



Introduction
Traditional population-level dietary guidelines, largely derived from Western epidemiological and clinical research, have substantially contributed to global public health.(Reference Antwi1,Reference Ordovas, Ferguson and Tai2) However, these guidelines implicitly assume that the physiological responses to dietary components are broadly uniform across populations.(Reference Bailey and Stover3,Reference Ma and Chan4) Growing evidence challenges this assumption, demonstrating that metabolic responses to macronutrient composition, micronutrient quality, and overall dietary patterns differ considerably according to genetic architecture, gut microbiome composition, habitual diet, lifestyle environments, and cultural factors.(Reference Ramos-Lopez, Martinez and Milagro5–7) These biological and environmental determinants shape postprandial glucose excursions, lipid handling, inflammatory pathways, and metabolic flexibility, all of which critically influence long-term metabolic health outcomes.(Reference Berry, Valdes and Drew8,Reference van Ommen, Keijer and Heil9)
The advent of omics technologies has transformed our understanding of interindividual variability.(Reference Tebani and Bekri10,Reference de Toro-Martín, Arsenault and Després11) Rapid advances in genomics, metabolomics, proteomics, and metagenomics have enabled the high-resolution characterization of molecular pathways linking diet to health outcomes.(Reference Ramos-Lopez, Martinez and Milagro5,7,Reference Wu, Oniani and Shao12) Omics profiling has revealed nutrient-specific metabolic reactions, diet-sensitive gene–environment interactions, and early biochemical perturbations that precede clinically detectable diseases.(Reference Berry, Valdes and Drew8,Reference van Ommen, Keijer and Heil9,Reference Qiu, Cai and Yao13) These tools provide unprecedented opportunities to identify individuals who may benefit from targeted dietary strategies and to refine public health recommendations based on biological responsiveness.(Reference Antwi1,Reference Bailey and Stover3)
These advances are particularly relevant for Asian populations, who face a disproportionately high burden of metabolic diseases despite having a lower average BMI than Western populations.(7,14,Reference Yoon, Lee and Kim15) East Asians typically exhibit greater visceral adiposity, reduced β-cell reserve, and heightened insulin resistance at comparatively lower BMI thresholds.(Reference Ma and Chan4,7,Reference Yoon, Lee and Kim15) South Asians experience an even earlier onset of cardiometabolic risk, frequently driven by high hepatic fat sensitivity, impaired mitochondrial oxidation, and carbohydrate-dominant dietary patterns.(Reference Misra and Vikram16,Reference Bineid, Ventura and Samidoust17) Southeast Asian populations show additional variability, reflecting diverse dietary traditions, food processing methods, and environmental exposures.(Reference Popkin, Adair and Ng18)
Asia’s exceptional dietary diversity – from rice-centred patterns in Northeast and Southeast Asia to lentil-, wheat-, and millet-based diets in South Asia – contributes to population-specific metabolic signatures.(Reference de Toro-Martín, Arsenault and Després11,Reference Popkin, Adair and Ng18) Likewise, genomic studies have identified Asian-enriched variants affecting lipid metabolism, adipogenesis, cholesterol transport, and glucose regulation.(Reference Bineid, Ventura and Samidoust17,Reference Ma, Lee and Kim19–Reference Huang, Han and Jang23) These genetic and cultural factors underscore the importance of population-specific evidence in guiding precision nutrition strategies.
In this review, we synthesize current metabolomic and genomic evidence from Asian populations, describe nutrient–metabolite mechanisms relevant to dietary guidance, and outline a translational framework for implementing precision nutrition in diverse Asian healthcare and community settings.
Biological basis for precision nutrition in Asians
Developing precision nutrition strategies tailored to Asian populations requires a detailed understanding of their distinct metabolic and physiological characteristics. Compared with Western populations, Asian populations often exhibit a constellation of traits that increase susceptibility to obesity, metabolic syndrome, and type 2 diabetes.(Reference Ma and Chan4,7) These traits include increased visceral adiposity, altered adipokine secretion, lower lean mass, reduced metabolic flexibility, and diminished β-cell functional reserve.(Reference Ma and Chan4,7,Reference Yoon, Lee and Kim15)
Visceral adiposity and inflammatory signalling
Even at BMI levels considered normal by Western standards, many Asians accumulate a greater proportion of visceral adipose tissue.(7,Reference Popkin, Adair and Ng18) Visceral adipocytes are metabolically active and are characterized by elevated lipolytic activity and high secretion of pro-inflammatory cytokines, such as tumour necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6).(Reference Hotamisligil24,Reference Shoelson, Lee and Goldfine25) These cytokines impair the phosphorylation of insulin receptor substrate-1 (IRS-1) and disrupt downstream phosphatidylinositol 3-kinase (PI3K)–protein kinase B (Akt) signalling, initiating early insulin resistance and promoting hepatic gluconeogenesis.(Reference Shoelson, Lee and Goldfine25,Reference Samuel and Shulman26) Elevated circulating free fatty acids (FFAs) from visceral lipolysis further exacerbate hepatic lipid accumulation and impair muscle glucose uptake.(Reference Samuel and Shulman26,Reference Boden27)
Reduced β-cell reserve and glycemic vulnerability
Multiple studies have indicated that East and South Asians have lower β-cell compensatory capacity, rendering them more vulnerable to postprandial hyperglycaemia when exposed to high-glycemic diets.(Reference Ma and Chan4,Reference Misra and Vikram16) Declines in early-phase insulin secretion appear earlier and more prominently than in Western populations,(Reference Misra and Vikram16) suggesting a pathophysiological trajectory characterized by impaired secretion rather than insulin resistance alone.(Reference DeFronzo28)
Genetic susceptibility and pathway modulation
Genetic architecture contributes substantially to the distinct metabolic vulnerability observed in many Asian populations. In addition to previously described Asian-enriched variants, such as polymorphisms in TMEM182, NPC1L1, APOA5, CETP, and inflammatory pathway genes,(Reference Ma, Lee and Kim19–Reference Huang, Han and Jang23) multiple loci robustly associated with type 2 diabetes and glycemic traits further clarify the biological underpinnings of cardiometabolic risk in this region. Variants in KCNQ1, TCF7L2, and SLC30A8, repeatedly replicated in East Asian and multi-ancestry genome-wide association studies (GWASs), are particularly informative in this context.(Reference Yasuda, Miyake and Horikawa29–Reference Sladek, Rocheleau and Rung31) Their utility in Asian cohorts is further shaped by ancestry-related differences in allele frequencies and linkage disequilibrium, which may affect effect-size transferability from European studies.
