



Introduction
In recent decades, warming temperatures and a lengthened fire season have led to increased fire activity in the western US (Westerling et al., Reference Westerling, Hidalgo, Cayan and Swetnam2006). Annual area burned and fire size are expected to increase with continued warming (Westerling et al., Reference Westerling, Turner, Smithwick, Romme and Ryan2011). Higher elevations have disproportionately felt the effects of warming temperatures (Mountain Research Initiative EDW Working Group, 2015; Pepin et al., Reference Pepin, Arnone, Gobiet, Haslinger, Kotlarski, Notarnicola and Palazzi2022), and fires have increasingly advanced upslope in recent decades (Alizadeh et al., Reference Alizadeh, Abatzoglou, Luce, Adamowski, Farid and Sadegh2021, Reference Alizadeh, Abatzoglou, Adamowski, Modaresi Rad, AghaKouchak, Pausata and Sadegh2023). Subalpine forests of the Rocky Mountains have been burning more than ever in recent history (Higuera et al., Reference Higuera, Shuman and Wolf2021), particularly due to factors such as earlier spring snowmelt (Westerling, Reference Westerling2016; Alizadeh et al., Reference Alizadeh, Abatzoglou, Adamowski, Modaresi Rad, AghaKouchak, Pausata and Sadegh2023). To contextualize recent fire trends, fire records from subalpine forests are necessary to understand how fire activity responded to past periods of warm temperatures.
Within the Rocky Mountains, the Greater Yellowstone Ecosystem (GYE) measures approximately 22 million acres and is one of the world’s largest, nearly intact temperate ecosystems (Marcus et al., Reference Marcus, Meacham, Rodman, Steingisser, Menke and West2022). The environmental history of the GYE is among the best studied in the U.S., with records from all elevations (Whitlock, Reference Whitlock1993; Fall et al., Reference Fall, Davis and Zielinski1995; Whitlock et al., Reference Whitlock, Bartlein and Van Norman1995, Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012; Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000; Huerta et al., Reference Huerta, Whitlock and Yale2009; Mumma et al., Reference Mumma, Whitlock and Pierce2012; Krause and Whitlock, Reference Krause and Whitlock2013, Reference Krause and Whitlock2017; Iglesias et al., Reference Iglesias, Krause and Whitlock2015, Reference Iglesias, Whitlock, Krause and Baker2018; Rust and Minckley, Reference Rust and Minckley2020). Pollen and charcoal analyses from these sites indicate that the vegetation and fire history generally track millennial-scale climate variations. Most of these studies have focused on the subalpine forest history in the central and southern GYE, while sites in the northern GYE are in low-elevation steppe and xeric forests (Huerta et al., Reference Huerta, Whitlock and Yale2009; Mumma et al., Reference Mumma, Whitlock and Pierce2012; Krause and Whitlock, Reference Krause and Whitlock2013; Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015), other than one site in a high-elevation subalpine forest in the Bridger Range (Benes et al., Reference Benes, Iglesias and Whitlock2019).
The vegetation and fire histories of mixed-conifer forests in the northern GYE are not well known in the absence of paleoecological records from middle elevations. This study provides a 13,500-year-long record from Langohr Wetland (45.52425°N, 111.01807°W, 1898 m asl, ∼2 ha), located in a mixed-conifer forest in the northern Gallatin Range adjacent to the Gallatin Valley and the city of Bozeman. The primary objective of this study was to reconstruct the long-term climate–fire–vegetation interactions at Langohr Wetland and compare its vegetation and fire history with sites in other ecological settings in the region.
Study site
The Gallatin Range spans ∼120 km north to south and averages 32 km in width, hosting several peaks above 3000 m asl, the highest of which is Electric Peak (3433 m) in Yellowstone National Park (YNP). Present-day vegetation in the region varies with elevation and aspect. Artemisia (sagebrush) steppe and grassland (mainly Festuca idahoensis, Poaceae spp.) cover valley floors, and in the northern Gallatin Range (closest to Langohr Wetland), lower treeline is at approximately 1650 m asl. Juniperus scopulorum (Rocky Mountain juniper), Pinus flexilis (limber pine), and Pseudotsuga menziesii (Douglas fir) constitute low-elevation dry forests from ∼1650 to 1800 m asl, and forest density increases with elevation. Subalpine conifer forests of Picea engelmannii (Engelmann spruce), Abies lasiocarpa (subalpine fir), and Pinus contorta (lodgepole pine) occur between 1800–2100 m asl, with A. lasiocarpa increasing in abundance with elevation. Forests dominated by Picea engelmannii and A. lasiocarpa are generally above ∼2100 m asl, and Pinus albicaulis (whitebark pine) forests are found at the highest elevations above 2400 m asl. Alpine tundra is found on the highest peaks, above ∼3000 m asl (Arno, Reference Arno1979; Whitlock, Reference Whitlock1993; Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018).
Langohr Wetland is in the Custer Gallatin National Forest in a glacially formed depression (Weber, Reference Weber1965) within the Hyalite Creek watershed. It has a small inlet spring entering from the south and flowing to the northeast (Fig. 1). The wetland supports Carex spp., Eleocharis spp. (sedges), and Poaceae spp. (grasses). Salix spp. (willow) and Betula spp. (birch) shrubs grow along the margins. A closed mixed-conifer forest of Picea engelmannii, Pinus contorta, and lesser amounts of A. lasiocarpa surrounds the wetland. Pseudotsuga menziesii and J. scopulorum grow on nearby south-facing slopes, and Pinus albicaulis dominates at higher elevations in the watershed. Juniperus communis (common Juniper) is found in the forest understory along with other shrubs, including species of Rosaceae (i.e., Rosa woodsi (Woods’ rose), Physocarpus malvaceus (ninebark), Potentilla gracilis (cinquefoil), Amelanchier alnifolia (serviceberry), and Spiraea betulifolia (birch leaf spirea)), Symphoricarpos albus (snowberry), Berberis repens (creeping mahonia), Shepherdia canadensis (buffaloberry), and Arctostaphylos uva-ursi (bearberry). The adjacent grassy, west-facing hillslope supports Juniperus horizontalis (creeping juniper) and a diverse array of herbs from the Asteraceae, Rosaceae, Amaranthaceae, Brassicaceae, Ranunculaceae, and Caryophyllaceae. Artemisia tridentata shrubs are present in a meadow ∼0.25 km NW of the wetland, and Alnus viridis (green alder), Populus tremuloides (quaking aspen), and Populus balsamifera (black cottonwood) grow on the wet banks of Hyalite Creek ∼0.5 km NW of the wetland.
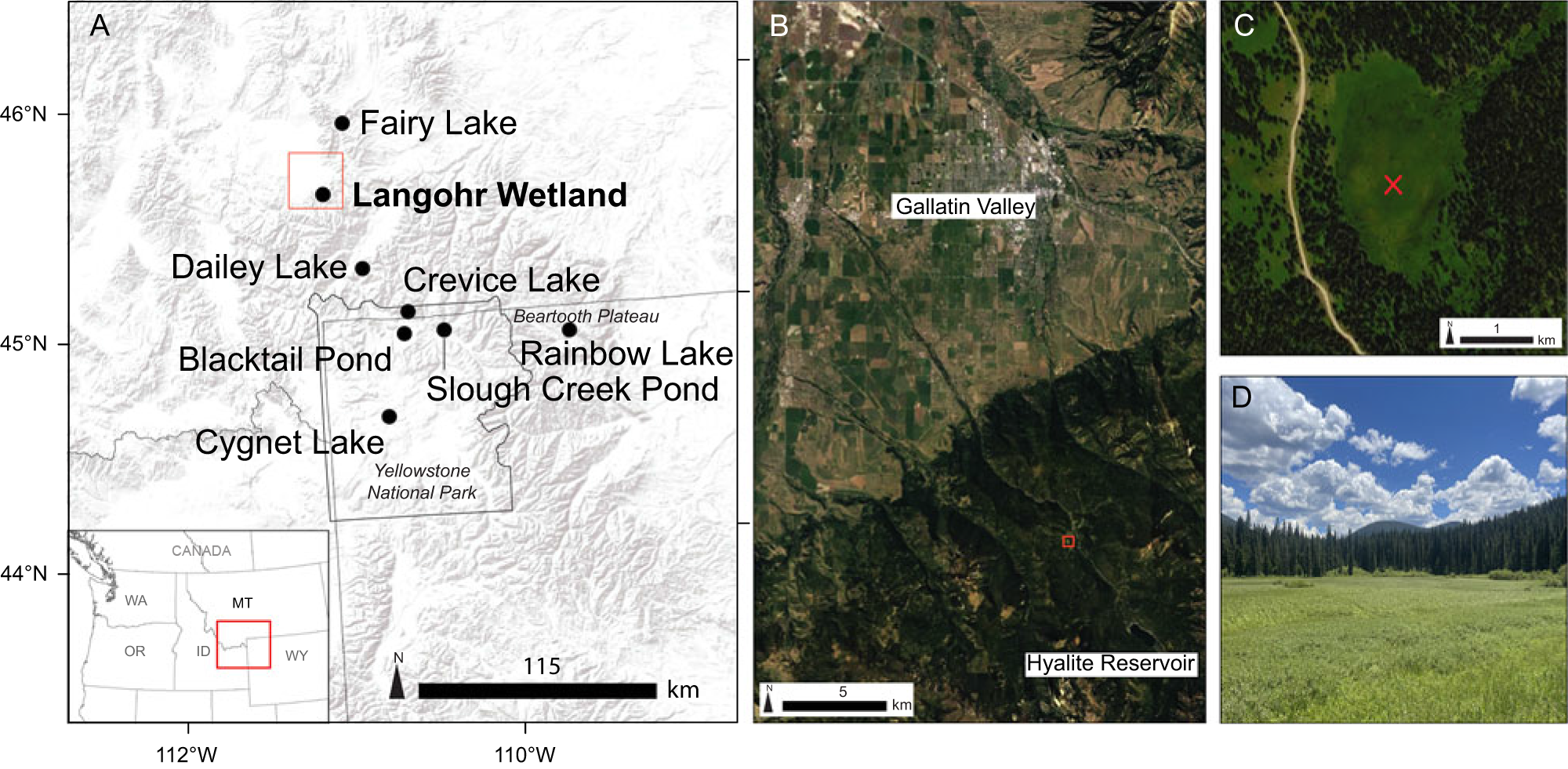
Figure 1. (A) Regional map showing Langohr Wetland and other study sites discussed in the text. (B) Location of Langohr Wetland (red square) in relation to the Gallatin Valley. (C) Coring location (red X) in Langohr Wetland. (D) Langohr Wetland.
Extensive logging in the Hyalite Creek watershed started in the late 1800s and primarily targeted large merchantable Pseudotsuga menziesii trees. Post-logging forest openings have since been colonized by Pinus contorta and Pseudotsuga menziesii. Secondary forests tend to be dense and have a homogeneous structure, similar to other logged forests in Montana (Naficy et al., Reference Naficy, Sala, Keeling, Graham and Deluca2010).
Large fires (> 400 ha) have occurred historically throughout the Custer Gallatin National Forest between the mid-1500s and early 1900s (Barrett et al., Reference Barrett, Arno and Menakis1997). Large fires were infrequent between 1940 and 1980 due to a combination of cooler temperatures (Littell et al., Reference Littell, McKenzie, Peterson and Westerling2009) and intensified fire-suppression efforts (Agee, Reference Agee1993; Barrett, Reference Barrett1993; Erickson, Reference Erickson2022). In recent decades, fire activity in the western U.S. has increased due to rising temperatures and drought (Abatzoglou and Williams, Reference Abatzoglou and Williams2016). Contrary to this trend, the total acreage burned in the Custer Gallatin National Forest has decreased since the large Yellowstone fires of 1988, and xeric forests that historically experienced high- (0- to 35-yr fire return interval) and moderate- (35–100+ yr fire return interval) frequency fire regimes are assumed to have a fire deficit at present (Erickson, Reference Erickson2022).
