


Introduction
Caves are abundant throughout the karst landscape of Cretaceous limestone (e.g., Edwards and Trinity groups) that forms the bedrock of the Edwards Plateau in central Texas (Kastning, Reference Kastning, Beck and Wilson1987; Veni, Reference Veni, Elliott and Veni1994). The sheltered settings of caves are well known for their potential to preserve detailed records of paleoclimate and the diversity of past biotic communities (Hadly, Reference Hadly1999; Lundelius, Reference Lundelius2006; Jass and George, Reference Jass and George2010; Schubert and Mead, Reference Schubert, Mead, White, Culver and Pipan2019). On the Edwards Plateau, records of late Quaternary climate, flora, and fauna derived from caves were instrumental in reconstructing regional natural history. Exploration of Pleistocene fossils in regional caves began early in the twentieth century (Hay, Reference Hay1920), but the richness of the record came into focus later, beginning in the late 1950s (Lundelius, Reference Lundelius, Schubert, Mead and Graham2003). Vertebrate fossils from caves on the Edwards Plateau of central Texas contributed to developing understanding of Late Pleistocene community dynamics throughout North America and played a key role in framing the debate over the cause(s) of end-Pleistocene extinctions (Lundelius, Reference Lundelius, Martin and Wright1967, Reference Lundelius and Black1974, Reference Lundelius and White2009; Graham and Lundelius, Reference Lundelius, Genoways and Dawson1984; Toomey, Reference Toomey1993, Reference Toomey, Elliott and Veni1994).
Fossil assemblages from more than 37 caves and rock-shelters were studied by Ernest L. Lundelius, Jr. (Lundelius, Reference Lundelius, Martin and Wright1967, Reference Lundelius, Genoways and Dawson1984, Reference Lundelius, Woodruff, Snyder, De La Garza and Slade1985; Sansom and Lundelius, Reference Sansom and Lundelius2005), his graduate students (Semken, Reference Semken1961; Patton, Reference Patton1963; Graham, Reference Graham1976a; Toomey, Reference Toomey1993; Winkler and Gose, Reference Winkler, Gose, Schubert, Mead and Graham2003; Sagebiel, Reference Sagebiel2010), and colleagues (Meade, Reference Meade1961; Slaughter, Reference Slaughter1966a; Holman, Reference Holman1968; Dalquest et al., Reference Dalquest, Roth and Judd1969; Bell et al., Reference Bell, Sagebiel, Lundelius, Czaplewski, Lively, Minatra, Lenamond, Barcelo and Gelnaw2022). The resulting dataset documents patterns of biogeography and species diversity across the late Quaternary, from the last glacial maximum (LGM) to the Late Holocene (ca. 20,000 to ca. 200 years ago). Extensive sampling of cave deposits produced a robust database of fossil occurrences that permitted regional syntheses. Remains of mammoths (Mammuthus), bison (Bison), horse (Equus), camel (Camelops), and extralimital small mammals were widespread, while other taxa, including tapir (Tapirus) and mastodon (Mammut) were rare.
Those patterns of vertebrate diversity supported pollen and other lines of evidence indicating the presence of a relatively dry, open grassland in a cool climate with reduced seasonality (relative to the present-day; Lundelius, Reference Lundelius, Martin and Wright1967; Hall and Valastro, Reference Hall and Valastro1995; Cordova and Johnson, Reference Cordova and Johnson2019). The absence of megafauna typical of other regions of Texas and North America, such as giant beaver (Castoroides) or pampathere (Holmesina), was interpreted as the result of biogeographic variation and the absence of appropriate environmental and/or climatic conditions (Lundelius, Reference Lundelius, Martin and Wright1967, Reference Lundelius, Schubert, Mead and Graham2003). Those regional data contributed to documenting broader patterns of occurrence across North America (e.g., FAUNMAP Working Group 1994, 1996) and were pivotal to recognizing and understanding the non-analog composition of Late Pleistocene communities (Graham, Reference Graham1976b, Reference Graham, Diamond and Case1985; Lundelius, Reference Lundelius1989). Today that foundational understanding of the fauna of the Edwards Plateau is regularly utilized in meta-analyses of Late Pleistocene community dynamics (e.g., Smith et al., Reference Smith, Tomé, Elliot Smith, Lyons, Newsome and Stafford2016; Seersholm et al., Reference Seersholm, Werndly, Grealy, Johnson, Keenan Early, Lundelius and Winsborough2020).
We initiated new, ongoing investigations of cave deposits and associated fossils from the Late Pleistocene across the Edwards Plateau that revealed a subset of caves with a distinct fossil biota. The record of Late Pleistocene faunas from regional caves was based on previous studies of fossils from sedimentary deposits preserved in 16 rockshelters and dry caves and one site, the Avenue Site, in surface alluvial deposits (Lundelius, Reference Lundelius, Schubert, Mead and Graham2003; Fig. 1; Table 1). Other caves in the region contain active underground rivers or streams and are, in Texas, colloquially referred to as water caves. Regional water caves also contain the remains of Pleistocene vertebrates, but those fossils have never been formally studied and are largely unknown. Paleontological exploration of regional water caves is still in early stages, but our initial work led to the discovery of fossils in Bender’s Cave that represent extinct megafauna that were previously unknown from the Edwards Plateau or are rare in the 17 other regional Late Pleistocene sites. In this paper we provide the first report of the Late Pleistocene fossil assemblage from Bender’s Cave and describe the occurrence of regionally novel megafauna taxa.
Texas and the Edwards Plateau with relevant Late Pleistocene sites indicated. See Table 1 for corresponding details of each site.

Late Pleistocene sites in Texas shown in Figure 1 and discussed in the text.
Study area and taphonomy
The fossils we describe come from Bender’s Cave in Comal County (Figs. 1–3), a cave within the limestone bedrock of the lower Glen Rose Formation (Veni, Reference Veni1997). Although the paleontology of the cave has never been investigated, Bender’s Cave was explored and mapped by cavers from the 1990s to the present as a part of investigations into the hydrogeology of local groundwater systems (Veni, Reference Veni1997).
Bender’s Cave is a groundwater conduit system that contains an active underground stream. Water flowing through Bender’s Cave is a component of the Trinity Aquifer system (Ashworth and Hopkins, Reference Ashworth and Hopkins1995) and the cave is one of many that contribute groundwater into the Guadalupe River basin. Incision of the Guadalupe River system during the Middle Pleistocene exposed the lower Glen Rose Formation, allowing groundwater to discharge and initiating the development of conduit cave passages (Woodruff and Abbott, Reference Woodruff and Abbott1979; Veni, Reference Veni1997).
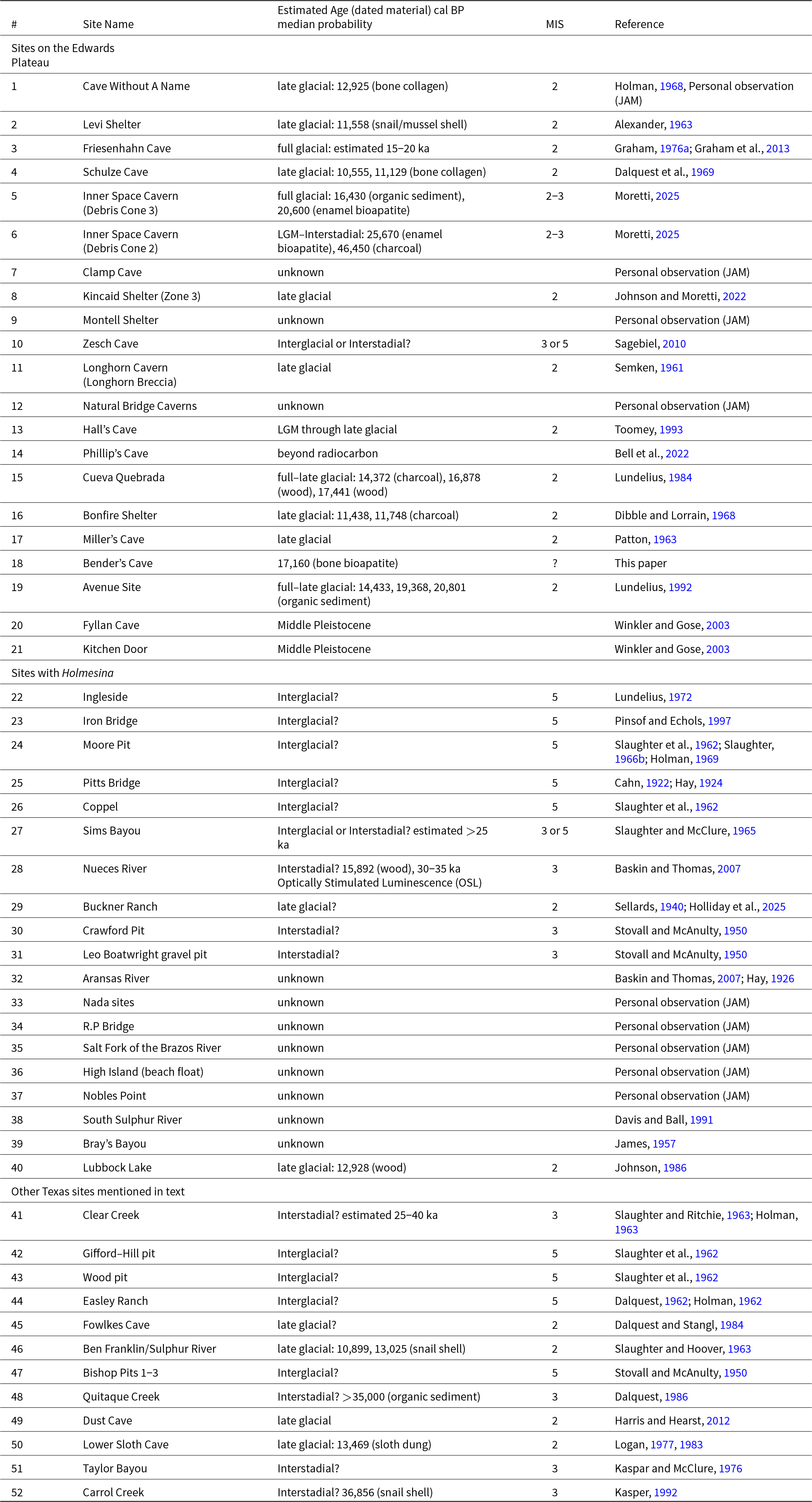
Bender’s Cave, and others like it, have three basic structural components. Vertical sinkholes extend from the upland surface and intersect the conduit, or stream, passages, allowing surface water (and surface materials) to enter the cave system and directly recharge the aquifer. Horizontal stream passages (Figs. 2 and 3) act as drainage conduits, facilitating the movement of groundwater within limestone bedrock. Groundwater and recharge water flow downstream through the conduit stream passages and eventually discharge at natural springs in the Guadalupe River system.
Stream passage in Bender’s Cave. (A) Low ceilings and shallow water; (B) collecting fossils underwater in one of two large rooms; (C) snorkel survey in deeper water; note the mud coating on the ceiling and walls (All units are in millimeters).