These loci underscore the prominent contribution of impaired β-cell function to diabetes susceptibility at lower BMI levels in many Asian groups. KCNQ1 has demonstrated strong and reproducible associations with type 2 diabetes in East Asian populations and is thought to influence insulin secretion through modulation of β-cell electrophysiology and incretin-related pathways.(Reference Yasuda, Miyake and Horikawa29) TCF7L2, one of the most consistently replicated genetic signals for type 2 diabetes across ancestries, affects incretin action and insulin secretion, thereby contributing to postprandial glycemic dysregulation.(Reference Grant, Thorleifsson and Reynisdottir30) SLC30A8, which encodes the β-cell zinc transporter ZnT8, plays a central role in insulin granule formation and glucose-stimulated insulin secretion; functional variation at this locus may modify β-cell resilience under metabolic stress.(Reference Sladek, Rocheleau and Rung31) In populations characterized by limited β-cell compensatory capacity, these variants may amplify glycemic vulnerability when dietary patterns are dominated by refined carbohydrates and high glycemic load.
In addition to glycemic regulation, loci related to micronutrient handling and lipid metabolism provide additional mechanistic anchors for population-tailored precision nutrition. Variants in FUT2 and FUT6 influence intestinal glycosylation patterns and host–microbiome interactions, with downstream effects on nutrient absorption, microbial metabolite production, and inflammatory signalling.(Reference Hazra, Kraft and Selhub32) Given the marked regional diversity in fermented foods, fibre sources, and carbohydrate staples across Asia, such variation may contribute to interindividual differences in postprandial metabolic responses. BCMO1, which regulates the conversion efficiency of β-carotene to retinoids, may alter retinoid-mediated transcriptional pathways involved in lipid oxidation and glucose metabolism.(Reference Leung, Hessel and Méplan33) In dietary contexts where provitamin A carotenoids constitute a major source of vitamin A, genetic variability in BCMO1 could meaningfully influence metabolomic profiles and metabolic risk trajectories.
Similarly, polymorphisms in the fatty acid desaturase cluster (FADS1/2) affect endogenous long-chain polyunsaturated fatty acid (PUFA) biosynthesis, circulating lipidomic signatures, and inflammatory tone.(Reference Schaeffer, Gohlke and Müller34) Because dietary fat composition varies considerably across Asian regions – ranging from marine-derived n−3 fatty acids to seed- and palm-oil–based fats – FADS-related variation may modify cardiometabolic responses to fat quality and n−3/n−6 balance, supporting a genotype-informed approach to dietary fat modulation.
Translationally, these genetic determinants gain clinical relevance when integrated with dynamic omics-derived intermediate phenotypes, including postprandial glycemic excursions assessed by continuous glucose monitoring (CGM), lipidomic profiles reflecting PUFA balance, and biomarkers of oxidative stress.(Reference Ramos-Lopez, Martinez and Milagro5–7,Reference Tebani and Bekri10) Genotype-informed risk stratification may therefore help identify biologically coherent responder subgroups. For example, individuals with elevated β-cell–related genetic susceptibility (e.g. KCNQ1-, TCF7L2-, or SLC30A8-implicated profiles) may derive particular benefit from dietary strategies emphasizing glycemic load reduction and improved carbohydrate quality. Conversely, those carrying variants affecting PUFA biosynthesis (FADS1/2) or nutrient absorption and metabolism (e.g. FUT2/6, BCMO1) may warrant more precise modulation of fatty acid composition or carotenoid-rich food intake. Such integrative approaches strengthen the mechanistic and translational foundation of precision nutrition strategies tailored to heterogeneous Asian populations.
Collectively, these genetic determinants illustrate how population-specific variation in β-cell function, micronutrient handling, and lipid metabolism can shape metabolic responses to diet in Asian populations. The integration of genotype information with intermediate omics phenotypes provides a mechanistic basis for developing biologically stratified precision nutrition strategies, as summarized in Figure 1.
Multi-omics precision nutrition framework for Asian populations. Asian populations exhibit distinct metabolic features, including increased visceral adiposity, reduced β-cell reserve, and culturally specific dietary patterns. Multi-omics profiling (genomics, metabolomics, microbiome, and oxidative stress markers) captures these traits and their responses to diet. The integration of nutrient–metabolite mechanisms and genetic–metabolomic risk models enables the stratification of individuals into differential risk and response groups, supporting culturally adapted precision nutrition interventions in Asian healthcare and community settings.

Figure 1. Long description
The diagram illustrates the multi-omics precision nutrition framework for Asian populations. It starts with distinct Asian metabolic features such as increased visceral adiposity, reduced beta-cell reserve, and cultural dietary patterns. This leads to multi-omics profiling, which includes genomics, metabolomics, microbiome, and oxidative stress markers. The nutrient-metabolite mechanisms section describes how dietary components like fiber, legumes, refined grains, and oils affect metabolic processes. The integrated genetic-metabolomic risk models section explains the use of genetic risk scores and metabolite panels for risk classification and early detection of metabolic dysfunction. Finally, the precision nutrition interventions in Asia section outlines personalized dietary patterns, omics-informed control, digital health integration, and culturally adapted strategies.
Metabolomic biomarkers in Asian populations
Metabolomics provides a dynamic representation of an individual’s biochemical state, reflecting recent dietary intake, metabolic stress, microbiome activity, and environmental exposure.(Reference van Ommen, Keijer and Heil9,Reference Tebani and Bekri10,Reference Qiu, Cai and Yao13,Reference Vo and Trinh35) This sensitivity to short- and medium-term metabolic fluctuations makes metabolomic profiling particularly valuable for investigating the risk of metabolic diseases in Asian populations.
