


Barley and wheat production trends
The Statistics Division of the Food and Agriculture Organisation of the United Nations collates data on the world production of various commodities, which can be downloaded for inspection( 1 ). The average yields in tonnes per hectare over time provide an overview of any perturbations in production for individual countries and the world total. These data show a reasonably steady increase in the average yield at a world level for barley and wheat from 1·41 and 1·31 t/ha, respectively, in 1961 to 2·96 and 3·16 t/ha, respectively, in 2012 (Fig. 1(a) and (b)). Average yields per hectare in Australia are less than the world average and also show more variation with frequent sharp reductions in the average yield per hectare that correspond to significant drought events in the country( 2 , 3 ). By contrast, average yields per hectare for the two crops in the UK are much higher than the world average, sometimes exceeding 8 t/ha for wheat. With few exceptions, e.g. 1976, UK production is not limited by drought. In fact too much water is more often a limiting factor with the drops in wheat production in 2001 and 2007 corresponding to rainfall above the 1961–1990 average in some of the major wheat growing areas( 4 ). When there is a marked increase in rainfall over most of the country as in 2012, the effect on wheat production can be severe, being over 1 t/ha less than the average of the five previous years.
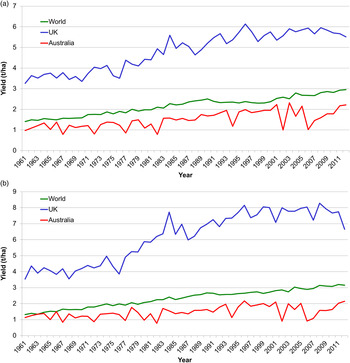
Fig. 1. (colour online) Trends in average yield in metric tonnes per hectare (t/ha) of barley (a) and wheat (b) for the Australian, UK and World crops from 1961 to 2012. Data obtained from FAOSTAT( 1 ).
Abiotic stress and grain composition
The UK Home Grown Cereals Authority conducts an annual survey of some cereal quality parameters and these are available online( 5 ). Among these data for wheat and barley, the fluctuations in mean grain nitrogen values for each year from 1977 to 2013 can provide some information on the influence of climate changes upon grain quality. The overall trend for wheat is an increase in grain nitrogen over the past 36 years. This trend conceals a pattern of 12 years with a protein content lower than 11 % in the period 1981–2000 whereas the protein content exceeded 12 % in 8 of the 13 years since 2000 (Fig. 2). While climate effects will have influenced these figures, the changes are more likely a reflection of a recent increase in the production of high protein wheat types for flour milling compared with a greater emphasis on high yielding feed wheat production in the previous period. By contrast, there has been a downward trend in the protein content of barley over the period (Fig. 2), which again reflects changes within the crop with spring barley forming a greater part of overall production and the concentration on production of low protein for use in malting within the spring crop. Nevertheless, there are some similarities in the responses of the two crops, notably the high production year of 2008 where both showed a marked drop in protein content.
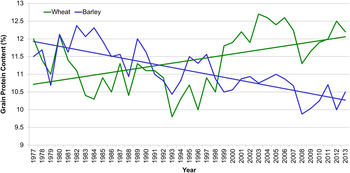
Fig. 2. (colour online) Trends in the grain protein content (% dm) of the UK barley and wheat crops from 1977 to 2013. Data available from the Home Grown Cereals Authority of the Agriculture and Horticulture Development Board( 5 ).
The relationship between yield and protein content is important for the barley and wheat crops as it can be critical for the market destination. Typically, the malting barley industry requires low protein content to maximise the yield of fermentable sugars whereas the wheat flour and baking industries require a high protein content to provide the right baking characteristics. While nitrogen fertilisers can provide an increase in grain protein content, protein content of both crops generally declines in a linear fashion as yield increases and much of this is the so-called yield dilution effect where the grains of higher yielding crops tend to have a larger starchy endosperm content and hence a lower protein content (Fig. 3). This effect can be modified to some degree by altering the timing of nitrogenous fertiliser applications when a late summer application to wheat can help ensure adequate protein for breadmaking( Reference Sylvester-Bradley, Blake and Kindred 6 ).

Fig. 3. (colour online) Relationship between grain yield per unit area (t/ha) and the grain nitrogen content (% dm) of a range of winter wheat varieties (individual points on the graph) grown with three different levels of nitrogen available to the plant over five site × season combinations. Data derived from the NUE Crops project( 28 ).
