


Introduction
Rhizoliths (from Greek “rhizo” meaning root and “lithos” meaning stone) also known as rhizoconcretions, rhizomorphs, root casts, and osteokollen, are mineralized and calcified structures that develop around roots (e.g., Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011). These structures, typically cylindrical in shape, are composed of minerals such as calcite, opaline silica, fluorite, and analcime (Owen et al., Reference Owen, Owen, Renaut, Scott, Jones and Ashley2008). The formation of rhizoliths occurs through the accumulation and crystallization of minerals, predominantly calcium carbonate (CaCO3), around decaying roots or root channels (Klappa, Reference Klappa1980; Jones and Kahle, Reference Jones and Kahle1986; Alonso-Zarza et al., Reference Alonso-Zarza, Sanz, Calvo and Estévez1998). They can be oriented vertically, sub-vertically, or horizontally in soils, reflecting root growth patterns or the effects of erosion (Sun et al., Reference Sun, Huguet and Zamanian2021, Reference Sun, Zamanian, Huguet, Bayat, Wang and Badawy2022). While identifiable root materials are generally absent, occasional preservation does occur (McLaren, Reference McLaren1995; Gocke et al., Reference Gocke, Peth and Wiesenberg2014; Sun et al., Reference Sun, Xue, Zamanian, Colin, Duchamp-Alphonse and Pei2019b, Reference Sun, Zamanian, Huguet, Fa and Wang2020; Liu et al., Reference Liu, Li, Dong, Deng, Liu and Gao2024). The sizes of rhizoliths vary considerably, ranging from approximately 0.5 mm to over 30 cm in diameter, with lengths reaching several meters (Löscher and Haag, Reference Löscher and Haag1989; Alonso-Zarza et al., Reference Alonso-Zarza, Genise, Cabrera, Mangas, Martín-Pérez, Valdeolmillos and Dorado-Valiño2008). These structures illustrate the complex interplay between biological activity and geochemical processes (Rodríguez-Aranda and Calvo, Reference Rodríguez-Aranda and Calvo1998; Polymeris et al., Reference Polymeris, Kitis, Kiyak, Theodosoglou, Tsirliganis, Ertek and Erginal2016). As roots decay, the inner parts of rhizoliths may be filled with clay particles and minerals, further contributing to their intricate internal structures (Gocke et al., Reference Gocke, Peth and Wiesenberg2014; Liu et al., Reference Liu, Li, Dong, Deng, Liu and Gao2024).
Rhizoliths are valuable paleoenvironmental indicators, providing insights into past soil and vegetation conditions (Gao et al., Reference Gao, Li, Wang and Li2019). Their presence in alluvial sediments signals the existence of paleosols (Retallack, Reference Retallack1990) and paleo-vegetation (Wang and Greenberg, Reference Wang and Greenberg2007). The δ13C values of carbonates within rhizoliths, derived from root-respired CO2, offer clues about historical soil environments and climatic conditions (Smith and Epstein, Reference Smith and Epstein1971; Wang et al., Reference Wang, Ambrose and Fouke2004; Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011). Rhizoliths in aeolian deposits, such as loess (Huguet et al., Reference Huguet, Wiesenberg, Gocke, Fosse and Derenne2012) and sand dunes (Cramer and Hawkins, Reference Cramer and Hawkins2009; Sun et al., Reference Sun, Zamanian, Huguet, Wiesenberg, Zhao and Lei2023), help differentiate among deposition phases, stabilization, plant colonization, and soil formation (Retallack, Reference Retallack1990). Rhizolith formation requires water, making them indicators of past hydrological conditions and climates (Alonso-Zarza et al., Reference Alonso-Zarza, Genise, Cabrera, Mangas, Martín-Pérez, Valdeolmillos and Dorado-Valiño2008; Sun et al., Reference Sun, Zamanian, Huguet, Bayat, Wang and Badawy2022). They can reveal fluctuations in paleo-water tables (Mount and Cohen, Reference Mount and Cohen1984; Hembree and Hasiotis, Reference Hembree and Hasiotis2007; Greenbaum et al., Reference Greenbaum, Schwartz, Benito, Porat, Cloete and Enzel2014) and paleo-sea levels (Froede, Reference Froede2002). Additionally, rhizoliths are useful for radiocarbon dating (Cramer and Hawkins, Reference Cramer and Hawkins2009; Tabor and Myers, Reference Tabor and Myers2015).
Rhizolith formation is evidenced by centripetal CaCO3 accumulation, where the CaCO3 content decreases outward from the rhizolith into the surrounding sediment or soil (Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011, Reference Gocke, Kessler, van Mourik, Jansen and Wiesenberg2016). This pattern supports the idea that CaCO3 precipitates preferentially near roots during periods of water uptake, with root-derived CO2 serving as the primary carbon source (Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011; Zamanian and Kuzyakov, Reference Zamanian and Kuzyakov2019; Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021). Radiocarbon data further suggest that rhizoliths form predominantly during the root’s active growth phase, utilizing carbon from root respiration (Jaillard, Reference Jaillard1992; Becze-Deàk et al., Reference Becze-Deàk, Langohr and Verrecchia1997).
The timing and mechanisms of rhizolith formation, however, are subjects of ongoing debate. They may form during the root’s life, driven by water uptake and Ca2+ precipitation (Hinsinger et al., Reference Hinsinger, Gobran, Gregory and Wenzel2005; Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011), or post-mortem, influenced by microbial decomposition and nutrient (such as Ca2+) release (Braissant et al., Reference Braissant, Cailleau, Aragno and Verrecchia2004; Cailleau et al., Reference Cailleau, Braissant and Verrecchia2011, Reference Cailleau, Mota, Bindschedler, Junier and Verrecchia2014; Sun et al., Reference Sun, Huguet and Zamanian2021). This ambiguity complicates their use in paleoenvironmental reconstructions and dating, as discrepancies in radiocarbon ages between root remnants and cementing carbonates, as well as between rhizoliths and their hosting soil/strata persist (Sun et al., Reference Sun, Wang and Zamanian2019a; Zhao et al., Reference Zhao, Zhao, Stahr, Kuzyakov and Wei2020).
Understanding the formation rates and conditions of rhizolith development is therefore crucial for their application in environmental and chronological studies. This study applies numerical simulations with the HYDRUS-1D model to identify and quantify these processes, examining how varied boundary conditions influence root–soil–water interactions and rhizolith formation in calcareous soils under semi-arid environments.
Methods
HYDRUS-1D, coupled with the PHREEQC model, was used to reproduce realistic Ca2+ distributions. The model accounts for dynamic variables affecting Ca2+ accumulation and redistribution in the soil profile. The coupled HYDRUS-1D algorithms that compute dynamic plant water uptake make it effective in simulating evapotranspiration-induced Ca2+ redistribution and its effects on calcification around the root on daily timescale and in high spatial resolution (Šimůnek et al., Reference Šimůnek, Šejna, Saito, Sakai and van Genuchten2009b). Calcite precipitation occurs when the solution becomes super-saturated with Ca2+ (Drever, Reference Drever1982). The maximum Ca2+ concentration, [Ca2+], at the rhizosphere was assumed to be 3 mmol/L due to relatively higher CaCO3 solubility at higher CO2 partial pressure (pCO2 = 2%) close to the root (Zamanian et al., Reference Zamanian, Pustovoytov and Kuzyakov2016). Therefore, to calculate the period necessary for rhizolith formation we modeled changes in [Ca2+] distant to the root over time. When [Ca2+] reaches 3 mmol/L, then CaCO3 precipitates and rhizolith formation begins. HP1, the coupled model, uses an operator-splitting approach to integrate the hydrological and geochemical models (Jacques and Šimůnek, Reference Jacques and Šimůnek2005; Šimůnek et al., Reference Šimůnek, Jacques, van Genuchten and Mallants2006, Reference Šimůnek, van Genuchten and Šejna2008). HYDRUS-1D simulates water, solutes, and heat transport through fully saturated, partially saturated, or unsaturated porous media (Šimůnek et al., Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009a, Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009b) by calculating pressure head changes, water fluxes, and solute concentrations at each time step. Once these transport processes are computed, PHREEQC-2 (Parkhurst and Appelo, Reference Parkhurst and Appelo1999) models the geochemical reactions, such as changes in [Ca2+] and calcite precipitation (Šimůnek et al. Reference Šimůnek, Jacques, van Genuchten and Mallants2006, Reference Šimůnek, van Genuchten and Šejna2008) within the HYDRUS–PHREEQC (HP1) coupling framework (Jacques et al., Reference Jacques, Šimůnek, Mallants and van Genuchten2006).
We used a 100 cm deep silty loam profile, representing soil texture formed on loess deposits (Liu, Reference Liu1985) as a one-dimensional domain (Fig. 1). The profile was divided into 101 nodes, providing high spatial resolution to effectively resolve steep gradients with minimal computational errors. Root water uptake (Feddes et al., Reference Feddes, Kowalik and Zaradny1978) was modeled across nodes 0–51 (50-cm rootzone depth), with homogenous root distribution (Jackson et al., Reference Jackson, Canadell, Ehleringer, Mooney, Sala and Schulze1996; Cai et al., Reference Cai, Vanderborght, Couvreur, Mboh and Vereecken2017). This constrained homogenous root water uptake was applied to simulate field-observed radial/centripetal CaCO3 accumulation (McLaren, Reference McLaren1995) around a single root (Sun et al., Reference Sun, Zamanian, Huguet, Wiesenberg, Zhao and Lei2023). Calculations at specified nodes of interest (Fig. 1) were used to monitor Ca2+ redistribution and calcification at 0.2-, 1-, 2-, 3-, 4-, and 10-cm radius references to the root surface (at node 51).
Scheme of the manipulated conditions tested for rhizolith formation simulation. Root suction (green) is defined at 0–50 cm of a 100 cm profile (brown vertical line). The dual moisture profile shows drier topsoil (0–20 cm, blue line, = −1000 hPa) overlying wetter subsoil (20–100 cm, light green line, ψ = −100 hPa). Four initial [Ca2+] were homogenized in the soil (shades of gray vertical lines). Ca2+ fluxes were driven to the root at 0.03 cm/d and 1 cm/d (red arrows). Observations at 0.2, 1, 2, 3, 4, and 10 cm (dots) monitored calcification reference to the root surface (see also Fig. 8 for overview).