Lipid-derived metabolites and oxidative stress
A consistent finding across Asian metabolomics studies is the elevation of lipid-derived metabolites, particularly lysophosphatidylcholines (lysoPCs).(Reference Kim, Yoo and Ko36–Reference Knuplez and Marsche38) LysoPCs are generated through phospholipase A2–mediated cleavage of phosphatidylcholines and serve as biomarkers of oxidative stress, membrane turnover, and inflammatory activation.(Reference van Ommen, Keijer and Heil9,Reference Law, Chan and Marathe37,Reference Liu, Zhu and Chen39) In multiple Korean cohorts, metabolically unhealthy overweight individuals exhibited elevated lysoPC concentrations, higher levels of F2-isoprostanes, and reduced adiponectin levels.(Reference Park, Yoo and Jee22,Reference Kim, Yoo and Ko36) Mechanistically, lysoPCs impair mitochondrial efficiency, promote endothelial dysfunction, and activate proinflammatory cytokine pathways, contributing to early insulin resistance and increased vascular stress.(Reference Law, Chan and Marathe37–Reference Hung, Chan and Chan40)
Carbohydrate-related metabolites and glycemic-excursion markers
Carbohydrate-related metabolites can provide sensitive readouts of postprandial dysglycemia, which is particularly relevant in Asian settings, where refined grains and high glycemic-load staples remain common, and β-cell reserve may be comparatively limited. In addition to fasting glucose and HbA1c, 1,5-anhydroglucitol (1,5-AG) is a well-validated short-term marker of glycemic excursions because it decreases when plasma glucose exceeds the renal threshold, capturing postprandial spikes that may be missed by longer-term averages.(Reference Ku, Zhong and Song41) Evidence from East Asian clinical studies further supports its usefulness in reflecting postprandial hyperglycaemia and glycemic variability.(Reference Won, Park and Park42,Reference Seok, Huh and Kim43) Additional markers linked to early glycemic stress and reduced metabolic flexibility include lactate, α-hydroxybutyrate (α-HB), and plasma mannose, which have been associated with insulin resistance and incident type 2 diabetes in prospective analyses.(Reference Gall, Beebe and Lawton44–Reference Mardinoglu, Stančáková and Lotta46) Reactive dicarbonyls (e.g. methylglyoxal and 3-deoxyglucosone) and polyol-pathway metabolites (e.g. sorbitol and fructose) may also indicate carbonyl/redox stress under sustained hyperglycaemia.(Reference Brings, Fleming and Freichel47,Reference Yan48) Integrating excursion-sensitive biomarkers (e.g. 1,5-AG) with intermediary metabolism and glycation/redox-stress readouts can help identify ‘carbohydrate responders’ and prioritize interventions targeting carbohydrate quality and meal structure to blunting glucose peaks.
Microbiome-related metabolites
Microbiome-derived metabolites are particularly informative in Asia because fermented-food practices and heterogeneous fibre sources can reshape gut microbial ecology and downstream metabolite profiles.(Reference Leeuwendaal, Stanton and O’Toole49) SCFAs – including acetate, propionate, and butyrate – are produced through microbial fermentation of dietary fibre and can influence host metabolism via host–microbe signalling and enteroendocrine pathways.(Reference Kimura, Ozawa and Inoue50,Reference Koh, De Vadder and Kovatcheva-Datchary51) In parallel, trimethylamine N-oxide (TMAO), generated from microbial metabolism of dietary choline- and carnitine-containing substrates followed by hepatic oxidation, has been reported to be associated with adverse cardiovascular outcomes in experimental models and prospective human studies, including growing evidence from Asian cohorts.(Reference Qi, You and Li52–Reference Wang, Klipfell and Bennett54) In addition, bile acid pools are increasingly recognized as diet- and microbiome-sensitive endocrine signals through receptors such as the farnesoid X receptor (FXR) and Takeda G-protein–coupled receptor 5 (TGR5), which regulate lipid and carbohydrate metabolism and inflammatory tone, making them attractive candidates for targeted metabolite panels in precision nutrition.(Reference Staley, Weingarden and Khoruts55,Reference Fleishman and Kumar56) To avoid redundancy, detailed dietary determinants (e.g. specific fibres and fermented foods) are discussed further in the dietary-mechanisms section below.
Micronutrient-related biomarkers within metabolomics
Micronutrient-related biomarkers can be integrated into metabolomic panels or assayed alongside them to reflect nutrient status that is clinically actionable and regionally relevant, particularly in Asia, where dietary sources and food environments vary substantially across subregions. Among micronutrients, vitamin A derivatives – particularly retinal and retinoic acid – provide a compelling example of nutrient-linked biochemistry that can inform early metabolic risk stratification; in Korean adults, they have emerged as predictive biomarkers.(Reference Han, Yang and Kim57) These metabolites regulate hepatic lipid oxidation, gluconeogenesis, adipocyte differentiation, and insulin sensitivity through RAR/RXR-mediated transcriptional control.(Reference Desvergne, Michalik and Wahli58,Reference Li, Cai and Boyer59) Elevated retinoid metabolite levels predict the progression from impaired fasting glucose to type 2 diabetes more accurately than conventional markers, such as fasting glucose or HbA1c.(Reference Han, Yang and Kim57) When combined with oxidative stress markers such as oxidized LDL and 8-epi-prostaglandin F2α (8-epi-PGF2α), retinoid profiles offer a sensitive readout of metabolic stress that precedes clinical deterioration.(Reference Park, Yoo and Jee22,Reference Kim, Yoo and Ko36,Reference Han, Yang and Kim57)
In addition to vitamin A, vitamin B12-related biomarkers may be especially relevant in South Asian settings, where vegetarian dietary practices are common, and functional deficiency can co-occur with hyperhomocysteinemia.(Reference Yajnik, Deshpande and Lubree60–Reference Kurpad, Varadharajan and Aeberli62) Evidence from Indian cohorts (e.g. the Pune Maternal Nutrition Study) links low vitamin B12 and functional deficiency markers (including methylmalonic acid [MMA]) with higher offspring insulin resistance, supporting a life-course role of one-carbon metabolism in cardiometabolic risk.(Reference Yajnik, Deshpande and Jackson63,Reference Gammon, von Hurst and Coad64) Functionally, MMA and homocysteine can complement serum B12 to capture functional B12 insufficiency, strengthening risk stratification in settings where dietary exposure patterns differ markedly across Asian subregions.(Reference Wolffenbuttel, Wouters and de Jong65,Reference Vashi, Edwin and Popiel66) Finally, iodine status – highly salient in parts of Asia due to seafood/seaweed exposures and salt iodization variability – interfaces with thyroid hormone biology and can influence lipid metabolism; emerging reviews also highlight metabolomic shifts under iodine perturbation.(Reference Zhang, Shang and Liu67,Reference Gräfe, Scheibe and Schwarz68)
Fatty acid profiles as diet-responsive metabolomic readouts
Fatty acid profiles provide diet-responsive metabolomic readouts that capture both dietary fat exposure and endogenous remodelling, with clear relevance for Asian populations, where staple diets and fat sources vary widely. The circulating and erythrocyte membrane eicosapentaenoic acid (EPA) + docosahexaenoic acid (DHA) (the Omega-3 Index) reflect longer-term marine n−3 exposure and have been proposed as risk-relevant indices for coronary outcomes, while mechanistic and clinical literature supports the anti-inflammatory and pro-resolving actions of marine n−3 fatty acids.(Reference Harris and Von Schacky69,Reference Calder70) Importantly, fatty acid signatures can inform precision nutrition not only through ‘intake-status’ markers (e.g. EPA/DHA) but also through endogenous pathway markers that link carbohydrate-heavy diets to lipid dysregulation. Excess carbohydrates can be converted in the liver into palmitate (16:0) and desaturated to palmitoleate (16:1n–7); accordingly, de novo lipogenesis (DNL) proxy indices derived from very LDL-triacylglycerol fatty acid patterns have been used to approximate hepatic lipogenic remodelling and tend to increase under high-carbohydrate feeding, alongside increases in plasma triglycerides.(Reference Chong, Hodson and Bickerton71–Reference Lee, Lai and de Oliveira Otto73) However, these proxies should be interpreted in the context of the diet (e.g. palm-oil-derived palmitate) and, where feasible, complemented by postprandial designs or hepatic-output–enriched fractions.(Reference Rosqvist, McNeil and Pramfalk74) In practice, combining omega-3 status markers with DNL-related signatures can help distinguish individuals, whose risk biology is driven more by fat quality versus carbohydrate-to-fat conversion, thereby supporting culturally tailored recommendations in Asian populations.