We accessed Bender’s Cave from the upland surface of the plateau by rappelling into vertical sinkhole shafts. Two of three sinkhole entrances to Bender’s Cave were filled naturally with debris and were subsequently dug open by cavers. The sinkhole shafts are narrow (≤ 1 m) at the surface but widen into elongate fissures in the subsurface. Sinkhole #2, the primary entrance shaft, extends down 11 m below the surface and leads to a horizontal crawlspace 10–15 m in length that eventually intersects the flowing stream passage. That crawlspace contains soil, leaf litter, and other recent organic remains, including Late Holocene and/or modern skeletal elements of raccoons (Procyon lotor) and other extant vertebrates.
Stream passages range in size and form from ovaloid tunnel-like passages, 2–5 m wide by 0.5–2 m tall, to relatively narrow (∼2.5 m), canyon-like passages with high ceilings (≥ 3.5 m) (Fig. 2). Smaller side passages, some of which are not passable, branch off from the main stream passage (Fig. 3). Bender’s Cave contains two relatively large rooms (≥ 10 m tall) with domed ceilings and talus slopes of sediment (Fig. 2B). Those rooms appear to be potential past connections to the surface, but aside from three known sinkhole entrances (one of which, Sinkhole #1, is plugged with cave fill and garbage) and a spring entrance, there are no known connections to the surface. Water levels in Bender’s Cave fluctuate as a function of moisture abundance on the surface but the cave maintains permanent pools and flowing water, including during drought. We measured water levels ranging 22–99 cm between September 2022 and November 2024 with some areas, including large pools, being far deeper. The distribution and condition of objects in the stream bed as well as material coating the ceilings illustrate that the passages experience intervals with large volumes of turbulently flowing water.
Overview of Bender’s Cave illustrating location of entrances, layout of the stream passage, and placement of sampling zones. Sinkhole #3 and adjacent passage have been explored but not mapped and are not shown here. Cave map drafted by M. Miller and reproduced with permission (All units are in millimeters).
Skeletal remains of extinct, Pleistocene megafauna (e.g., Mammuthus, Camelops, Equus) are common in Bender’s Cave, with the spatial distribution and physical modification of the fossils indicating prolonged exposure to calcite-rich groundwater and fluvial transport as part of a lag assemblage. Skeletal elements of Pleistocene taxa are commonly polished, rounded, and battered. Those fossils are typically stained with iron-rich minerals and manganese and are dark in color, heavy in weight, and exhibit coatings of calcite and/or cemented matrix. The remains of smaller vertebrates (e.g., turtles, rabbits, rodents, small carnivorans) also exhibit these characteristics and are also Pleistocene in age. Fossils occur primarily as isolated elements scattered loose across the stream bed and in gravel bars. Pleistocene fossils also occur in speleothem formations, within isolated cobble-sized clasts of breccia, and in intact layers of bone breccia.
Fossils are not uniformly distributed and occur in sections of the cave where there are banks of sedimentary deposits as well as in areas with exposed bedrock floors. Sedimentary deposits, similar to cut-banks and point-bars, contain in-situ wood fragments and have some stratification. We have not yet observed vertebrate fossils in-situ in those deposits and the relationship between the sediment banks and fossil bone is unclear. Some fossils of extinct megafauna were recovered incidentally from a buried, submerged deposit of blue-green gleyed clay. Those fossils also exhibit polishing, rounding, and coatings of manganese.
Methods and materials
We collected faunal remains (i.e., Pleistocene fossils and twentieth century bones/teeth) as part of initial fieldwork designed to search for, document, and investigate Pleistocene materials in Bender’s Cave. Faunal remains were collected opportunistically from the surface of gravel bars, sediment banks, and the stream bed floor during six sampling episodes from March 2023 to November 2024. Faunal remains were collected underwater using snorkels and dive masks. A limited sample was made of fossils found protruding from underwater mud in a ∼30 × 30 cm area. Those fossils had coatings of, and cavities filled with, a blue-green gleyed clay, suggesting the existence of a buried, intact pond deposit. We divided the stream passage into 21 arbitrary sampling zones, each ∼70 m in length (Fig. 3), and recorded the horizontal location of all specimens collected to document the distribution and relative abundance of faunal remains. All recovered materials were donated by the landowners to the Jackson School Museum of Earth History Vertebrate Paleontology Laboratory at The University of Texas at Austin.
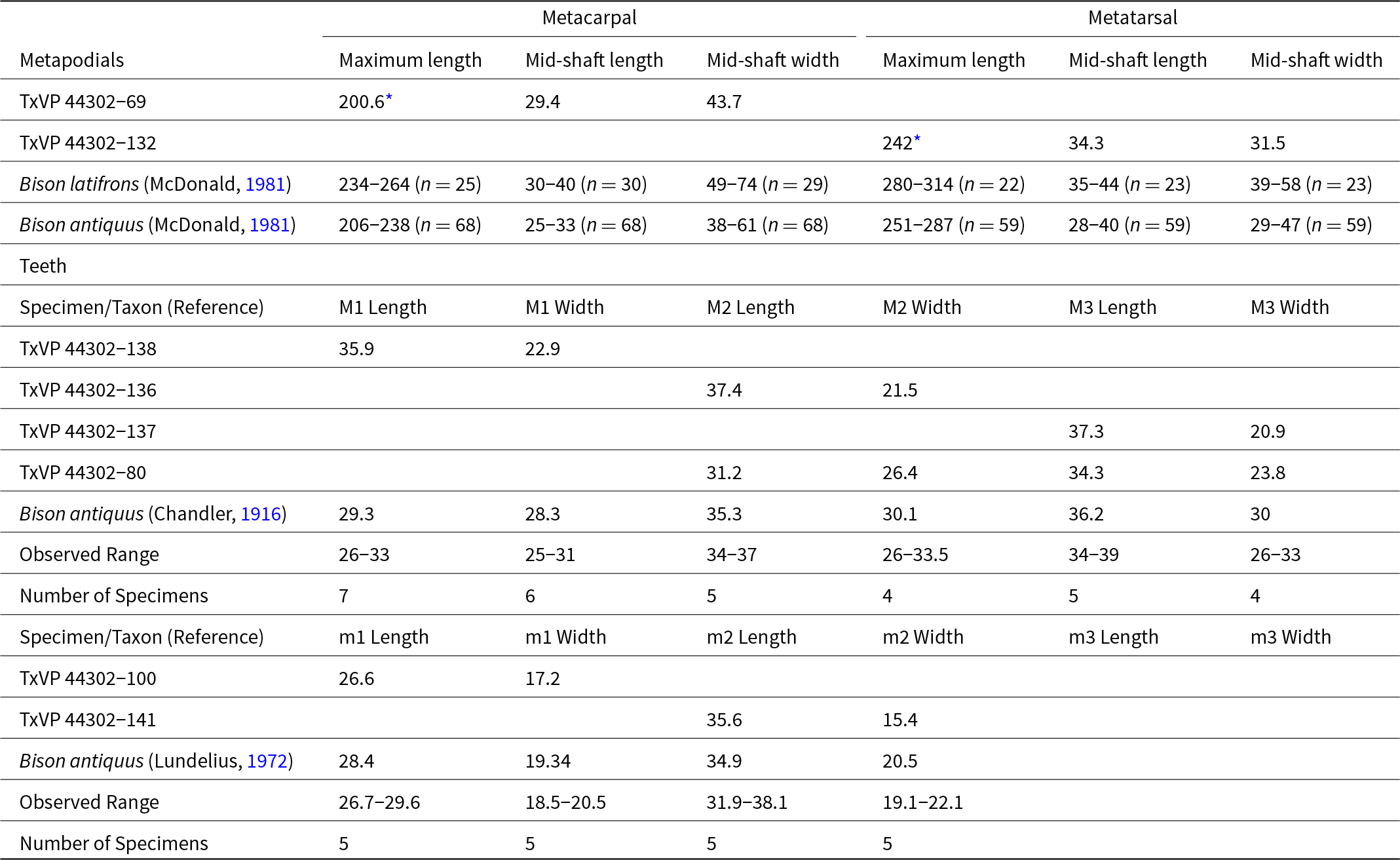
Taxonomic assignment of the recovered faunal remains was based on the presence of diagnostic or distinctive morphological traits. Distinctive traits are relative differences in size and form that allow us to recognize a given taxon but lack an evolutionary context. Diagnostic traits reflect unique morphological states, some (but not all) of which are apomorphies. Teeth are referred to by an upper-case (teeth of the premaxilla and maxilla) or lower-case (teeth of the dentary) letter in combination with a number indicating tooth position. For example, I3, c1, P3, and m2 referred to upper incisor 3, lower canine 1, upper premolar 3, and lower molar 2, respectively. Specimen measurements in tables are reported in millimeters.
Two fragments of Late Pleistocene bone from Bender’s Cave were sampled for collagen by the Center for Applied Isotope Studies at the University of Georgia for radiocarbon analysis. Those samples came from a fragment of tibia (Cetartiodactyla [indeterminate]; TxVP 44302-36) collected loose on the floor of the stream bed and a transverse process from a thoracic vertebra (TxVP 44302-33) of a large mammal recovered from a buried, submerged deposit of blue-green clay. Those specimens were chosen because they came from the two depositional contexts sampled in the cave and represented megafaunal mammals. We did not select more taxonomically significant specimens because those specimens are relatively few and we could not justify destructively sampling those specimens given the high probability of significant post-mortem, geochemical alteration of the fossils. Both samples lacked collagen.
TxVP 44302-36 was subsequently sampled for bioapatite. That specimen was cleaned, washed in an ultrasonic bath, and crushed into fragments. The bone was then treated with diluted 1N acetic acid to remove secondary carbonates. The sample was reacted under vacuum with 100% phosphoric acid to dissolve the bone mineral and release CO2, which was then cryogenically purified and catalytically converted to graphite. The resulting radiocarbon age was calibrated using CALIB Rev. 8.2 (Stuiver and Reimer, Reference Stuiver and Reimer1993) using the IntCal20 calibration curve (Reimer et al., Reference Reimer, Austin, Bard, Bayliss, Blackwell, Bronk Ramsey and Butzin2020) at the 95% confidence interval.
The similarity between the composition of samples of fossil megafauna from the water caves and other Late Pleistocene sites in Texas was explored using hierarchical cluster analysis in R 4.41 (R Core Team, 2024). We observed similarity and dissimilarity between lists of taxa identified from Bender’s Cave and those identified from other Late Pleistocene sites in Texas. A hierarchical cluster analysis is one method of evaluating those similarities in a more critical and quantifiable manner than simply comparing faunal lists. Hierarchical cluster analysis is an exploratory tool, used here to identify general patterns in taxon occurrence and relationships between sites across geographic space and geologic time. Visualization and quantification of those relationships offers a basis for developing hypotheses regarding large scale patterns in species diversity across space and time.
A variation of the Raup–Crick index (Raup and Crick, Reference Raup and Crick1979) was used for the cluster analysis because that method emphasizes the occurrence of rare taxa over the occurrence of common taxa when estimating similarity. That emphasis is desirable here because we expect little change in megafaunal diversity in one region of North America throughout the duration of the Late Pleistocene and, accordingly, many taxa (e.g., Equus, Camelops, Bison) may be common to most, or all, analyzed samples. Rare taxa, in contrast, appear to have a greater potential to highlight compositional differences that reflect individualistic species responses to latitudinal and/or temporal variation in paleoenvironments and associated community diversity. Raup–Crick’s index is also preferred here because it has a greater capacity, relative to Simpson’s and other indices, to accurately group similar samples despite uneven sampling, unsampled taxa, variation in species richness, and other issues related to incomplete data that are inherent to paleontological samples (Travouillon et al., Reference Travouillon, Archer, Legendre and Hand2007). For those reasons, the Raup–Crick index was used previously to discern biochronologic patterns in fossil samples (e.g., Smith and Polly, Reference Smith and Polly2013) and is increasingly utilized in modern ecological studies to explore patterns of β-diversity (e.g., Chase et al., Reference Chase, Kraft, Smith, Vellend and Inouye2011).
We used the ‘raup’ function in the package ‘vegan’ to perform a cluster analysis of a presence/absence matrix of 31 taxa of Late Pleistocene megafauna (Supplementary Table 1) using the ‘ward.D2’ method. Microfauna genera were excluded because they are, so far, poorly sampled from Bender’s Cave. The presence/absence matrix was generated based on published records as well as taxon occurrences observed in samples housed at the Texas Vertebrate Paleontology Collections from 43 Late Pleistocene sites in Texas (sites 1–11, 13–19, 22–32, 37–38, and 40–52 in Table 1). Published radiocarbon ages (n = 24) from those sites were obtained previously from a variety of materials, including bone collagen (n = 3), bone bioapatite (n = 3), charcoal (n = 4), wood (n = 4), sloth dung (n = 1), organic sediment (n = 5), and snail shells (n = 4), using a variety of methods and field sampling procedures and, as a result, are not equally reliable and may not be directly comparable (Table 1). We accepted radiocarbon ages as published. We calibrated those radiocarbon ages using CALIB Rev. 8.2 with the IntCal20 calibration curve and they are listed in Table 1 as median probabilities.
We assessed the reliability of the results of the cluster analysis by calculating a coefficient of cophenetic correlation. We employed a multiscale bootstrap resampling of the data to determine clusters. AU (i.e., approximately unbiased) p-values were calculated using multiscale bootstrap resampling. Those values indicated the probability of recovering the topology of sites within a cluster. The arrangement of sites in clusters with low (e.g., 0%) AU values indicated that those arrangements were poorly resolved and not reliable. High (e.g., 95–100%) AU values indicated that the arrangement of sites in a cluster was relatively well resolved and stable. AU values do not, however, indicate a measure of probability of the existence of individual clusters.
Results
Systematic paleontology
Class Reptilia Laurenti, Reference Laurenti1768
Order Testudines Batsch, Reference Batsch1788
Family Testudinidae Batsch, Reference Batsch1788
Testudinidae (indeterminate)
Material and provenience
TxVP 44302-18, shell (carapace or plastron) fragment; TxVP 44302-145, shell (carapace or plastron) fragment; TxVP 44302-146, carapace fragment; TxVP 44302-147, shell (carapace or plastron) fragment. Zone 13–14 (TxVP 44302-147), Zone 15 (surface: TxVP 44302-18; blue-green clay: TxVP 44302-145, TxVP 44302-146).
Description and discussion
Four large, thick (e.g., 23 mm) fragments of giant tortoise (Testudinidae) shell collected from the blue-green clay (n = 2) and the stream bed surface (n = 2), represent some form of giant tortoise. Abrasion and other taphonomic modifications have removed most of the cortical surfaces, obscuring the morphology of the shell fragments and preventing assignment to genus or species.
Genus Hesperotestudo Williams, Reference Williams1950
Hesperotestudo sp. (giant form)
Material and provenience
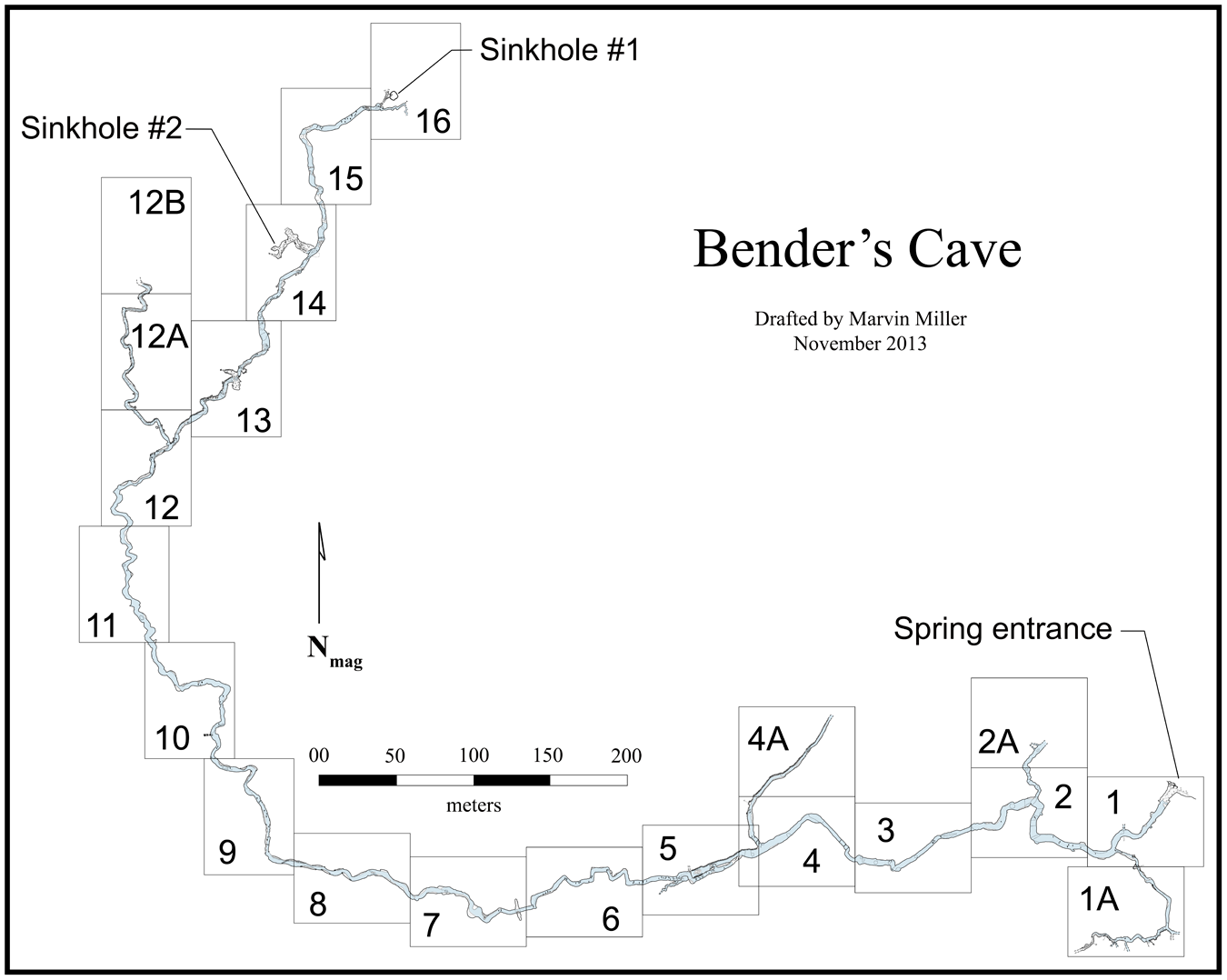
TxVP 44302-76, scapula, left (Fig. 4). Zone 15 (blue-green clay).
Left scapula of Hesperotestudo sp. (giant form) (TxVP 44302-76). (A) Posterior view; (B) anterior view. Scale bar is 5 cm.
Description and discussion
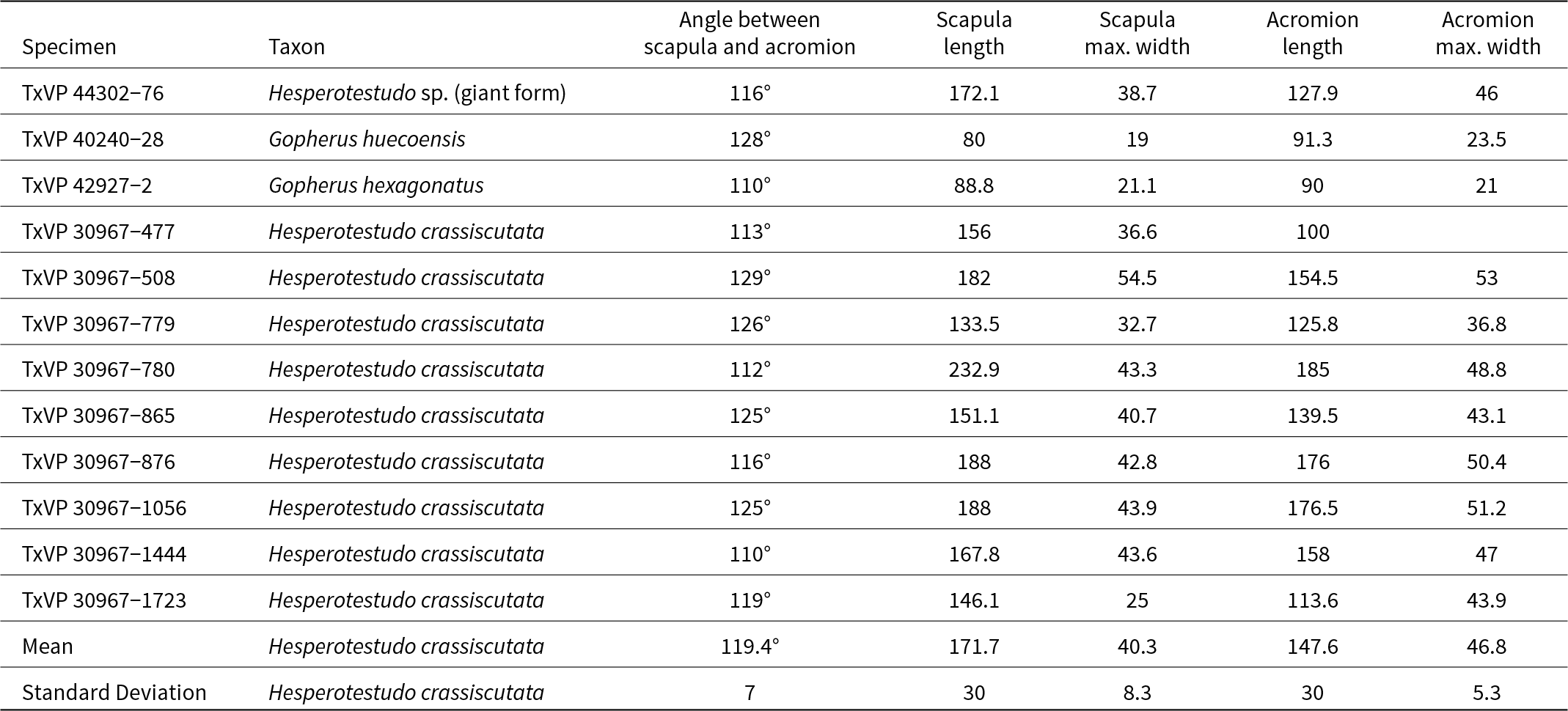
A scapula of a large turtle with the ends of the scapula and acromion processes, the cortical surfaces of the glenoid fossa, and articulation for the procoracoid all broken off and missing (Fig. 4). The angle between the acromion and scapula processes is obtuse, not acute as in Emydidae (Auffenberg, Reference Auffenberg1974). Within Testudinidae, Hesperotestudo is reportedly distinct from Gopherus in having a greater angle between the scapula and acromion processes and in having a scapula that is round in cross-section (Meylan and Sterrer, Reference Meylan and Sterrer2000). We encountered substantial variation in the angle of the processes in a sample of extinct tortoises from the Pliocene and Pleistocene of Texas. Two scapulae from extinct Gopherus (G. huecoensis Strain, Reference Strain1966, and G. hexagonatus Westgate, Reference Westgate, Hall, Collins and Prewitt1987) exhibited angles of 110° and 128°, respectively (Table 2). The angles in a large sample of Hesperotestudo crassiscutata from Ingleside, Texas (Lundelius, Reference Lundelius1972), largely overlapped with the Gopherus specimens, ranging 110–129°, with a mean of 119.4° (n = 9; Table 2). Although those species of Gopherus are giant (e.g., carapace length ≥ 50 cm; scapula length: 80 and 88.8 mm; Table 2), the Hesperotestudo specimens were consistently far larger (scapula length observed range: 133.5–232.9 mm; Table 2). In Hesperotestudo, the scapula is compressed antero-posteriorly where it diverges from the acromion process but widens dorsally to a consistently round cross-section while the scapula in Gopherus remains compressed throughout and is ovaloid or crescentic in cross-section. TxVP 44302-76 has an angle of 116°, the scapula process has a round cross-section, and the size of the specimen fits within the range documented by the sample of Hesperotestudo crassiscutata from Ingleside (scapula length: 172.1 mm; Table 2).
Measurements of scapulae in some giant species of Gopherus and Hesperotestudo All measurements are in millimeters.
Class Mammalia Linnaeus, Reference Linnaeus1758
Superorder Xenarthra Cope, Reference Cope1889
Order Pilosa Flower, Reference Flower1883
Family Megalonychidae Ameghino, Reference Ameghino1889
Genus Megalonyx Jefferson, Reference Jefferson1799
Megalonyx jeffersonii Desmarest, Reference Desmarest1822
Material and provenience
TxVP 44302-41, lower caniniform, left; TxVP 44302-42, ungual; TxVP 44302-43, ungual; TxVP 44302-44, phalanx, proximal and medial, co-ossified, digit III, pes, right (Fig. 5A–C). Zone 15 (blue-green clay: TxVP 44302-41, TxVP 44302-42, TxVP 44302-44; surface: TxVP 44302-43).
Fossils of Megalonyx jeffersonii and Holmesina septentrionalis. (A, B) Left lower caniniform of Megalonyx jeffersonii, TxVP 44302-41 (A) in lingual view and (B) in occlusal view; (C) ungual of Megalonyx jeffersonii, TxVP 44302-43; (D, E) buckler osteoderm of Holmesina septentrionalis, TxVP 44302-30 (D) in dorsal view and (E) in ventral view. Scale bar is 2 All units are in millimeters.