To ascertain this concentric calcification assumption, root water uptake was mathematically resolved at a point (node 31) and compared to the calcification around the single root surface (node 51). Feddes parameters were described as −15, −120, −1500, and −8000 cm, representing the matric potential head thresholds marking the onset (hVo), peak (hVmax), maximum sustained rate (hVmax!), and cessation (hԑ) of root water extraction, respectively. Therefore, the simulation becomes hydraulically constrained when atmospheric demand via evapotranspiration depletes the water reservoir and the matric potential head approaches −8000 cm across all points within the 100 cm profile. Note that the hԑ value constraining root water uptake may vary based on plant physiology, age, and type (Bouma, Reference Bouma2018). The hVmax! value was set at −1500 cm, taking into account the drier profile (ho = −1000 cm) considered, such that ho > hVmax!.
Water flow rates for evapotranspiration were set at 0.03 cm/d (low) and 1 cm/d (high) to simulate conditions observed in semi-arid environments (Szutu and Papuga, Reference Szutu and Papuga2019). The dependence of root water and solute uptake on solute concentration, soil moisture dynamics, and temperature gradient can be described using Cauchy/Neumann (flux conditions) and/or Dirichlet-type models (van Genuchten and Parker, Reference van Genuchten and Parker1984). We imposed atmospheric conditions with a surface layer (Neumann Type 3) and a constant pressure head (Dirichlet Type 1) for the upper and lower boundaries, respectively. The former was used to define ETo and specify solution types, whereas solution compositions were defined in the coupled HP1 model. For solution composition in HP1 definitions, two solutions were defined. Rhizospheric soil solution (pH 6.5) constrained rhizospheric biogeochemistry and was set in equilibrium with atmospheric O2 (log pO2 = −0.68) and average soil CO2 (log pCO2 = −2.22) to simulate soil (root + microbial) respiration-driven bicarbonate formation (Jacques and Šimůnek, Reference Jacques and Šimůnek2005; Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025). At this pH, dissolved CO2 undergoes hydrolysis to form carbonic acid (Equation 1), which dissociates into bicarbonate and protons (Equation 2).
 \begin{equation}{\text{C}}{{\text{O}}_{\text{2}}}{\text{ + }}{{\text{H}}_{\text{2}}}{\text{O}} \rightleftharpoons {{\text{H}}_{\text{2}}}{\text{C}}{{\text{O}}_{\text{3}}}\end{equation}
\begin{equation}{\text{C}}{{\text{O}}_{\text{2}}}{\text{ + }}{{\text{H}}_{\text{2}}}{\text{O}} \rightleftharpoons {{\text{H}}_{\text{2}}}{\text{C}}{{\text{O}}_{\text{3}}}\end{equation} \begin{equation}{{\text{H}}_{\text{2}}}{\text{C}}{{\text{O}}_{\text{3}}} \rightleftharpoons {\text{HC}}{{\text{O}}_{\text{3}}}^{-}{{ + }}{{\text{H}}^{+}}\end{equation}
\begin{equation}{{\text{H}}_{\text{2}}}{\text{C}}{{\text{O}}_{\text{3}}} \rightleftharpoons {\text{HC}}{{\text{O}}_{\text{3}}}^{-}{{ + }}{{\text{H}}^{+}}\end{equation}Since pH 6–7 favors HCO3– as the dominant dissolved inorganic carbon (DIC) species (namely CO2-solute (aq), H2CO3 (aq), HCO3− (aq), CO32− (aq)) in the carbonate equilibria, the system was initialized with 1 mmol/L HCO3− to reflect this balance (Jacques and Šimůnek, Reference Jacques and Šimůnek2005). The boundary bulk soil solution (pH 7.5) contained four predetermined initial homogeneous [Ca2+] concentrations of 0.13, 0.15, 0.3, and 1 mmol/L (Fig. 1), transported towards the rhizosphere via root water uptake, representing 4%, 5%, 10%, and 33% of the threshold [Ca2+] for calcification (Zamanian et al., Reference Zamanian, Pustovoytov and Kuzyakov2016). As Ca2+ accumulates over time, it reacts with bicarbonate to form CaCO3 (Equation 3).
 \begin{equation}{\text{C}}{{\text{a}}^{{{2 + }}}}{\text{ + 2HC}}{{\text{O}}^{{{3 - }}}} \rightleftharpoons {\text{CaC}}{{\text{O}}_{{3}}} + {\text{C}}{{\text{O}}_{{2}}} + {{\text{H}}_{\text{2}}}{\text{O}}\end{equation}
\begin{equation}{\text{C}}{{\text{a}}^{{{2 + }}}}{\text{ + 2HC}}{{\text{O}}^{{{3 - }}}} \rightleftharpoons {\text{CaC}}{{\text{O}}_{{3}}} + {\text{C}}{{\text{O}}_{{2}}} + {{\text{H}}_{\text{2}}}{\text{O}}\end{equation}The equilibrium phase of the geochemical model was initialized with zero calcite to allow dynamic precipitation when Ca2+ approaches the 3 mmol/L threshold for onset of rhizolith formation. Isothermal conditions at 25°C were assumed to evaluate [Ca2+] and moisture constraints. Initial conditions considered constant matric potential heads of −1000 cm and −100 cm to represent profile conditions at intermediate soil-water (hI) and field capacity (hFC), respectively, for the presumably drought-resistant roots in (semi) arid conditions. A dual moisture layer profile with drier (h = −1000 cm) topsoil (z = 0–20 cm) and optimal moisture (h = −100 cm) at deeper zones (z = 20–100 cm) often observed in semi-arid environments (Fig. 1; Sanchez‐Mejia and Papuga, Reference Sanchez-Mejia and Papuga2014) was also considered. The hydraulic properties of the loess were described by van Genuchten parameters (Table 1) and were defined using the (silty loam) class pedotransfer function of Carsel and Parrish (Reference Carsel and Parrish1988).
van Genuchten parameters that describe the hydraulic properties of the loess, namely the residual water content θr, the water content at saturation θs, the air entry suction (α), the shape fitting parameter (n), and the saturated hydraulic conductivity (Ks); with m constrained as 1−1/n.