Wet Picea–Pinus forests, like those surrounding Langohr Wetland, currently experience a high-severity fire regime with a 35–200+ year return interval. The south-facing Pseudotsuga forests experience a higher frequency mixed/low-severity fire regime (every 35–100+ years) (LANDFIRE, 2014; Erickson, Reference Erickson2022). No major fires have occurred in the Hyalite Creek watershed in the last 100 years (Erickson, Reference Erickson2010), although small, lightning-caused fires were immediately suppressed in 2013 (Horsetail fire) and 2014 (Sleeping Giant and Lick Creek fires). In the Gallatin Range as a whole, large fires last occurred in 2001 (Fridley and Purdy fires) and 2012 (Millie Creek fire; MTBS, 2021). Because Picea–Pinus forests have a relatively long natural fire return interval, fire-suppression efforts of the last 100 years have likely had minimal effect on the forest around Langohr Wetland (Schoennagel et al., Reference Schoennagel, Veblen and Romme2004).
Interpolated climate data based on 30-year mean monthly temperature and precipitation data (PRISM Climate Group, 2022) indicate that mean summer (July–August) temperature at Langohr Wetland is 14°C, with a maximum of 25.6°C, and mean winter temperature (December–February) is −5.6°C, with a minimum of −12°C. Langohr Wetland receives an average of 720 mm annual precipitation, most of which is received in late spring to early summer (April–June = 280 mm of precipitation on average). Late summer and early fall months are driest (July–September = 120 mm on average), and ∼35% of total precipitation falls as snow (November–March = 250 mm on average). The ratio of summer (June–August) precipitation to winter (December–February) precipitation is ∼1.20, suggesting the high amount of June precipitation causes it to receive slightly more precipitation in the early summer than in the winter.
Methods
Field and laboratory
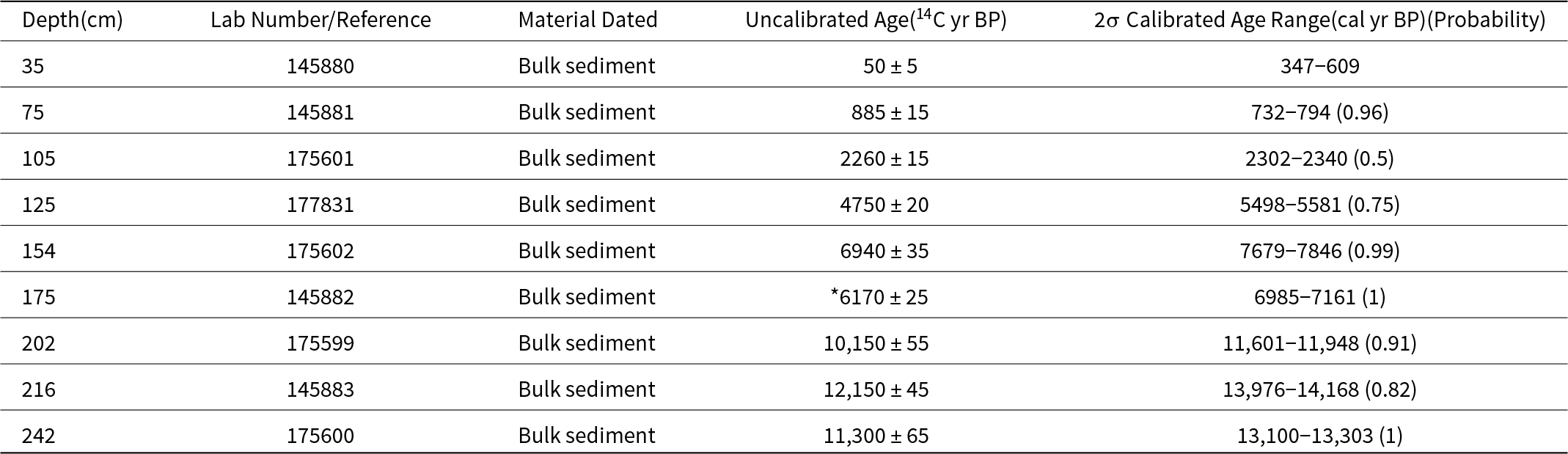
A 250-cm-long sediment core was collected from Langohr Wetland using a Russian D-section Peat Borer. Cores were taken at continuous 50-cm-long intervals until the borer hit bedrock. Sediment cores were wrapped in plastic and aluminum foil in the field and stored in the cold room at Montana State University until they were analyzed. In the laboratory, core lithology was described by sediment type (Schnurrenberger et al., Reference Schnurrenberger, Russell and Kelts2003) and subsampled. Nine bulk sediment samples were taken throughout the core and submitted to the National Ocean Sciences Accelerator Mass Spectrometry facility at Woods Hole Oceanographic Institution for accelerator mass spectrometry (AMS) radiocarbon dating (Table 1). AMS C14 dates were calibrated with the IntCal20 calibration curve (Reimer et al., Reference Reimer, Austin, Bard, Bayliss, Blackwell, Ramsey and Butzin2020), and an age-depth model was created with BACON age-depth modeling software, which uses a Bayesian statistical approach (Blaauw and Christen, Reference Blaauw and Christen2011).
Table 1. Radiocarbon information for Langohr Wetland; *denotes a rejected date
Charcoal analysis
To reconstruct the local fire history, samples that were 2 cm3 in volume were taken from contiguous 1-cm intervals of the entire core for charcoal analysis. Samples were processed following standard methods and gently washed through a 125-μm sieve (Whitlock and Larsen, Reference Whitlock, Larsen, Smol, Birks, Last, Bradley and Alverson2001). The residue was examined under a stereomicroscope, and charcoal particles were counted at 50–100× magnification. Charcoal counts were analyzed with CharAnalysis software (Higuera et al., Reference Higuera, Brubaker, Anderson, Hu and Brown2009). Charcoal concentrations (particles/cm3) were binned into 47-yr time intervals (median sampling interval) to minimize differences in charcoal accumulation due to changes in sedimentation rate, and charcoal accumulation rates (CHAR, particles/cm2/yr) were calculated by dividing the charcoal concentration by the sediment accumulation rate (yr/cm). The charcoal record was separated into a background component (background CHAR = BCHAR) and a higher frequency peaks component. BCHAR was calculated using a 600-yr LOWESS smoother robust to outliers, which optimized the signal-to-noise ratio (SNI > 3; Kelly et al., Reference Kelly, Higuera, Barrett and Hu2011). A Poisson minimum count test identified charcoal peaks that exceeded a 90% threshold as significant fire episodes.
Charcoal data register local fire episodes, one or more fires within 3 km of the site that occurred during the time span of the peak (Whitlock and Larsen, Reference Whitlock, Larsen, Smol, Birks, Last, Bradley and Alverson2001; Higuera et al., Reference Higuera, Brubaker, Anderson, Hu and Brown2009). Charcoal peaks are usually a record of stand-replacement fires rather than surface fires, which do not produce much charcoal and carry lesser amounts aloft. BCHAR is considered either (1) a representation of regional biomass burned, (2) a result of secondary charcoal deposition and sediment mixing (Whitlock and Larsen, Reference Whitlock, Larsen, Smol, Birks, Last, Bradley and Alverson2001), and/or (3) a measure of arboreal fuel biomass on the landscape (Marlon et al., Reference Marlon, Bartlein and Whitlock2006). Peak magnitude measures the total charcoal deposition per fire event (particles/cm2/peak) and can potentially measure changes in fuel consumption or fire size (Higuera et al., Reference Higuera, Brubaker, Anderson, Hu and Brown2009). Fire-episode frequency was determined using a smoothed 1000-year moving window, and the mean fire-episode return interval is the years between significant fire episodes.
To analyze regional charcoal trends, a composite charcoal record was created with records from nearby sites in the GYE that extended through the late-glacial period. Published age-depth models and counts were first analyzed in CharAnalysis with the same parameters as the Langohr record analysis, except that charcoal peaks that crossed a 95% threshold were considered significant fire episodes. To standardize the data for the composite record, CHAR results were subjected to statistical minmax, Box–Cox, and finally Z-score transformations as described in Power et al. (Reference Power, Marlon, Ortiz, Bartlein, Harrison, Mayle and Ballouche2008).
Pollen
One-cm3-volume samples were processed for pollen following the procedures of Bennett and Willis (Reference Bennett, Willis, Smol, Birks and Last2001). A Lycopodium tablet of known concentration was added to each sample to calculate pollen concentration (grains/cm3) and pollen accumulation rates (PAR; grains/cm2/yr). A minimum of 300 terrestrial pollen grains were counted per sample at 400× and 1000× magnification. Pollen reference slides at Montana State University and pollen keys (McAndrews et al., Reference McAndrews, Berti and Norris1973; Faegri and Iversen, Reference Faegri and Iversen1989; Kapp et al., Reference Kapp, Davis and King2000) were used to identify the grains. Terrestrial pollen counts were converted to percentages based on the terrestrial pollen sum. Aquatic pollen counts were converted to percentages with the combined terrestrial and aquatic pollen sum. Carex spp. is a dominant Langohr Wetland plant; accordingly, Cyperaceae is considered an aquatic taxon in this study.
Based on the conifer species present in our study region, Pinus subg. Strobus pollen was attributed to Pinus flexilis or Pinus albicaulis and Pinus subg. Pinus pollen was ascribed to Pinus contorta. Damaged Pinus grains that were missing distal membranes were classified as Pinus undifferentiated. Abies grains were attributed to A. lasiocarpa, Picea grains to Picea engelmannii, and Juniperus-type pollen to J. communis or J. scopulorum. Species in the “Total Rosaceae” category include undifferentiated Rosaceae and Spiraea, Potentilla, and Amelanchier-type pollen grains. Shrubs and herbs under 1% were grouped in an “other shrubs and herbs” category. Non-arboreal pollen amounts are higher in non-forested landscapes and increase with distance from forest boundaries (Whitlock, Reference Whitlock1993; Fall, Reference Fall1994; Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018), and the ratio between arboreal pollen (AP) and non-arboreal pollen (NAP) was used as an indicator of forest cover. Arboreal pollen is the sum of all tree and shrub species, excluding Artemisia, since Artemisia is predominantly found in open landscapes and forest meadows. Non-arboreal pollen is the sum of Artemisia, herbs, and grasses. The AP:NAP ratio was calculated by (AP−NAP)/(AP+NAP). CONISS was used to identify pollen zones through stratigraphically constrained cluster analysis (Grimm, Reference Grimm1987).
Results
Lithology
The Langohr core consisted of four lithological units (Fig. 2). Unit 1 (250–240 cm depth) was inorganic gray clay and sand. Unit 2 (240–175 cm depth) was dark brown fragmental granular peat. Unit 3 (175–105 cm) was primarily fine-detritus sapropel and contained two layers with increased levels of inorganic silt and clay (silty-clay sapropel), one at 150–138 cm and one at 120–115 cm depth. Unit 4 (105 cm depth–surface) consisted of fragmental sedge herbaceous peat that increased in coarseness towards the top.