The effects of abiotic stress on dietary fibre contents of wheat and barley appear to be variable as one report suggests that β-glucan content of barley declines under temperature stress( Reference Savin, Stone and Nicolas 7 ) whereas another reports an increase in β-glucan content under drought stress( Reference Jansen, Schliephake and Kopahnke 8 ). While the stresses are different, one would have expected some consistency in response. Indeed, studies of three wheat cultivars under heat, drought and the combined effects of heat and drought found that all the three stress condition resulted in a decrease in grain β-glucan content coupled with a decrease in the ratio of short to long chain glucooligosaccharides( Reference Rakszegi, Lovegrove and Balla 9 ). The last study also found that arabinoxylan content increased with the three stress conditions, together with the protein content.
The quality of the barley crop is also liable to be affected by any drought stresses that may result from climate change. A study of the effect of an increase of 2·5°C in soil temperature on the spring barley variety Quench found that yield and starch content decreased with the increase in temperature. In contrast, the amounts of some proteins increased substantially, notably aspartate, glycine, alanine, arginine, valine and tryptophan( Reference Hoegy, Poll and Marhan 10 ). The effects of increased temperature on wheat grain properties were studied in more detail by applying a 4 h heat stress at 18 d after anthesis. The study sampled grains from control and stress treatments at four developmental stages from the beginning of the stress treatment up to physiological maturity and found that the heat stress reduced grain mass by 23 %. The heat stress also reduced the amount of nitrogen in each grain by 16 % but, because this was less than the reduction in grain mass, the net effect was an increase in nitrogen concentration in the grain. Interestingly, there was a significant increase in the amount of protein in a grain shortly after the stress was removed but the control treatment appeared to continue to accumulate protein up to maturity whereas the stress treatment did not( Reference Majoul-Haddad, Bancel and Martre 11 ). The authors also studied the effects on different wheat protein components and found that the responses of the gliadin and glutenin fractions were very similar to those of the total protein. By contrast, there were no significant differences between the control and stress treatments for the albumin/globulin and amphilipid fractions for the three pre-maturity sampling stages with a noticeable decline in the amount present in each grain at maturity compared with the third sampling stage.
A review of gene expression studies on the effects of heat stress on key genes involved in starch metabolism found that nearly all were down-regulated in barley with just two (a starch synthase and a β-amylase) being up regulated. While fewer genes had been studied in wheat, all were down-regulated in the heat stress experiments surveyed. Rice also showed a general pattern of down regulation of starch synthesis under heat stress( Reference Thitisaksakul, Jimenez and Arias 12 ). These findings are consistent with the reduction in grain mass as found in the wheat study aforementioned( Reference Majoul-Haddad, Bancel and Martre 11 ) and indicate that the effect on starch synthesis is more marked than on protein deposition in the grain. The gene expression review did note that the various heat stress experiments had identified a trend to increased amylose content together with a greater proportion of A granules of starch and thus an overall effect on starch quality( Reference Thitisaksakul, Jimenez and Arias 12 ).
Clearly, climate change is going to affect barley quality as well as yield, which will in turn affect the efficiency of major end-users such as maltsters, brewers and distillers. This in turn may affect the sourcing of their crop with a possible trend to buy from more distant markets in times when the quality of the local market has been adversely affected by climate change effects. This could then lead to a reduction in barley area in the local area and hence a withdrawal from barley breeding by the private sector in some areas. Escape mechanisms such as a switch to winter malting barley is an option in some traditional spring barley growing areas but it is also essential that sources of drought tolerance that can readily be incorporated into elite germplasm are identified and a public–good breeding effort is set up to facilitate the development of drought tolerant barley germplasm.
Breeding for improved abiotic stress tolerance
Currently breeders feel that the effects of climate change will be reflected in yield and therefore continuing to select for high yield, whether phenotypically or genotypically, will continue to produce yield gains. This strategy presumes that climate change effects are effectively incremental changes in the same direction and some pragmatists suggest that moving germplasm from lower latitudes to higher latitudes may well provide the right breeding material for the higher latitudes. A study of barley production in Finland under a range of climate change models suggested that production in such a Northern region is currently limited by the short growing season due to delayed sowing to avoid cold damage so the extended growing season that a warmer climate in such regions would mean an increase in production under all but one of the scenarios evaluated, although the increase was only predicted for the more favourable soil types( Reference Rotter, Hohn and Trnka 13 ). Moving germplasm to higher latitudes does not solve the problem for the lower latitudes and may result in market failure for these regions. The increased daylength at the higher latitude can be expected to interact with the typical flowering time genes found in lower latitude germplasm and this may well result in a completely different growth pattern that may not provide a sustainable escape from drought. It is important to remember that Mediterranean varieties effectively escape drought rather than tolerate it. Indeed, evidence from a study of four genotypes suggests that wheat varieties from more Mediterranean regions are not inherently more drought tolerant than those from the UK but that they are earlier and more likely to escape severe effects( Reference Semenov, Stratonovitch and Alghabari 14 ).