Simulations held boundary conditions constant with no flux through the boundaries preventing Ca2+ leaching. The limiting factors for rhizolith formation were first evaluated in 1- and 5.5-year simulation runs, as presented in the supplementary material (S), followed by the extensive simulation detailed here.
Results
Hydraulic and time constraints of the model
The model was resolved to the time point when the suction head gradient approached zero to assess the maximum time constraints and the behavior of the calcification curve at this point. The profile information in Figure 2a and 2b demonstrates a distinctive suction gradient, where the matric potential heads along the root (|z| = 0–50 cm) were more negative compared to the underlying soil, establishing a suction head gradient. This gradient drives positive fluxes from below towards the root. Over time, the matric potential head progressively becomes even more negative, reaching a value of −8000 cm at ca. 500 years. Throughout the simulation run, a suction head gradient was induced towards the root (|z| = 0–50 cm) except at the 500-year mark, where the matric potential heads uniformly become −8000 cm across the entire profile. This uniformity results in no net force to drive water within the profile. Consequently, the time-dependent equilibrium matric potential head (hԑ) was identified as −8000 cm, with the corresponding time (tԑ) marked at 500 years. Water content shown in Fig. 2c and 2d follows a similar trend to the pressure heads, with the equilibrium moisture content (θԑ) recorded at 0.089 cm3/cm3 which is 137% of the residual water content (Table 1). Additionally, the hydraulic capacity curves displayed in Fig. 2e and 2f have a similar trend to the above (matric potential heads and water retention profiles).
Profile information illustrating the development of matric potential heads following root water uptake (|z| = 0–50 cm) from h = −100 cm in panel (a) and h = −1000 cm in panel (b) over time, from T0 to equilibrium matric potential head, hε of −8000 cm across the profile at T500 yrs as the hydraulic capacity equalizes. Panels (c) and (d) show the corresponding water content profiles, while (e) and (f) display the hydraulic capacity curves along the profile. See also Supplementary Material (Fig. S1) for zoomed-in root pressure effect.

When the model was resolved for point root water uptake (|z| = 30 cm), similar hydraulic redistribution patterns emerged (Fig. 3). Here, negative fluxes from above and positive fluxes from below were forced toward the induced suction at a 30-cm depth. However, equilibrium took longer to achieve, extending to 1000 years due to greater water availability around the point root than in the deep root scenario. This approach allowed for comparative analysis of calcification surrounding the point and single root validating the concentric calcification assumption.
Profile information illustrating the development of matric potential heads following cycles of point root water uptake (|z| = 30 cm) from h = −100 cm in panel (a) and h = −1000 cm in panel (b) over time, from T0 to equilibrium matric potential head, hε of −8000 cm across the profile at T1000 yrs as the hydraulic capacity (in panels a and b) equalizes. Panels (c) and (d) show the corresponding water content profiles, while (e) and (f) display the hydraulic capacity curves along the profile.

Water fluxes were driven across a suction head gradient from the soil toward the root, with higher fluxes observed under optimal conditions (h = −100 cm; Fig. 4a and 4c) compared to arid conditions (h = −1000 cm; Fig. 4b and 4d). After one year, fluxes to the root considerably decrease, stabilize over 50 years, and approach zero by the end of 500 years.
Hydraulic and temporal constraints within the model. In the upper panel, water fluxes are forced towards the surfaces (RS) of deep root (a and b) and point root (c and d) over time until reaching 500 years, at which point zero-suction gradient is attained (indicated by horizontal arrows). As fluxes diminish, root water uptake drastically declines (in panel e) even in the short term and stabilize for both 0.03 and 1 cm/d water flow rates. In the long term (panel f), water uptake approaches zero. Note different scales on the x-axes.

In the short term, under optimally moist soil profiles (h = −100 cm), water fluxes to the root have a significant impact on root water uptake, particularly at higher evapotranspiration rates (ETo = 1 cm/d). However, root water uptake decreases drastically for ETo = 1 cm/d compared to ETo = 0.03 cm/d. By 30 days, root water uptake at ETo = 1 cm/d falls and equalizes with that of ETo = 0.03 cm/d (Fig. 4e), after which it further declines below that of ETo = 0.03 cm/d. By 220 days, root water uptake gradually decreases to 0.01 cm/d for ETo = 0.03 cm/d, reaching equilibrium with that of ETo = 0.03 cm/d over the 400-day simulation period.
Under dry conditions (h = −1000 cm), water flow rates show no influence on root water uptake. Considering the 500-year simulation run, root water uptake at ETo = 0.03 cm/d equalizes for both optimally moist profiles (h = −100 cm) and arid conditions (h = −1000 cm) at 30 years (Fig. 4f), becomes negligible at 50 years, and reaches zero from ca. 374 years onwards as the water reservoir depletes. In the deep root scenario, root water uptake is homogeneous across the 0–50 cm depth, hence, negative/downward fluxes across this range are irrelevant, while upward/positive fluxes towards the root dominate. This upward movement enables Ca2+ redistribution around the root surface at 50 cm depth.
Rhizolith formation and pore-jamming effect at equilibrium simulation run
Knowledge about the finite hydraulic and time constraints (Table 2) of the model provided insights into the shape and behavior of the calcification curve in the long run. During hydraulic redistribution, calcification gradually jams pore spaces, reflecting calcite cementation, and inversely affecting hydraulic properties (Table 3; Fig. S4). The equilibrium water content (θԑ = 0.089 cm3/cm3) defines the maximum soil pore volume available for calcification. The porosity (ɸ) of loess with bulk (ρb) and particle (ρs) densities of 1.8 and 2.65 g/cm3, respectively is 0.32 cm3/cm3 (Table 2). The maximum pores available for calcite saturation (ɸCasat = ɸ − θԑ) equals 0.23 cm3/cm3, which is 72% (i.e., ɸCasat % = (ɸ − θԑ)/ɸ) of the total pore volume (Table 2 and 3). This implies that more pores are available for calcium saturation for larger volume of soil. For instance, for 0.2-cm proximity around the root, total pore volume, ɸz=0.2, of 0.0128 cm3/cm2 provides maximum pore volume of 0.009 cm3 for calcite saturation. Therefore, at equilibrium (θԑ = 0.089 cm3/cm3), up to 0.0025 g of calcite with particle density 2.7 g/cm3 completely saturates the pores at 0.2-cm proximities to the root. Calcite saturation concentration ([Casat]) at any given proximity to the root (i.e., 0.2, 1, 2, 3, 4, 10 cm) was then deduced (Table 3).
Initial matric potential (ho), time to a halt in root water uptake (Troot), matric potential (hԑ) and water content (θԑ) at equilibrium, time to zero-suction gradient (Tԑ), loess porosity (ɸ), and threshold porosity for calcite saturation (ɸCasat).

Area at a given radius (z, cm) from the root surface (Az, cm2), total pore volume at a given radius (ɸvz, cm3), threshold pore volume for calcite saturation (ɸv Casat, cm3), threshold cumulative calcite saturation mass, moles, and concentration, respectively, m Casat (g), n Casat (mol), and [Casat] (mol/ L).

At the equilibrium simulation run (tԑ = 500 years), three shapes of the calcification curve are observed: the hydraulically constrained equilibrium/equilibrium departure (Dԑ), the pre-saturation equilibrium (So), and the saturation equilibrium (Sԑ) (Fig. 5). Considering a drier profile (h = −1000 cm) with 0.15 mmol/L initial [Ca2+], no rhizolith forms over the 500-year simulation run (i.e., equilibrium departure, Dԑ; Fig. 5a). Doubling the initial [Ca2+] to 0.3 mmol/L initiated the formation of a rhizolith at least 1 cm in radius in ca. 275 years (Fig. 5b). For initially high [Ca2+] ([Ca2+]o = 1 mmol/L), a rhizolith with a radius of 2 cm forms within 165 years under dry or semi-arid soil moisture conditions (Fig. 5c; Table 4), whereas the point-root model produced a 3-cm rhizolith in 150 years (Fig. 7), highlighting the role of localized water availability and intensified Ca2+ uptake associated with root-thinning. This demonstrates the dual role of [Ca2+]o and root geometry in enhancing Ca2+ exploitation and promoting rhizolith growth. In contrast, under optimal moisture conditions (h = −100 cm), the same rhizolith size (r = 3 cm) develops within 5 years, illustrating the limiting effect of hydraulic constraints on rhizolith formation under drier conditions (Fig. 5d).
Temporal evolution of rhizolith at 0.2-, 1-, 2-, and 3-cm radius around the root surface for 500-year equilibrium simulation run under arid conditions (ho = −1000 cm) except for d (ho = −100 cm). The equilibrium departure (in a) represents the limit of rhizolith formation by both low initial [Ca2+] and moisture/hydraulic constraints. The pre-saturation equilibrium (i.e., So, in ‘a’ and ‘b’) denotes a standstill in calcification function at 0.2-cm radius due to preferential redistribution/deposition of Ca2+ farther from 0.2-cm radius due to flux halt at 1-cm radius close to zero suction. The true saturation equilibrium (Sԑ, in ‘c’ and ‘d’) represents a standstill in the calcification function due to calcite saturation. In (e and f), the changes in CaCO3 are shown. See also Fig. 7 for point root and Supplementary Materials (S2 Figs. S2–S4) for the limiting factors for rhizolith formation simulated over 1- and 5.5-year runs.