Population-specific metabolomic signatures
Metabolomic fingerprints vary considerably across Asian subpopulations due to differences in genetic background, gut microbiome composition, habitual dietary substrates (including staple carbohydrates, fermentation practices, and regional fat sources), and environmental exposure.(Reference Popkin, Adair and Ng18,Reference van Valkengoed, Argmann and Ghauharali-van der Vlugt75,Reference Kim, Lee and Hur76) Importantly, these differences extend beyond lipid- and amino-acid patterns to include microbiome-derived metabolites (e.g. SCFAs, bile-acid derivatives, and TMAO),(Reference Koh, De Vadder and Kovatcheva-Datchary51,Reference Wang, Klipfell and Bennett54,Reference Fleishman and Kumar56) carbohydrate-related markers of glycemic stress,(Reference Ku, Zhong and Song41,Reference Gall, Beebe and Lawton44–Reference Mardinoglu, Stančáková and Lotta46) and fatty acid profiles reflecting both intake (omega-3/omega-6 balance) and endogenous remodelling (e.g. DNL-related signatures).(Reference Harris and Von Schacky69–Reference Rosqvist, McNeil and Pramfalk74) In some settings, these metabolite signatures can be further contextualized by micronutrient-related functional indicators that track one-carbon metabolism (e.g. vitamin B12–related MMA/homocysteine patterns).(Reference Yajnik, Deshpande and Lubree60,Reference Yajnik, Deshpande and Jackson63,Reference Wolffenbuttel, Wouters and de Jong65)
-
• East Asians often exhibit metabolite–nutrient interaction patterns at the population level, including phospholipid and amino acid signatures that track rice-and fermented food-rich diets; these patterns may coincide with microbiome-derived metabolite differences.(Reference Kim, Lee and Hur76,Reference Shibutami, Ishii and Harada77)
-
• South Asians exhibit characteristic elevations in branched-chain amino acids (BCAAs) and acylcarnitines in multi-ethnic analyses, consistent with early metabolic inflexibility and mitochondrial substrate-handling differences.(Reference van Valkengoed, Argmann and Ghauharali-van der Vlugt75,Reference Muilwijk, Goorden and Celis-Morales78,Reference Tai, Tan and Stevens79) In subgroups, functional vitamin B12 insufficiency and elevated homocysteine levels may further modify risk profiles, aligning with long-standing vegetarian dietary traditions in parts of South Asia.(Reference Yajnik, Deshpande and Lubree60,Reference Gammon, von Hurst and Coad64)
-
• Southeast Asians can display distinct lipidomic and fatty acid features in multi-ethnic settings, and regional culinary fat sources (e.g. coconut- and palm-based oils) have been evaluated in human studies for their effects on cardiometabolic markers,(Reference Swarnamali, Ranasinghe and Jayawardena80–Reference Begum, Torta and Narayanaswamy82) requiring careful metabolomic interpretation to distinguish dietary saturated fat intake from endogenous, carbohydrate-driven DNL where relevant.(Reference Rosqvist, McNeil and Pramfalk74)
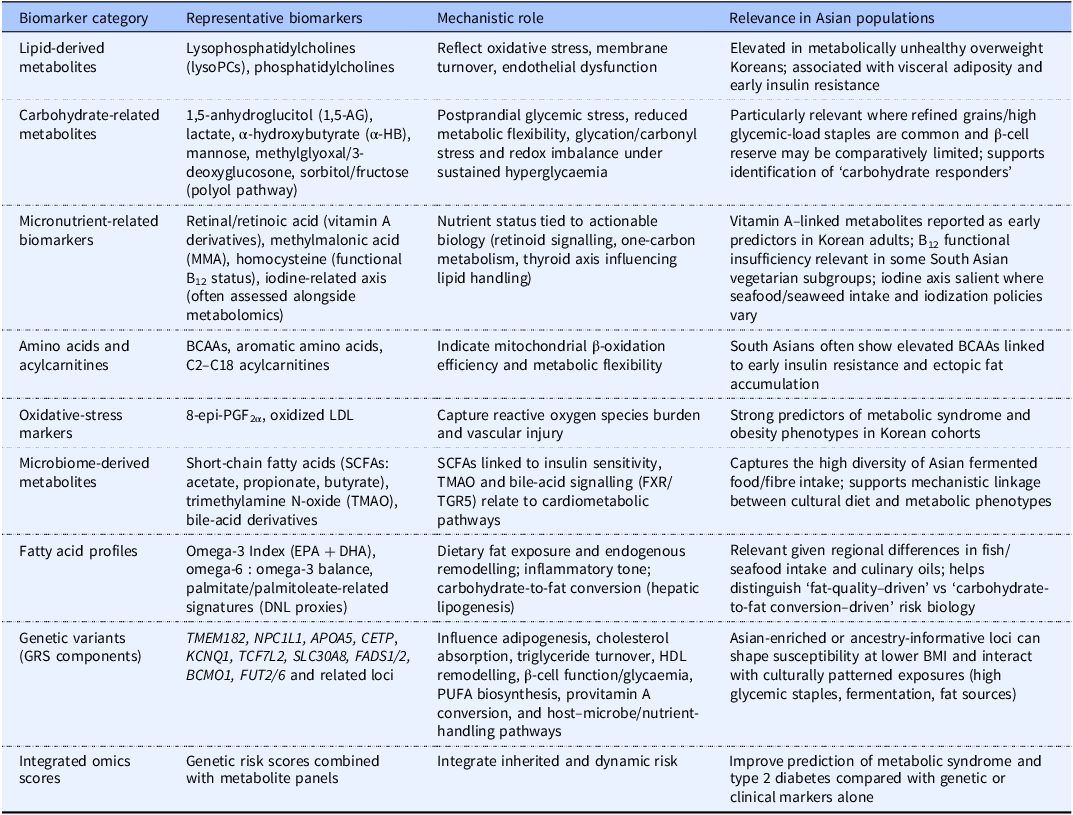
These population-specific patterns highlight the need to establish regionally appropriate reference ranges rather than relying on Western-centric metabolomic interpretations of the data. Table 1 summarizes the key metabolomic and genomic biomarkers that are particularly relevant to precision nutritional approaches in Asian populations.