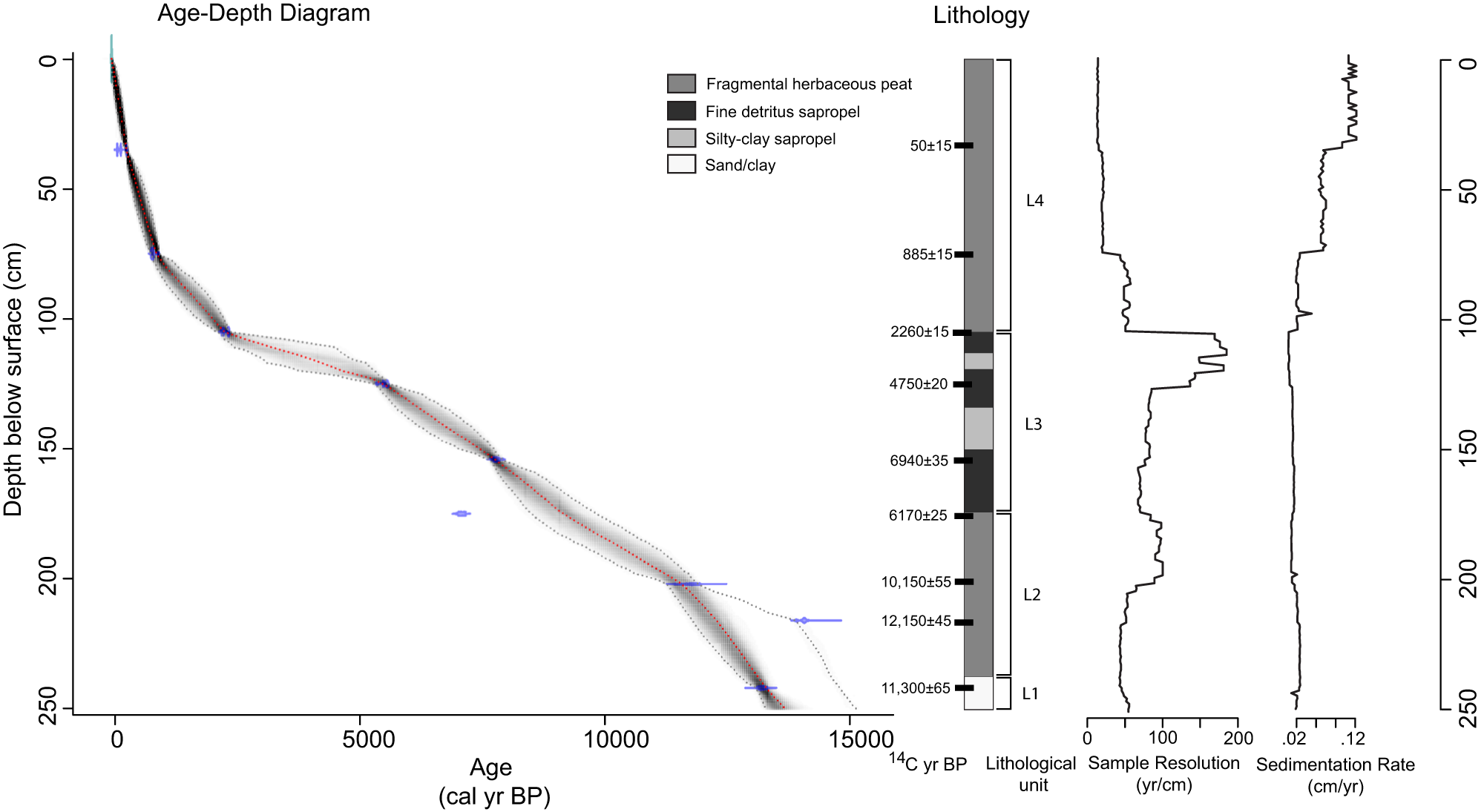
Figure 2. Age-depth diagram and lithology of Langohr Wetland core. The age-depth diagram is based on nine radiocarbon dates. The red dotted line is the median probability age for each depth, and the black dotted lines indicate the 95% probability range. The lithology includes sediment types, sample depths for radiocarbon dates, and lithologic units discussed in the text.
Chronology
The Langohr age-depth chronology was based on nine AMS 14C dates from bulk sediment (Table 1, Fig. 2). Extrapolated ages from the bottom two dates suggest sediment accumulation began between ca. 14,000 and 13,000 cal yr BP. One age reversal was noted at the base of the core. The age of 14,168–13,976 cal yr BP at 216 cm depth may be anomalously old due to incorporating ancient carbon in the sediments. However, the date at 242 cm depth could be anomalously young, therefore, the basal age primarily influenced by the 13,300–13,100 cal yr BP date at 242 cm depth is considered a minimum age. An anomalously young age at 175 cm depth was rejected in the age-depth model.
The sedimentation rate in units 1 and 2 averaged 0.02 cm/yr. Unit 3 had the slowest sedimentation rate of 0.01 cm/yr average. The low sedimentation rate in unit 3 may suggest undetected hiatuses in deposition, but we assume unit 3 had uninterrupted sedimentation. It is also possible that the two silty-clay layers in unit 3 represent rapid depositional events, but without distinct boundaries, they were not conclusively identified as such and retained in the age-depth model as continuous deposition. Unit 4 had the fastest accumulation rate of the core, with an average of 0.07 cm/yr.
Pollen and charcoal
The pollen record was divided into four pollen zones and three subzones based on constrained cluster analysis (Grimm, Reference Grimm1987; Fig. 3). Pollen percentages are reported as zone ranges, and the PAR, AP:NAP, and Pinus subg. Strobus to Pinus subg. Pinus ratios are reported as entire zone averages unless otherwise noted.
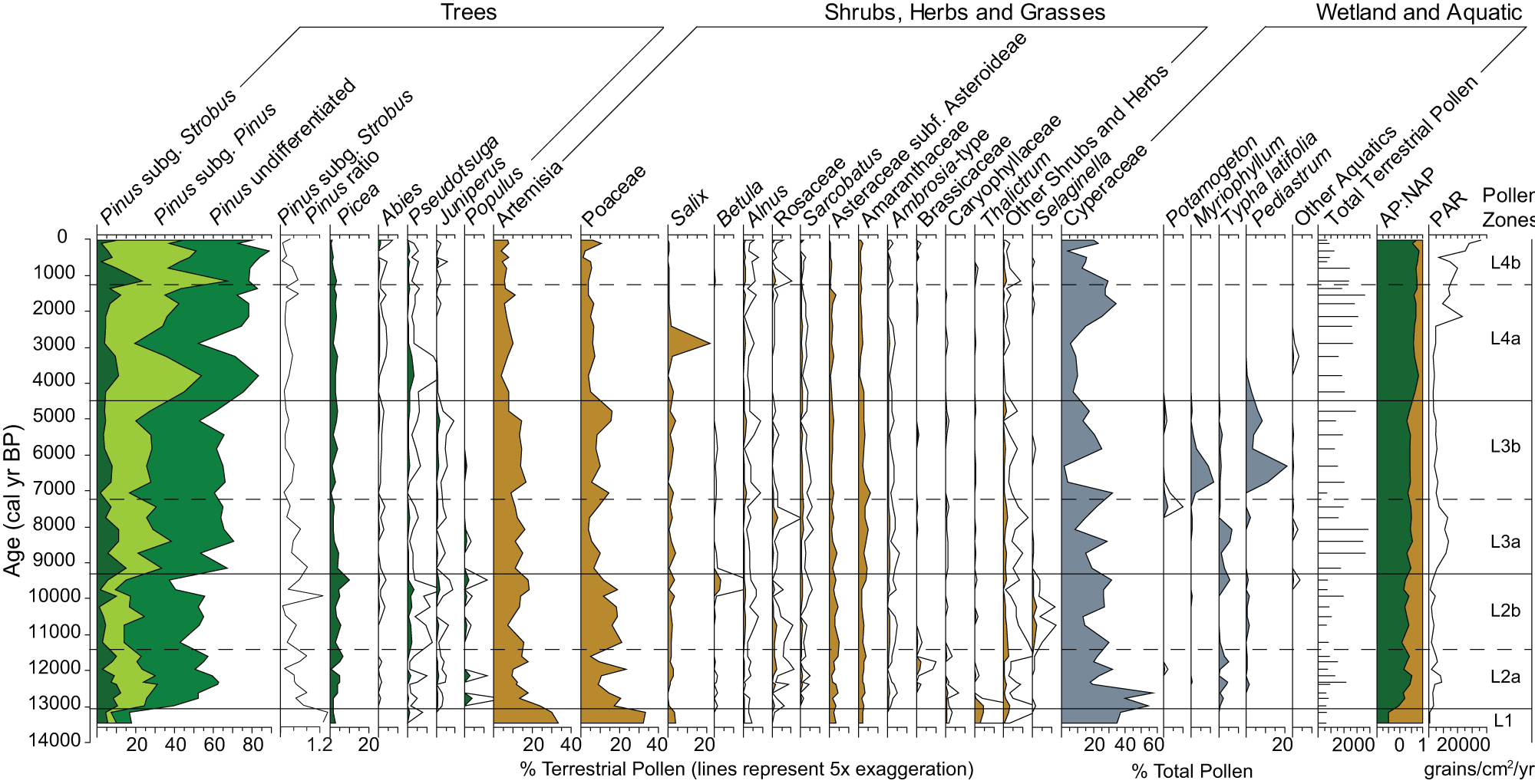
Figure 3. Pollen percentage diagram of selected taxa for Langohr Wetland, Montana. Terrestrial pollen percentages are based on the terrestrial pollen sum. Wetland and aquatic pollen percentages are based on the total pollen sum.
Zone L1 (250–238 cm depth; 14,000–13,000 cal yr BP) consists of two pollen samples at the base of the core that were characterized by high levels of nonarboreal pollen taxa. Both samples had high Artemisia and Poaceae percentages (30–33% and 32–34%, respectively), and a diverse assemblage of shrubs and upland herbs, including Salix (3–4%), Thalictrum (3–5%), Asteraceae subfam. Asteroideae (2–3%), Amaranthaceae (2%), Caryophyllaceae (< 1%), Ranunculaceae (< 1%), Fabaceae (< 1%), and Polygonum bistortoides-type (< 1%). Values of Pinus pollen (16–18%) were low, and the Pinus subg. Strobus to Pinus subg. Pinus ratio (1.38) was high. Picea values (2–3%) were low, and other conifer percentages were negligible (0–1%).
The low total pollen accumulation rate (PAR; 314 grains/cm2/yr) suggests a sparsely vegetated environment (Davis et al., Reference Davis, Brubaker, Webb, Birks and West1973), and the lowest AP:NAP value of the record (−0.49 vs. long-term average = 0.43) indicates an open landscape (Fall, Reference Fall1994). Cyperaceae pollen was abundant (34–37%) in the zone and may have been part of the upland vegetation; no other aquatic species were present. Charcoal was absent in this zone, suggesting minimal to no fires (Fig. 4). Overall, the assemblage suggests a cold tundra–steppe environment dominated by Artemisia and Poaceae with alpine herbs and scattered Pinus albicaulis/flexilis and Picea.