Time (TRH) to the onset of rhizolith formation at a given radius (r) from root surface at slow water flow rates (ETo) in an arid profile (h = –1000 cm) with varying [Ca2+] in the soil solution. When “–”, rhizolith never formed at the given ecohydrological constraints. Water flow rates (fast or slow) have no influence on rhizolith formation rate under the relatively dry condition (h = –1000 cm). See also Figs. S1–S3.

Under both scenarios (Fig. 5c and 5d), the 0.2-cm proximity around the root was completely saturated with calcite at TCasat = 465 years. In both profiles, the saturation threshold ([Casat]z = 0.2) is observed to be 0.023 mol/L CaCO3. The agreement between the simulated saturation threshold ([Casat]z = 0.2 = 0.023 mol/L CaCO3) against the analytically calculated [Casat]z = 0.2 = 0.0245 mol/L CaCO3 (Table 3) is remarkable given such long simulation run. These distinct rhizolith layers exhibit increasing cementation from the outer to inner zones, reflecting calcite accumulation along water flux gradients during hydraulic redistribution (Figs. 3 and 5). This progressive cementation pattern highlights the capillary-driven nature of rhizolith formation and its role in pore occlusion over time. There was no tracked history of calcite dissolution after rhizolith formation (Fig. 5e and 5f).
Temporal evolution of rhizoliths and the time-capsule perspective
At initial time To, Ca2+ in the soil is driven along with water fluxes to the root. Ca2+ accumulation proceeds across time until Ca2+ concentration reaches 3 mmol/L, at which threshold (RhT) CaCO3 precipitation occurs and rhizoliths form at time TRh. Upon calcification, if drought stress does not constrain Ca2+ transport to the root, the CaCO3 even grows with time until the saturation threshold [Casat] for the soil pores at time TCasat is attained. After that, an inert ecosystem forms, where organic C (i.e., root remains), inorganic C, Ca, and O2 (as rhizolith), and soil minerals (e.g., carbonates and silicates) are conserved and transferred across a series of time t ≥ tRh, ti-1 until present, ti. Therefore, in our model, rhizoliths and their formation processes are perceived as a time capsule (TCap) along which CaCO3 masses are transferred across a series of time (Fig. 6). Root water uptake constrained at a single point reproduced similar spatio-temporal Ca2+ redistribution (Fig. 7) compared to that around the surface of the single-root scenario presented above.
Temporal evolution of rhizoliths under (semi-) arid stress and varying [Ca2+]o. Top panels (a–d) show rhizoliths (2 mm radius) forming under dual-moisture conditions, where optimal soil moisture (b, d) and high [Ca2+]o (c, d) influence calcification. The time capsule (TCap) concept is shown by branched arrows, representing CaCO3 transfer over time. Bottom panels (e–h) display a 500-year simulation of rhizoliths (10 mm radius). Panel (e) highlights equilibrium departure Dԑ under low [Ca2+]o and arid stress, while (f) shows a hydraulically constrained equilibrium. Panels (g and h) contrast rhizolith formation under (semi-) arid versus optimal soil moisture, with progressive calcification reflected in the y-scale expansion. Curved arrows in (h) emphasize time-dependent CaCO3 accumulation.

Summary of rhizolith formation model
Suction-induced fluxes towards the root redistribute Ca2+ around the root. When [Ca2+] reaches 3 mmol/L, calcification around the root surface occurs, and rhizolith formation begins. The time TRh required to form various radii of rhizolith differs across proximities to the root. Smaller radii/proximities closest to the root form first followed by wider proximities, thus resulting in layers of concentric rings around the root (Figs. 7 and 8 ).
Rhizolith formation profile summarizing concentric CaCO3 accumulation around the point root surface. Segments |a1a2|, |b1b2|, |c1c2|, and |d1d2| correspond to rhizolith diameters of 0.4 cm, 2 cm, 4 cm, and 6 cm, forming at ca. 1.5, 10, 100, and 150 years, respectively. The reference point is the root surface, with a 150-year simulation for visualization.

Conceptual framework for rhizolith formation modelling integrating hydraulic processes, redox equilibrium, and root water uptake using the HYDRUS-1D model. Hydraulic fluxes (green arrows) transport Ca2+ from initial time To to equilibrium time Tԑ, following a decreasing trend. As root water uptake  $v_t\left(\theta\right)$ depletes soil moisture
$v_t\left(\theta\right)$ depletes soil moisture 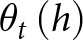 $\theta_t\left(h\right)$, the system reaches equilibrium water content (
$\theta_t\left(h\right)$, the system reaches equilibrium water content (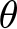 $\theta$ԑ) at matric potential head (hԑ), which defines the calcite saturation threshold
$\theta$ԑ) at matric potential head (hԑ), which defines the calcite saturation threshold 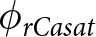 $\phi_{rCasat}$. Rhizolith formation occurs at varying radii (r1, r2, …, rn) when Ca2+ precipitates, illustrated by concentric rings representing effective pore volume. The interdependence of root water uptake and soil moisture is indicated by blue arrows, while curved gray arrows highlight calcite saturation at low pore volume. Upward/downward fluxes apply to point roots, whereas only upward fluxes occur in deep-root systems. Both scenarios exhibit similar hydraulic and Ca2+ redistribution patterns from the reference point to the root surface.
$\phi_{rCasat}$. Rhizolith formation occurs at varying radii (r1, r2, …, rn) when Ca2+ precipitates, illustrated by concentric rings representing effective pore volume. The interdependence of root water uptake and soil moisture is indicated by blue arrows, while curved gray arrows highlight calcite saturation at low pore volume. Upward/downward fluxes apply to point roots, whereas only upward fluxes occur in deep-root systems. Both scenarios exhibit similar hydraulic and Ca2+ redistribution patterns from the reference point to the root surface.