Key metabolomic and genomic biomarkers relevant to precision nutrition in Asian populations

Table 1. Long description
The table presents key metabolomic and genomic biomarkers relevant to precision nutrition in Asian populations, emphasizing the need for regionally appropriate reference ranges. It includes categories such as lipid-derived metabolites, carbohydrate-related metabolites, micronutrient-related biomarkers, amino acids and acylcarnitines, oxidative-stress markers, microbiome-derived metabolites, fatty acid profiles, genetic variants, and integrated omics scores. Each category lists representative biomarkers, their mechanistic roles, and their relevance in Asian populations. For example, lipid-derived metabolites like lysophosphatidylcholines are elevated in metabolically unhealthy overweight Koreans and are associated with visceral adiposity and early insulin resistance. Carbohydrate-related metabolites such as 1,5-anhydroglucitol and lactate reflect postprandial glycemic stress and reduced metabolic flexibility. The table also highlights the importance of micronutrient-related biomarkers like vitamin A derivatives and methylmalonic acid, which are tied to nutrient status and actionable biology. Amino acids and acylcarnitines indicate mitochondrial beta-oxidation efficiency, while oxidative-stress markers capture reactive oxygen species burden and vascular injury. Microbiome-derived metabolites like short-chain fatty acids are linked to insulin sensitivity and bile-acid signaling. Fatty acid profiles, genetic variants, and integrated omics scores further contribute to understanding dietary fat exposure, endogenous remodelling, and inherited risk. The table underscores the diversity of Asian populations and the need for tailored nutritional approaches.
Dietary phenotypes and nutrient–metabolite mechanisms in Asian populations
Dietary exposure in Asian populations is not defined solely by macronutrient composition but by culturally embedded patterns that shape staple food selection, preparation techniques, fermentation practices, meal structure, and habitual food combinations. These dietary phenotypes influence glycemic load, fatty acid balance, sodium intake, fibre availability, and phytochemical exposure, thereby generating population-specific metabolomic and microbiome configurations. In carbohydrate-dominant dietary settings common across East and South Asia, metabolomics provides mechanistic insight into how these culturally patterned nutrient exposures translate into lipidomic shifts, oxidative stress signatures, and alterations in amino acid metabolism.(Reference Koh, De Vadder and Kovatcheva-Datchary51,Reference den Besten, van Eunen and Groen83,Reference Nicholson, Holmes and Kinross84) Therefore, understanding dietary phenotypes at the pattern level is essential before interpreting nutrient-level mechanisms.(Reference Ordovas, Ferguson and Tai2) Pattern-level exposures provide the ecological context in which nutrient–metabolite interactions unfold.
Fish- and seafood-rich dietary patterns
Fish and seafood consumption is culturally central in many coastal East and Southeast Asian populations, often representing long-term habitual exposure rather than occasional intake. Marine-derived long-chain n−3 PUFAs, particularly EPA and DHA, modulate hepatic triglyceride synthesis, membrane phospholipid remodelling, and inflammatory resolution pathways.(Reference Calder85,Reference Jump, Depner and Tripathy86) Lipidomic profiling demonstrates that dietary n−3 intake is reflected in circulating triglyceride and phosphatidylcholine species, influencing downstream insulin signalling and inflammatory mediator balance.(Reference Quehenberger, Armando and Brown87)
However, metabolic consequences depend substantially on preparation practices. Salted, dried, or deep-fried seafood products may increase sodium intake(Reference Wu, Micha and Imamura88) and introduce oxidized lipid species, potentially attenuating cardiometabolic benefits.(Reference Falade, Oboh and Okoh89) Precision-nutrition frameworks in Asian contexts should therefore differentiate fresh marine-based dietary patterns from highly processed seafood preparations when interpreting lipidomic and inflammatory biomarkers.
Soy and traditional soy-based dietary exposures
Soybeans and traditional soy-based foods, including tofu, soy milk, tempeh, miso, doenjang, and natto, are culturally salient protein sources in East Asia. Epidemiological and clinical studies indicate that soy intake is associated with improved lipid profiles and reduced cardiovascular risk markers.(Reference Messina90,Reference Zhan and Ho91) Isoflavones, such as genistein and daidzein, influence endothelial function, lipid metabolism, and insulin sensitivity through oestrogen receptor–mediated transcriptional pathways and antioxidant effects.(Reference Messina90)
A defining feature of soy in precision nutrition is interindividual variability in isoflavone metabolism. Gut microbial conversion of daidzein to equol varies widely across individuals and populations.(Reference Setchell and Clerici92) Equol production status has been associated with differential metabolic and vascular responses to soy intake.(Reference Guadamuro, Delgado and Redruello93) Integration of dietary data with microbiome and metabolomic profiling thus enables the stratification of responders and non-responders within culturally consistent soy-based dietary patterns, illustrating how identical food exposure can generate divergent metabolomic phenotypes.
Plant-forward dietary patterns and carbohydrate quality
Many traditional Asian dietary patterns are plant-forward and characterized by a substantial intake of grains, vegetables, and legumes. When minimally processed, these diets provide dietary fibre and polyphenols that promote microbial fermentation and SCFA production.(Reference Koh, De Vadder and Kovatcheva-Datchary51,Reference Reynolds, Mann and Cummings94,Reference Sonnenburg and Bäckhed95) SCFAs bind to GPR41 and GPR43 and enhance insulin sensitivity and reduce hepatic lipogenesis,(Reference Nicholson, Holmes and Kinross84) and stimulate glucagon-like peptide-1 (GLP-1) secretion.(Reference Tolhurst, Heffron and Lam96)
However, carbohydrate quality and degree of refinement critically shape metabolic outcomes. Diets dominated by polished white rice and refined wheat products may increase postprandial glycemic excursions and exacerbate β-cell stress, particularly in Asian populations with reduced β-cell reserve.(Reference Ma and Chan4,Reference Hills, Arena and Khunti97) Metabolomic studies have linked high glycemic-load dietary patterns to elevations in BCAAs, acylcarnitines, and lipid species associated with impaired mitochondrial β-oxidation and early insulin resistance.(Reference Newgard98,Reference Wang, Larson and Vasan99) Thus, the classification of diets as ‘plant-based’ without accounting for processing level and meal composition may obscure important metabolic heterogeneity.
Nutrient–Metabolite mechanisms within cultural contexts
Within these broader dietary phenotypes, specific nutrient-level mechanisms provide mechanistic insights into how culturally congruent modifications can improve metabolic resilience.