Description and discussion
TxVP 44302-41 is a high-crowned tooth, ovaloid in cross-section with a rootless base and an open pulp cavity (Fig. 5A, B). The tooth is consistent with descriptions by Leidy (Reference Leidy1853) and Stock (Reference Stock1925) of the caniniform teeth of Megalonychidae. The occlusal surface is worn into a depression in the center with transversely oriented striations and scars (Fig. 5B). The labial face of the tooth exhibits longitudinal striations. The lingual face exhibits a lingual bulge bounded anteriorly and posteriorly by distinct longitudinal grooves (Fig. 5A, B). The presence of the lingual bulge and its demarcation by parallel grooves are diagnostic of Megalonyx and M. jeffersonii, respectively (McDonald, Reference McDonald1977; McDonald et al., Reference McDonald, Harington and De Iuliis2000).
The two unguals match the descriptions of Megalonychidae and Megalonyx jeffersonii by Leidy (Reference Leidy1853), McDonald (Reference McDonald1977), and Hoganson and McDonald (Reference Hoganson and McDonald2007). Most of the nail sheath is broken and missing from both specimens. TxVP 44302-43 is nearly complete, missing only the distal and proximal-most extremities of the bone (Fig. 5C). TxVP 44302-42 is missing the distal end of the ungual process and nearly 1/3 of the left side (when viewed distally) from the dorsal to the ventral border. Both specimens exhibit vascular grooves and associated foramina extending dorsally and distally from the ventral base of the ungual process. The ungual process is large and slightly asymmetric in cross-section in both specimens, indicating that neither specimen is from digit 1 or digit 3 of the manus or pes. The left face of TxVP 44302-42 is slightly convex and the right face is relatively straight, while the reverse morphology is present in TxVP 44302-43. The straight face of the ungual process would have faced digit 3. Those details suggest that TxVP 44302-42 represents the ungual of left digit 2 or right digit 4 in the manus or pes and that TxVP 44302-43 represents the ungual of right digit 2 or left digit 4 in the manus or pes.
TxVP 44302-44 is a large, robust phalanx with a broad proximal end bearing a deep central groove for articulation with metatarsal III and a relatively narrow distal end with two large, round condyles separated by a wide, flattened groove. Co-ossification of phalanges 1 and 2 in digit III is a common feature of the pes in multiple clades of extinct ground sloths. The co-ossified phalanx in digit III of Megatheriidae is distinct in being proximo-distally compressed and having a wedge-like shape (Owen, Reference Owen1860; De Iuliis, Reference De Iuliis1996). Comparison with illustrations (Stock, Reference Stock1917) and isolated, non-fused proximal and medial phalanges of Paramylodon (TxVP 30967, Ingleside, Texas) indicate that, in this taxon, the proximal central groove is narrower relative to TxVP 44302-44, the articulation for the medial sesamoid is narrower, and the distal trochleae are tilted medially relative to the proximal-distal long axis of the bone. TxVP 44302-44 is larger than the corresponding element in Nothrotheriops and is further distinguished from that genus by a central groove on the proximal end that is wider dorsally than ventrally (Stock, Reference Stock1925). The size and morphology of TxVP 44302-44 matches seminal descriptions (Leidy, Reference Leidy1853; Stock, Reference Stock1925; McDonald, Reference McDonald1977) of co-ossified proximal and medial phalanges of pedal digit III in Megalonyx. Within Megalonychidae, the co-ossification of the proximal and medial phalanges in pedal digit III is known to occur only in Megalonyx jeffersonii and is absent in earlier genera and species (Hirschfeld and Webb, Reference Hirschfeld and Webb1968; McDonald, Reference McDonald1977; McDonald et al., Reference McDonald, Miller and Morris2001).
Order Cingulata Illiger, Reference Illiger1811
Family Pampatheriidae de Paula Couto, Reference de Paula Couto1954
Genus Holmesina Simpson, Reference Simpson1930
Holmesina septentrionalis Leidy, Reference Leidy1889a
Description and discussion
TxVP 44302-30 is polygonal (five-sided; Fig. 5D and E). The ventral surface is concave and exhibits, at least, six large neurovascular foramina and many smaller foramina, pits, and fissures. The dorsal surface of the osteoderm is weakly convex and exhibits a marginal band (terminology of Edmund, Reference Edmund1985) that ranges from 1–5.2 mm wide along the borders of the osteoderm. The marginal band surrounds an elevated rim termed the submarginal band, sharply defined and extending up to 1.4–1.8 mm above the surface of the marginal band. The submarginal band surrounds a central figure, a rounded elongate ridge oriented antero-posteriorly, which is bordered medially and laterally by a shallow concavity, or sulcus. The posterior end of the central figure abuts the submarginal band. The dorsal surface is marked by dozens of small foramina, pits, and fissures. Piliferous foramina, occur along the edges of the submarginal band and are concentrated throughout the marginal band.
Pampathere taxa are diagnosed primarily on the morphology of isolated osteoderms, and that morphology is variable within and between individual animals (Edmund, Reference Edmund, Stewart and Seymour1996). The size and form of buckler osteoderms varies ontogenetically in an extant analog, the nine-banded armadillo (Dasypus novemcinctus; personal observation, JAM), and similar ontogenetic variation presumably occurred in extinct pampatheres. This variation suggests isolated, individual osteoderms may not accurately reflect the morphology of all osteoderms in a carapace (De Iuliis and Edmund, Reference De Iuliis and Edmund2002) and the emphasis in pampatheriid taxonomy on osteoderm morphology may bias our understanding of the diversity of pampathere taxa (de Porta, Reference de Porta1962). Still, the morphology preserved in the osteoderm from Bender’s Cave is sufficient to assign this specimen to species based on current practices in pampatheriid taxonomy.
The osteoderm from Bender’s Cave is assigned to Holmesina based on the presence of a relatively wide, continuous, and depressed marginal band, an elevated submarginal band, and the presence of a linear, ridge-like central figure that meets the submarginal band (Simpson, Reference Simpson1930; Holmes and Simpson, Reference Holmes and Simpson1931; Edmund, Reference Edmund1985, Reference Edmund, Stewart and Seymour1996; Scillato-Yané et al., Reference Scillato-Yané, Carlini, Tonni and Noriega2005, Góis, Reference Góis2013). The central figure is distinct and elevated relative to the rest of the dorsal surface (in contrast to Pampatherium and Vassalia; Scillato-Yané et al., Reference Scillato-Yané, Carlini, Tonni and Noriega2005) and lacks lateral projections (as in Tonnicinctus; Góis et al., Reference Góis, González-Ruiz, Scillato-Yané and Soibelzon2015). The adjacent sulci are shallow (in contrast to Kraglievichia; Jiménez-Lara, Reference Jiménez-Lara2020). The piliferous foramina are round and set within broadly rounded grooves and pits, but the anterior foramina are not connected by a channel (as in Scirrotherium; Edmund and Theodor, Reference Edmund, Theodor, Kay, Madden, Cifelli and Flynn1997). The form of these foramina and their concentration throughout the marginal band combined with the traits listed above support assignment to Holmesina septentrionalis (Leidy, Reference Leidy1889b; Sellards, Reference Sellards1915; Simpson, Reference Simpson1930). The morphology of TxVP 44302-30 fits within the range of qualitative variation present in a large sample of buckler osteoderms (n = 52) of Holmesina septentrionalis from Ingleside, Texas (Lundelius, Reference Lundelius1972).
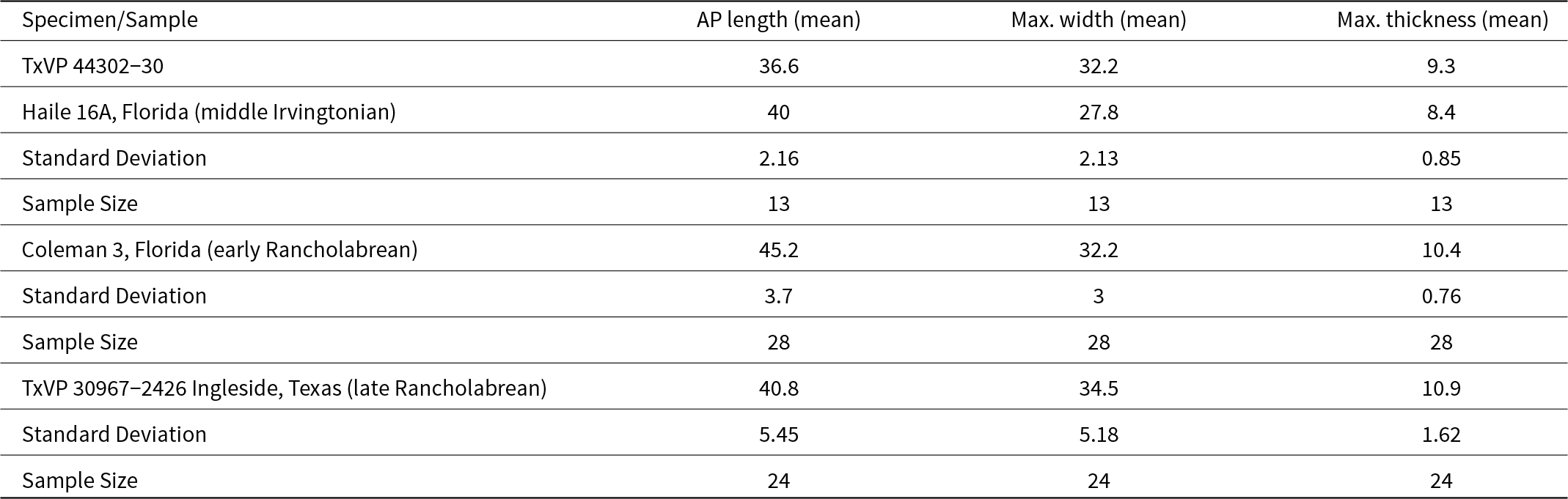
The size of TxVP 44302-30 (Table 3) also supports assignment to Holmesina septentrionalis. Large samples of Holmesina from a time series of Plio-Pleistocene sites in North America document an increase in body size from Holmesina floridanus in Blancan and early Irvingtonian faunas to Holmesina septentrionalis in the middle Irvingtonian to Rancholabrean faunas (Edmund, Reference Edmund and Montgomery1987; Hulbert and Morgan, Reference Hulbert, Morgan, Martin and Barnosky1993). Although that size change is apparent in large samples (n = 200+) of skeletal elements (Hulbert and Morgan, Reference Hulbert, Morgan, Martin and Barnosky1993), size change through time is difficult to distinguish from ontogenetic size change in small samples. Nonetheless, the largest, geologically youngest specimens do not overlap in size with the smallest, geologically oldest specimens. TxVP 44302-30 falls (Table 3) within a chronocline of buckler osteoderm size between middle Irvingtonian (Haile 16A) and early Rancholabrean (Coleman 3) samples of Holmesina septentrionalis from Florida (Hulbert and Morgan, Reference Hulbert, Morgan, Martin and Barnosky1993). That osteoderm size provides a quantitative basis for assigning TxVP 44302-30 to Holmesina septentrionalis.
Size of the buckler osteoderm of Holmesina septentrionalis from Bender’s Cave in comparison to other samples. Data from Florida (Hulbert and Morgan, Reference Hulbert, Morgan, Martin and Barnosky1993) and Texas samples is for 5-6 sided buckler osteoderms only All measurements are in millimeters.
Order Carnivora Bowdich, Reference Bowdich1821
Family Felidae Fischer, Reference Fischer1817
Subfamily Machairodontinae Gill, Reference Gill1872
Genus Homotherium Fabrini, Reference Fabrini1890
Homotherium serum Cope, Reference Cope1893
Material and provenience
TxVP 44302-31, P4, right; TxVP 44302-39, parietal-occipital; TxVP 44302-53, atlas (Fig. 6). Zones 11 (TxVP 44302-31) and 12 (TxVP 44302-39). TxVP 44302-53 was collected prior to the start of research and lacks provenience data.
Fossils of Homotherium serum. (A, B) Right P4, TxVP 44302-31 (A) in labial view and (B) in lingual view; (C, D) parietal-occipital, TxVP 44302-39 (C) in left lateral view and (D) in posterior view; E–G) atlas, TxVP 44302-53 (E) in anterior view, (F) in dorsal view, and (G) in ventral view. Scale bar is 2 cm. Abbreviations: lc = lambdoid crest, mc = metacone, pc = paracone, prez = prezygapophysis, sc = sagittal crest, vc = vertebarterial canal, vmt = ventral median tubercle.