Under steady-state conditions, the finite water reservoir across the profile diminishes with time (To), water fluxes to the root decrease in response, and so does root water uptake. Equilibrium in the calcification function is attained at time Tԑ, at which point the matric potential heads are everywhere the same (hԑ) across the profile. At equilibrium, no net force exists because the suction head gradient dh/dz becomes zero, resulting in zero hydraulic fluxes and root water uptake. Therefore, the water content θԑ at Tԑ is infinitely occupied in the porous medium and defines the threshold pore volume for calcite saturation.
In our numerical experiment, equilibrium matric potential heads were attained at −8000 cm H2O, corresponding to 0.089 cm3/cm3 volumetric water content (θԑ). The porosity ɸ of the loess is 0.32 cm3/cm, therefore accounting for the equilibrium water content (θԑ = 0.089 cm3/cm3) results in 0.23 cm3/cm3 available for calcite saturation (i.e., ɸCasat), which is 72% of the porosity of loess (i.e., ɸCasat = 0.72ɸ). The equilibrium water content (θԑ) has not yet reached the residual water content (θr) listed in Table 1 due to its dependency on the selected Feddes parameters and the semi-arid reference evapotranspiration values, including surface evaporation and year-round transpiration dynamics (Szutu and Papuga, Reference Szutu and Papuga2019). For the purpose of simulating rhizolith formation, the model’s approach to hydraulic redistribution through root water uptake is simplified in Fig. 8 .
Discussion
Ecohydrological constraints for rhizolith formation
Pedogenic CaCO3 precipitation from a soil solution at thermodynamic equilibrium can be driven by an increase in [Ca2+] in soil solution under several biogeochemical constraints (Monger et al., Reference Monger, Cole, Buck and Gallegos2009). However, physical processes such as hydraulic redistribution and soil water content alone are sufficient (and certainly quintessential) for driving pedogenic carbonate accumulation (McFadden and Tinsley, Reference McFadden and Tinsley1985; Košir, Reference Košir2004). Alongside plant phenology, root water uptake is influenced by soil moisture (Bouma, Reference Bouma2018) and the physical processes dominating soil-water movement (Richards and Caldwell, Reference Richards and Caldwell1987). The investigation into these factors influencing rhizolith formation in semi-arid environments yielded valuable insights into the interplay between water availability (Fig. 2c and 2d), potential evapotranspiration rates (Fig. 4e and 4f; Table 4), and initial [Ca2+] (Fig. 5; see also Table 5). Thus, limited soil water availability (h = −1000 cm) under (semi) arid climate (Sanchez‐Mejia and Papuga, Reference Sanchez-Mejia and Papuga2014; Sardans et al., Reference Sardans, Peñuelas and Estiarte2006) constrains root water uptake and impedes Ca2+ fluxes toward the roots (Corwin et al., Reference Corwin, Rhoades and Šimůnek2007; Šimůnek et al., Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009a, Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009b).
Time (TRH) to the onset of rhizolith formation at a given radius (r) from root surface at fast and slow water flow rates (ETo) in an optimally moist profile (h = –100 cm) with varying [Ca2+] in the soil solution. When “–”, rhizolith never formed. See also Figs. S1–S3.

This limitation explains the inability of Ca2+ fluxes to exceed the threshold necessary for rhizolith formation within the 5.5-year simulation period, even with an initial 0.3 mmol/L Ca2+ (Drever, Reference Drever1982). When water is limiting, the calcification curve remains insensitive to increases in water flow rate under arid conditions. However, under optimally moist conditions (h = −100 cm), increasing the water flow rate considerably accelerates calcification leading to rhizolith formation (Table 5). This observation underscores the critical role of soil moisture content in regulating rhizolith formation (Alonso-Zarza et al., Reference Alonso-Zarza, Genise, Cabrera, Mangas, Martín-Pérez, Valdeolmillos and Dorado-Valiño2008; Sun et al., Reference Sun, Zamanian, Huguet, Bayat, Wang and Badawy2022), providing important insights into pedogenic processes in ‘calcium-rich’ soils (Rodríguez-Aranda and Calvo, Reference Rodríguez-Aranda and Calvo1998). In line with our findings, the prerequisite for calcification is the amount of Ca2+ in the soil solution, therefore, even if soil moisture does not constrain root water uptake, Ca2+ accumulation around the root and rate of calcification are determined by the initial [Ca2+] in the profile.
The development and evolution of rhizoliths depending on soil moisture and [Ca2+] scenarios shed light on the complex dynamics governing rhizolith formation. From our simulations, rhizolith formation is influenced by [Ca2+]o and ETo, as well as water content, which limits the process. In dryland ecosystems, drought stresses are the principal ecohydrological constraints (IPCC, Reference Houghton, Ding, Griggs, Noguer, van der Linden, Dai, Maskell and Johnson2001; Sardans et al., Reference Sardans, Peñuelas and Estiarte2006) that influence root water uptake (Bouma Reference Bouma2018), and transpiration (Kurc and Small, Reference Kurc and Small2007; Cavanaugh et al., Reference Cavanaugh, Gosnell, Davis, Ahumada, Boundja, Clark and Mugerwa2014; Bouma, Reference Bouma2018). In such conditions, Ca2+ fluxes to the root do not yield sufficient Ca2+ accumulation for rhizolith formation, even under presumably high [Ca2+]o (0.3 mmol/L) at the end of the 5.5-year simulation run. However, an optimally moist profile (h = −100 cm) following, for instance, a few days after a large storm (Cavanaugh et al., Reference Cavanaugh, Kurc and Scott2011), favors the rhizolith formation process.
Rhizolith formation as a mechanism in Earth system processes and deep-time C sink
Rhizolith formation is intrinsically linked to the soil-C cycle through biogenic CO2 consumption, DIC production, carbonate precipitation, and root-C occlusion. In the rhizosphere, elevated pCO2 from root and microbial respiration enhances CO2 hydrolysis, forming carbonic acid (Lambers et al., Reference Lambers, Mougel, Jaillard and Hinsinger2009; Zamanian et al., Reference Zamanian, Pustovoytov and Kuzyakov2016; Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025; Equation 1). At equilibrium, carbonic acid dissociates into bicarbonate and protons (Equation 2), and as Ca2+ accumulates via root water uptake, bicarbonate reacts with Ca2+ to precipitate CaCO3 (Equation 3) immobilizing C as pedogenic CaCO3 while simultaneously occluding root C, effectively mitigating atmospheric CO2 concentrations over geological timescales. By stabilizing belowground-C, rhizolith formation contributes to long-term C storage and mitigates atmospheric CO2 concentrations (Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021; Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025). These findings provide critical insights into the role of pedogenic carbonate formation in global carbon cycling and its sensitivity to climate variability.
Climatic aridity and plant lifespan shape rhizolith formation in long-term equilibrium simulations