-
• Dietary fibre and fermentation-derived metabolites. Soluble fibre is fermented by gut microbiota to generate SCFAs, including acetate, propionate, and butyrate, which regulate hepatic lipid metabolism, immune signalling, and insulin sensitivity.(Reference Koh, De Vadder and Kovatcheva-Datchary51) In Korean metabolomics studies, higher fibre intake has been associated with increased fatty acid β-oxidation, reduced lipid peroxidation, and improved phospholipid balance.(Reference Kim, Yoo and Ko36,Reference Hung, Chan and Chan40) These findings suggest that increasing fibre intake within rice-dominant dietary contexts may partially counteract oxidative and inflammatory metabolic phenotypes.
-
• Legume substitution and glycemic modulation. Legumes, including soybeans, lentils, and chickpeas, are traditional components of many Asian dietary patterns and represent culturally feasible substitution strategies. Randomized controlled trials (RCTs) in Korean women have demonstrated that replacing one-third of daily rice intake with legumes reduces oxidative stress biomarkers, increases adiponectin concentrations, and promotes modest weight reduction.(Reference Han, Kim and Lee100,Reference Kim, Jeung and Jeong101) Mechanistically, these changes are associated with enhanced AMP-activated protein kinase-mediated fatty acid oxidation and improved antioxidant capacity.(Reference Hardie, Ross and Hawley102) Such culturally aligned substitutions illustrate how traditional foods can be leveraged within precision nutrition frameworks to mitigate glycemic and lipid-related risk.
Conceptual integration
Collectively, fish-rich, soy-centred, and plant-forward dietary phenotypes generate distinct nutrient–metabolite configurations that shape cardiometabolic risk trajectories in Asian populations. By explicitly linking staple foods, preparation methods, and fermentation practices to candidate omics biomarkers, such as lipidomic signatures, SCFAs, amino acid profiles, and oxidative-stress indicators, precision-nutrition approaches can move beyond generalized dietary advice toward culturally embedded, mechanism-informed personalization strategies.
Integrated genetic–metabolomic risk models
Multi-omics integration provides a refined understanding of metabolic risk by combining genetic predisposition and dynamic metabolomic states. In Asian populations, where specific genetic variants and characteristic metabolic phenotypes frequently co-occur, integrated models offer substantial advantages over single-parameter assessments.
Genetic variants enriched in Asian populations
GWASs conducted in Korean, Chinese, and broader Asian cohorts have identified population-enriched variants that influence adiposity, lipid metabolism, and glycemic regulation.(Reference Oh, Lee and Shin21,Reference Ng, Park and Oh103–Reference Mahajan, Taliun and Thurner105) Variants in TMEM182, NPC1L1, APOA5, and CETP affect adipogenesis, cholesterol absorption, triglyceride metabolism, and high-density lipoprotein (HDL) remodelling.(Reference Ma, Lee and Kim19,Reference Cho, Huang and Han20,Reference Teslovich, Musunuru and Smith106,Reference Thompson, Lira and Durham107) These variants interact with nutrient-sensitive transcriptional regulators, including peroxisome proliferator-activated receptors (PPARα/γ), sterol regulatory element-binding proteins, and the liver X receptor, thereby shaping metabolic responses to dietary fats and high-glycemic carbohydrates.(Reference Desvergne, Michalik and Wahli58,Reference Ordovas and Corella108)
In East Asian populations, genetic risk scores (GRSs) incorporating population-enriched variants have demonstrated improved risk discrimination for cardiometabolic outcomes, such as type 2 diabetes, compared with scores derived from European datasets.(Reference Liu, Wang and Cui109,Reference Furukawa, Hara and Nishida110) In addition, weighted GRS approaches that account for variant-specific effect sizes have been shown to further enhance predictive accuracy in Asian cohorts,(Reference Park, Yoo and Jee22,Reference Huang, Han and Jang23) supporting the use of ancestry-informed weighting and calibration for risk stratification.
Metabolomic biomarkers as dynamic indicators
Metabolomic profiles complement genetic data by capturing dynamic and time-sensitive metabolic perturbations. Elevated lysoPC concentrations reflect phospholipid turnover, mitochondrial stress, and systemic inflammation.(Reference Law, Chan and Marathe37,Reference Knuplez and Marsche38) Altered amino acid signatures, including BCAAs, aromatic amino acids, and acylcarnitines, provide insights into mitochondrial β-oxidation efficiency, nitrogen metabolism, and microbiome–host interactions.(Reference Newgard, An and Bain111,Reference Shah, Kraus and Newgard112) Oxidative stress markers such as 8-epi-PGF2α offer an additional dynamic layer that reflects lifestyle, environmental exposure, and nutritional status.(Reference van Ommen, Keijer and Heil9,Reference Park, Yoo and Jee22)
Synergistic value of integrated models
Studies in Korean cohorts have demonstrated that genetic–metabolomic integration improves the prediction of metabolic syndrome, type 2 diabetes, and obesity phenotypes.(Reference van Ommen, Keijer and Heil9,Reference Oh, Lee and Shin21–Reference Huang, Han and Jang23) Multi-omics models capture the interactions between inherited susceptibility and modifiable metabolic conditions, enabling the precise identification of individuals likely to benefit from specific dietary interventions. For example, individuals with a high GRS for dyslipidemia may respond more favourably to legume substitution or reduced glycemic load when their metabolomic profiles concurrently exhibit markers of phospholipid dysregulation or heightened oxidative stress.(Reference Zeevi, Korem and Zmora6,Reference Park, Yoo and Jee22,Reference Ordovas and Corella108)
These findings underscore the value of moving beyond single-biomarker frameworks toward integrated, mechanistically informed risk assessments.
Translational framework for omics-guided precision nutrition in Asia
Translating multi-omics findings into clinical and public health practices requires the coordinated development of analytical infrastructure, workforce capacity, technological tools, and equity considerations. Asian countries vary widely in terms of laboratory resources, healthcare systems, and dietary environments, necessitating regionally adaptive approaches. A stepwise translational pipeline for omics-guided precision nutrition in Asia is shown in Figure 2.
Translational pipeline for omics-guided precision nutrition in Asia. The pipeline begins with discovery omics studies in Asian cohorts, followed by the validation and standardization of biomarker panels. Clinically feasible assays and decision support tools are then integrated into healthcare and digital health platforms. Implementation and scale-up efforts must address workforce training, cost-effectiveness, and equity to ensure that the benefits of precision nutrition are accessible across diverse Asian populations.