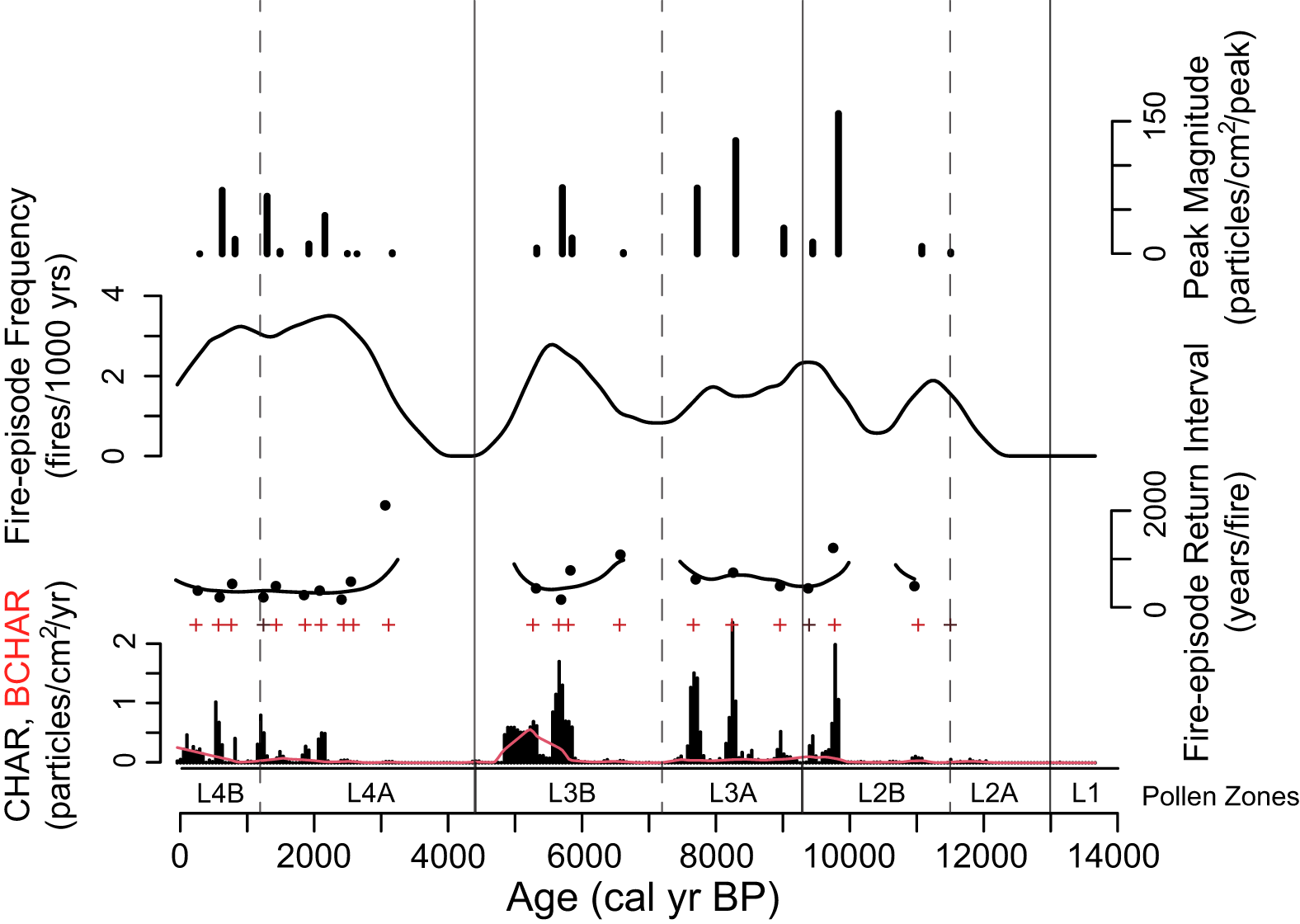
Figure 4. Charcoal accumulation rates (CHAR) from Langohr Wetland with background CHAR (BCHAR) in red. Significant fire peaks (+ signs) likely represent stand-replacing fire episodes. Fire-episode return interval is the years between significant peaks, fire-episode frequency is the number of peaks smoothed over a 1000-year moving window, and peak magnitude is the number of charcoal pieces per cm2 from all samples in a peak.
Zone L2 was characterized by increased arboreal pollen taxa and separated into two subzones. Subzone L2a (238–200 cm depth; 13,000–11,500 cal yr BP) had increased Pinus (40–61%) and Picea (to 7%), and a low appearance of Pseudotsuga (0.2–2%), Juniperus-type (1%), and Populus (1–4%) pollen. The Pinus subg. Strobus to Pinus subg. Pinus ratio decreased (0.53), suggesting an increase in Pinus contorta, although moderate levels of Pinus subg. Strobus pollen (avg = 8.5%) indicate Pinus albicaulis/flexilis was still present (Fall, Reference Fall1994). Artemisia and Poaceae declined (9–17% and 5–24%, respectively), as did Salix (< 3%) and Thalictrum (< 1%). Other shrubs and herbs (such as Asteraceae subfam. Asteroideae, Amaranthaceae, and Caryophyllaceae) remained at similar levels as in Zone L1, and Betula, Ambrosia-type, Sarcobatus, Rosaceae, and Brassicaceae pollen appeared in this subzone in low amounts (< 2%). Cyperaceae percentages were high at the base of the zone (high of 57%) but declined towards the top (low of 17%), and Typha pollen was present in moderate amounts (1–5%). CHAR remained negligible at the bottom of the subzone until 201 cm depth (ca. 11,500 cal yr BP) when values increased (to 0.06 particles/cm2/yr-) with the first small significant fire peak (peak magnitude = 1.9 particles/cm2/peak).
Subzone L2b (200–177 cm depth; 11,500–9300 cal yr BP) was distinguished from Subzone 2a by a slight decline in Pinus pollen (37–54%) and an increase in Picea (3–11%) and Pseudotsuga (1–4%). Artemisia and Poaceae pollen percentages remained unchanged (7–19% and 11–22%, respectively), and Juniperus and Populus percentages were low (< 2%). Betula and Amaranthaceae percentages increased at the top of the subzone (up to 4%), but other shrubs and herbs were the same as in Subzone L2a. Selaginella densa-type spore percentages increased during this subzone (1–2%), and after an initial decline from Subzone L2a, Cyperaceae and Typha increased towards the top of the subzone (from 13 to 32% and 0 to 5%, respectively). CHAR and BCHAR remained low at the base of the zone (< 0.1 particles/cm2yr), although one fire episode with a small peak magnitude (8.3 particles/cm2/peak) was recorded at 196 cm depth (ca. 11,000 cal yr BP). After a ca. 1200-year period of minimal CHAR, values increased to 2 particles/cm2/yr at the top of the subzone, and two fire episodes were recorded: one with a peak magnitude of 158.7 particles/cm2/peak (ca. 9800 cal yr BP) and the other with 13.2 particles/cm2/peak (ca. 9400 cal yr BP). Other than a ca. 1200-year gap with no fire, the mean return interval for fire episodes was approximately 380 years. Increases in PAR (1850 grains/cm2/yr) and AP:NAP (0.28) during Zone L2 suggest a parkland (Davis et al., Reference Davis, Brubaker, Webb, Birks and West1973; Fall, Reference Fall1992, Reference Fall1994), and the pollen and charcoal data suggest Picea was the dominant conifer with some Pinus albicaulis/flexilis surrounding the wetland at a time when stand-replacing fires started to occur.
Zone L3 was characterized by another increase in arboreal pollen taxa and decrease in nonarboreal pollen taxa. Subzone L3a (177–147 cm depth; 9300–7200 cal yr BP) registered an increase in Pinus pollen (55–71%) and decreases in Picea (1–4%) and Pseudotsuga (0–1%) pollen. The Pinus subg. Strobus to Pinus subg. Pinus ratio decreased (to 0.32), suggesting the increased presence of Pinus contorta. All shrubs, herbs, and grasses (such as Artemisia, 11–16%; Poaceae, 4–10%; and Betula, < 1%) decreased, other than Amaranthaceae, which increased (3–5%). Cyperaceae decreased (8–22%), and Typha latifolia increased to the highest values of the record (up to 7%). CHAR increased during this subzone to 2.4 particles/cm2/yr, and three significant charcoal peaks with magnitudes of 29.1, 128.1, and 74.5 particles/cm2/peak (ca. 9000, 8200, and 7700 cal yr BP, respectively). The mean return interval between fire episodes averaged approximately 570 years, or 1–2 episodes per 1000 years.
Subzone L3b (147–118 cm depth; 7200–4400 cal yr BP) was distinguished from Subzone L3a primarily by the increase in aquatic taxa. Cyperaceae pollen percentages reached the lowest values of the record (2%), Typha latifolia declined (< 1%), and Pediastrum (0–21%) and Myriophyllum (1–11%) increased. CHAR levels decreased from the previous subzone (up to 1.7 particles/cm2/yr), although four fire episodes were recorded with peak magnitudes of 1.4, 17. 8, 74.9, and 6.5 particles/cm2/peak (ca. 6600, 5835, 5690, and 5300 cal yr BP, respectively). The mean fire-episode return interval averaged approximately 900 years between ca. 8000 and 6000 cal yr BP and decreased to approximately 260 years between 6000 and 5000 cal yr BP. BCHAR increased for the first time in the record during this same period (to 0.5 particles/cm2/yr), suggesting more fires in the region. The increase in AP:NAP (0.44) and PAR (3704 grains/cm2/yr) in Zone L3 provides evidence of forest closure (Davis et al., Reference Davis, Brubaker, Webb, Birks and West1973; Fall Reference Fall1992). A forest with Pinus, Picea, and Pseudotsuga surrounded Langohr Wetland, and fires were likely stand-replacement events. The Subzone L3b aquatic pollen percentages suggest that the wetland may have deepened into a shallow pond.
Zone 4, which is characterized by the highest levels of arboreal pollen in the record, is split into two subzones. Subzone L4a (118–84 cm depth; 4400–1200 cal yr BP) had an overall increase in Pinus pollen (70–82%), although one sample showed a decrease to 52% at 109 cm depth (ca. 3000 cal yr BP). This decrease in Pinus values may be an artifact of percentage calculations since a large increase in Salix percentages (to 27%) occurs at this level. Pinus PAR, in fact, increased from 1083 grains/cm2/yr at 111 cm depth to 1266 grains/cm2/yr at 109 cm depth, and 1720 grains/cm2/yr at 106 cm depth (data not shown). Continued decrease in the Pinus subg. Strobus to Pinus subg. Pinus ratio (0.23) suggests the dominance of Pinus contorta over Pinus albicaulis/flexilis. Picea values were the same as in the previous zone (1–3%), while those of Pseudotsuga declined to the top of the zone following an initial increase (up to 4%), and those of Juniperus were negligible (< 1%). Artemisia and Poaceae levels decreased (4–12% and 3–8%, respectively). Cyperaceae percentages increased towards the top of the subzone (from 5 to 34%), and Myriophyllum, Potamogeton, Typha, and Pediastrum values dropped to 0%. CHAR was low at the base of the subzone (∼0 particles/cm2/yr until 102 cm depth, ca. 2000 cal yr BP) and increased at the top of the subzone (to 0.5 particles/cm2/yr), recording seven charcoal peaks with magnitudes of 1.2, 0.1, 0.3, 43.4, 11.1, 2.8, and 65.1 particles/cm2/peak (at ca. 3150, 2600, 2475, 2140, 1900, 1470, and 1275 cal yr BP, respectively). Following a period of 1700 years with no fire episodes (from ca. 4700–3000 cal yr BP), fire-episode frequency increased to ∼2–3 episodes per 1000 years, with a mean return interval of approximately 312 years.