Alongside drought stress, which limits Ca2+ fluxes to the root at a certain root water requirement, time emerges as an important factor for rhizolith formation. Considering a semi-arid profile containing 0.3 mmol/L Ca2+ in the soil solution, rhizoliths never formed over a 5.5-year simulation run, whereas extending simulation runs resulted in the formation of rhizoliths of radius 1 cm in 275 years (Fig. 5b). Again, using a similar moisture profile with 1 mmol/L Ca2+, a rhizolith of radius 0.2 cm forms in ca. 1.5 years and it takes ca. 165 years to form a bigger rhizolith of 2-cm radius (Fig. 5c), while under a root-thinning scenario, the same rhizolith size develops in ca. 150 years (Fig. 7).
The implication is that plant lifespan may influence rhizolith size, with short-lived plants, such as Artemisia sphaerocephala, forming smaller rhizoliths compared to longer-lived species, such as Larrea tridentata (creosote bush). However, ecohydrological constraints, such as root water uptake rate or local soil and climatic conditions, are the limiting factors influencing rhizolith onset and size (Fig. 5; see also Fig. S1). Another implication is that root water uptake may accumulate Ca2+ (in amounts) just below the rhizolith formation threshold, potentially inducing post-mortem rhizolith formation from an addendum of Ca2+ topped up by the cytosol of dead roots, microorganisms, and soil minerals (Cramer and Hawkins, Reference Cramer and Hawkins2009; Monger et al., Reference Monger, Cole, Buck and Gallegos2009; Sun et al., Reference Sun, Zamanian, Huguet, Fa and Wang2020). Importantly, during redistribution, the movement of water is towards the drier region as a result of adsorption and capillary attraction (Cornelis et al., Reference Cornelis, Gabriels and Hartman2004). Similarly, in our model, as long as there is a hydraulic head gradient between the root zone and the surrounding soil, Ca2+ is driven toward the root along with water. The implication is that once rhizolith of any size is formed, the matric head becomes more negative around the root than the surroundings, inducing redistribution and/or internal drainage towards the former. Therefore, Ca2+ continues to accumulate along with water fluxes, consequently forming a thicker rhizolith whether the root is active or inactive until the pores are jammed by calcite saturation. Even at 0.2 cm radius, Ca2+ saturation occurs in the long run (TCasatz = 0.2 cm = 465 years) at 0.023 mol/L CaCO3, implying that for wider radii, Ca2+ will continually fill the pores over a much longer time until saturation.
In our model, as matric potential head develops and approaches −8000 cm, the hydraulic capacity at proximities close to the root equilibrates with that of the root (Fig. 2e and 2f). This phenomenon presents enormous constraints on water fluxes (Feddes and Zaradny, Reference Feddes, Zaradny, Penning de Vries and van Laar1978), which drive Ca2+ redistribution and pedogenic calcification. For instance, as the hydraulic capacity at 1 cm proximities to the root equilibrates with that at the root, Ca2+ fluxes halt at 1 cm without reaching the 0.2 cm radius. This can be observed in the so-called pre-saturation equilibrium (Fig. 5a and 5b) where more Ca2+ deposition preferentially occurs at 1 cm around the root than at 0.2 cm proximities. The reason is simply because fluxes driven towards the root upon reaching 1 cm proximity, z1, must be redistributed between the root and z1. However, because the hydraulic capacities are almost in equilibrium across the two points, fluxes are left behind at 1-cm proximity while negligibly small amounts reached 0.2-cm proximity. This preferential Ca2+ deposition at 1-cm proximity becomes evident (Fig. 5b) where rhizolith formed a 1-cm radius in ca. 275 years while a 0.2-cm radius still lags below the rhizolith formation threshold. Under normal conditions of distinctive suction gradient, accumulated Ca2+ at a radius of 0.2 cm should suffice to initiate rhizolith formation prior to any formation at 1 cm. However, from a 100-year simulation run onwards, flux partitioning between the root and 1-cm proximities becomes irrelevant as suction head gradient across them approaches zero (Feddes and Zaradny, Reference Feddes, Zaradny, Penning de Vries and van Laar1978). In this study, we refer to this state as a ‘pre-saturation equilibrium,’ indicating that Ca2+redistribution ceases even though the maximum porosity available for calcification (ɸCasat = 0.23 cm3/cm3) has not yet been exceeded; the standstill results solely from hydraulic constraints (Hillel, Reference Hillel2003; Šimůnek et al., Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009a, 2009b). True saturation equilibrium was attained at 0.023 mol/L calcite (Fig. 5c and 5d).
These modeled equilibrium outcomes align with Quaternary deposits, where rhizoliths exhibit cementation patterns reflecting variations in soil chemistry, moisture regimes, and plant physiology. The prevalence of pedogenic carbonate cementation in semi-arid environments suggests that calcite precipitation around roots follows similar water redistribution patterns as observed in paleosol systems. Cementation structures vary radially, with an outer zone (2–3 cm radius) showing weak cementation, dispersed calcite patches occupying ∼13.9% of the loess pore volume, and evidence of intermittent Ca2+ deposition (Fig. 5). The intermediate zone (1–2 cm radius) exhibits moderate cementation, where continuous Ca2+ deposition follows water fluxes toward the root, resulting in calcite filling ∼36.2% of the loess pore volume. The inner zone (0.2–1 cm radius) is strongly cemented, reaching calcite saturation at 0.023 mol/L CaCO3 after 465 years, occupying ∼72.2% of loess porosity, where pore jamming restricts water permeability, hydraulic conductivity, and aeration (Fig 3).
These cementation gradients are characteristic of rhizolith formation in loess deposits, where pedogenic carbonate cementation is dominant (Kraus and Hasiotis, Reference Kraus and Hasiotis2006; Li et al., Reference Li, Wang, Cheng, Ning, Zhao and Li2015a). The degree of cementation influences the structural integrity (Fig. 5e and 5f) and internal architecture of rhizolith ichnofacies after root mortality (see also Fig. 9). However, in loess–paleosol sequences, alternative cementation processes such as silicification, ferruginization, gypsification, and organic cementation also contribute to rhizolith development (Kraus and Hasiotis, Reference Kraus and Hasiotis2006). The modeled rhizolith sizes, ages, and morphologies correspond to field observations, where rhizoliths range from less than 1 mm to 30 cm in diameter (Löscher and Haag, Reference Löscher and Haag1989; Alonso-Zarza et al., Reference Alonso-Zarza, Genise, Cabrera, Mangas, Martín-Pérez, Valdeolmillos and Dorado-Valiño2008; Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021).
Conceptualization of (i) root encasement types (a, b, and c) during rhizolith formation, (ii) fate of encapsulated root types, and (iii) the concomitant rhizolith transformations (in types b and c) enhanced by diagenesis, hydrological, and biogeochemical processes. Dark horizontal arrows indicate the processes driving rhizolith transformation. In type (a) (ii), the brown root represents remnants due to physical occlusion. In types (b and c) (ii), central voids (white) form following root decay. Blue arrows indicate the direction of diagenesis and macropore transport into these voids. The concretionary ring shows increasing calcification from the outer edge (light brown) toward the center (shades of gray; a, b, and c) and the reformed internal structure (dark blue) in types (b and c) (iii).