Figure 2. Long description
The flowchart outlines the translational pipeline for omics-guided precision nutrition in Asia. It starts with discovery and cohort studies involving multi-omics in Asian cohorts, including genomics, metabolomics, and microbiome analysis. The second stage focuses on biomarker selection and validation, identifying reproducible markers such as lysoPC, retinoid, amino-acid, SCFA markers, and Asian-specific genetic variants and GRS. The third stage involves clinical panel development, creating targeted metabolite panels and standardized assays and cut-offs. The fourth stage integrates clinical decision-support and digital health tools, including EHR-integrated decision tools, apps, CGM, and wearables for feedback. The final stage emphasizes implementation and scale-up in Asia, addressing training for clinicians and dietitians, cost-effectiveness, reimbursement, and equity across regions such as East, South, and Southeast Asia. The process includes an outcomes and feedback loop to reduce metabolic disease burden and continuously feed data back into discovery.
Analytical standardization and feasibility
A major barrier to the clinical adoption of metabolomics is the lack of cross-platform standardization.(Reference Nicholson and Lindon113,Reference Wishart114) Variability in sample processing, analytical platforms, and data interpretation limits the comparability of studies.(Reference Gika, Theodoridis and Wilson115–Reference Want, Masson and Michopoulos117) The development of standardized, clinically validated targeted metabolite panels offers a feasible solution. Such panels should prioritize biomarkers with established mechanistic relevance in Asian populations, including lysoPC species, retinoid metabolites, amino acid ratios, and oxidative stress markers.(Reference van Ommen, Keijer and Heil9)
Clinician training and decision-support systems
The effective use of omics data requires clinicians to understand metabolic pathways, gene–diet interactions, and biomarker interpretation. However, surveys indicate that many healthcare professionals report limited confidence in applying omics data to nutritional decision-making.(Reference Haga, Burke and Ginsburg118,Reference Manolio, Chisholm and Ozenberger119) Integrating omics education into medical and dietetics curricula, coupled with decision-support systems embedded within electronic health records, can reduce interpretive barriers and support the consistent implementation of precision nutrition.(Reference Williams, Taylor and Walton120,Reference Marsolo and Spooner121)
Digital-health integration
Wearable biosensors, CGM, mobile diet-tracking applications, and artificial intelligence (AI)-based diet analysis tools offer scalable platforms for personalized nutrition.(Reference Williams, Taylor and Walton120–Reference van den Brink, van den Broek and Palmisano122) Emerging microbiome-enabled extensions of these platforms now include app-linked, at-home gut microbiome testing with standardized reporting pipelines, enabling microbiome features to be incorporated into digital precision-nutrition workflows.(Reference Porcari, Mullish and Asnicar123,Reference Servetas, Gierz and Hoffmann124) These technologies can incorporate omics-derived biomarkers to generate adaptive dietary recommendations that are updated in response to real-time metabolic feedback. In particular, AI-driven microbiome–diet response prediction models that integrate gut microbiome profiles with dietary records and CGM data have been used to predict individualized postprandial responses and to support algorithm-guided dietary recommendations.(Reference Zeevi, Korem and Zmora6,Reference Berry, Valdes and Drew8) Digital feedback systems can also integrate microbiome-derived metabolites (e.g. SCFAs and related host–microbe co-metabolism signatures) to provide actionable, phenotype-linked coaching aligned with culturally patterned exposures such as fermented foods and variable fibre sources.(Reference Koh, De Vadder and Kovatcheva-Datchary51,Reference van den Brink, van den Broek and Palmisano122) Evidence and clinical validation levels vary across platforms, highlighting the need for harmonized standards and rigorous evaluation before broad implementation. Given current variability in consumer-facing microbiome testing, implementation should emphasize analytical standardization, clinical validation, privacy protection, and equitable access.(Reference Porcari, Mullish and Asnicar123,Reference Servetas, Gierz and Hoffmann124) Digital health integration is particularly promising in Asia, where mobile technology adoption is high and health systems are increasingly embracing telehealth.(Reference Lee, Cho and Kim125–Reference Macariola, Santarin and Villaflor127)
Equity and regional diversity
Despite these rapid advances, omics research in Asia remains concentrated in Korea, Japan, and China. South and Southeast Asian populations are underrepresented despite their high metabolic disease burden and diverse dietary exposure. Limited laboratory capacity, socioeconomic barriers, and variable access to healthcare resources further challenge the equitable implementation.(Reference Popejoy and Fullerton128–Reference Rotimi, Abayomi and Abimiku132) Ensuring affordability, cultural compatibility, and adequate representation in omics datasets is essential to prevent the widening of metabolic health disparities.
Future research priorities
Advancing precision nutrition in Asian populations requires strategic investments in multiple research areas. Several high-priority areas have emerged from recent evidence.
Longitudinal multi-omics cohorts
First, there is a clear need for longitudinal cohort studies that track metabolomic, genomic, microbiomic, and proteomic changes over time.(114,Reference Nicholson, Lindon and Holmes133–Reference Hasin, Seldin and Lusis136) Such studies will help determine the stability of metabolomic signatures, identify early biochemical shifts preceding metabolic deterioration, and clarify temporal relationships among diet, metabolic pathways, and disease risk. Continuous or repeated sampling is particularly important in Asian populations, where glycemic responses and oxidative stress can fluctuate dramatically according to dietary composition and environmental exposure.(Reference Zeevi, Korem and Zmora6,Reference Berry, Valdes and Drew8)
Systems-level integration of omics datasets
Integrating genomics, metabolomics, and microbiome data into cohesive system-level models will enable deeper mechanistic insights into nutrient–metabolite interactions.(Reference van Ommen, Keijer and Heil9,Reference Argilés, Busquets and López-Soriano137,Reference Misra, Langefeld and Olivier138) Computational approaches, such as network modelling, machine learning, and causal inference frameworks, can help map dietary exposures onto biological pathways and identify interaction nodes that may serve as targets for tailored dietary interventions.(Reference Libbrecht and Noble139,Reference Pearl140)
Representation of under-studied Asian populations