Description and discussion
The P4 (TxVP 44302-31) is elongate and blade-like with a steeply angled shearing surface on the lingual face of the crown (Fig. 6A and B). The posterior root is long (25.4 mm) relative to the anterior root (11.1 mm). There is a wear facet on the lingual face of the mesial end of the tooth, in the location of the parastyle (Fig. 6B). The paracone is relatively short and the metacone is long (Fig. 6A, B). There is no protocone or an associated antero-internal root (i.e., protoradix; Fig. 6B).
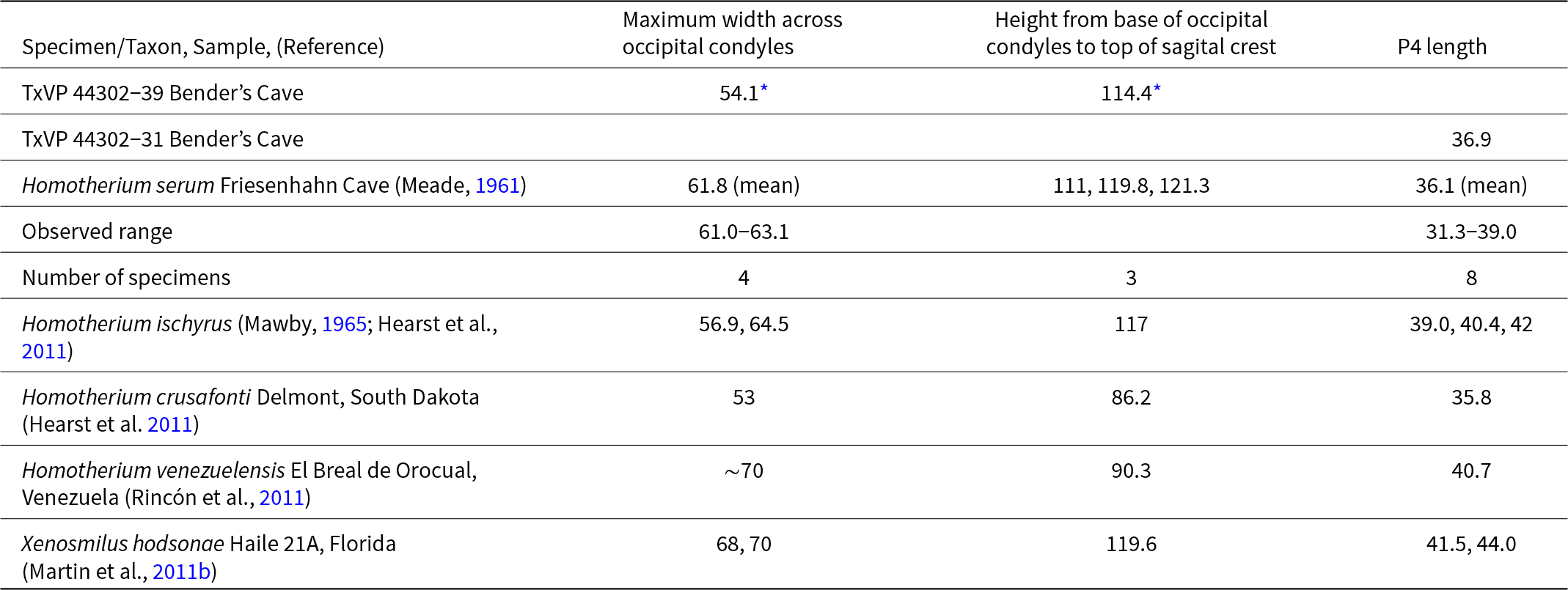
The reduction of the protocone on P4 is a synapomorphy of Homotherini (Martin et al., Reference Martin, Naples, Barbiarz, Naples, Martin and Barbiarz2011a; Christiansen, Reference Christiansen2013). A reduced protocone and/or protoradix is present on the P4 of Homotherium crusafonti, H. ischyrus, H. latidens, and Xenosmilus hodsonae (Ballesio, Reference Ballesio1963; Martin and Harksen, Reference Martin and Harksen1975; Hearst et al., Reference Hearst, Martin, Babiarz, Naples, Naples, Martin and Babiarz2011; Martin et al., Reference Martin, Naples, Barbiarz, Naples, Martin and Barbiarz2011b). The P4 of Homotherium serum lacks a protocone and protoradix (Cope Reference Cope1893), but the protocone is also absent in H. venezuelensis (Rincón et al., Reference Rincón, Prevosti and Parra2011). TxVP 44302-31 is smaller than H. venezuelensis but consistent in size and morphology with H. serum from Friesenhahn Cave (Table 4).
Measurements of the P4 and occipital of Homotherium serum All measurements are in millimeters.
* specimen damaged and measurement incomplete
TxVP 44302-39 is a fragmentary skull with portions of the parietal and occipital preserved (Fig. 6C and D). A large, prominent (albeit damaged) sagittal crest extends across the antero-posterior length of the preserved parietal (Fig. 6D). A large lambdoid crest projects posteriorly but is broken and abbreviated. Viewed posteriorly, the skull is tall dorso-ventrally, narrow medio-laterally, and sub-triangular in outline (Fig. 6C). A pronounced median ridge extends dorsally from the foramen magnum, with deep concavities on either side of the ridge. The occipital is broken and fragmentary but the occipital condyles, foramen magnum, and immediately adjacent areas are relatively well preserved (Fig. 6C). The ventral surface of the occipital is largely broken away and missing but the hypoglossal foramen and the posterior border of the left jugular foramen are present.
The preserved portions of TxVP 44302-39 are sufficient to support assignment to Homotherium serum. The tall, narrow, sub-triangular occipital is distinctive of Machairodontinae (Emerson and Radinsky, Reference Emerson and Radinsky1980; Christiansen, Reference Christiansen2008). Within Machairodontinae, the extreme dorso-ventral elongation of the occipital distinguishes the specimen from Smilodon and is typical of Homotherium (Meade, Reference Meade1961; Churcher, Reference Churcher1966). Occipital condyles are narrow, as in Homotherium crusafonti and H. serum (Table 4). The fragmentary skull from Bender’s Cave is taller (measured from the occipital condyles to the sagittal crest) than in H. crusafonti but matches the size and form of H. serum from Friesenhahn Cave (Table 4).
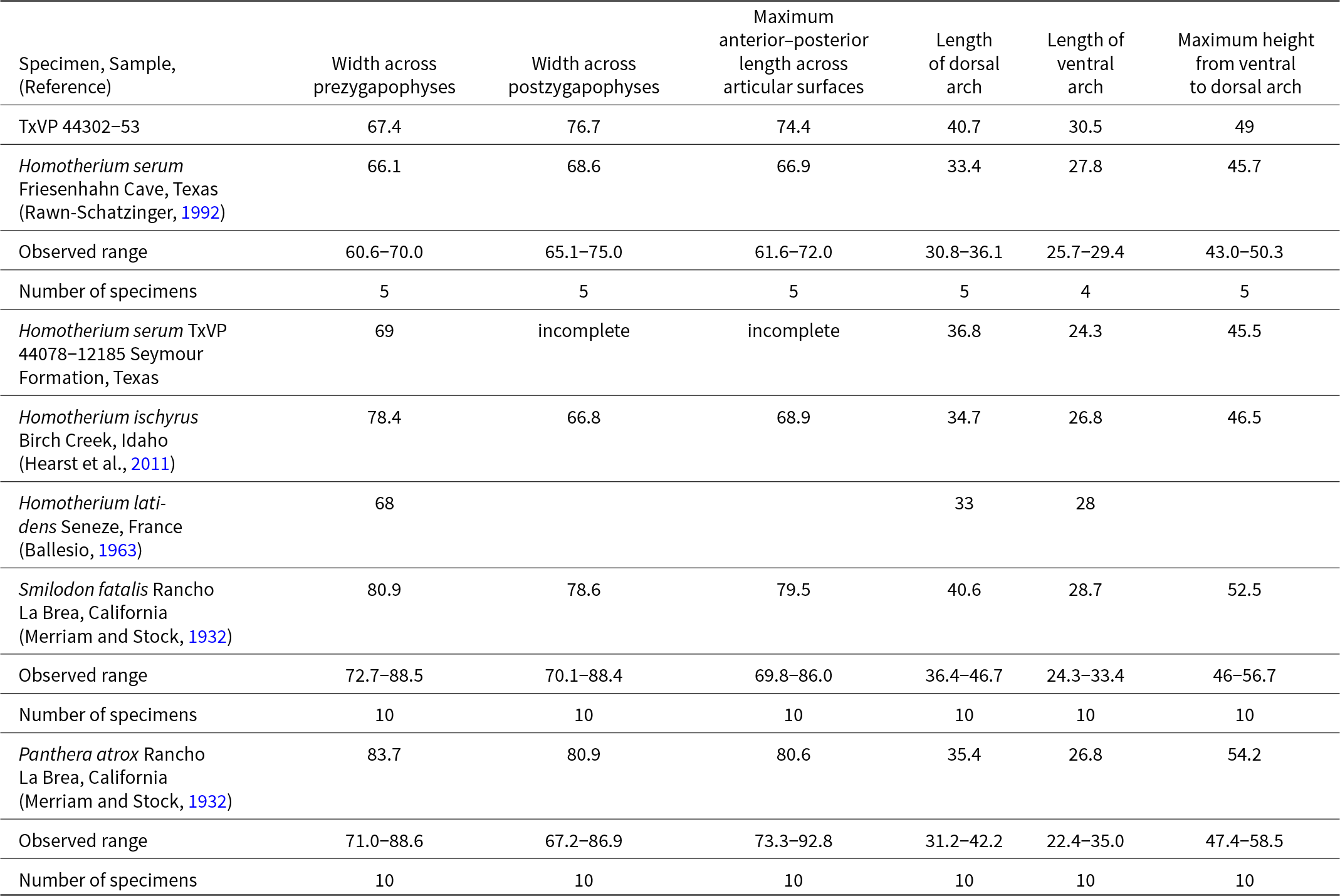
TxVP 44302-53 is assigned to Homotherium based on a prominent and ridge-like median tubercle on the ventral arch, posterior openings to the vertebarterial canal that are ventrally located, and the medial expansion of the ventral articular surfaces of the prezygapophyses (Fig. 6E–G). Comparisons with published descriptions (e.g., Merriam and Stock, Reference Merriam and Stock1932; Ballesio, Reference Ballesio1963) supplemented with modern (Panthera leo: TxVP M-874, M-916; P. onca: TxVP M-6646; P. tigris: TxVP M-6574, M-8778, M-15762) and fossil (Smilodon fatalis from Rancho La Brea: TxVP 31021-8304, TxVP 31021-8305; Homotherium serum from Friesenhahn Cave: TxVP 933-2276, TxVP 933-3382, TxVP 933-3007, TxVP 933-2280, TxVP 933-544) specimens illustrated consistent differences between the atlas of Panthera, Smilodon, and Homotherium and suggest that the traits outlined above are, in combination, diagnostic for Homotherium. The atlas of Xenosmilus is not known (Martin et al., Reference Martin, Naples, Barbiarz, Naples, Martin and Barbiarz2011b).
Homotherium has a median tubercle that extends along the posterior half of the ventral arch and projects posteriorly beyond the posterior border of the arch. That tubercle is a prominent ridge in Homotherium, but is smaller in Panthera, present only as a triangular prong on the posterior border of the ventral arch. The tubercle also is simple and small in Smilodon (Merriam and Stock, Reference Merriam and Stock1932); there is no ventral ridge, instead this attachment for the longus colli muscle occurs as a blunt, posteriorly facing tubercle. The ventral median tubercle in Homotherium ischyrus was reported to be larger than in Homotherium serum from Friesenhahn Cave (Hearst et al., Reference Hearst, Martin, Babiarz, Naples, Naples, Martin and Babiarz2011). The ventral tubercle in TxVP 44302-53 (Fig. 6G) is indistinguishable from the same tubercle in specimens from Friesenhahn Cave.
In all three genera of large felids examined here, the vertebarterial canal terminates posteriorly immediately lateral to the postzygopophysis. The position of the posterior opening to the vertebarterial canal differs among the three genera, with the opening positioned near the center (dorso-ventrally) of the postzygapophyses in Panthera and Smilodon fatalis, but near the ventral edge of the postzygapophyses in Homotherium serum. The prezygapophyses extend farther medially in Homotherium serum than in Panthera or Smilodon, with the two articular surfaces nearly meeting along the midline.
TxVP 44302-53 exhibits the morphology of Homotherium (Fig. 6E–G), but some measurements, especially the length of the dorsal arch of the atlas, extend outside of the range of variation observed in Homotherium (Table 5). We interpret the larger size of the specimen as representing an aspect of qualitative variation that has not previously been sampled in this rare felid. That hypothesis is supported by relatively large samples of atlas vertebrae from Rancho La Brea (Merriam and Stock, Reference Merriam and Stock1932) that document a wide range of size variation (Table 5) in both the extinct lion Panthera atrox and the machairodontine Smilodon fatalis.
Measurements of the atlas of Homotherium, Smilodon, and Panthera atrox All measurements are in millimeters.
Order Perissodactyla Owen, Reference Owen1848
Family Equidae Gray, Reference Gray1821
Genus Equus Linnaeus, Reference Linnaeus1758
Equus sp.
Material and provenience
TxVP 44302-21, metatarsal III, right; TxVP 44302-105, P3 or P4, right; TxVP 44302-113, DP3 or DP4, right; TxVP 44302-114, M3, left; TxVP 44302-115, P3 or P4, left; TxVP 44302-116, P3 or P4, left; TxVP 44302-117, M3, right; TxVP 44302-118, M3, right; TxVP 44302-119, M1 or M2, right; TxVP 44302-120, P3 or P4, right; TxVP 44302-121, DP2, left; TxVP 44302-122, M1 or M2, right; TxVP 44302-123, upper cheek tooth fragment, right; TxVP 44302-124, upper cheek tooth fragment, unworn, right; TxVP 44302-125, P3 or P4, in fragment of maxilla, right; TxVP 44302-126, dp3, right; TxVP 44302-127, m2, left; TxVP 44302-128, lower cheek tooth fragment; TxVP 44302-130, metacarpal III, right (Fig. 7).
Fossils of Equus sp. (A, B) right P3 or P4, TxVP 44302-105 (A) in labial view and (B) in occlusal view; (C, D) right M1 or M2, TxVP 44302-119 (C) in labial view and (D) in occlusal view; (E, F) left DP2, TxVP 44302-121 (E) in labial view and (F) in occlusal view; (G, H) right M1 or M2, TxVP 44302-122 (G) in labial view and (H) in occlusal view; (I–J) right dp3, TxVP 44302-126 (I) in occlusal view, (J) in labial view; (K, L) left m2, TxVP 44302-127 (K) in occlusal view, (L) in labial view. Scale bar is 2 All units are in millimeters. Abbreviations: ecf = ectoflexid, hyg = hypoconal groove, lif = linguaflexid, mtc = metaconid, mts = metastylid, plih = pli hypostyle, plip = pli protoloph, prc = protocone, prs = protoselene.

