachiopod genera such as Leptaena Dalman, Reference Dalman1828, Coolinia Bancroft, Reference Bancroft1949 (=Fardenia, Lamont, Reference Lamont1935; per Rong et al., Reference Rong, Huang, Zhan and Harper2013), Aphanomena Bergström, Reference Bergström1968 (=Eostropheodonta, Bancroft, Reference Bancroft1949; per Rong and Cocks, Reference Rong and Cocks1994), co-occur in a variety of different assemblages throughout the Ellis Bay Formation (see Jin and Copper, Reference Jin and Copper2008), either stratigraphically above or outside the geographic range of H. notiskuani (Jin and Copper, Reference Jin and Copper2008; Copper et al., Reference Copper, Jin and Desrochers2013), a pattern similar to the diachronous temporal and spatial distribution of the Hirnantia fauna observed in other Upper Ordovician sections (Rong et al., Reference Rong, Chen and Harper2002). These genera, inclusive of Hirnantia, range up to the top of the reefal Laframboise Member (Jin and Zhan, Reference Jin and Zhan2008; Copper et al., Reference Copper, Jin and Desrochers2013). On this basis, the brachiopod assemblages of the Ellis Bay Formation should be considered a part of the Hirnantia fauna.
Here we consider the H. notiskuani association of the basal Ellis Bay Formation on eastern Anticosti Island to represent a moderate-diversity Hirnantia fauna, associated with surviving brachiopod taxa from late Katian (Richmondian) epeiric seas of Laurentia, such as Hesperorthis Schuchert and Cooper, Reference Schuchert and Cooper1931, Gnamptorhynchos Jin, Reference Jin1989, Plaesiomys, Furcitella Cooper, Reference Cooper1956, Nasutimena, Hypsiptycha Wang, Reference Wang1949, and Rostricellula Ulrich and Cooper, Reference Ulrich and Cooper1942 (see Jin and Zhan, Reference Jin and Zhan2008). A similar assemblage, although lacking Hirnantia, has been observed in Hirnantian strata in southern Ontario (Stott and Jin, Reference Stott and Jin2007), as well as in marginal Laurentian Hirnantian strata in Girvan, Scotland (Harper, Reference Harper1981). Together, these assemblages show the presence of a unique Laurentian-affinity fauna composed of taxa typical of epeiric seas as well as the typical Hirnantia fauna. The H. sagittifera association in the Laframboise Member, however, represents a more “typical” Hirnantia fauna, associated with common Vinlandostrophia Zuykov and Harper, Reference Zuykov and Harper2007, Mendacella Cooper, Reference Cooper1930, Leptaena, Coolinia (=Fardenia), Aphanomena (=Eostropheodonta), Eospirigerina, and Hindella (Copper et al., Reference Copper, Jin and Desrochers2013). In this regard, if the entire Ellis Bay brachiopod fauna is to be treated as a Hirnantia fauna, it would be a highly diverse assemblage (see faunal list in Copper et al., Reference Copper, Jin and Desrochers2013). Finally, across the Ellis Bay/Becscie formational contact, the brachiopod assemblages abruptly transition to an assemblage with an Edgewood–Cathay affinity (sensu Rong et al., Reference Rong, Harper, Huang, Li, Zhang and Chen2020a), consisting of genera such as Koigia Modzalevskaya, Reference Modzalevskaya1985, Mendacella, Diceromyonia Wang, Reference Wang1949, Platystrophia King, Reference King1850, Leptaena, and Biparetis Amsden, Reference Amsden1974, and higher up in the Becscie Formation, Viridita Jin and Copper, Reference Jin and Copper2000 (Li and Copper, Reference Li and Copper2006). Biostratigraphy based on brachiopod assemblages would therefore ascribe an early to middle Hirnantian age to the entire Ellis Bay Formation, inclusive of the Laframboise Member, and a late Hirnantian age to the lowermost Becscie Formation. Given its potentially unique and well-preserved Hirnantian fossil and stratigraphic records, future studies of Upper Ordovician strata from Anticosti Island should focus on the incorporation of paleobiological and chemostratigraphic data from Anticosti Island into our global understanding of the sequence of climate (e.g., Finnegan et al., Reference Finnegan, Bergmann, Eiler, Jones, Fike, Eiseman, Hughes, Tripati and Fischer2011) and extinction (e.g., Copper et al., Reference Copper, Jin and Desrochers2013) events across the Ordovician/Silurian boundary.
Acknowledgments
J.B.Z. thanks the Anticosti-UNESCO Steering Committee for financial and logistical support. We thank A. Desrochers and P. Copper for invaluable field guidance, E. Bōgner and J. Tseng for their assistance in collecting and processing CT data, and T. Goldhawk for his assistance in producing the SEM images included in this study. J.B.Z.'s fieldwork is funded by the Marland Pratt Billings and Katharine Fowler-Billings Research Award from the Geological Society of America (grant no. 12320-19), the William B.N. Berry Fund from the University of California Museum of Paleontology, and the Lerner–Gray Memorial Fund of the American Museum of Natural History. J.J.'s work is funded by the Natural Sciences and Engineering Research Council of Canada. Collections were made under authorization issued by Ministère de l'environnement et de la Lutte contre les changements climatiques (MELCC) under the Natural Heritage Conservation Act (permit no. 7912-09-01-0000201; reference 402036698). The constructive comments of two journal reviewers helped greatly improve the clarity of our discussion.
Declaration of competing interests
The authors declare none.











 Open access
Open access