Subzone L4b (84–0 cm depth; 1200 cal yr BP to present day) was distinguished from Subzone L4a by the highest levels of Pinus pollen in the record (73–90%), whereas Picea (1–3%), Pseudotsuga (< 1%), and Juniperus (< 1%) pollen remained low. Abies reached the highest levels of the record at the top of the zone, although still minimal (1.5%). Artemisia and Poaceae values were the same as in the previous subzone, although Poaceae percentages increased at the top of the subzone (to 10%). Other shrubs and herbs were in low abundance throughout the subzone. Cyperaceae was the only aquatic pollen type present in this subzone and its percentages increased at the top (from 4 to 23%). CHAR levels increased (high of 0.8 particles/cm2/yr), and three significant peaks had magnitudes of 16.4, 71.9, and 0.02 particles/cm2/peak (ca. 800, 600, and 270 cal yr BP, respectively). Mean return interval of fire episodes remained similar to before, at 336 years, which equals 2–3 episodes per 1000 years. BCHAR also increased towards the top of this subzone (to 0.26 particles/cm2/yr). The highest AP:NAP (0.70) ratio and PAR (7070 grains/cm2/yr) suggest Zone L4 (and particularly Subzone L4b) was a period of dense forest (Davis et al., Reference Davis, Brubaker, Webb, Birks and West1973; Fall, Reference Fall1994). A closed mixed-conifer forest of predominantly Pinus contorta with Picea and Pseudotsuga surrounded the Cyperaceae-covered Langohr Wetland, and episodes of stand-replacing fires were approximately 300 years apart with the most recent fire episode occurring approximately 330 years ago. Elevated levels of CHAR and BCHAR at the very top of the core suggest that fire activity in the region increased between 600 years ago and present day.
Discussion
The Langohr Wetland record provides a postglacial vegetation and fire history of a mixed-conifer subalpine forest in the northern Greater Yellowstone Ecosystem (GYE) and highlights the relative stability of these wet forests through time. Vegetation structure and fire activity generally followed millennial-scale climate trends driven by changes in the seasonal cycle of insolation (Berger, Reference Berger1978).
Here, we describe the Langohr Wetland record and compare it with other pollen and charcoal records from sites across the northern GYE (Table 2, Fig. 1). Background charcoal levels were minimal throughout the Langohr record, and it is likely that fine charcoal particles (< 120 microns) were not preserved in wetland sediments (due to degradation, or other factors). As a result, charcoal peaks are likely limited to representing very local (< 1 km) fires within the Langohr watershed. Hence, we assume that the fire record likely focuses on fire events immediately adjacent to Langohr Wetland and does not include fire events that lacustrine records would capture from a larger landscape.
Table 2. Sites in the Greater Yellowstone Ecosystem used for pollen and charcoal comparison. JJA/DJF is the summer to winter precipitation ratio.
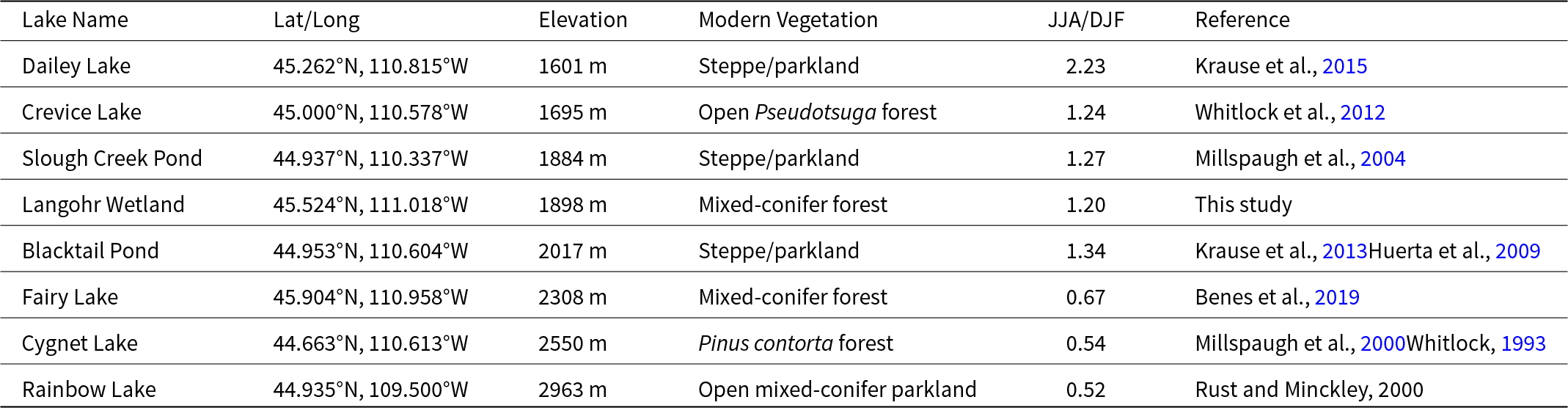
The nearest study sites to Langohr Wetland are high-elevation Fairy Lake (∼40 km NE from Langohr), a subalpine lake surrounded by mixed-conifer forest in the Bridger Mountain Range (Benes et al., Reference Benes, Iglesias and Whitlock2019), and low-elevation Dailey Lake (∼30 km SE from Langohr) in the Yellowstone River Valley surrounded by steppe (Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015). These three sites allow for comparison across an elevational gradient, although Dailey Lake’s published vegetation and charcoal record only covers ca. 13,900–7500 cal yr BP. Other sites located farther south are Slough Creek Pond (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004), Blacktail Pond (Huerta et al., Reference Huerta, Whitlock and Yale2009; Krause and Whitlock, Reference Krause and Whitlock2013), and Crevice Lake (record spans 9400 yr BP to present day; Whitlock et al., Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012) located in steppe/Pseudotsuga parkland in northern YNP. We also considered data from Cygnet Lake (Whitlock, Reference Whitlock1993; Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000), which lies in the Pinus contorta forest on the rhyolite plateau of central YNP. The highest elevation record for comparison is Rainbow Lake (Rust and Minckley, Reference Rust and Minckley2020), located in Picea, Abies, and Pinus albicaulis parkland on the Beartooth Plateau in the northeastern GYE.
Four of the sites (Dailey Lake, Crevice Lake, Slough Creek Pond, and Blacktail Pond) receive less than 500 mm annual precipitation and are considered summer-wet sites, meaning they receive more precipitation during summer than winter months (Whitlock and Bartlein, Reference Whitlock and Bartlein1993). The other three sites receive over 500 mm of annual precipitation and are located in the summer-dry region, which receives more precipitation in winter. It has been proposed that during the Early Holocene, summer-wet areas became wetter as a result of strengthened monsoonal circulation, whereas summer-dry regions became drier as a result of a stronger-than-present subtropical high (Whitlock and Bartlein, Reference Whitlock and Bartlein1993). A comparison of these records offers insight into how different forest ecosystems responded to late-glacial and Holocene climate variations and provides new information to compare with a regional study by Iglesias et al. (Reference Iglesias, Whitlock, Krause and Baker2018).
Late-glacial (> 14,000–11,500 cal yr BP): transition from tundra–steppe to Picea parkland
During the last glacial maximum (LGM), a large glacial complex covered the Greater Yellowstone Ecosystem, and alpine glaciers covered adjacent mountain ranges (Weber, Reference Weber1965; Pierce, Reference Pierce1979; Licciardi and Pierce, Reference Licciardi and Pierce2018). Terminal moraines approximately 20 km to the east of Langohr Wetland retreated from their maximum position after ca. 17,900 ka (Licciardi and Pierce, Reference Licciardi and Pierce2018). Recessional moraines ∼40 km to the southeast in the Yellowstone Valley that date to ca. 15,100 ka (Licciardi and Pierce, Reference Licciardi and Pierce2018) suggest that the basal inorganic clay at Langohr Wetland, dating to a minimum of ca. 13,500 cal yr BP, accumulated in an ice-free depression as part of deglaciation.
Artemisia tundra–steppe vegetation developed initially at Langohr Wetland and was replaced by Picea-dominated parkland with lesser amounts of Pinus at ca. 13,000 cal yr BP. Picea percentages above 6% suggest that Picea trees were abundant (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018), and though the rapid increase in Pinus at ca. 13,000 cal yr BP likely marks its arrival in the Hyalite Creek watershed, the fact that percentages were below 60% suggests that Pinus trees were not abundant locally (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). The high Pinus subg. Strobus to Pinus subg. Pinus ratio indicates Pinus albicaulis/flexilis dominance initially, consistent with a colder-than-present climate. The transition from steppe to parkland, along with the appearance of Typha latifolia and the formation of organic peat sediment (lithological unit L2), implies the development of a productive wetland and warmer conditions than before by 13,000 cal yr BP.
In addition to Picea and Pinus, the pollen record indicates that Pseudotsuga trees were also present at Langohr Wetland as early as 13,000 cal yr BP. Pseudotsuga pollen fluctuated between 0 and 1% until 11,500 cal yr BP before increasing in abundance during the Holocene. Pseudotsuga is often underrepresented in pollen records due to its poor production, dispersal, and preservation, and pollen levels as low as 0.5% can suggest its presence on the landscape (Gugger and Sugita, Reference Gugger and Sugita2010). Thus, it seems likely that Pseudotsuga was present in low numbers, probably on south-facing slopes where it grows today. Abies pollen percentages were low throughout the entire record until the very top of the core, and Abies was probably not well represented in the local vegetation until recently.
The late-glacial transition from tundra–steppe to parkland is recorded in sites across the region (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018) and supports paleoclimate model simulations, which show that following a cold, dry late-glacial period, rising summer insolation and a northward shift of the jet stream from its full-glacial position, increased summer temperatures and brought increased moisture to the region (Bartlein et al., Reference Bartlein, Anderson, Anderson, Edwards, Mock, Thompson, Webb, Webb and Whitlock1998; Alder and Hostetler, Reference Alder and Hostetler2015). Landscapes dominated by Artemisia, Poaceae, and herbs were replaced primarily by Picea parkland at all but the highest elevations by the end of the late-glacial period. Dailey Lake and Fairy Lake differed in the timing of conifer arrival as a consequence of their elevations. Similar to Langohr Wetland, Picea parkland established at low-elevation Dailey Lake at ca. 13,300 cal yr BP (Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015), whereas high-elevation Fairy Lake supported tundra–steppe until 11,000 cal yr BP (Benes et al., Reference Benes, Iglesias and Whitlock2019). Picea parkland was established at Blacktail and Slough Creek ponds in northern YNP by 12,500 cal yr BP (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004; Krause and Whitlock, Reference Krause and Whitlock2013), and similar to Fairy Lake, higher-elevation Rainbow and Cygnet lakes in central YNP supported tundra–steppe throughout the late-glacial period until ca. 10,500 and 11,500 cal yr BP, respectively (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000; Rust and Minckley, Reference Rust and Minckley2020). Thus, the development of early parkland was influenced by glacial recession, distance to refugial conifer populations, and the rate of warming and soil development (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). Tree establishment occurred first at low elevations and last at high elevations and in central YNP (Krause and Whitlock, Reference Krause and Whitlock2017).
No fire episodes were recorded at Langohr Wetland in the late-glacial period, suggesting patchy woody fuel cover and low temperatures at this time likely limited fire spread. Fire activity remained low throughout the parkland period between ca. 13,000-11,500 cal yr BP, when conditions were still cool. It is possible that low-severity surface fires, which do not produce much charcoal, occurred but were not detected in the charcoal record.
Fire activity was generally low across the region during the late-glacial period (Fig. 5). Unlike Langohr Wetland, however, fire activity did increase with the appearance of Picea parkland at Dailey Lake, Slough Creek, and Blacktail Pond at ca. 13,200, 12,250, and 12,000 cal yr BP, respectively (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004; Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015). These records suggest biomass burning increased in the late-glacial period as Picea established and expanded, but generally low charcoal influx from regional records indicates biomass burning was limited.