Zone 7–8 (TxVP 44302-128), Zone 9 (TxVP 44302-113), Zone 11–12 (TxVP 44302-123), Zone 13 (TxVP 44302-122), Zone 13–14 (TxVP 44302-121, TxVP 44302-130), Zone 14 (TxVP 44302-105), Zone 15 (blue clay: TxVP 44302-114 through TxVP 44302-120, TxVP 44302-124 through TxVP 44302-126; surface: TxVP 44302-127, TxVP 44302-130).
Diagnosis and description
Assignment of fossil specimens to Equus sp. follows MacFadden (Reference MacFadden, Janis, Scott and Jacobs1998) and Barrón-Ortiz et al. (Reference Barrón-Ortiz, Avilla, Jass, Bravo-Cuevas, Machado and Mothé2019) in diagnosing this genus based on large size in combination with upper cheek teeth that have the protocone attached to the protoselene, the consistent presence of a single pli hypostyle and a pli protoloph, lower cheek teeth with the metaconid and metastylid well separated from each other on p3–m3, a shallow ectoflexid that does not penetrate the isthmus between the metaconid and metastylid on p2–p4, and monodactyl, mesaxonic foot morphology.
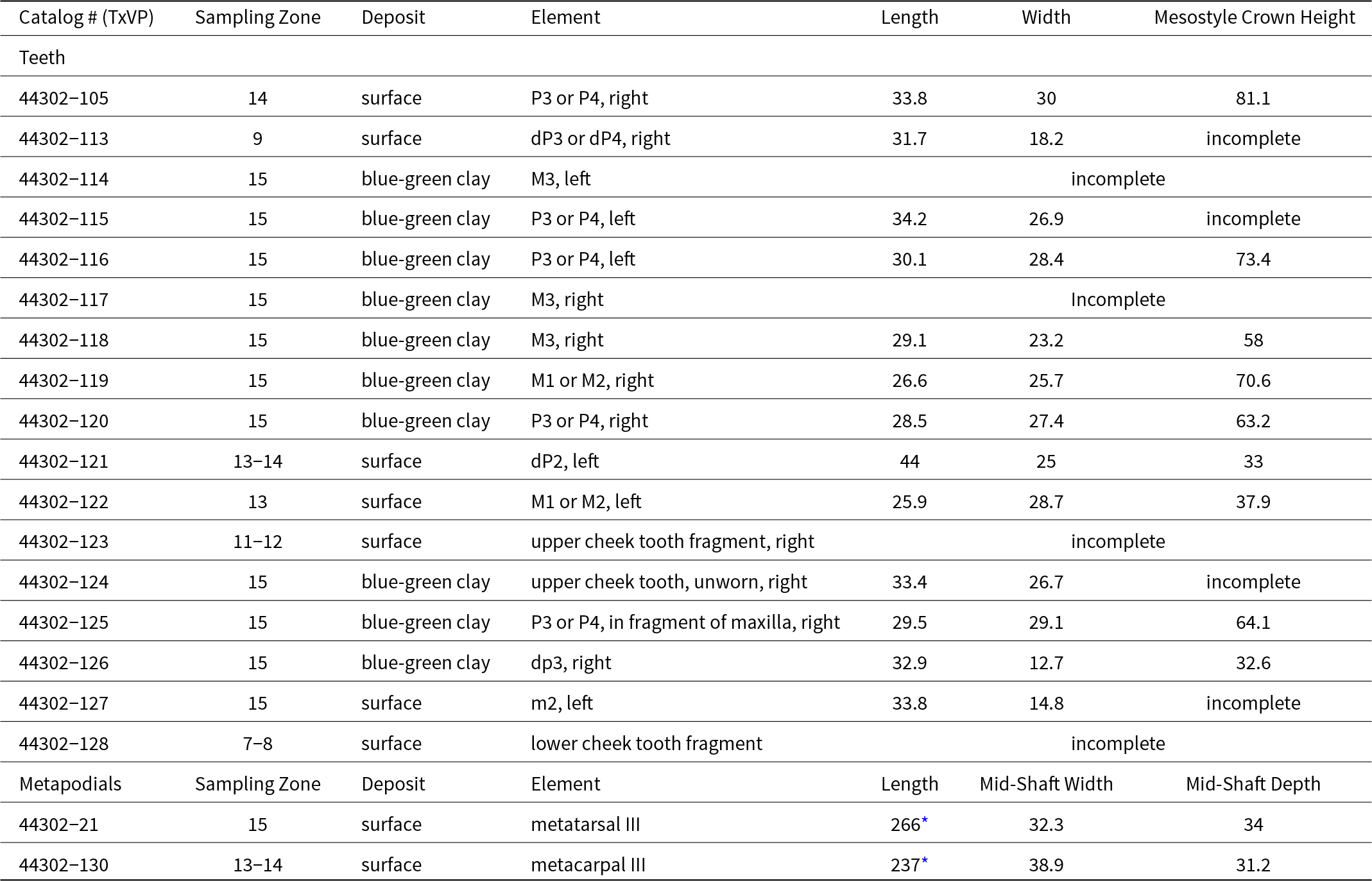
The complete, worn, adult upper cheek teeth from Bender’s Cave are relatively large (Table 6) and have plicated fossette borders with a pli hypostyle and a pli protoloph (Fig. 7A–D, G-H). Most (6/7 specimens) upper cheek teeth have simple, single pli caballin (Fig. 7B and D), except in TxVP 44302-122, where the pli caballin is absent (Fig. 7H). The hypoconal groove is shallow but varies from “U”- to “V”-shaped (Fig. 7B, D, H), except in an M3 (TxVP 44302-118) where it occurs as an isolated enamel lake immediately distal to the postfossette. The protocone is large, oblong, and is attached to the protoselene (Fig. 7B, D, H). The lingual border of the protocone is grooved in most specimens (5/7; Fig. 7B) but is straight in TxVP 44302-119 and TxVP 44302-122 (Fig. 7D and H). The only complete, adult lower cheek tooth is an m2 (TxVP 44302-127), which has a shallow ectoflexid that does not penetrate the isthmus, a pli caballinid, and crenulated enamel borders on the labial faces of the entoflexid, metaflexid, and metastylid (Fig. 7K and L). The metaconid and metastylid are separated by a wide, rounded, and generally “U”-shaped linguaflexid (Fig. 7K and L). A dp3 (TxVP 44302-126) exhibits an ectoflexid that penetrates deeply into the isthmus, a “V”-shaped linguaflexid, a pli caballinid, crenulated enamel on labial border of the metaflexid only, and lacks a protostylid and plicated enamel on the isthmus (Fig. 7I). A DP2 (TxVP 44302-121) is complete but is so lightly worn that the details of the enamel pattern are unclear (Fig. 7E and F).
Size of teeth and metapodials of Equus sp. from Bender’s Cave All measurements are in millimeters.
* specimen damaged and measurement incomplete
Two metapodials (TxVP 44302-21, TxVP 44302-130) are modified by taphonomic processes (weathering, polishing, abrasion, calcite growth) and incomplete, but represent a large (Table 6), monodactyl equid.
Discussion
Upper molar TxVP 44302-122 is distinct in being relatively small (Table 6), in lacking a pli caballin, and having a straight lingual border of the protocone (Fig. 7G and H). The remainder of the upper cheek teeth are morphologically similar, however three specimens (TxVP 44302-105, TxVP 44302-115, TxVP 44302-124) stand out as being relatively long in the mesial-distal direction (Table 6). The variation observed in the dentition may indicate the presence of multiple species in the Bender’s Cave sample, but more material is needed to confidently discern such a pattern.
Order Cetartiodactyla Montgelard, Catzeflis, and Douzery, 1997
Cetartiodactyla (indeterminate)
Material and provenience
TxVP 44302-36, tibia diaphysis, left (Fig. 8A). Zone 13.
Limb elements of artiodactyl taxa. (A) Left tibia diaphysis of Cetartiodactyla (indeterminate), TxVP 44302-36, in posterior view; (B) left metacarpal III–IV of Camelops hesternus, TxVP 44302-28, in anterior view; (C) left metacarpal III–IV of Bison sp., TxVP 44302-69, in anterior view; (D, E) right metatarsal III–IV of Bison sp., TxVP 44302-132 (D) in proximal view and (E) in anterior view. Scale bar is 5 cm. Abbreviations: cvg = central vascular groove, df = distal foramen, fg = foramen and groove.

Description and discussion
The tibia is weathered, damaged, and incomplete, missing the proximal and distal ends (Fig. 8A). The posterior face of the diaphysis is flattened antero-posteriorly and has a foramen that opens proximally into a prominent groove just medial to the lateral border (Fig. 8A). There is a ridge along the length of the medial border of the posterior face. The diaphysis is 346 mm long and, at the location of the foramen, is 64.5 mm wide. The size of the specimen, combined with the morphology of the foramen, suggests Camelops or Bison (Webb, Reference Webb1965). Both genera have a foramen located near the lateral border of the diaphysis. A similar foramen is present in Equus, but is situated more centrally on the diaphysis, farther from the lateral border, relative to Camelops or Bison. The specimen is too incomplete to support a more refined taxonomic assignment. A 2.7-g fragment of cortical bone from TxVP 44302-36 was destructively sampled for a radiocarbon age.
Family Camelidae Gray, Reference Gray1821
Genus Camelops Leidy, Reference Leidy1854
Camelops hesternus Leidy, Reference Leidy1873
Material and provenience
TxVP 44302-28, metacarpal III–IV, left (Fig. 8B). Zone 14.
Description and discussion
TxVP 44302-28 is a large, fused metacarpal III–IV with the divergent distal articular condyles typical of Camelidae (Fig. 8B). The proximal end exhibits articular facets for the magnum and trapezoid. The facet for the magnum is larger than the facet for the trapezoid and it projects proximally beyond the lateral facet for articulation with the cuneiform. The facets for the trapezoid and the cuneiform are separated by a triangular fossa that is perforated by several foramina. The anterior face of the metacarpal is convex and lacks the longitudinal groove that is typical of extant Camelus (Webb Reference Webb1965; Fig. 8B). The posterior face of the diaphysis bears a pair of foramina within a deep central groove that is bounded medially and laterally by rugose ridges. The morphology of TxVP 44302-28 matches descriptions of the metacarpal of Camelops hesternus (Webb, Reference Webb1965; Zazula et al., Reference Zazula, MacPhee, Hall and Hewitson2016). The length of TxVP 44302-28 (381 mm) is consistent with the range observed for Camelops hesternus from Rancho La Brea (374–380 mm; Webb, Reference Webb1965). This specimen is more robust than the elongate metacarpals of llama-like camelids (e.g., Lama, Hemiauchenia, Palaeolama; Webb, Reference Webb1965; Breyer, Reference Breyer1974; Harrison, Reference Harrison1985) and is smaller than the giant forms Gigantocamelus, Megacamelus, and Megatylopus (Breyer, Reference Breyer1974; Zazula et al., Reference Zazula, MacPhee, Hall and Hewitson2016).
Family Cervidae Goldfuss, Reference Goldfuss1820
Odocoileini Pocock, Reference Pocock1923
Material and provenience
TxVP 44302-143, dentary fragment with m1 or m2 (Fig. 9A and B). Zones 8–9 (TxVP 44302-143).
Teeth of deer (Odocoileini) and Bison sp. (A, B) Left dentary fragment with m1 or m2 of Odocoileini, TxVP 44302-143 (A) in lateral view and (B) in occlusal view; (C) left maxilla fragment with M2–M3 of Bison sp., TxVP 44302-80, in occlusal view. Scale bar is 2 cm. Abbreviations: ect = ectostylid, ent = entostyle, mts = metastylid.