Future research should expand beyond East Asian cohorts to include South and Southeast Asian populations, who face disproportionately high metabolic disease burdens.(Reference Yoon, Lee and Kim15,Reference Hills, Arena and Khunti97,Reference Misra and Ganda141) These populations often exhibit early metabolic impairment, unique fat distribution patterns, and distinct dietary exposures that are not adequately represented in current omics datasets. Increasing the representation improves the accuracy, generalizability, and cultural relevance of precision nutrition models.(Reference Popejoy and Fullerton128)
Randomized controlled trials using omics endpoints
There is a critical need for dietary intervention RCTs that incorporate omics biomarkers as exposure measures and mechanistic outcomes.(Reference Zeevi, Korem and Zmora6,Reference de Toro-Martín, Arsenault and Després11) As a key example, a randomized, open-label trial conducted in China demonstrated that baseline gut microbiome features could predict individualized glycemic benefits from dietary fibre supplementation in adults with prediabetes, supporting microbiome-informed responder stratification for precision nutrition.(Reference Song, Feng and Ma142) Building on this emerging evidence, future RCTs in Asian populations should prioritize culturally relevant and scalable interventions, with a focus on fermentable dietary fibres, legumes, soy-based foods, omega-3–rich fish and seafood, and fermented foods, paired with multi-omics endpoints (e.g. microbiome features, SCFAs, metabolomics, and postprandial glycemic readouts such as CGM) to identify biological responders versus non-responders and to define minimal effective ‘doses’ of dietary modification. Such trials can provide mechanistic evidence necessary for personalized dietary guidelines.(Reference Berry, Valdes and Drew8)
In addition to published efficacy trials, protocol papers and trial registries of ongoing or planned studies can provide timely evidence on emerging study designs, research priorities, and upcoming precision-nutrition interventions in Asian populations. For example, the New Zealand SYNERGY study – a fully diet-controlled, parallel, residential intervention conducted in Asian Chinese and European Caucasian adults with prediabetes – illustrates the value of controlled feeding designs for isolating diet effects while enabling cross-population comparisons without weight-loss confounding.(Reference Sequeira-Bisson, Fraser and Leiu143)
Implementation science and real-world application
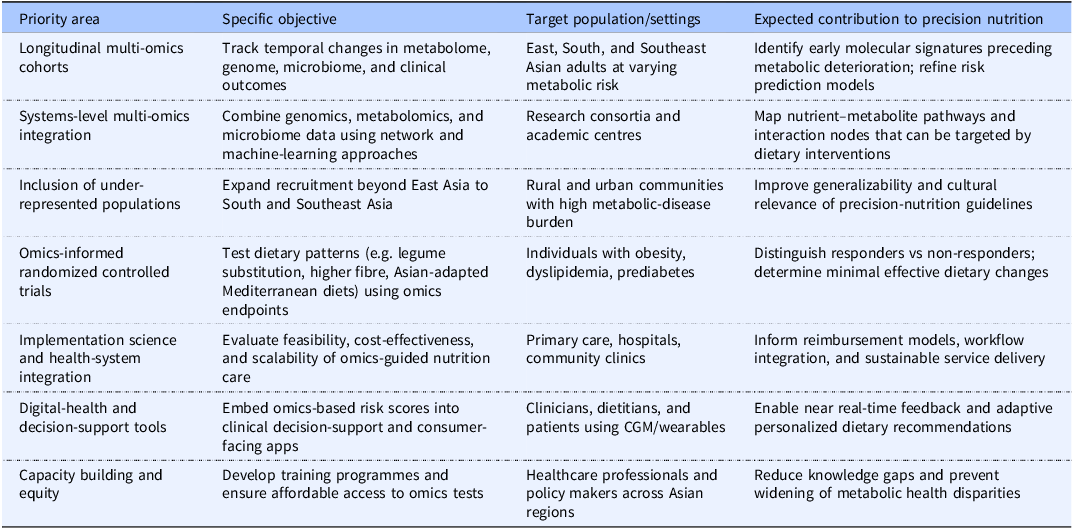
Finally, implementation-science frameworks are needed to evaluate the feasibility, cost-effectiveness, and long-term sustainability of omics-guided dietary interventions in diverse healthcare settings.(Reference Khoury, Gwinn and Yoon131,144,Reference Roberts, Kennedy and Chambers145) These frameworks should address practical factors, including laboratory capacity, digital health integration, clinician training, reimbursement structures, and patient acceptability. The major research and implementation priorities for omics-guided precision nutrition in Asia are summarized in Table 2.
Research and implementation priorities for omics-guided precision nutrition in Asian populations

Table 2. Long description
The table outlines research and implementation priorities for omics-guided precision nutrition in Asian populations. It consists of four columns: Priority area, Specific objective, Target population/settings, and Expected contribution to precision nutrition. The table has six rows, each detailing specific objectives, target populations, and expected contributions. The priority areas include longitudinal multi-omics cohorts, systems-level multi-omics integration, inclusion of under-represented populations, omics-informed randomized controlled trials, implementation science and health-system integration, digital-health and decision-support tools, and capacity building and equity. Each row provides specific objectives, target populations or settings, and the expected contributions to precision nutrition. For example, the first row focuses on tracking temporal changes in metabolome, genome, microbiome, and clinical outcomes among East, South, and Southeast Asian adults at varying metabolic risk, aiming to identify early molecular signatures preceding metabolic deterioration and refine risk prediction models.
Conclusion
Precision nutrition provides a promising approach for improving metabolic health across Asia by integrating genetic, metabolomic, microbiome, and environmental data into personalized dietary recommendations. The Asian populations are characterized by distinct metabolic phenotypes, including heightened visceral adiposity, reduced β-cell reserve, increased oxidative stress, and culturally shaped dietary exposures, which influence susceptibility to metabolic diseases. Multi-omics biomarkers, such as lysoPCs, retinoid metabolites, amino acid signatures, oxidative stress indicators, and emerging markers of postprandial glycemic excursions, microbiome activity, and fatty acid remodelling, offer sensitive tools for the early detection of metabolic dysfunction and may guide tailored dietary interventions.
Genetic–metabolomic integration further enhances risk stratification by accounting for both inherited susceptibility and dynamic metabolic states. Translating these insights into practice will require robust analytical standardization, improved clinician training, digital health solutions capable of supporting adaptive personalized guidance, and a commitment to equity across diverse Asian regions. Future progress will also depend on longitudinal multi-omics cohorts and omics-endpoint dietary RCTs, complemented by protocol papers and trial registries that provide early visibility into ongoing and upcoming interventions. By aligning mechanistic evidence with culturally informed dietary strategies, omics-guided precision nutrition has the potential to contribute meaningfully to early disease prevention and reduce the growing burden of metabolic disorders in Asia.
Acknowledgements
The author gratefully acknowledges the contributions of the research participants, graduate students, and collaborating researchers whose work informed this review. Appreciation is also extended to the Korean Nutrition Society, American Society for Nutrition, and International Union of Nutritional Sciences for their ongoing support of precision nutrition research and capacity building.
Financial support
This research was supported by the National Research Foundation of Korea grant funded by the Korean government (MSIT) (RS-2024-00355298) and by the 2025 Hannam University Research Fund.. The funder had no role in the design, analysis, interpretation, or writing of the manuscript.
Competing interests
The author declares no conflicts of interest.
Ethical standards
This is a narrative review. All studies that involved human participants were conducted in accordance with the ethical standards of the institutional or national research committees and the 1964 Declaration of Helsinki and its later amendments.
Authorship
M.K. conducted the literature search, performed data synthesis, prepared the manuscript, supervised all aspects of the review, secured funding, critically revised the manuscript, and approved the final version.



 Open access
Open access