The root pressure effect and its implications on rhizolith formation
The root pressure effect plays a pivotal role in understanding the processes underlying root water uptake and its implications for rhizolith formation, particularly in arid environments where soil-water availability is limited. Vascular bundles within the root system are composed of xylem vessels, which facilitate water transport and maintain hydrostatic equilibrium with the soil matrix, allowing water influx via osmosis and active transport (Zimmermann, Reference Zimmermann1978; Tyree and Ewers, Reference Tyree and Ewers1991; Jackson et al., Reference Jackson, Sperry and Dawson2000). The phenomenon of root pressure (Fig. 3; see also Fig. S4) illustrates how the matric potential head (hRoot) at the root vicinity (taken at 0.2 cm proximity) initially exceeds the equilibrium matric potential head (hԑ). This mechanism is crucial for preventing fatal physiological stress, such as embolism resulting from hydraulic failure (blockage of the xylem vessels) induced by the ingress/build-up of air bubbles into the root (Tyree and Ewers, Reference Tyree and Ewers1991). The influence of hRoot per unit area of soil is more pronounced for thicker than for fine roots, indicating a greater potential for closer redistribution of ions including Ca2+ around the latter (Jackson et al., Reference Jackson, Canadell, Ehleringer, Mooney, Sala and Schulze1996; McCully, Reference McCully1999) whereas the former invests in water selectivity to quench its water affinity, maintain hydration, and overcome osmotic/salinity stress (Cochard and Tyree, Reference Cochard and Tyree1990).
In arid conditions, where precipitation surplus is scarce and soil salinity is a concern due to lack of leaching, selective water uptake by semi-permeable thicker taproots serves as an adaptation without succumbing to osmotic and salinity stress (van den Honert, Reference van den Honert1948; Carlquist, Reference Carlquist1975). This is evident in species such as Welwitschia mirabilis, which can survive for millennia without evidence of rhizolith formation, due to its thick taproot adapted for selective water exploitation and preservation. In contrast, fine roots can navigate through fissures, intricate soil pore networks, and cracks, tapping otherwise inaccessible moisture reservoirs and driving pedogenic calcification by transporting Ca2+ along with water. This concept partly explains why the fine roots of species with even relatively short lifespans, such as Artemisia sphaerocephala and Larrea tridentata, have been studied to reflect rhizolith formation (Cleveland et al., Reference Cleveland, Hildebrand, MacLean and Hargrave2015; Sun et al., Reference Sun, Zamanian, Huguet, Fa and Wang2020). Thus, the interplay among root size, water uptake mechanisms, and soil conditions influences the formation and characteristics of rhizoliths in various plant species within (semi-) arid environments.
Rhizoliths as paleoindicators of paleosol hydrology and redox dynamics
Rhizoliths, formed at varying diameters, undergo diagenetic transformations (Sun et al., Reference Sun, Zamanian, Huguet, Bayat, Wang and Badawy2022, Reference Sun, Zamanian, Huguet, Wiesenberg, Zhao and Lei2023), contingent upon the degree of concretionary encasement surrounding the root. Three primary scenarios emerge (Fig. 9): full encasement, tip encasement with exposed collar, and length encasement with exposed tip and collar.
Full encasement (type a) entails homogeneous Ca2+ accumulation along the entire root length (Fig. 9a), resulting in tubular rhizolith formation encompassing the entirety of the root from the tip to the collar. Because the suction head gradient driving Ca2+ fluxes varies across elevations in the profile (Darcy, Reference Darcy1856; Šimůnek et al., Reference Šimůnek, Jacques, Twarakavi and van Genuchten2009a), full encasement rarely occurs. However, when it does occur, the longevity of the buried root activity hinges on factors such as concretion degree of available pore volume for aeration and access to soil water and nutrients (Jim, Reference Jim1993; McLaren, Reference McLaren1995). Following root death, microbial access from the surrounding soil is obstructed due to the physical occlusion enhanced by the concretion, leading to indefinite root remnants within an ‘inert’ concretion.
In tip encasement with exposed collar (type b), the root tip is encapsulated within a concretionary mass, while the collar remains exposed to the surrounding sediment (Fig. 9b). This configuration creates a unidirectional hotspot for microbial access, facilitating decomposition of the root material within the concretionary mass through the exposed collar. As a result, organic substances (e.g., acids) and ions including Ca2+ are released, leading to the formation of a fossilized root channel and the creation of a one-way void/macropore within the concretion. On the other hand, the closed or concreted bottom of the encasement potentially leads to water stagnation creating a reducing microenvironment. We emphasize that the degree and duration of the reducing microenvironment are influenced by amount and composition of transported sediments into the central void, and the availability of alternative electron acceptors for microbial respiration. Weathering of minerals such as CaCO3 and silicates is therefore accelerated and induces the formation of secondary carbonates and the release of Ca2+. Reactions between the secondary carbonates and the released species (such as Ca2+ from root decomposition and calcite weathering) further facilitate the reprecipitation/recrystallization of CaCO3.
For length encasement with exposed tip and collar (type c), because bilateral microbial access to the partially encased root fosters extensive microbial activity (Fig. 9c) the preserved root remains are generally absent (Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011; Sun et al., Reference Sun, Wang and Zamanian2019a). This leads to the formation of organic acids and the creation of two-way voids within the concretionary mass. These voids serve as macropores, facilitating preferential pathways for hydrological and diagenetic processes. Type c in contrast to type b fosters aerobic conditions facilitated by the contact soil acting as a filter at the bottom, allowing for water passage while soil particles and colloids refill the void. Hydrological processes drive organic and inorganic substances into the void within the concretionary mass via colloid-facilitated transport and chemical dissolution fluids. However, the clastic clay and silt particles can be driven into the central voids by the forces of the diagenesis process, or the mechanical shift of soil and/or sediments. Negative surfaces of mobilized organic substances and mineral soil particles such as clays act as carriers for Ca2+ and hydration by water (e.g., Brady, Reference Brady1984). Consequently, these voids serve as reservoirs for water and soil particles including Ca2+-bearing colloids.
Therefore, the resulting internal structures of rhizoliths, formed under encasement types a, b, and c, display a spectrum of complexity and heterogeneity. This heterogeneity echoes the dynamic interplay among roots, minerals, and microorganisms over time. Importantly, the refilling of void types b and c is not merely a physical phenomenon but also a biogeochemical process (Klappa, Reference Klappa1980; Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011, Reference Gocke, Huguet, Derenne, Kolb, Dippold and Wiesenberg2017). Microbial communities, mineral deposits, and Ca2+ transported by colloids collectively contribute to calcification within the void (Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025), shaping the internal structure of rhizoliths through a multifaceted interplay of hydrological, physical, microbial, and biogeochemical processes (Fig. 9). We emphasize that these processes unfold gradually in arid environments, often triggered by infrequent heavy storms capable of inducing macropore flow.
The reformed internal structure of rhizoliths holds important implications for paleoenvironmental reconstructions. The process of organic acid attack on carbonate minerals results in the alteration or removal of primary carbonate structures (Schimel et al., Reference Schimel, Balser and Wallenstein2007; Schimel and Schaeffer, Reference Schimel and Schaeffer2012). This alteration leads to reduction in CaCO3 percentage towards the biogeochemical hotspot, reshaping the morphological features and isotopic signatures of the rhizolith. Additionally, the release of ions, particularly Ca2+ from decomposed roots, may play a role in the formation of secondary carbonate minerals. These secondary carbonates can contribute to modern/younger 14C values (from coprecipitated CO2 released from the organic acids following root decomposition) within the reformed internal structure compared to the older concretionary edge (Kuzyakov et al., Reference Kuzyakov, Schevtzova and Pustovoytov2006; Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025). The contrasting hydrological regimes governing encasement types a, b, and c highlight the potential of rhizoliths as paleoindicators of ancient drainage conditions and redox states, offering insights into moisture regimes and biogeochemical cycling in paleosols (Zwertvaegher et al., Reference Zwertvaegher, Finke, De Reu, Vandenbohede, Lebbe, Bats and De Clercq2013; Li et al., Reference Li, Wang, Li, Ning, Cheng and Zhao2015b). Well-preserved rhizoliths, characterized by a lack of fluid exchange, reflect well-drained paleosol conditions with minimal redox fluctuations, where prolonged oxic conditions inhibit Fe and Mn mobilization and limit pedogenic gleying. In contrast, oxidized halo rims in types b and c may indicate paleosols with fluctuating drainage, where periodic water saturation drives alternating redox cycles, Fe mobilization, and partial gleying (Kraus and Hasiotis, Reference Kraus and Hasiotis2006). Diffuse gray rhizohaloes within a concretionary mass reflect groundwater gleying in poorly drained paleosols where persistent water saturation promotes Fe reduction.
Rhizolith as a ‘time capsule’ and pathway for multi-layer paleoclimate proxy development
The model conceptualizes the process of rhizolith formation as a time capsule (TCap) in which root systems, soil minerals, and CaCO3 masses are preserved across multiple temporal scales (Figs. 5). This supports the concept of ‘mass flow’ of Ca2+ driven towards the root (Alonso-Zarza, Reference Alonso-Zarza2018; Luo et al., Reference Luo, Wang, An, Zhang and Liu2020) along with water fluxes during transpiration (Caldwell et al., Reference Caldwell, Dawson and Richards1998; Cardon and Gage, Reference Cardon and Gage2006). The transportation of CaCO3 occurs over successive time windows, resulting in the formation of concentric rhizoliths with different diameters at various stages: a 0.2-cm radius rhizolith forms in ca. 1.5 years, followed by a 1-cm radius rhizolith in ca. 9.5 years, and eventually a ∼2- to 3-cm radius rhizolith in ca. 150–165 years (Figs. 5c and 7). This temporal evolution implies that rhizoliths comprise layers of calcareous species (Ca2+, CO2) transported across various time periods (Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021). These calcified layers, composed of an organo-geochemical matrix of root remnants, CaCO3, and mineral particles, serve as chronological records of paleoenvironmental conditions. Each successive layer forms under unique hydrological, geochemical, and biological setting, effectively capturing variations in soil water , atmospheric CO2, and biotic activity (Retallack, Reference Retallack1990; Mattucci et al., Reference Matteucci, Belluomini and Manfra2007; Gao et al., Reference Gao, Li, Wang and Li2019, Reference Gao, Li, Zhu and Wang2020). Alongside calcification, the structural integrity of rhizoliths can be enhanced by Si for indefinite inertness (Gocke et al., Reference Gocke, Kuzyakov and Wiesenberg2010).
Thus, rhizoliths are not only pedogenic strcutures but time capsules of organo-geochemical memory storing paleoclimate proxies (Figs. 5 and 10). These ‘inert organo-geochemical materials’ are composed of distinct submatrices, each originating from specific time frames (To, Ti−1, Ti, as past, recent, and present respectively; Fig. 6). Rhizolith layers, hence, are witnesses of evolving ecosystems, develop independently over time, and interact at various points before converging in the present. In essence, the present (Ti) serves as the common time point, where the interactions of these evolving layers intersect, encapsulating a comprehensive record of recent and past environmental conditions and soil dynamics (Retallack, Reference Retallack1990). Rhizolith growth is governed by soil moisture, evapotranspiration, and root activity (Tables 4 and 5), which respond dynamically to precipitation, temperature, and CO2 levels. In arid climates, higher evapotranspiration rates concentrate Ca2+, accelerating CaCO3 precipitation, whereas humid conditions promote leaching, slowing rhizolith formation. By analyzing their size, distribution, and isotopic composition, researchers can infer pedogenic duration, climatic intensity, and ecosystem evolution crucial for understanding the evolution of ancient ecosystems and for reconstructing paleoclimatic scenarios (Li t al., Reference Li, Gao and Han2017).
Schematic representation of rhizolith formation as a mass-flow concentration mechanism. Geochemical and hydraulic processes channel Ca2+ towards root surfaces via water uptake, where supersaturation triggers CaCO₃ precipitaton with rhizospheric-derived dissolved inorganic carbon. This produces concentric mineral bands that form successive growth rings (r1, r2, r3) with distinct CaCO3 gradients. These rings archive multiple proxies including root carbon, biomarkers, stable isotopes ((δ13C, δ18O, δ44Ca), and trace elements, preserved across successive temporal frames from the past (To) trough the present (Ti). Rhizoliths thus operate as “time capsules” of paleoenvironmental information, recording soil CO2 dynamics, temperature, precipitation, and vegetation histories