Description and discussion
TxVP 44302-143 is a fragment of a dentary with m1 or m2 (Fig. 9A, B). The molar is small (length 14.9 mm, width 9.9 mm), brachyodont, and selenodont. A small ectostylid is present and there is a narrow anterior cingulum (Fig. 9A). The parastylid, metaconid, and metastylid are prominent; the metastylid curves sharply posteriorly and lingually (Fig. 9B). The entoconid and entostylid are broken off and missing. Dentine in the anterior crest is confluent with dentine in the posterior crest (Fig. 9B). The molar is consistent with m1/2 of extant Odocoileus virginianus, however the morphology of lower molars in late Neogene–Quaternary cervids is conservative (Gustafson, Reference Gustafson1985; Dong, Reference Dong, Ohtaishi and Sheng1993) and size varies with geography and geologic time (Purdue, Reference Purdue1989; Lister, Reference Lister1996). In the Western Hemisphere, lower molars of species of Odocoileus and Bretzia overlap broadly in size and form (Wheatley and Ruez, Reference Wheatley and Ruez2006; Gustafson, Reference Gustafson2015). That variation prevents a genus-level assignment in this case and the specimen from Bender’s Cave is assigned generally to the Western Hemisphere tribe Odocoileini (Heckeberg, Reference Heckeberg2020).
Family Bovidae Gray, Reference Gray1821
Genus Bison Smith, Reference Smith, Griffith, Smith and Pidgeon1827
Bison sp.
Material and provenience
TxVP 44302-69, metacarpal III–IV, left; TxVP 44302-80, maxilla fragment with M2–M3, left; TxVP 44302-100, m1, left; TxVP 44302-132, metatarsal III–IV, right; TxVP 44302-136, M2, left; TxVP 44302-137, M3, left; TxVP 44302-138, M1, right; TxVP 44302-141, m2, left (Fig. 8C-E, 9C). Zone 10 (TxVP 44302-138), Zones 13–14 (TxVP 44302-132, TxVP 44302-137), Zone 15 (blue-green clay: TxVP 44302-80, TxVP 44302-136, TxVP 44302-141; surface: TxVP 44302-69, TxVP 44302-100).
Description and discussion
Teeth assigned to Bison sp. are large (Table 7), hypsodont and selenodont (Fig. 9C). In the upper molars, the paracone and metacone are situated between a prominent parastyle, mesostyle, and metastyle with the protocone and metaconule occurring as curved cusps on the lingual face mesial and distal, respectively, to a large central, pillar-like entostyle (Fig. 9C). Lower molars have the metaconid and entoconid positioned between a parastylid and entostylid with a prominent ectostylid occurring between the protoconid and hypoconid.
Size of teeth and metapodials of Bison sp. from Bender’s Cave All measurements are in millimeters.
* Specimen incomplete/broken
The proximal end of a left metacarpal (TxVP 44302-69) is nearly crescent-shaped in proximal view and preserves the antero-medial corner of the facet for the fused magnum-trapezoid. A pair of distal articular condyles separated by a notch are present but incomplete. The diaphysis is compressed antero-posteriorly, the anterior face is rounded with a central vascular groove (Fig. 8C), and the posterior face is flattened. Both faces of the diaphysis have large foramina at the distal and proximal ends of the diaphysis, set in the central vascular groove (Fig. 8C).
A right metatarsal (TxVP 44302-132) is missing the distal end and the posterior portions of the proximal articular surfaces (Fig. 8D and E). Articular facets for the fused ectocuneiform–middle cuneiform and the naviculo-cuboid are large, broad, and separated by a central pit with two large foramina. Large foramina are present at the distal end of the diaphysis on the anterior and posterior faces, the foramen on the anterior face is set within a central vascular groove (Fig. 8D).
The morphology of these elements is consistent with extinct species of Bison, as exemplified by large samples from Lubbock Lake (Johnson, Reference Johnson1987) and Blackwater Draw (Stock and Bode, Reference Stock and Bode1936). The specimens from Bender’s Cave are distinct from all other bovids native to the Western Hemisphere in having large, elongate metapodials (metapodials of “ovibovine” caprines are relatively short and wide; see White et al., Reference White, Mead and Morgan2025) with relatively deep central vascular grooves with associated large distal foramina and large, hypsodont cheek teeth with large, prominent entostyles and ectostylids (Neas, Reference Neas1991). Skeletal form does not, however, provide a consistent or reliable means of distinguishing between Bison and domesticated forms of Bos. Assignment of the material from Bender’s Cave to Bison sp. relies on the size of the specimens and their concordance with extinct species of Bison (Table 7).
We follow McDonald (Reference McDonald1981) and Scott (Reference Scott2010) in recognizing two Pleistocene species of Bison in North America, B. latifrons and B. antiquus. The taxonomy of Bison is based almost exclusively on skull and horn-core characters (Skinner and Kaisen, Reference Skinner and Kaisen1947), but the two species exhibit some differences in size. Teeth from Bender’s Cave align with Bison antiquus (Table 7), but measurements of molar rows illustrate that the teeth of B. latifrons and B. antiquus overlapped broadly in size (Miller, Reference Miller1971; McDonald, Reference McDonald1981). Metapodial data appears to be more clear-cut, with measurements of the diaphysis indicating that specimens from Bender’s Cave fall at the lower end of size variation for Bison antiquus and outside of the observed range for Bison latifrons (Table 7). Measurements suggest Bison antiquus is represented by the fossils identified here, but we refrain from identifying the species because the sample lacks a skull or horn-cores.
The giant Bison latifrons is typical of faunas from the last interglacial (“Sangamonian,” Marine Isotope Stage 5/5e; Pinsof, Reference Pinsof, Stewart and Seymour1996) and is often regarded as absent from assemblages younger than ca. 20 ka (McDonald, Reference McDonald1981). Bison antiquus does, however, occur in sites interpreted as MIS 5 in age (e.g., Lundelius, Reference Lundelius1972; Scott, Reference Scott2010), there is some evidence for post-LGM occurrences of Bison latifrons (Baskin et al., Reference Baskin, Walls, Thomas, Morgan, Baskin, Czaplewski, Lucas, Mead and White2022), and the two species may have been contemporaneous from MIS 5 into MIS 3 (Lundelius et al., Reference Lundelius, Downs, Lindsay, Semken, Zakrzewski, Churcher, Harington, Schultz, Webb and Woodburne1987; Scott, Reference Scott2010).
Order Proboscidea Illiger, Reference Illiger1811
Mammutidae Hay, Reference Hay1922
Material and provenience
TxVP 44302-34, molar fragment (Fig. 10A). Zone 15 (blue-green clay).
Fossils of Proboscidea. (A) Molar fragment of Mammutidae, TxVP 44302-34, in anterior view; (B) deciduous premolar of Mammuthus sp., TxVP 44302-70, in anterior or posterior view; (C, D) deciduous premolar of Mammuthus sp., TxVP 44302-71 (C) in occlusal view and (D) in anterior or posterior view. Scale bar is 2 cm. Abbreviation: c = crescentoid.

Description and discussion
The fragment represents the anterior half of a pretrite cusp, complete from the median sulcus to near the labial/lingual border (Fig. 10A). The cusp is broad and triangular in cross-section. An enamel crest (crescentoid of Tobien, Reference Tobien1975) extends from the apex to the base of the cusp. The crest ends near a series of conelets (Fig. 10A). The conelets are largest at the base of the crest. Those large conelets are relatively discrete but smaller conelets are only weakly separated or merged together (Fig. 10A). The orientation and arrangement of the enamel features indicate that the fragment is the first pretrite cusp and that the crest is crescentoid 1 where it connects with the anterior cingulum (terminology of Tobien, Reference Tobien1975). The presence of a linear, crest-like (as opposed to conical) crescentoid, the broad cusp and its triangular cross-section, and the overall morphology of this anterior cusp distinguish the specimen from Gomphotheriidae and indicate a member of Mammutidae. The specimen is, however, too fragmentary for a genus-level assignment. The specimen was recovered from an apparently intact deposit of blue-green clay but exhibits polishing and calcite formations that cement the tooth to a fragment of turtle shell (Fig. 10A).
Family Elephantidae Gray, Reference Gray1821
Genus Mammuthus Brookes, Reference Brookes1828
Mammuthus sp.
Material and provenience
TxVP 44302-70, isolated enamel plate of deciduous premolar (Fig. 10B); TxVP 43407-71, fragment of deciduous premolar (Fig. 10C and D). Zone 9 (TxVP 44302-70) and Zone 12 (TxVP 44302-71).
Description and discussion
Both teeth are incomplete fragments of deciduous premolar crowns (Fig. 10B–D). TxVP 44302-70 is a single unworn enamel plate (Fig. 10B). TxVP 44302-71 is a fragment with two complete plates and a portion of a third (Fig. 10C and D). Assignment to Elephantidae and Mammuthus is based on the presence of thin (1.0–1.1 mm), transverse enamel plates filled with cement (Maglio, Reference Maglio1973). The width of individual cheek teeth varies according to their position in the tooth row, offering a means of identifying isolated teeth and estimating the ontogenetic age of the corresponding individual (Laws, Reference Laws1966; Graham, Reference Graham, Frison and Todd1986a). The fragmentary teeth from Bender’s Cave are complete and intact in their transverse dimensions (Fig. 10B–D). Comparison with tooth widths in a large sample of cheek teeth of Mammuthus columbi from Friesenhahn Cave (Graham, Reference Graham1976a) and other Late Pleistocene sites (Graham, Reference Graham, Frison and Todd1986a) gives context to the fragmentary teeth from Bender’s Cave. The width of TxVP 44302-70 (29.8 mm) is consistent with DP3 (31.8–44.8 mm; Graham Reference Graham1976a) and dp3 (32.0–44.0 mm; Graham, Reference Graham1976a). The width of TxVP 44302-71 (53.5 mm) aligns with DP4 and dp4 (51.2–59.5 mm; Graham, Reference Graham1976a). Wear patterns observed in extant African elephants (Loxodonta, Laws, Reference Laws1966; Haynes, Reference Haynes1991), provide an indication of the general ontogenetic age of the mammoth teeth from Bender’s Cave with TxVP 44302-70 corresponding to individuals 6–18 months in age and the more heavily worn TxVP 44302-71 suggesting an individual 3–13 years in age.
Spatial distribution of faunal remains
The fossils from Bender’s Cave were recovered from the surface of the stream bed (underwater or on gravel bars) and from a submerged, buried deposit of blue-green gleyed clay. The fossils from the stream bed surface occur as a lag assemblage and exhibit a common physical appearance and mineralization. Fossils from blue-green clay are physically similar to fossils obtained loose from the surface of the stream bed (e.g., Fig. 10), but typically exhibit less mineralization and less evidence of fluvial transport. Fossils in the blue-green clay appear to also represent a lag assemblage, preserved in a pond deposit. All described fossils were collected from Zones 15–7 (Table 8; Fig. 3). The blue-green clay occurs near the middle of sampling Zone 15, between Sinkholes #1 and #2 (Fig. 3).
The spatial distribution of fossils specimens based on collecting zone (Figure 3) and depositional setting.
The spatial distribution of the described fossils (Table 8) illustrates that most fossils (57.8%) were concentrated in the blue-green clay with the remaining fossils (42.2%) relatively evenly distributed across the surface of the stream bed, with a slightly greater abundance in the upstream zones (i.e., toward Sinkhole #1; Fig. 3). The tortoise Hesperotestudo sp. (giant form) and the mastodon Mammutidae (indeterminate) were recovered only from the blue-green clay, while Holmesina septentrionalis, Mammuthus sp., Camelops hesternus, Odocoileini, and Homotherium serum were recovered only from the stream bed surface (Table 8). Remains of Testudinidae (indeterminate), Megalonyx jeffersonii, Equus sp., and Bison sp. occurred in both the blue-green clay and the stream bed surface. Fossils of Megalonyx have been found only in Zone 15. Greater sampling effort was invested in collecting fossils from the surface of the stream bed than from the blue-green clay.
All known Pleistocene fossils occur downstream (i.e., toward the spring entrance) of Sinkhole #1 (Table 8, Fig. 3). That sinkhole was used as a trash dump during the twentieth century (personal communication with cavers and landowners, observation of production stamps on associated glass objects). On the surface, Sinkhole #1 is filled with sediment and historic garbage. Within the cave, dense quantities of historic garbage (i.e., glass, metal, and skeletal remains) extend from interior crawlspaces within Sinkhole #1 through the upper reaches of the stream passage (i.e., Zones 15–16) and occur as isolated scatter in Zones 14–6 (Fig. 3). The contents of that garbage pit included skeletal remains. Those twentieth century skeletal remains are found throughout Zones 16–6 and are taphonomically distinct from Pleistocene fossils, recognizable based on their light color, light weight, unstable condition, and the general absence of calcite formations (Fig. 11). Some specimens have distinct straight cut edges, presumably from saw or knife blades (Fig. 11). No extinct or extralimital taxa have been observed among this material. Instead, the assemblage includes common members of the extant native fauna (e.g., Dasypus, Procyon, Odocoileus) and domesticated species (e.g., Sus, Bos, Caprinae/Capra, Equus; Fig.. 11).
Faunal remains associated with the twentieth century trash pit in Sinkhole #1. (A) Anthropogenic cut-bone, TxVP 44302-87; (B) anthropogenic cut-bone, TxVP 44302-88; (C) right maxilla with DP2–DP4 and unerupted adult premolars of Sus scrofa, TxVP 44302-86, in occlusal view; (D) right tibia of Suidae?, TxVP 44302-144, in anterior view, note exfoliating cortical surfaces. Scale bars are 2 cm.

