


Introduction
Industrialized farming systems such as intensive monoculture and reduced tillage systems have encouraged a heavy reliance on herbicides with the same site of action (SOA) for weed management. This reliance on herbicides is particularly evident in the Republic of South Africa, where corn (Zea mays L.) is one of the most important grain crops produced in all nine provinces (Fanadzo et al. Reference Fanadzo, Chiduza and Mnkeni2009; Masehela et al. Reference Masehela, Rhodes, Groenewald, Poole, Van den Berg, Gouse, Skowno, Barros, Seymour and Mandivenyi2021). Approximately 85% to 95% of the maize grown in South Africa is genetically modified to have tolerance to glyphosate, resistance to certain insects, or both (Masehela et al. Reference Masehela, Rhodes, Groenewald, Poole, Van den Berg, Gouse, Skowno, Barros, Seymour and Mandivenyi2021). Globally, a clear link exists between the adoption of glyphosate-tolerant crops such as maize and soybeans [Glycine max (L.) Merr.] and the accelerated evolution of glyphosate-resistant (GR) weeds (Kniss Reference Kniss2018; Powles Reference Powles2008).
The evolution of herbicide resistance is driven primarily by the reproductive biology and genetic variability of weeds (Torra et al. Reference Torra, Montull, Calha, Osuna, Portugal and De Prado2022). In South Africa, nine different weed species have evolved resistance to multiple SOAs. Among the Amaranthus species present in South Africa, Palmer amaranth (Amaranthus palmeri S. Watson) was first reported to have evolved resistance to inhibitors of acetolactate synthase (Group 2 herbicides as categorized by the Herbicide Resistance Action Committee [HRAC and Weed Science Society of America [WSSA]) and 5-enolpyruvlshikimate-3-phosphate synthase (Group 9) by Reinhardt et al. (Reference Reinhardt, Vorster, Küpper, Peter, Simelane, Friis, Magson and Aradhya2022), whereas smooth pigweed populations have been reported to have evolved resistance to atrazine (Group 5) (Heap Reference Heap2025; Sereda et al. Reference Sereda, Erasmus and Coetzer1996).
The increase in biotypes demonstrating resistance to multiple SOAs increases the selection pressure on the remaining effective SOAs, especially in the absence of integrated weed management strategies that include nonchemical weed control methods (Braz and Takano Reference Braz and Takano2022). Nevertheless, some management practices have been employed to reduce selection pressure and help manage the current resistance issues in Amaranthus species. Incorporating preemergence herbicides is essential for controlling Amaranthus species and ensuring an effective and sustainable management program (Braz and Takano Reference Braz and Takano2022). Weeds are more vulnerable to herbicides during the germination and emergence stages and are less likely to express target-site resistance and metabolic resistance during this stage (Kaundun et al. Reference Kaundun, Hutchings, Dale, Howell, Morris, Kramer, Shivrain and Mcindoe2017; Vennapusa et al. Reference Vennapusa, Faleco, Vieira, Samuelson, Kruger, Werle and Jugulam2018; Wuerffel et al. Reference Wuerffel, Young, Matthews and Young2015). Postemergence herbicides should be applied early (i.e., when weeds are at the 2- to 4-leaf stage) for optimum control. Late application of herbicides (>8-leaf stage) typically requires herbicide combinations or sequential applications (Braz and Takano Reference Braz and Takano2022; Kaundun Reference Kaundun2021).
Recently, glyphosate-resistant (GR) smooth pigweed populations in the Bergville and Winterton districts of KwaZulu-Natal Province (KZN) were confirmed to possess a triple mutation (TAP-IVS) in the EPSPS gene (Shabangu et al. Reference Shabangu, Simelane, Reinhardt, Marais and Vorster2024). This target-site mutation confers high levels of glyphosate resistance (García et al. Reference García, Palma-Bautista, Rojano-Delgado, Bracamonte, Portugal, Alcántara-de la Cruz and De Prado2019; Perotti et al. Reference Perotti, Larran, Palmieri, Martinatto, Alvarez, Tuesca and Permingeat2019; Shabangu et al. Reference Shabangu, Simelane, Reinhardt, Marais and Vorster2024; Sulzbach et al. Reference Sulzbach, Turra, Cutti, Kroth, Tranel, Merotto and Markus2024). This study aimed to evaluate the effectiveness of preemergence and postemergence herbicide products with different SOAs as alternative options for controlling GR smooth pigweed populations in maize and soybeans in South Africa. We hypothesized that herbicide resistance would vary based on herbicide rate and population (i.e., location) for the herbicide products that represent different SOAs. Therefore, the response of smooth pigweed populations to various herbicide rates and their interaction (herbicide rates × populations) was evaluated to determine differential susceptibility to the tested herbicide products.