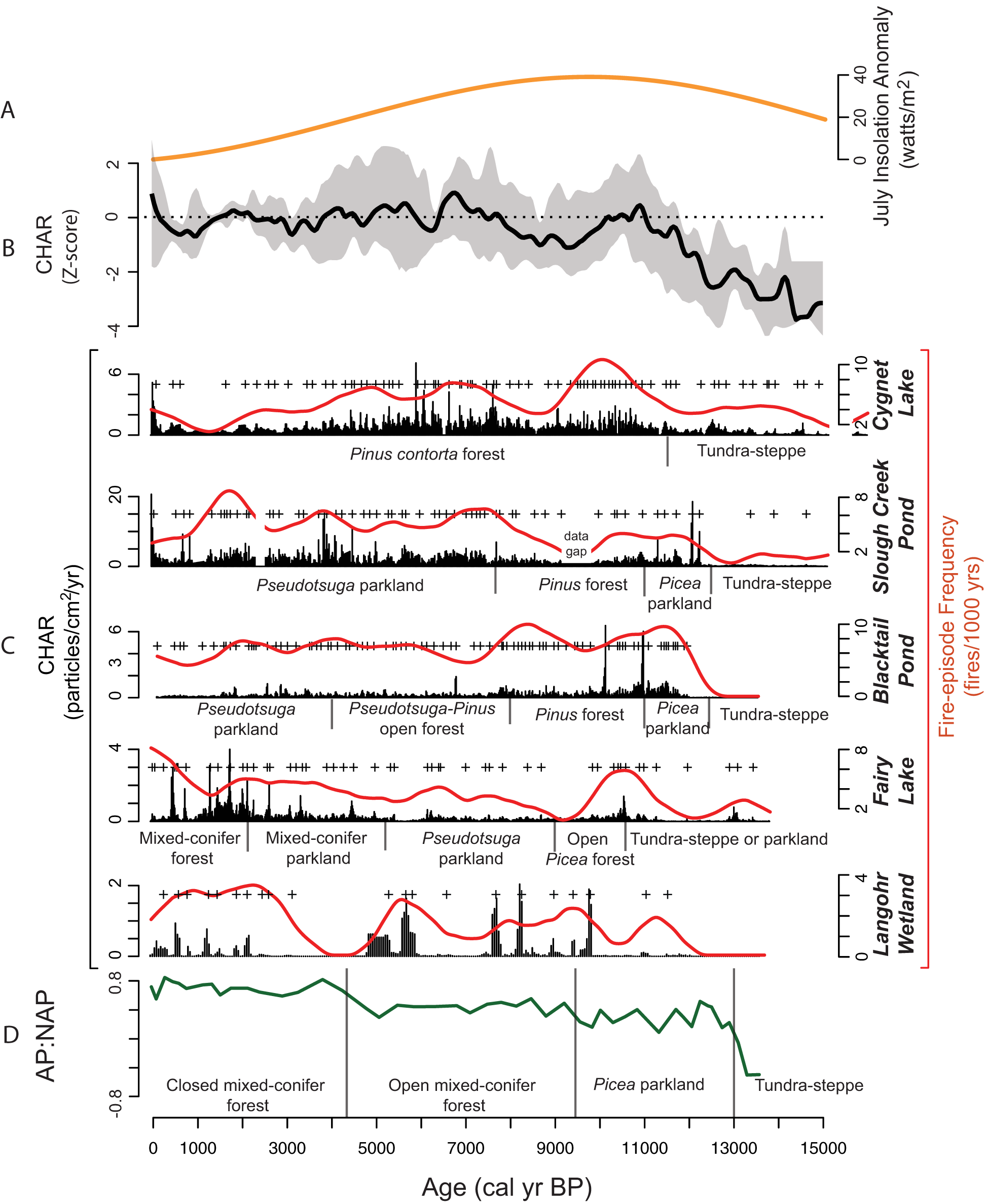
Figure 5. Select regional charcoal accumulation (CHAR) records and a CHAR composite from sites in the GYE that extend into the late glacial period (13,500–15,000 cal yr BP), arranged by their proximity to Langohr Wetland: Fairy Lake (Benes et al., Reference Benes, Iglesias and Whitlock2019), Blacktail Pond (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004), Slough Creek Pond (Huerta et al., Reference Huerta, Whitlock and Yale2009), and Cygnet Lake (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000). (A) July insolation anomaly at 45°N (Berger, Reference Berger1978). (B) Regional fire history based on Z-score anomalies of composited CHAR data from the five sites. Gray shading represents 95% confidence interval. (C) CHAR (black bars), fire-episode frequency reconstruction (red lines), and pollen-inferred vegetation for each site. Small black bars denote the timing of vegetation changes, and (+) signs signify significant fire episodes. (D) The arboreal-to-non-arboreal pollen (AP:NAP) ratio, which indicates the abundance of forest versus open vegetation, and the inferred vegetation at Langohr Wetland. Black bars between text denote the timing of vegetation changes.
Early Holocene (ca. 11,500–7000 cal yr BP): transition from parkland to open mixed-conifer forest
The Picea-dominated parkland persisted at Langohr Wetland at the start of the Early Holocene, indicated by the high percentages of Poaceae and Selaginella densa-type (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). The increase and consistent presence of Pseudotsuga pollen after ca. 11,500 cal yr BP indicates Pseudotsuga trees became more established on the landscape, suggesting the continued warming conditions of the Early Holocene. Populus spp. and patches of Salix likely grew in the riparian areas along the stream and wetland.
The parkland became more forested at ca. 9300 cal yr BP, indicated by an increase in arboreal pollen percentages and decreases in Poaceae and Selaginella densa-type values. Picea and Pseudotsuga were still present but less abundant, and Pinus dominated the forest. A low Pinus subg. Strobus to Pinus subg. Pinus ratio suggests the presence of both Pinus flexilis/albicaulis and Pinus contorta, with Pinus contorta dominating, also suggesting warming conditions. The presence of Artemisia in moderate amounts implies areas of steppe at lower and/or upper elevations and in dry forest openings.
Open and closed forests replaced parkland across the region in response to the continued warming and increased effective moisture during the late-glacial and Holocene transition (Bartlein et al., Reference Bartlein, Anderson, Anderson, Edwards, Mock, Thompson, Webb, Webb and Whitlock1998; Alder and Hostetler, Reference Alder and Hostetler2015). The timings of forestation varied (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018), with most sites becoming forested before Langohr Wetland. At Dailey Lake, forestation occurred at ca. 12,300 cal yr BP, with increases in Abies, Picea, and Pinus albicaulis/flexilis (Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015), and Fairy Lake became more closed with conifer forest development at ca. 11,000 cal yr BP (Benes et al., Reference Benes, Iglesias and Whitlock2019). Upper treeline and subalpine parkland moved upslope while lower- and middle-elevation forests became denser and Pinus dominated in the other areas of the GYE as well. Pinus forest was established at Blacktail Pond and Cygnet Lake by 11,500 cal yr BP, and Pinus and Juniperus trees were present at Slough Creek Pond at ca. 11,000 cal yr BP (Millspaugh et al., Reference Millspaugh, Whitlock and Bartlein2000, Reference Millspaugh, Whitlock, Bartlein and Wallace2004; Krause and Whitlock, Reference Krause and Whitlock2013). High-elevation Rainbow Lake transitioned from tundra–steppe to open Pinus forest by ca. 10,000 cal yr BP (Rust and Minckley, Reference Rust and Minckley2020).
Summer (winter) insolation levels continued to increase (decrease) into the early Holocene, reaching a maximum (minimum) at 11,000-10,000 cal yr BP (Berger, Reference Berger1978). Higher summer insolation increased summer temperature and transpiration (reducing effective moisture), and indirectly strengthened the northeastern Pacific subtropical high-pressure system. These conditions increased summer drought in the region (Bartlein et al., Reference Bartlein, Anderson, Anderson, Edwards, Mock, Thompson, Webb, Webb and Whitlock1998; Alder and Hostetler, Reference Alder and Hostetler2015); and at some sites, this led to more open forests. Dailey Lake recorded an open Pseudotsuga and Pinus forest at ca. 10,200 cal yr BP (Krause et al., Reference Krause, Lu, Whitlock, Fritz and Pierce2015), and Fairy Lake supported a similar forest after ca. 9000 cal yr BP (Benes et al., Reference Benes, Iglesias and Whitlock2019). A record from an ice patch on the Beartooth Plateau also suggests more open vegetation at high elevations at ca. 10,000 cal yr BP (Alt et al., Reference Alt, Puseman, Lee, Pederson, McConnell, Chellman and McWethy2024). Evidence of drier conditions also comes from declining lake levels in the GYE at Moran Bay in Grand Teton National Park in the southern GYE at ca. 10,400 cal yr BP (Dilworth et al., Reference Dilworth, McGlue, Thigpen, Brown, Yeager, Woolery, Johnson, Whitehead, Cortese and Matocha2024), Rainbow Lake on the Beartooth Plateau at ca. 10,000 cal yr BP (Shuman and Serravezza, Reference Shuman and Serravezza2017), and Lake of the Woods, Wyoming, just south of YNP, at ca. 9000 cal yr BP (Shuman et al., Reference Shuman, Henderson, Colman, Stone, Fritz, Stevens, Power and Whitlock2009). These vegetation and hydrological changes seem explained by climate conditions created by the summer insolation maximum.
Other sites, like Langohr Wetland, remained forested during the Early Holocene, despite warmer and drier conditions, but probably for different reasons. The wet setting of Langohr Wetland may have provided a refugium for mesophytic taxa and thus was not as responsive to intensified summer drought as other locations. Cygnet Lake was surrounded by a closed Pinus contorta forest because of its location on the infertile rhyolitic Central Plateau, which is most conducive to Pinus contorta growth (Whitlock, Reference Whitlock1993). High-elevation Rainbow Lake showed an increase in forest cover as a result of an upslope shift in treeline under warming conditions. Parkland surrounding Fairy Lake and alpine tundra at the ice patch suggests some high-elevation areas did not support closed forests despite an overall upward shift in treeline, which could be attributed to local topographical features (such as locations in large open basins and high plateaus) causing them to be more susceptible to summer drought.
Slough Creek Pond, Blacktail Pond, and Crevice Lake show evidence of more forest cover in the Early Holocene followed by a shift to more open forest and steppe after ca. 8000 cal yr BP. This later timing for forest opening has been attributed to their location in a summer-wet precipitation regime that may have been wetter during the Early Holocene insolation maximum and subsequently became more open as summer insolation decreased (Whitlock and Bartlein, Reference Whitlock and Bartlein1993). At the other summer-wet site, Dailey Lake, the transition toward open vegetation occurred at ca. 10,000 cal yr BP (2000 years earlier), with the increase in summer insolation levels, suggesting it was not wetter during the Early Holocene, despite its location in a summer-wet region.
The earliest evidence of Pseudotsuga occurred at Langohr Wetland (pollen percentage of 1.5% at ca. 13,100 cal yr BP), and its sustained presence after 11,500 cal yr BP (continuous percentages between 1 and 5%) also pre-dates other sites. Dailey Lake first registered Pseudotsuga pollen at ca. 12,500 cal yr BP with a sustained presence after ca. 10,200 cal yr BP, and Slough Creek Pond, Blacktail Pond, and Fairy Lake do not have pollen evidence of Pseudotsuga until ca. 9000 cal yr BP. Both Dailey Lake and Langohr Wetland provide support for the hypothesis of a glacial age refugial population of Pseudotsuga in the unglaciated parts of the Gallatin or upper Yellowstone valleys close to the Yellowstone Ice Cap. As conditions warmed, Pseudotsuga populations at low and middle elevations were able to colonize higher deglaciated elevations (Krause and Whitlock, Reference Krause and Whitlock2017).