The longer term prospects of breeding for drought tolerance were assessed through consultation with a panel of experts from a range of European countries ranging from Scandinavia to the Mediterranean. Experts were asked to rate the current climatic challenges to crop production in each region for different crops and then what would be the best adaptation strategies to meet various climatic scenarios. For spring barley, the threats most consistently identified as causing problems were rain at sowing and drought with the former being more important in Northern regions and rarely a problem in Southern ones whereas drought problems were only considered rare around the French and Iberian Atlantic coasts. Curiously, introduction of ‘climate-proof’ spring barley cultivars was considered to be of minor importance as an adaptation strategy in three of the twelve climatic regions with none considering such a development to be of more than moderate importance. Measures such as monitoring of pests and diseases and seasonal agrometeorological forecasts were, however, considered to be of major importance( Reference Olesen, Trnka and Kersebaum 15 ).
The Focused Identification of Germplasm Strategy , in which mathematical and statistical approaches have been developed over a number of years to combine environmental and Geographic Information Systems data to predict regions that are most likely to contain germplasm with specific attributes e.g. tolerance to mid-season drought stress, and thus select appropriate subsets from germplasm collections. For example, this approach was used to screen over 17 000 accessions of wheat in germplasm collections to identify a subset of 510 for field evaluation of resistance to Russian wheat aphid, a reduction of 97 % in testing. From this pool, twelve promising new sources of resistance to the pest were identified( Reference El Bouhssini, Street and Amri 16 ). This approach could also be adopted to search and test for sources of drought tolerance and has been applied to identify drought tolerance characters such as canopy temperature and relative water content in faba bean( Reference Khazaei, Street and Bari 17 ).
Strategies for improving drought tolerance
Until the advent of DNA fingerprinting techniques, plant breeders relied on direct observations of characters, phenotypic selection, to select improved plant varieties and release them to growers. By combining DNA fingerprints of different genotypes of a crop with phenotypic measurements of characters, researchers have been able to associate specific chromosomal regions, called quantitative trait loci (QTL), with the control of those characters. In doing so, they have identified specific DNA markers that can be used to select for favourable alleles of the QTL, a technique known as marker assisted selection. The advantages of using marker assisted selection are that the markers can be deployed at any stage in the breeding programme and they are not subject to the interactions with the environment that direct phenotypic measurements can be. Marker assisted selection can therefore be used to select a pool of germplasm that has a high genetic potential for key target characters, an attribute that is especially useful when applied to characters that show very marked interactions with the environment, such as drought tolerance.
Much research effort has therefore been directed at the detection of QTL and searching the bibliographic database Web of Science with the key words QTL and Map showed that over 38 000 scientific refereed papers on the topic have been produced between 1990 and 2012 inclusive. Much of this work was not, however, taken up by the plant breeding community as, until recently, the studies were not conducted on the typical elite germplasm being used by breeders or the molecular marker density was insufficient to produce a tightly linked marker to a QTL that could be used reliably in selection. The development of high density marker assays in the mid 2000s coupled with developments in association mapping methodologies has meant that these problems have largely been solved so that markers can be found that are closely linked to QTL. Moreover, developments in genome sequencing mean that the gene content of key crops such as barley and wheat are now available, which facilitate the identification of the underlying genes controlling the characters, offering the prospect of identifying ‘perfect markers’ that target the functional sequence polymorphism that contributes to a high or a low expression of a character. The limiting factor is now the ability to generate high quality phenotypic data on the populations.
A review examined a range of possibilities to improve the water use efficiency and hence the drought tolerance of cereal crops. The characters ranged from establishment characteristics, including rooting behaviour, through physiological mechanisms such as transpiration efficiency to leaf morphological characters( Reference Richards, Rebetzke and Watt 18 ). All of these characters have been the subject of QTL mapping programmes and markers linked to most traits had been identified, offering the prospect of marker-assisted selection to improve water use efficiency, but the authors concluded that phenotypic selection remained the most efficient way of improving these characters. This group had focused on the application of carbon isotope discrimination as a surrogate measure for transpiration efficiency and had applied it in a breeding programme to develop two new Australian wheat cultivars that had markedly improved water use efficiency. Drysdale and Rees were released in the early 2000s and were recommended for growing in low rainfall areas due to their relatively better performance under such situations.