Over decades to centuries, rhizoliths accumulate calcified layers that chronicle shifts in soil water and pCO2 dynamics, root activity, and carbonate precipitation (Fig. 7). These timescales directly link rhizoliths to paleosol development. Thicker rhizoliths (2–3 cm), formed within decades (Fig. 5d; Table 5), may indicate prolonged periods of soil water availability and enhanced biological activity, whereas thinner or discontinuous rhizoliths suggest episodic moisture or unstable pedogenic conditions (Fig. 5a–c; Table 4).
As tangible time capsules of paleoenvironmental change, rhizoliths serve as valuable proxies for reconstructing past climates. Their geochemical and isotopic signatures (e.g., δ13C, δ1⁸O, δ⁴⁴Ca) of the modeled environmental variables (Equation 3) and elemental profiles (e.g., Sr, Ba) refine paleoclimatic interpretations (Pustovoytov and Terhorst, Reference Pustovoytov and Terhorst2004; Agnelli, Reference Agnelli, Ascher, Corti, Ceccherini, Pietramellara and Nannipieri2007; Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021; Liu et al., Reference Liu, Li, Dong, Deng, Liu and Gao2024). Shifts in δ13C values indicate transitions between C3- and C4-dominated vegetation, while variations in δ18O track past precipitation and evapotranspiration trends (Marion, Reference Marion1989; Liutkus et al., Reference Liutkus, Wright, Ashley and Sikes2005; Kelson et al., Reference Kelson, Huth, Andrews, Bartleson, Cerling, Jin, Salinas and Levin2025). Decadal climate fluctuations during the Late Pleistocene can thus be reconstructed using CaCO3-rhizolith records (Wang and Greenberg, Reference Wang and Greenberg2007; Li et al., Reference Li, Wang, Cheng and Li2016).
Our concentric CaCO3 accumulation model (RhizoCalc) highlights isotopic stratigraphy within rhizolith layers, encoding long-term environmental changes (Fig. 10). These signals in loess–paleosol sequences support chrono-sequential reconstructions of late Quaternary paleoenvironments, where root-driven Ca2+ fluxes control calcification (Agnelli, Reference Agnelli, Ascher, Corti, Ceccherini, Pietramellara and Nannipieri2007; Gocke et al., Reference Gocke, Pustovoytov, Kühn, Wiesenberg, Löscher and Kuzyakov2011; Zamanian et al., Reference Zamanian, Lechler, Schauer, Kuzyakov and Huntington2021; Liu et al., Reference Liu, Li, Dong, Deng, Liu and Gao2024). To enhance paleoclimatic interpretations, cross-sectional rhizolith analysis should focus on isolating concentric TCap layers to decode discrete isotopic bands, improving insights into soil–plant interactions, root growth patterns, and nutrient cycling in ancient landscapes (Retallack, Reference Retallack1990; Morris et al., Reference Morris, Leake, Stein, Berry, Marshall, Wellman and Milton2015; Liu et al., Reference Liu, Li, Dong, Deng, Liu and Gao2024).
Limitations and applicability of the dynamic RhizoCalc model
While the RhizoCalc model provides insights into rhizolith formation mechanisms in loess soils, its applicability to other environments is dynamic. The model was calibrated for silty loam loess soils, and its transferability to coarser soils, such as sand dunes, may be constrained due to differences in hydraulic properties and water retention. The assumptions of a homogenous root distribution and simplified water uptake do not account for variability in root architecture or species-specific differences. However most (desert) plants in (semi-) arid regions are adapted to the equilibrium pressure heads (−8000 cm H2O) considered in the model. To isolate rhizolith formation constraints, simulations held key environmental conditions (soil temperature, moisture, pCO2, [Ca2+]) constant, omitting diurnal and seasonal fluctuations. Applying the model to sandy or clay-rich soils would require recalibration of hydraulic and geochemical parameters to reflect distinct water and solute transport processes. Key model parameters including dual moisture profiles, initial [Ca2+], evapotranspiration rates, and pCO2, remain robust for (semi-) arid conditions. In this way RhizoCalc provides a transferable framework for understanding the fundamental dyanmics of rhizolith ‘time capsule’ development, while retaining flexibility for broader pedogenic and paleoenvironmental applications when appropriately modified.
Conclusions and outlook
The RhizoCalc model provides important insights into the steps and factors influencing mass flow towards the root, root calcification, and rhizolith development in ‘calcium-rich’ soils. By elucidating the roles of soil water, initial [Ca2+], and potential evapotranspiration rates, this work supports the postulate of ‘mass flow concentrating’ as the fundamental/leading mechanism of rhizolith formation.
Firstly, the root water uptake model, based on the hydraulic flux principle, underscores the importance of soil moisture in driving Ca2+ fluxes toward the root. As root water uptake progresses over time, the finite water reservoir diminishes, leading to a decline in hydraulic capacity and eventually reaching a zero-suction gradient in 500 years. This phenomenon has important implications for understanding the constraints on root water uptake under varying soil moisture conditions and the subsequent effects on plant physiology and ecosystem dynamics.
This investigation into the processes of rhizolith formation elucidates the complex interplay among soil-water availability, initial [Ca2+], and potential evapotranspiration rates. We demonstrate that these factors play critical roles in determining the rate and extent of rhizolith formation over time scales from years to decades (up to 1–2 centuries). Particularly under arid conditions, limited soil-water availability constrains Ca2+ fluxes towards the roots, impeding the accumulation necessary for rhizolith formation. This underscores the importance of considering environmental variables in predicting rhizolith formation patterns and understanding pedogenic processes in calcium-rich soils.
Our simulations highlight the remarkable sensitivity of rhizolith formation to environmental shifts and plant lifespan. Through extensive long-term modeling, we unravel the intricate temporal dynamics of rhizolith development, revealing its potential to span years, decades, and even centuries. These processes culminate in the creation of rhizoliths of increasing radii, ranging from 0.2 cm to 3 cm, as they interact and evolve over time until reaching a convergence at certain intervals, thereafter, transferring through time to the present.
This model conceptualizes rhizoliths as time capsules, encapsulating essential environmental indicators, including C, O, and Ca isotopes, as well as organic compounds from root remnants, and Si isotopes from soil minerals. Each concentric ring within a rhizolith captures a snapshot of recent and past environmental conditions and ecosystem dynamics, providing invaluable insights into soil history across distinct time intervals. Furthermore, our study underscores the dynamic nature of rhizoliths over time, a process influenced by the degree of concretionary encasement surrounding the root. These transformations are driven by a combination of biogeochemical and energetic physical processes, including diagenesis and macropore-/colloid-facilitated transport. As a result, rhizoliths develop diverse internal structures with implications for paleoenvironmental reconstructions.
Lastly, investigation into the root pressure effect unveils the role of root size dynamics on water uptake mechanisms in arid environments and their implications for rhizolith formation. The large surface area to volume ratio of fine roots permits them to navigate through fissures, intricate soil pore networks, and cracks, extensively exploiting otherwise inaccessible moisture reservoirs and driving pedogenic calcification by transporting Ca2+ along with water.
Root water uptake is therefore dynamic and depends on plant phenology and physiology, moisture dynamics influenced by rainfall, surface evaporation, and capillary rise (from a phreatic zone). These dynamics present enormous constraints for the biophysical mechanisms highlighted for rhizolith formation. For instance, fluctuations in rainfall events might cause fluctuations in moisture content and pCO2, hence affecting the calcification function. These factors may slow down the driving mechanisms and present uncertainties/variability at the field scale. Despite these complexities, the RhizoCalc model offers a robust mechanistic framework for understanding rhizolith formation under diverse environmental conditions.
Supplementary material
The supplementary material for this article can be found at https://doi.org/10.1017/qua.2025.10021.
Acknowledgments
The authors would like to thank the Chinese-German Centre (Mobility project M0069), the German Research Foundation (DFG ZA1068/4), the Ministry of Science and Culture of Lower-Saxony, Germany (15-76251-2—Stay-8/22- 5947/2022), and the RUDN University Strategic Academic Leadership Program for their support.






















 Open access
Open access