Age of the fossil assemblage
The presence of Bison alongside other extinct, Pleistocene megafauna suggests assignment to the Rancholabrean Land Mammal Age (Bell et al., Reference Bell, Lundelius, Barnosky, Graham, Lindsay, Ruez, Semken, Webb, Zakrzewski and Woodburne2004; Moretti and Bell, Reference Moretti, Bell and Elias2025) and a Late Pleistocene age (129,000–11,700 cal BP; Head et al., Reference Head, Pillans and Zalasiewicz2021) for the fossil assemblage. Lag assemblages are often time averaged, and the fossils described here may not be the same geologic age, a possibility highlighted by the co-occurrence of faunal remains from the Pleistocene and twentieth century. Accordingly, it is possible that not all of the identified fossils represent members of a contemporaneous Rancholabrean fauna. In attempt to clarify the geologic age of the fossils, samples of megafauna bone from the blue-green clay and the stream bed surface were submitted for radiocarbon analysis (see Methods section). Both specimens lacked collagen and an age of 17,160 cal BP (median probability; 17,330–17,030 cal BP age range, 95.4% probability; uncalibrated age: 14,090 ± 40 14C yr BP) was obtained from the bioapatite of a fragment of tibia (TxVP 44302-36) from the stream bed surface assemblage.
The water caves are carbonate-rich environments and those settings present the strong possibility that the fossil bone sampled was contaminated with exogenous carbonates. Ages on bioapatite are often erroneously young as the result of the presence of secondary carbonates (Tamers and Pearson, Reference Tamers and Pearson1965; Stafford et al., Reference Stafford, Hare, Currie, Jull and Donahue1990; Hedges et al., Reference Hedges, Lee-Thorp and Tuross1995; Zazzo and Saliège, Reference Zazzo and Saliège2011; Hopkins et al., Reference Hopkins, Snoeck and Higham2016). A high percent modern carbon value of 17.31% (± 0.08) suggests a significant level of carbonate contamination (Hedges et al., Reference Hedges, Lee-Thorp and Tuross1995; Surovell, Reference Surovell2000) in the fossil sample and that the age of 17,160 cal BP is too young. However, radiocarbon ages on bioapatite can also be falsely old (Irving and Harington, Reference Irving and Harington1973; Nelson et al., Reference Nelson, Morlan, Vogel, Southon and Harington1986), especially in carbonate environments (Cherkinsky, Reference Cherkinsky2009). Errors in either direction, too young or too old, can be on the scale of 20 ka (Nelson et al., Reference Nelson, Morlan, Vogel, Southon and Harington1986; Hopkins et al., Reference Hopkins, Snoeck and Higham2016). The current radiocarbon age cannot be clearly interpreted and does not clarify the geologic age of the fossils.
Cluster analysis
Taphonomy of the fossils from Bender’s Cave indicates the potential for significant time-averaging, yet the observed patterns of preservation and spatial distribution point to the possibility that the fossils may represent a contemporaneous fauna. That possibility is supported by the fact that the megafauna taxa from Bender’s Cave also co-occur in Late Pleistocene fossil samples from fluvial/alluvial settings in north and east Texas and along the Gulf Coast of Texas that are interpreted as representations of contemporaneous past vertebrate communities (e.g., Stovall and McAnulty, Reference Stovall and McAnulty1950; Lundelius, Reference Lundelius1972). We used hierarchical cluster analysis to explore and quantify the similarity between Bender’s Cave, treating it as a single fauna, and other Late Pleistocene sites in Texas based on the richness of 31 megafauna taxa.
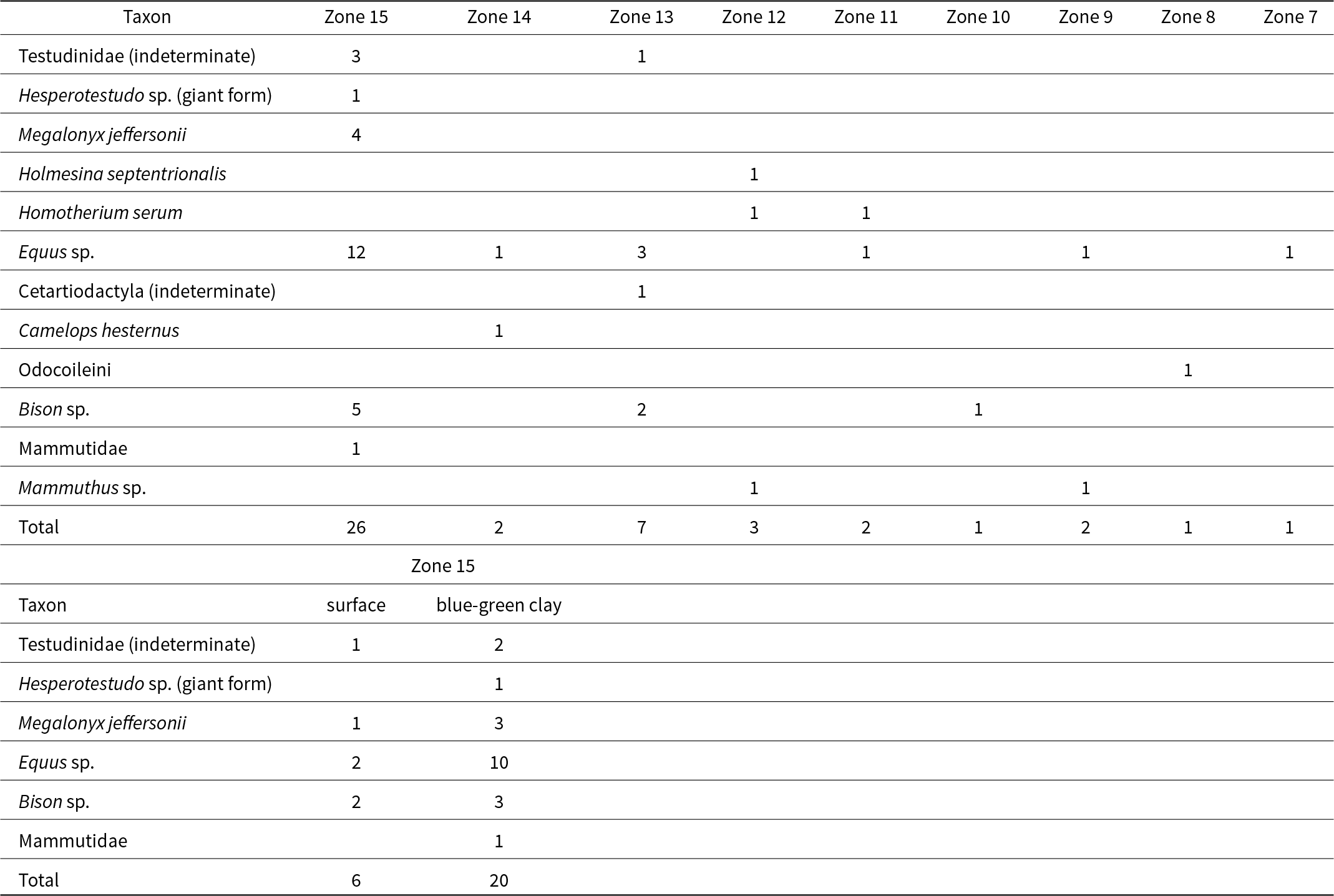
Cluster analysis divided 43 sites into two broad groups (Fig. 12). Those broad groups (i.e., Groups 1 and 2; Fig. 12) were well supported and stable (AU value of 100%), but, in general, the placement of individual sites into smaller, more specific groups was variable. The results were generally consistent with our understanding of the age relationships among sites. Groups 1 and 2 generally corresponded to full/late glacial (MIS 2) and interglacial (MIS 5) sites, respectively, however, that age distribution was not consistent. Group 1 included a mix of interstadial (n = 4) and full/late glacial sites (n = 13). Group 2a (Fig. 12) included a mix of interglacial (MIS 5, n = 2), interstadial (MIS 3, n = 4) and full/late glacial (MIS 2, n = 4) sites. Group 2b was composed of sites interpreted as dating to MIS 5 (n = 8), and Buckner Ranch (Fig. 12). Buckner Ranch contains Holmesina septentrionalis (Supplementary Table 1) and was interpreted as interglacial in age (e.g., Lundelius, Reference Lundelius1972). Recent stratigraphic and geochronologic studies, however, indicated that the associated deposits are Late Pleistocene (< 14.4 ka) and Holocene, and the Late Pleistocene fauna from Buckner Ranch may be a time-averaged composite (Holliday et al., Reference Holliday, Brown, Calame, Frederick, Goode and Nash2025).
Dendrogram results of a hierarchical cluster analysis examining the similarity between megafauna in 43 Late Pleistocene sites in Texas. Numbers at nodes are AU (i.e., approximately unbiased) p-values (0.05). Sites bounded by black boxes represent statistically significant clusters.

Although the arrangement of sites within each of the three major groups was not always well resolved, the topology of four sub-groups (outlined with black rectangles) appeared stable and reliable (AU values of 95% - 100%). Some of the sites in each of those four sub-groups were related. For example, cluster analysis found statistically significant similarity (AU value of 95%) between Gifford-Hill Pit, Moore Pit, and Wood Pit, three localities that were sampled separately (Slaughter et al., Reference Slaughter, Crook, Harris, Allen and Seifert1962), but were later interpreted as representing a single, contemporaneous fauna (Slaughter, Reference Slaughter1966b).
The general division of sites between MIS 2 and MIS 5 and the clustering of known, related sites indicate that the results of the cluster analysis reflect meaningful patterns in the spatio-temporal distribution of megafauna taxa in Texas throughout the Late Pleistocene. The cluster analysis placed Bender’s Cave into Group 2b with, mostly, other interglacial (MIS 5) sites. Notably, Bender’s Cave is the only site in Group 2b from the Edwards Plateau. That result suggests the megafauna identified from Bender’s Cave co-occur elsewhere and could, plausibly be interpreted as a contemporaneous fossil fauna. Further, cluster analysis indicates that the composition of megafauna sampled from Bender’s Cave is most similar, relative to 43 other sites from Texas, to sites interpreted as dating to the last interglacial (i.e., MIS 5).
Discussion
Fossils from Bender’s Cave represent taxa that are common (Mammuthus sp., Equus sp., Camelops hesternus, Bison sp., Odocoileini) to 17 other Late Pleistocene sites across the Edwards Plateau (Fig. 1; including the adjacent Stockton Plateau) alongside other megafauna that are rare or were previously unknown from the region (Lundelius, Reference Lundelius, Martin and Wright1967, Reference Lundelius, Schubert, Mead and Graham2003; Bell et al., Reference Bell, Sagebiel, Lundelius, Czaplewski, Lively, Minatra, Lenamond, Barcelo and Gelnaw2022; Clamp Cave, Natural Bridge Caverns, personal observation, JAM). Bender’s Cave provides the first records of the giant tortoise Hesperotestudo sp. and the pampathere Holmesina septentrionalis from the Late Pleistocene on the Edwards Plateau.
Mastodon and Megalonyx occur in only two or three, respectively, of the 16 regional caves sampled (Mammut americanum: Friesenhahn Cave and Cave Without A Name, Graham, Reference Graham1976a; Megalonyx jeffersonii: Inner Space Cavern Debris Cone 3, Sansom and Lundelius, Reference Sansom and Lundelius2005; Megalonyx sp.: Inner Space Cavern Debris Cone 2, Moretti, Reference Moretti2025; Megalonychidae: Zesch Cave, Sagebiel, Reference Sagebiel2010). A third record of mastodon (Mammut americanum) comes from alluvial plain deposits in the Avenue Site (Lundelius, Reference Lundelius1992; Table 1). Megalonyx does not co-occur with Mammutidae in any other regional site.
Interpreting these novel occurrences and integrating the fossil assemblage from Bender’s Cave into the broader regional record is challenged by uncertainty around the primary source of the fossils and a lack of associated geologic age data. Nonetheless, evidence from Bender’s Cave and from the broader regional record offer context for developing hypotheses regarding the taphonomy of the fossil assemblage and its temporal relationship to other sites.
The primary source of the fossils is not yet known, but Sinkhole #1, a large (∼4.7 m diameter), filled sinkhole at the upstream end of the cave (Fig. 3) is a plausible source of Pleistocene materials. Water in the stream passage eroded trash deposits from Sinkhole #1 and the resulting spatial distribution patterns offer a plausible model for considering the taphonomy of the Pleistocene fossils in Bender’s Cave. Glass, metal, and twentieth century faunal remains occur throughout the passage downstream, including alongside Pleistocene fossils. Erosion and transport acting similarly on hypothetical Pleistocene deposits in Sinkhole #1 could produce the fossil lag assemblage that we observed and sampled. One or multiple intact deposits within the sinkhole could be eroded during periods of flooding and turbulent water flow and the contents of those deposits scattered downstream. The bone breccia and submerged, buried blue-green clay may represent secondary deposits formed during stable, quiet periods of low water flow. Subsequent intervals of turbulent flow could have further eroded the primary deposits and ripped apart secondary deposits, again transporting and re-depositing associated materials and producing the observed pattern of fossils occurring both in isolation and within loose clasts of breccia. The greater concentration of identified fossils in the blue-green clay, including taxa (e.g., Megalonyx jeffersonii) also found in the surface sample, supports this depositional model.
Regardless of the source of the fossils, because erosion and fluvial transport create the potential to mix fossils from different intervals of the geologic past, the samples from Bender’s Cave may be vastly time-averaged and sourced from multiple, distinct primary depositional settings that span significant portions of the Pleistocene, a possibility we cannot fully discount. The taxa recognized so far support interpreting the assemblage as Rancholabrean in composition and Late Pleistocene in age. Bison occurs in samples from the surface and the blue-green clay. The size of the osteoderm of Holmesina septentironalis aligns with Rancholabrean samples (Table 3). Yet, the identified fossils could still be a mix of taxa from throughout the Late Pleistocene or even from the Middle and Late Pleistocene. Such vast time-averaging might help to explain the occurrence of the four regionally novel taxa. However, comparisons with the Late Pleistocene record from across Texas, including our hierarchical cluster analysis results, indicate that the identified taxa are known to co-occur and could be contemporaneous. Our current working hypothesis is that the fossil assemblage documents taxa that were generally contemporaneous and Late Pleistocene in age, as opposed to a mix of taxa from multiple land mammal ages.
Four taxa found in Bender’s Cave, Hesperotestudo sp. (giant form), Megalonyx jeffersonii, Holmesina septentrionalis, and Mammutidae, are novel in the context of the Edwards Plateau and set this site apart from all other Late Pleistocene sites in the region. The scarcity or absence of these taxa was previously interpreted as reflecting the absence of appropriate ecological and/or climatic conditions for those megafauna. The biogeography of Holmesina in North America suggests that it was limited to relatively warm, relatively mesic environmental settings (Edmund, Reference Edmund and Montgomery1987; Pinsof and Echols, Reference Pinsof and Echols1997). Extant tortoises are restricted to tropical and subtropical regions (Brattstrom, Reference Brattstrom1961) and although extinct tortoises may have had different temperature tolerances and/or behavioral mechanisms for dealing with temperature extremes, a poikilothermic physiology would presumably still constrain Hesperotestudo to relatively warm climates (Brattstrom, Reference Brattstrom1961; Graham, Reference Graham and Elliot1986b). Multiple lines of evidence indicate that Megalonyx (Schubert et al., Reference Schubert, Graham, McDonald, Grimm and Stafford2004; McDonald et al., Reference McDonald, Feranec and Miller2019) and Mammut americanum (Birks et al., Reference Birks, van Geel, Fisher, Grimm, Kuijper, van Arkel and van Reenan2018; Smith and DeSantis, Reference Smith and DeSantis2018) were browsers, reliant on C3 woody vegetation and forest habitats. Although the mammutid from Bender’s Cave is currently undetermined, the bunodont cheek teeth of other mammutid taxa suggest a browsing ecology (e.g., McDonald et al., Reference McDonald, Atwater, Dooley and Hohman2020). These four megafauna taxa appear ecologically compatible, a notion supported by their co-occurrence in Florida (Webb and Simons, Reference Webb, Simons and Webb2006) and south and east Texas (e.g., Lundelius, Reference Lundelius1972). We infer that the presence of these four taxa reflects forest habitats and relatively warm, perhaps mesic, conditions in the vicinity of Bender’s Cave, a paleoenvironmental setting that contrasts with the broader regional record.
Most of the 17 Late Pleistocene sites on the Edwards Plateau date to MIS 2 (Table 1; Toomey, Reference Toomey, Elliott and Veni1994; Lundelius, Reference Lundelius, Schubert, Mead and Graham2003) and the associated record of flora and fauna documents a relatively dry, open grassland from approximately 20,000–11,700 cal BP (Lundelius, Reference Lundelius, Martin and Wright1967; Cordova and Johnson, Reference Cordova and Johnson2019). Pollen and phytoliths from ca. 18.1–11.7 ka at Hall’s Cave (Cordova and Johnson, Reference Cordova and Johnson2019) and pollen from ca. 20–15 ka (estimated age, see Graham et al., Reference Graham, Lundelius, Meissner, Muhlestein, Hunt and Catlos2013, for details) at Friesenhahn Cave (Hall and Valastro, Reference Hall and Valastro1995) indicate open grasslands with sparse trees (primarily Pinus) on the uplands and more diverse tree communities (e.g., Picea, Juglans, Alnus) within river valleys. Multiple lines of evidence indicate that, from ca. 21–11 ka, the region experienced a general drying trend coupled with the erosion and loss of the upland soil at a rate of ca. 11 cm/1000 years (Toomey et al., Reference Toomey, Blum and Valastro1993; Musgrove et al., Reference Musgrove, Banner, Mack, McCombs, James, Cheng and Edwards2001; Cooke et al., Reference Cooke, Stern, Banner, Mack, Stafford and Toomey2003). The Bølling–Allerød marks a brief hiatus from these broader paleoenvironmental trends, with a shift to wetter conditions and evidence of woodland habitats (Joines, Reference Joines2011; Seersholm et al., Reference Seersholm, Werndly, Grealy, Johnson, Keenan Early, Lundelius and Winsborough2020). Aside from that short-lived return to interstadial conditions, the known floral and environmental trends of MIS 2 do not appear conducive for the presence and maintenance of widespread forest habitats.
The sole radiocarbon age (17,330–17,030 cal BP) available for Bender’s Cave is flawed but indicates that the fossil assemblage falls within MIS 2 and is contemporaneous with the rest of the Late Pleistocene record from the Edwards Plateau, a possibility that needs to be considered. Hesperotestudo sp. (giant form), Megalonyx jeffersonii, Holmesina septentrionalis, and Mammutidae may have been rare on the Edwards Plateau during any time interval, including MIS 2, and paleontological sampling of 17 regional sites may not be extensive enough to recover those rare taxa. Rather, those taxa may appear to be novel simply due to sampling biases. That scenario is likely for Homotherium serum, found in only two other regional sites (Friesenhahn Cave, Inner Space Cavern Debris Cone 2), but would be surprising for the megafaunal herbivores identified here, particularly since preservation of osteoderms from Hesperotestudo sp. (giant form) and Holmesina septentrionalis should be favored by taphonomic biases. The radiocarbon age is problematic, given the potential for geochemical contamination of the dated bioapatite. More importantly, the radiocarbon age appears to be discordant with the rest of the regional record.
Bias may be driving the observed patterns, but extensive sampling of regional Late Pleistocene deposits combined with a lack of evidence of forest habitats hints that the occurrence of the novel megafauna is better explained by environmental variation associated with the cyclical climatic oscillations of the Pleistocene. Climatic change associated with the glacial–interglacial cycle prompted regional changes in habitats and associated biota elsewhere in North America, shifting between interglacial/interstadial forests and glacial steppe or savanna (McDonald et al., Reference McDonald, Harington and De Iuliis2000; Saunders et al., Reference Saunders, Grimm, Widga, Campbell, Curry, Grimley, Hanson, McCullum, Oliver and Treworgy2010; Zazula et al., Reference Zazula, MacPhee, Southon, Nalawade-Chavan, Reyes, Hewitson and Hall2017). A similar glacial–interglacial dynamic in central Texas could explain the novel megafauna in Bender’s Cave.
Warmer and/or wetter conditions, often attributed to interglacial and interstadial intervals (Hibbard and Taylor, Reference Hibbard and Taylor1960; Kurtén and Anderson, Reference Kurtén and Anderson1980; Karrow, Reference Karrow1990; Otvos, Reference Otvos2015), appear consistent with the presence of Hesperotestudo sp. (giant form), Holmesina septentrionalis, and forest habitats, inferences suggesting that fossils from Bender’s Cave date to the last interglacial (i.e., MIS 5) or interstadial (i.e., MIS 3). Interglacial/interstadial faunas are often difficult to identify, especially because they lie near, or beyond, the limits of radiocarbon dating. The sole available radiocarbon age from Bender’s Cave could be interpreted as indicating an age within MIS 3 (57–29 ka). MIS 5 (130–71 ka) is entirely beyond the range of radiocarbon, but erroneous radiocarbon ages in the range of MIS 2 have been reported for MIS 5 sites (e.g., Smith and Polly, Reference Smith and Polly2013; Zazula et al., Reference Zazula, MacPhee, Southon, Nalawade-Chavan, Reyes, Hewitson and Hall2017).
Faunal correlation offers another means of recognizing interglacial/interstadial faunas, but any potential faunal distinction between MIS 3 and MIS 5 in Texas is unclear and fossil assemblages from both intervals have been conflated previously as “Sangamonian” in age (e.g., see discussions in Lundelius, Reference Lundelius1972; Pinsof, Reference Pinsof, Stewart and Seymour1996; Otvos, Reference Otvos2015). There are no known sites on the Edwards Plateau that date to MIS 5. Three adjacent sites at the north end of Inner Space Cavern contain components dating to interstadial MIS 3 (Moretti, Reference Moretti2025). Limited age data make it difficult to resolve the faunal composition of the interstadial fauna in Inner Space Cavern, but Megalonyx sp., Dasypus cf. D. bellus, and a large form of Antilocapridae all occur in deposits older than 20,000 cal BP (Moretti, Reference Moretti2025).
The co-occurrence in Bender’s Cave of Hesperotestudo sp. (giant form), Megalonyx, Holmesina, and Mammutidae is distinct from other Late Pleistocene faunas from the Edwards Plateau but is similar to faunas from elsewhere in Texas (e.g., Iron Bridge, Moore Pit, Ingleside, Inner Space Cavern) that are interpreted as dating to MIS 3 and MIS 5. Cluster analysis did not group Bender’s Cave with Inner Space Cavern, a result that may stem from the absence of Bison and Holmesina from Inner Space Cavern. Instead, cluster analysis highlighted the similarity between the assemblage from Bender’s Cave and interglacial (MIS 5) sites in north, east, and southern Texas (Table 1, Fig. 12). An interglacial (MIS 5) age, or interstadial (MIS 3) age, for Bender’s Cave offers the most parsimonious interpretation of the available data and allows us to interpret the distinctive fauna within the broader Late Pleistocene record from the Edwards Plateau and Texas.
The existing regional floral, faunal, and chronological data, taken at face value, indicate that Late Pleistocene fossils in Bender’s Cave may date to a warm climatic interval, prior to the LGM, potentially the last interglacial, MIS 5. New radiometric age data are clearly needed to test this hypothesis and ongoing field and lab work is focused on this critical area. Additional radiocarbon dating of fossils is likely to produce additional problematic results, so instead we are focusing on U–Th dating of speleothems associated with fossils, coupled U-series and electron spin resonance dating of mammal teeth (e.g., Withnell et al., Reference Withnell, Joannes-Boyau and Bell2020), and radiocarbon dating of other organics (if they exist) from the blue-green clay or other intact deposits.
The presence of the novel megafauna in Bender’s Cave is important, regardless of the underlying cause(s) of their occurrence. The faunal distinctions between Bender’s Cave and the 17 other sites on the Edwards Plateau have implications for understanding Late Pleistocene vertebrate biogeography and provincialism, as well as the biases associated with fossil samples. If the megafauna from Bender’s Cave appear novel only as the product of sample bias, then that sample bias is significant and recognizing it is essential to understanding regional natural history and to interpreting fossil assemblages as past biotic communities. Sample bias would imply that extensive sampling of 17 regional sites was insufficient to fully document regional Late Pleistocene paleoecology and megafaunal diversity. If the observed patterns of species occurrence in Bender’s Cave are not the product of sample bias, then this cave is providing previously unrecognized evidence of habitat diversity and/or intra-Pleistocene faunal change. Either outcome would be significant and an indication that our understanding of the fossil record of the Pleistocene and its implications for natural history remain incomplete.
Conclusions
Sampling of lag assemblages in Bender’s Cave produced a diverse sample of Pleistocene megafauna including taxa common to 17 Late Pleistocene sites elsewhere on the Edwards Plateau of central Texas. The co-occurrence of four taxa (Hesperotestudo sp. [giant form], Megalonyx jeffersonii, Holmesina septentrionalis, Mammutidae) that are novel in the region distinguishes Bender’s Cave from other regional sites and raises questions about the association and geologic age of the fossils. The common and novel taxa may be sourced from two (or more) separate deposits and may represent distinct, non-contemporaneous past animal communities.
Yet, the taphonomic modification and spatial distribution of the fossils encourage viewing the taxa as comprising a single fossil assemblage. Most other regional sites date to, in part or in whole, MIS 2 and are younger than 20,000 cal BP. The composition of the Bender’s Cave fauna is similar to sites from other regions of Texas interpreted as dating to MIS 3 or MIS 5. Regional climate, paleoenvironments, and fauna may have been different during those earlier interstadial/interglacial intervals, relative to MIS 2, and those differences could explain the novel record from Bender’s Cave.
Still, our understanding of the geochronology and biota of warm intervals prior to the LGM is relatively unresolved. Reliable age data are limited even for sites that fall within MIS 2, including on the Edwards Plateau, and the lack of radiometric ages and ongoing reliance on outdated, unreliable ages (e.g., Cave Without a Name, Friesenhahn Cave) make it difficult to resolve faunal dynamics and interpret the novel fauna of Bender’s Cave. Addressing key questions about the fauna from Bender’s Cave will require re-visiting the faunal composition and chronology of Late Pleistocene sites across Texas.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/qua.2025.10071.
Acknowledgments
We thank the Texas Cavers who came before us for their pioneering exploration and documentation of regional caves, this research would not be possible without their efforts. Pleistocene fossils in Bender’s Cave were first observed and reported by cavers. Cavers (including author J. Young) also developed the relationships with relevant landowners that granted us access and permission to collect fossils. We thank landowners L. and B. McAllister for providing access to their property and for their generous donation of the fossils discovered in Bender’s Cave. JAM thanks C. Bell, R. Martindale, B. Schubert, and D. Stockli for comments and suggestions that improved the quality of this research and manuscript. S. Skwarcan instructed us on specimen photography. C. Williams and J. Jacisin gave advice on statistical methods. Hierarchical clustering was performed, in part, using code developed and provided by D. Muscente. This research was facilitated by lab activities performed by B. Bustos, M. Frazier, C. Cabrera, E. Burton, and A. Smith. Funding for fieldwork and radiocarbon analyses was provided by support from the Jackson School of Geosciences of The University of Texas at Austin.