Fire activity at Langohr Wetland was low until ca. 10,000 cal yr BP, when an increase in charcoal peaks marks a shift towards more stand-replacing fires. After a large fire episode in the record (ca. 9800 cal yr BP), Pinus percentages decreased, and percentages of Juniperus, Artemisia, and Betula increased for approximately 500 years. The timing of the first large fires during the summer insolation maximum (ca. 10,000 cal yr BP) suggests that warm, dry conditions led to increased fire activity and prevented forest regeneration for hundreds of years. The slight increase in Juniperus and Pseudotsuga at that time suggests more drought-tolerant species after the fire episode.
As the forest around Langohr Wetland became more Pinus-dominated after ca. 9300 cal yr BP, stand-replacing fire episodes occurred regularly until ca. 7500 cal yr BP. The mean fire-episode return interval of 570 years between fire episodes suggests a long time between stand-replacing fire episodes, characteristic of a high-intensity fire regime, although the fire return interval was longer than the 150–300-year fire return interval based on tree ring studies in northern Rocky Mountain subalpine forests (Baker, Reference Baker2009). The coincidence of the first large fire episodes and a shift from Picea-dominated parkland to Pinus-dominated mixed-conifer forest suggests that an increase in fire activity may have fostered the transition from open to closed forest by favoring more fire-tolerant Pinus over Picea (Krause and Whitlock, Reference Krause and Whitlock2013).
Fire activity similarly increased across the GYE during the late-glacial to Early Holocene transition, likely in response to warming and increased woody fuel (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). Many sites register an increase in fire activity earlier than at Langohr Wetland, as part of their vegetation transition to more conifers on the landscape during the late-glacial (Fig. 5). Dailey Lake, for example, experienced increased fire activity during the shift from parkland to forest at ca. 12,000 cal yr BP, and Fairy Lake registered increased fire activity with the establishment of open Picea forest at ca. 11,000 cal yr BP. Rainbow Lake supported more fires when tundra–steppe transitioned to open forest at ca. 10,000 cal yr BP (Rust and Minckley, Reference Rust and Minckley2020). Similar to Langohr Wetland, Cygnet Lake recorded an increase in fire frequency at ca. 10,000 cal yr BP in response to the summer insolation maximum. Mean fire return intervals for all sites were shorter than at Langohr Wetland, which may reflect its uniquely mesic setting, or indicate that Langohr only registered fires immediately surrounding the wetland.
Similar to Langohr Wetland, a rise in fire activity at many sites also accompanied the subsequent transition from Picea parkland to closed Pinus forest (Fig. 5; Krause and Whitlock, Reference Krause and Whitlock2013). Stand-replacing fire activity increased at Blacktail Pond at 11,500 cal yr BP. Forest cover increased soon thereafter and became a closed Pinus forest at 10,000 cal yr BP. At Slough Creek Pond, fires increased at 12,250 cal yr BP, followed by forest development at 11,000 cal yr BP, and then closed Pinus forest at 10,750 cal yr BP. Fire activity increased at Cygnet Lake at 11,000 cal yr BP coincident with the development of Pinus contorta establishment, and closed Pinus forest was present at 10,750 cal yr BP. The synchronous rise in fire activity was initiated by a warming climate, and then Pinus albicaulis, Pinus flexilis, and Pinus contorta colonized burned areas at the expense of Picea engelmannii (Iglesias et al., Reference Iglesias, Krause and Whitlock2015; Krause and Whitlock, Reference Krause and Whitlock2013, Reference Krause and Whitlock2017), which has little adaptation to fire (Agee, Reference Agee1993). The predominantly Pinus forests were then maintained by more frequent fires than before (Krause and Whitlock, Reference Krause and Whitlock2013, Reference Krause and Whitlock2017; Iglesias et al., Reference Iglesias, Krause and Whitlock2015).
After ca. 10,000 cal yr BP, many forests that became more open experienced a decrease in stand-replacing fires. Fire activity decreased at Dailey and Fairy lakes when the vegetation transitioned to parkland at ca. 10,000 cal yr BP. However, fire activity increased at 10,000 cal yr BP with the increase in summer drought and forest cover at Blacktail Pond, Cygnet Lake, Rainbow Lake, and Langohr Wetland (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004; Rust and Minckley 2018). Fire activity at the forested sites likely increased due to a combination of abundant fuel and warm, dry conditions. Only open sites at upper and lower treeline had a decline in fire episodes, probably because of lack of fuel connectivity; however, we cannot rule out the possibility that these ecotones experienced low-severity fires not detected in the charcoal record. At Blacktail Pond and Crevice Lake, fires were less frequent after the forest transitioned to Pseudotsuga–Pinus parkland at ca. 8000 cal yr BP. This suggests that while vegetation at some of the “summer-wet” sites seems to suggest a wetter Early Holocene (forests at Blacktail Pond, Slough Creek Pond, and Crevice Lake), regional fire activity generally corresponds to the availability of woody biomass, increasing with Early Holocene warmth, and decreasing with the transition to parkland.
Mid-Holocene (ca. 7000–4500 cal yr BP): establishment of mixed-conifer forest
The mixed-conifer forest surrounding Langohr Wetland became less Pinus-dominated at ca. 7000 cal yr BP with the increased presence of Pseudotsuga and Picea, and changes in lithology and aquatic taxa suggest variations in wetland hydrology. At ca. 7000 cal yr BP, a lithological change from peat to fine-detritus sapropel (lithological unit 2 to 3), followed by a decline in emergent wetland taxa (Cyperaceae and Typha) and an increase in aquatic taxa (Myriophyllum and green algae Pediastrum) from ca. 7000–4500 cal yr BP imply areas of open water in the wetland. Myriophyllum, in particular, is a submerged aquatic plant (Lesica, Reference Lesica2012), and the increased abundance of both taxa suggests a deeper wetland and increased effective moisture.
As summer insolation declined into the Mid-Holocene, the high seasonality of the Early Holocene attenuated and the strength of the subtropical high-pressure system weakened, decreasing summer drought (Alder and Hostetler, Reference Alder and Hostetler2015). Vegetation conditions across the region responded to these changing climate conditions, although again the timing varied from site to site. Langohr Wetland maintained a mixed-conifer forest, but the increase in aquatic taxa suggests greater effective moisture at ca. 7000 cal yr BP, and Fairy Lake transitioned from Pseudotsuga parkland to a mixed-conifer parkland at ca. 5000 cal yr BP. There was an increase in Pseudotsuga abundance around Dailey Lake (Benes et al., Reference Benes, Iglesias and Whitlock2019), and tree density increased on the Beartooth Plateau at ca. 6000 cal yr BP (Alt et al., Reference Alt, Puseman, Lee, Pederson, McConnell, Chellman and McWethy2024). Low- and middle-elevation sites in other regions of the northern Rocky Mountains also transitioned to closed forests between ca. 6000 and 5500 cal yr BP (Alt et al., Reference Alt, McWethy, Everett and Whitlock2018; McWethy et al., Reference McWethy, Alt, Argiriadis, Battistel, Everett and Pederson2020). Lake levels in sites from the GYE and across the northern Rocky Mountains rose to present levels between ca. 6600 and 5500 cal yr BP, also indicating wetter conditions than before (Shuman et al., Reference Shuman, Henderson, Colman, Stone, Fritz, Stevens, Power and Whitlock2009; Shuman and Marsicek, Reference Shuman and Marsicek2016; Shuman and Serravezza, Reference Shuman and Serravezza2017; Dilworth et al., Reference Dilworth, McGlue, Thigpen, Brown, Yeager, Woolery, Johnson, Whitehead, Cortese and Matocha2024).
Other sites in this study did not experience a vegetation change reflective of cooler, wetter conditions at this time. For some of them, this may be attributed to their summer-wet precipitation regime (Whitlock and Bartlein, Reference Whitlock and Bartlein1993). After the transition to parkland at ca. 8000 cal yr BP, summer-wet sites Slough Creek Pond, Blacktail Pond, and Crevice Lake experienced conditions that became drier towards the present day (Whitlock and Bartlein, Reference Whitlock and Bartlein1993), and forest structure remained open (Huerta et al., Reference Huerta, Whitlock and Yale2009; Whitlock et al., Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012). The summer-dry sites that did not record vegetation change are Cygnet Lake, which was surrounded by closed Pinus contorta forest due to edaphic controls (Whitlock, Reference Whitlock1993), and high-elevation Rainbow Lake, which maintained open forest/parkland throughout the entire Holocene, similar to other high-elevation sites in the GYE (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018). Although there was no vegetation shift at these sites, a decrease in fire-episode frequency after ca. 6000 cal yr BP at Rainbow and Cygnet lakes is consistent with decreased summer temperatures and increased effective moisture (Alder and Hostetler, Reference Alder and Hostetler2015).
Fire activity at Langohr Wetland decreased between 7200 and 6000 cal yr BP, during the same time interval as the increase in aquatic taxa and lacustrine-derived sediment. Also, within the lacustrine sediment (lithological unit 3) was an interval of high inorganic silt/clay content (150–138 cm depth), suggesting the introduction of allochthonous material during flood events or periods of rapid snowmelt. These wet events could have contributed to the absence of fire during this time. An increase in CHAR and BCHAR at 6000 cal yr BP after approximately 1000 years without large fires indicates a return of stand-replacing fires, as well as an increase in fires regionally.
Fairy Lake and Langohr Wetland registered increased fire-episode frequency after 6000 cal yr BP, coincident with forest closure (Fig. 5). Higher fire activity may have been a result of greater fuel availability at those sites, but the increase is also noted throughout the GYE (Iglesias et al., Reference Iglesias, Whitlock, Krause and Baker2018), including at summer-dry site Crevice Lake (Whitlock et al., Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012), and coincides with an increase in fire-related sedimentation events at 5500 cal yr BP in northern YNP (Meyer et al., Reference Meyer, Wells and Timothy Jull1995). The overall vegetation changes of the Early and Mid-Holocene generally support the summer-wet and summer-dry hypothesis of the precipitation amplification in the Early Holocene (Whitlock and Bartlein, Reference Whitlock and Bartlein1993), but the fire histories seem to reflect more site-to-site variations in local vegetation and climate.
Late Holocene (ca. 4500–present day): increase in forest cover
After ca. 4500 cal yr BP, the present-day dense mixed-conifer forest of Pinus, Picea, and Pseudotsuga was established at Langohr Wetland, with Abies becoming more prevalent in the last 100 years. AP:NAP and PAR reached a maximum, and Artemisia, Poaceae, and other shrubs and herbs declined to their lowest levels of the record. Aquatic taxa also declined, and the lithology shifted from limnic back to sedge-dominated peat (lithological unit 4) at ca. 2500 cal yr BP, suggesting an infilling of the basin and marking establishment of the modern wetland. A slight increase in Pinus flexilis/albicaulis at ca. 1100 cal yr BP occurred at the onset of the Medieval Climate Anomaly (MCA; ca. 1050–650 cal yr BP; Cook et al., Reference Cook, Woodhouse, Eakin, Meko and Stahle2004). It may be evidence of an upslope shift in the range of low-elevation Pinus flexilis during warmer and drier conditions, but this hypothesis warrants further investigation.
The Late Holocene climate was characterized by continued cool and wet conditions but superimposed on this trend were warmer and cooler centuries, including during the MCA and the Little Ice Age (LIA; 550–200 cal yr BP; Mann et al., Reference Mann, Zhang, Rutherford, Bradley, Hughes, Shindell, Ammann, Faluvegi and Ni2009). Forest composition around Langohr Wetland and Fairy Lake changed little during the Late Holocene, but forest cover increased after ca. 3000 cal yr BP. At this same time, Crevice Lake registered a transition from open Juniperus–Pinus forest to open Pseudotsuga–Pinus forest, and pollen data at Blacktail Pond and Slough Creek Pond indicate the establishment of the present-day steppe and forest ecotone. Generally, summer-wet sites maintained an open xerophytic vegetation structure, also reflective of their minimal annual precipitation amounts. No change in vegetation was noted during the MCA or LIA, other than an increase in Pinus flexilis/albicaulis at Langohr Wetland during the MCA.