Such an outcome suggests that improvements in drought tolerance are possible and other variants such as the stay-green phenotype could also be deployed. Mutation of a wheat genotype was used to derive a line with delayed senescence of leaves, the stay-green phenotype. The mutant had a slower degradation of chlorophyll and appeared to have a greater stability of the thylakoid membrane under drought stress conditions, all of which led to a delayed senescence under drought treatment( Reference Tian, Gong and Zhang 19 ). While such findings are promising, the authors do not report any associated effects on yield, although studies on the performance of sorghum hybrids under drought stress indicated a significant yield advantage for those with the stay-green phenotype( Reference Borrell, Hammer and Henzell 20 ).
There are many reports in the scientific literature of transgenic approaches to improve drought tolerance. Dehydrins have been reported as important factors in stress tolerance( Reference Campbell and Close 21 ) and the genetic locations of some dehydrin family members co-locate with QTL for drought tolerance characters in barley( Reference Teulat, Zoumarou-Wallis and Rotter 22 ). Transcription factors such as the dehydration responsive element binding (DREB) factor also play a role in plant abiotic stress tolerance( Reference Agarwal, Agarwal and Reddy 23 ) and over-expression of some DREB genes produced improved survival under drought stress under the control of a constitutive promoter. This combination did, however, produce slower growing and lower yielding plants than the non-transformed controls, although this effect could be modified by using a drought inducible promoter to control the DREB factors under study( Reference Morran, Eini and Pyvovarenko 24 ). The same study also found that there was no difference in the plant yields of the control and drought-inducible DREB transgenic plants that survived the drought stress but that more of the transgenes did survive the stress and thus should provide a greater crop yield under stress conditions. The restrictions on growing transgenic plants in the field generally prevent a true test of the effectiveness of transgenic approaches to improve drought tolerance. To test this, another study selected transgenic DREB1A lines in the glasshouse on the basis of (1) improved survival and recovery after drought and (2) improved water use efficiency characteristics. While there was no difference between the yields under drought of the combined transgenic selection groups and controls, those selected for water use efficiency had an acceptable yield when water was not limiting but also had a stable performance across a range of different water regimes( Reference Saint Pierre, Crossa and Bonnett 25 ).
There are therefore a number of strategies that could be deployed to improve the drought tolerance of wheat and barley. Highly controlled experimental conditions with plants moving on conveyor belts (so-called phenomics platforms( Reference Furbank and Tester 26 )) could be deployed to screen out the most promising and then these could be more fully evaluated under a range of different field conditions to pick the best( Reference Munns, James and Sirault 27 ).
Conclusions
The threat of increased drought events poses not only a risk to the amount of crop produced but also its quality with reductions in the amount of carbohydrate and protein contained in each grain. Carbohydrate production in the grain is, however, more affected than protein production with the result that the protein concentration in the grain increases. While an increase in grain protein percentage might make the wheat crop more suitable for milling, the smaller grain would reduce the yield potential and increase the land area required for milling wheat and there may well be a reduction in flour yield (the proportion of the grain that is milled into flour) that would result in a further increase in the land area required for milling wheat. The effects of drought on soluble fibre compounds is varied with β-glucan decreasing and arabinoxlyan increasing but the balance of these factors and the consequent effect on the digestive system remains to be established. Breeding for improved drought tolerance in terms of yield is possible but the associated effects on grain quality have yet to be fully established. A multi-disciplinary research effort is needed to integrate genotypic, phenotypic, proteomic, physiological, transcriptomic and epigenetic data to understand the networks that lead to various quality attributes and then form strategies to make crops more resilient in the face of climate change.
Acknowledgements
The author acknowledges the support and contributions made by KWS Lochow and KWS (UK) for the associated text and Fig. 3 that was derived from the work conducted under a work package of the EU FP7 Improving nutrient use efficiency in major European food, feed and biofuel crops to reduce negative environmental impacts of crop production (NUE Crops): EU-FP7 222-645 (2009–2014) led by Carlo Leifert of the University of Newcastle.
Financial Support
The author thanks the Scottish Government's Rural and Environment Science and Analytical Services Division for funding.
Conflicts of Interest
None.
Authorship
W. T. B. Thomas reviewed the literature and wrote the manuscript.