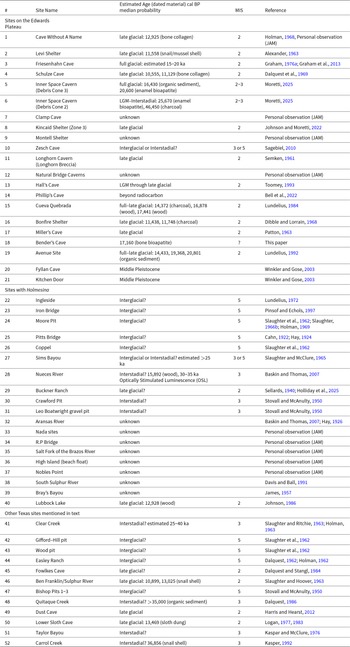

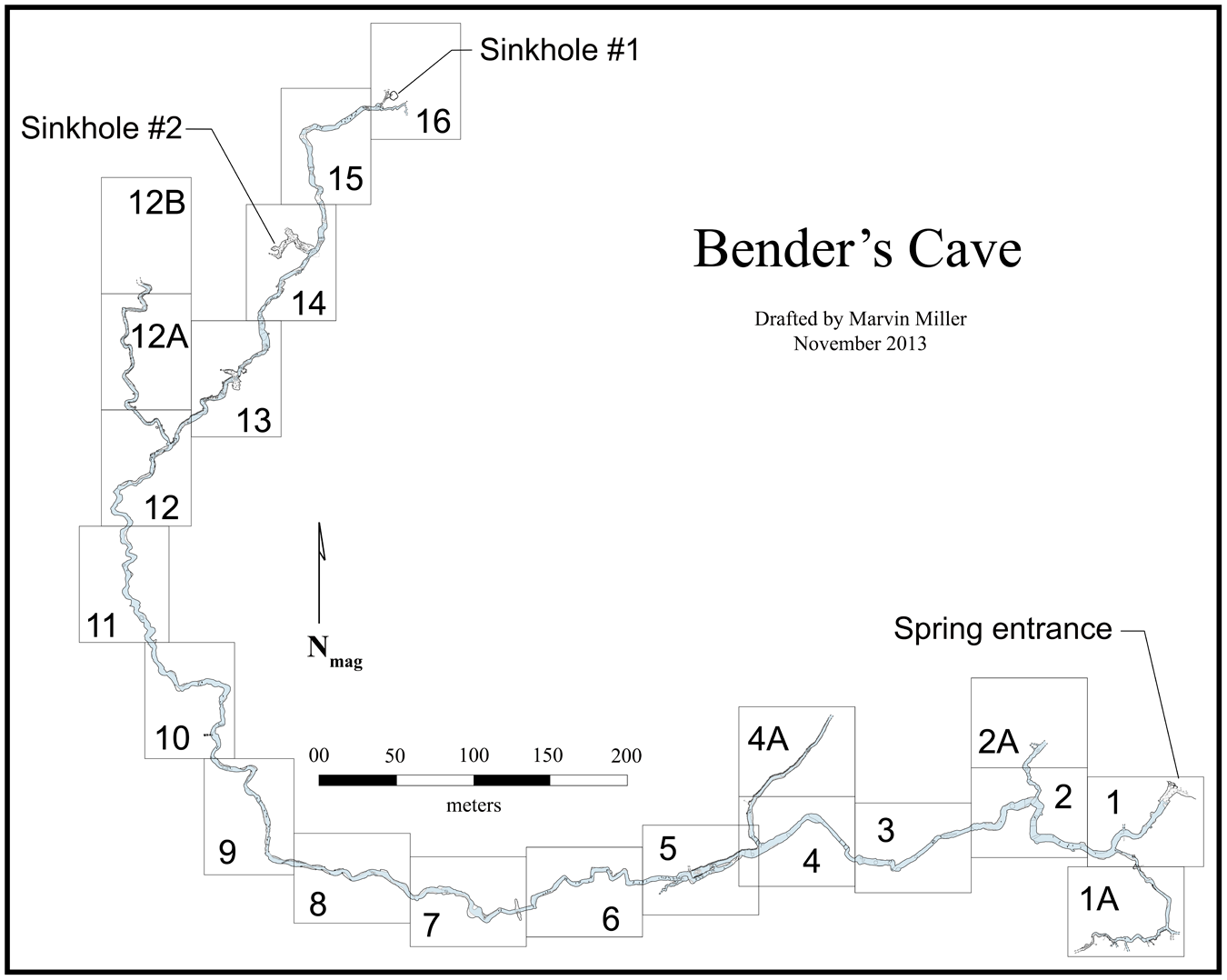
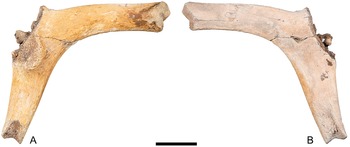
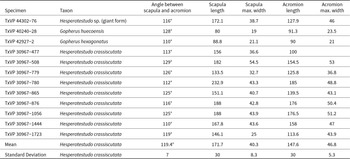

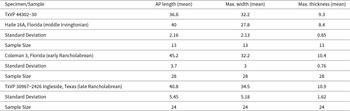

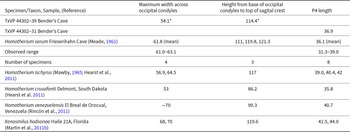
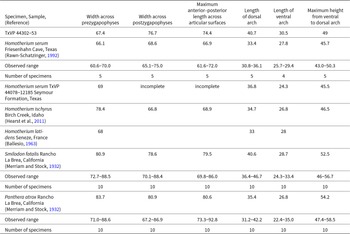

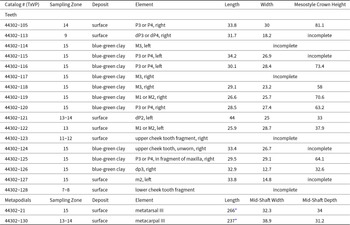
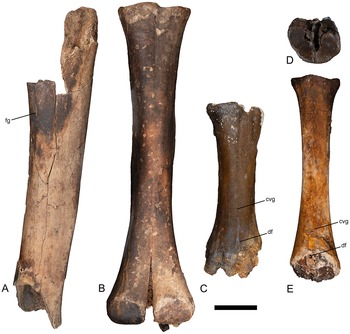

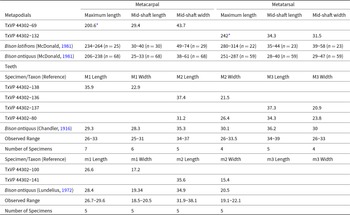

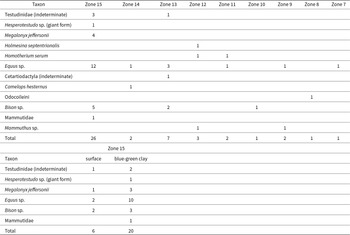





 Open access
Open access