A decrease in fire activity at Langohr Wetland from ca. 4700 to 3000 cal yr BP occurred during the deposition of silty-clay sediment (120–115 cm depth, unit L3), which could again suggest wetter conditions or a hiatus. Episodes of stand-replacing fires increased in frequency between ca. 2500 cal yr BP and the present day, and the shortest mean return interval of fire episodes was approximately 330 years. This interval is consistent with fire return intervals from dendrochronological studies in subalpine forests in western Montana (Baker, Reference Baker2009), although it is longer than the 35–200+ year fire return interval reported for the Custer–Gallatin National Forest (Erickson, Reference Erickson2022). Charcoal peak magnitudes during this period were lower than earlier in the record, but the long 330-year interval between fire episodes suggests that they were likely stand-replacing events. The most recent significant fire episode in the Langohr Wetland record occurred approximately 330 years ago (ca. 270 cal yr BP), and a general increase in BCHAR in the last century indicates increased fire activity in the region during Euro-American settlement.
Fire trends diverged across the region during the Late Holocene. From 3000 cal yr BP to present day, Fairy Lake and Langohr Wetland had the highest level of fire activity in their records, while other sites had decreases. The frequency of stand-replacing fire episodes increased for both sites at a time when surrounding forests were becoming denser and woody fuel load was increasing. Although fire-episode frequency at Fairy Lake was within the expected range for stand-replacing fires in subalpine forests (∼6 episodes/1000 yr), fire-episode frequency at Langohr (2–3 episodes/1000 years) was longer than the expected range (Baker, Reference Baker2009).
Charcoal records from Blacktail Pond, Slough Creek Pond, Cygnet Lake, and Rainbow Lake all register a decrease in fire frequency over the last 3000 years, likely reflecting a decline in stand-replacing fires (Huerta et al., Reference Huerta, Whitlock and Yale2009). At Slough Creek Pond and Blacktail Pond, a decrease in fire frequency was likely due to more surface fires in expanded steppe vegetation. Fire episodes at Cygnet Lake and Rainbow Lake continued to decrease in frequency from the Mid-Holocene, and over the last ca. 2000 years, the frequency was similar to that of Langohr Wetland (2–3 episodes/1000 yr), within current estimates of stand-replacing fires in subalpine forests (Baker, Reference Baker2009). Fire activity at Crevice Lake fluctuated throughout the Late Holocene but remained at about eight episodes/1000 yr until the last 300 years when large fires became more frequent (Whitlock et al., Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012). Regional CHAR also suggests an increase in fire activity over the last ca. 300 years (Fig. 5), mainly influenced by CHAR inputs from the 1988 fires in YNP. Despite increases in regional fire activity over the last ca. 300 years, Langohr shows little increase in charcoal influx in the most recent decades, suggesting that local site conditions may limit fire activity as conditions warm.
At ca. 2000 cal yr BP, a synchronous rise in fire-episode frequency and/or CHAR occurred at many sites regardless of overall trends in fire activity. In the northern GYE, Slough Creek Pond, Crevice Lake, Fairy Lake, Dailey Lake, and Langohr Wetland had increased CHAR and/or fire-episode frequency, coincident with increased fire-related debris flows in northern YNP (Meyer et al., Reference Meyer, Wells and Timothy Jull1995). Sites in northwestern Montana also had an increase in fire-episode frequency at this time (Brunelle et al., Reference Brunelle, Whitlock, Bartlein and Kipfmueller2005; Alt et al., Reference Alt, McWethy, Everett and Whitlock2018; McWethy et al., Reference McWethy, Alt, Argiriadis, Battistel, Everett and Pederson2020). The increase in fires occurs at the same time as increased marl production in Blacktail Pond between ca. 2900 and 1300 cal yr BP (Huerta et al., Reference Huerta, Whitlock and Yale2009), diatom assemblages at Crevice Lake that suggest shorter springs and drier conditions between ca. 2100 and 850 cal yr BP (Whitlock et al., Reference Whitlock, Dean, Fritz, Stevens, Stone, Power, Rosenbaum, Pierce and Bracht-Flyr2012), and tree-ring evidence of pronounced drought in the Upper Colorado River Basin at ca. 2000 cal yr BP (Gangopadhyay et al., Reference Gangopadhyay, Woodhouse, McCabe, Routson and Meko2022). This suggests a regional shift in climate to warm, dry conditions.
Indigenous peoples may have contributed to the increase in fire recorded at ca. 2000 cal yr BP as well. Archaeological evidence from Montana and the northern Rocky Mountains suggests an increase in human population in the Late Holocene (MacDonald, Reference MacDonald2012), and analysis of radiocarbon-dated cultural materials suggests a peak in population at ca. 2000 cal yr BP (Kelly et al., Reference Kelly, Surovell, Shuman and Smith2013, Reference Kelly, Mackie, Robinson, Meyer, Berry, Boulanger and Codding2022). The drier conditions would have facilitated fire spread for both human and natural ignitions.
Across the northern GYE region, fire-episode frequencies varied during the MCA and LIA, although there is little evidence for vegetation change associated with changes in fire activity (Fig. 5). Slough Creek Pond featured more stand-replacing fires during the MCA at ca. 1000–700 cal yr BP (Millspaugh et al., Reference Millspaugh, Whitlock, Bartlein and Wallace2004), and fire-related sedimentation events in northern YNP increased between 1300 and 750 cal yr BP as well (Meyer et al., Reference Meyer, Wells and Timothy Jull1995). In contrast, charcoal data from Langohr Wetland showed a decrease in fire episodes during the MCA between ca. 1200 and 800 cal yr BP, which is also noted at Blacktail Pond, Cygnet Lake, and Fairy Lake (Fig. 5). However, Slough Creek, Cygnet Lake, and Fairy Lake also saw slightly decreased levels of CHAR during the LIA, when cold conditions may have inhibited fires.
One period of interest in the Langohr fire record is the long approximately 1500-year period from ca. 4500–3000 cal yr BP with little to no fire activity. Fairy Lake also shows evidence of fewer fires during this time period (Fig. 5). Cooler, wetter winter conditions are inferred independently from 18O isotope and ice accumulation records from the Beartooth Plateau (Chellman et al., Reference Chellman, Pederson, Lee, McWethy, Puseman, Stone, Brown and McConnell2021), an increase in avalanches or landslides at Fairy Lake (Benes et al., Reference Benes, Iglesias and Whitlock2019), and glacial readvances in western Canada and the Canadian Rockies (Menounos et al., Reference Menounos, Osborn, Clague and Luckman2009). Beyond the GYE, fire activity also decreased in northwestern Montana (Alt et al., Reference Alt, McWethy, Everett and Whitlock2018; McWethy et al., Reference McWethy, Alt, Argiriadis, Battistel, Everett and Pederson2020). It is possible that the more northern and higher elevation locations of Langohr Wetland and Fairy Lake were more susceptible to the Late Holocene cooling conditions that were seen in northwestern Montana and the Canadian Rockies, whereas more southern and lower elevation sites in the GYE were not.
Conclusions
The pollen and charcoal data from Langohr Wetland document changes in vegetation and fire activity at middle elevations in the northern GYE in response to millennial-scale changes in climate variability over the last 13,500 years. The record highlights the relative stability of vegetation composition and structure across the late glacial and Holocene despite variability in fire activity. The sequence of vegetation changes starts with the early establishment of tundra–steppe following deglaciation and the development of predominantly Picea parkland after 13,000 cal yr BP. Open mixed-conifer forest of Pinus with some Picea and Pseudotsuga was established in the Early Holocene at ca. 9300 cal yr BP, when conditions were warmer and drier than today. This period was followed by establishment of a closed mixed-conifer forest of Pinus, Picea, and Pseudotsuga during the Mid-Holocene, when conditions were cooler and wetter than before. The forest became most dense during the Late Holocene.
Following the establishment of mixed-conifer forest at ca. 9300 cal yr BP, episodes of high-severity fire occurred 1–2 times per millennium, and the frequency increased during the Late Holocene when dense mixed-conifer forest was present. The mixed-conifer forest experienced infrequent stand-replacing fires throughout the entire record (one fire episode approximately every 600 years in the Early Holocene and approximately every 350 years in the Late Holocene). The Langohr fire return interval is longer than expected for most mixed-conifer forests in the northern Rocky Mountains (Baker, Reference Baker2009), although the charcoal record likely only recorded stand-replacing fires adjacent to the wetland and not broader fire trends.
The sequence of environmental change that occurred at Langohr Wetland in response to long-term climate trends generally corresponds to other sites in the region, and the Langohr record provides insights into the location of glacial refugia and fire-facilitated vegetation transitions (Krause et al., Reference Krause and Whitlock2017). The early appearance of Pinus, Picea, and Pseudotsuga during late-glacial warming suggests lower elevations near Langohr Wetland, such as the Gallatin Valley, served as refugia during the full-glacial period. As the climate warmed, these conifers were able to quickly colonize the foothills and mountains surrounding the valley. Additionally, increased fire activity during the late-glacial period facilitated a transition from Picea parkland to an open Pinus-dominated forest (mixed-conifer at Langohr), a pattern also observed at Dailey Lake, Blacktail Pond, Slough Creek Pond, and Cygnet Lake.
The Langohr Wetland vegetation history also has notable differences from reconstructions at other sites. Persistent high percentages of conifer pollen from the Early Holocene to the present contrasts with other sites, which show more dynamic forest responses through time, particularly from ca. 10,000–6000 cal yr BP. The complacency of the forests near Langohr Wetland suggests that the site was less affected by Early Holocene summer drought conditions than other locations. The Langohr record also indicates a shift towards wet conditions earlier than other sites during the Mid-Holocene at ca. 7000–4500 cal yr BP, consistent with its generally mesic setting.
Comparisons of a network of paleofire records across the region, while generally varying over smaller temporal scales, show some synchronous trends and times of high fire activity, such as an increase in fires at the start of the Early Holocene (ca. 11,000–10,000 cal yr BP), and in the Mid-Holocene (7000–5000 cal yr BP). They also recorded decreases in fire activity with the transition from forest to open forest/parkland (although at differing times). Variation in the timing of changes in vegetation and fire activity was likely influenced by atmospheric patterns and the associated spatial heterogeneity in the delivery of moisture, as well as site-to-site differences in local conditions (topography, substrate, elevation, and microclimate). The Langohr record shares most similarities in fire trends with nearby Fairy Lake, including an increase in fire activity as forests became more dense during the Mid-Holocene (after ca. 6000 cal yr BP), and an increase in fire activity when present-day closed mixed-conifer forest established (after ca. 3000 cal yr BP).
The fire record from Langohr Wetland is unique in that it generally records infrequent fire episodes compared to other sites in the region. The relatively low fire occurrence suggests that the forest surrounding this wetland may have been less influenced by droughts that affected the region more broadly. These mesic settings will likely serve as important fire refugia for fire-sensitive conifers, such as Picea and Abies, as temperatures increasingly warm into the future.
Acknowledgements
We are grateful for field assistance from S. Ewing, F. Miller, and J. Wendt and laboratory assistance from C. VanderMolen. We acknowledge funding for this project from Montana State University and National Science Foundation grant EAR-2149482 to C.W. and D.B.M and BCS-1832486 (www.nsf.gov). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could appear to have influenced the work reported in this paper.








 Open access
Open access