Materials and Methods
Seed Source and Experimental Site
Seeds of GR smooth pigweed collected in 2021 were used to screen for the efficacy of alternative herbicides. Seeds sourced from Bergville (28°43’23″S, 29°20’37″E) and Winterton (28°49’42″S, 29°32’5″E) districts in KwaZulu-Natal Province, together with their F1 seed lines, were confirmed to be highly resistant to glyphosate (Shabangu et al. Reference Shabangu, Simelane, Reinhardt, Marais and Vorster2024). The F1 seed lines were obtained by allowing the GR smooth pigweed plants from each population to grow to maturity and produce seeds in isolation from the other populations. While smooth pigweed plants are predominantly self-pollinated, the proximity of plants (approximately 10 plants m–1) might have allowed some degree of cross-pollination within each resistant population. A glyphosate-susceptible (GS) population from Hendrina (26°09’20″S, 29°44’02″E) district in Mpumalanga Province served as the control.
Preemergence and postemergence herbicide experiments were conducted concurrently in a greenhouse at the University of Pretoria (25°44’59″S, 28°15’19″E) using GR F1 seed lines from Bergville and Winterton and the GS population from Hendrina. Greenhouse conditions were maintained at an average temperature of 25 C under natural light, supplemented with high-pressure sodium light bulbs to simulate a 16/8-h day/night photoperiod for both experiments. The preemergence and postemergence herbicide experiments were conducted in 2023 and repeated in the 2024 growing season. We selected herbicide products with different SOAs registered for the control of smooth pigweed in both maize and soybean production in South Africa (Table 1).
Preemergence and postemergence herbicides evaluated in greenhouse experiments in 2023 and 2024. a

a Abbreviation: HRAC, Herbicide Resistant Action Committee. HRAC classifies herbicides into groups according to their mode of action.
b Trade names and recommended field-use rates are based on their registration in South Africa. The recommended field-use rates represent the standard label rate (1×) registered for field application in South Africa and is presented in grams of active ingredient per hectare (g ai ha−1).
c Where indicated, the following adjuvants were added: polyether-polymethylsiloxane-copolymer, 1,000 g L−1 (Complement Super, Syngenta); methylated vegetable oil, 700 g L−1 (Ballista, Bayer Crop Science), and alcohol ethoxylate, 250 g L−1 (Laeveld Agrochem).
Experiment 1: Preemergence Herbicides Study
For the preemergence herbicide experiment, natural mineral topsoil (field soil) classified as sandy loam with pH 4.61, 0.55% carbon, and 12% clay was used. Soil was collected from the Miertjie Le Roux Experimental Farm at the University of Pretoria (25°46’56″S, 28°33’07″E). During trial establishment, 900 cm3 of soil was filled into 25- by 40-cm clear plastic bags before being placed in 1,000 cm3 seedling pots. The seedling pots were equipped with drainage holes, thus the plastic bags prevented the herbicide from leaching out of the root zone by keeping it within the root zone. The soils were leveled to produce a smooth planting surface, and water was applied using a misting nozzle to field capacity (Westerveld et al. Reference Westerveld, Soltani, Hooker, Robinson, Tranel, Laforest and Sikkema2021), which was predetermined on the field soil. Thereafter, an average of 20 smooth pigweed seeds from each population were sown on the soil surface and covered with a 0.5- to 1-cm layer of topsoil. After sowing, the seedling pots were watered until the soil was moist.
The experiment was arranged in a completely randomized design with two factors: populations (GR populations from Bergville and Winterton districts, and a GS population from Hendrina district) and herbicide rates. GR and GS smooth pigweed populations were screened against different herbicide products representing various herbicide groups, with each product treated as an independent study. Preemergence herbicides were applied 12 h after sowing, and spraying was conducted in an enclosed room to prevent herbicide contamination with nontarget plants. A 1-m2 sampling quadrat was used, where five pots were systematically arranged inside this area. The herbicide rates were applied using an Oxford Small-Plot Precision Sprayer equipped with a TeeJet E80015 EVS nozzle. The spray volume was 250 L ha–1, and the sprayer was operated at 180 kPa. Each herbicide was applied at five different rates (0×, 0.5×, 1×, 2×, and 4× the recommended field-use rate) based on the soil characteristics for the various herbicide products (Table 1). The untreated control (0×) served as an indicator of seed dormancy and viability, and each herbicide rate was replicated five times.
Seedling pots were watered immediately after herbicide application with a volume of water equivalent to 10 mm of rain to incorporate and activate the herbicide active ingredients in the soil. Thereafter, seedling pots were watered to field capacity at 2- to 3-d intervals depending on the soil moisture content. Nutrition sourced from a hygroponic N-P-K 1.6:1:5 mixture (Hygrotech, South Africa) was applied 2 wk after emergence at a concentration of 1 g L−1. Thereafter, nutrition was applied 27 d after treatment (DAT). The efficacy of preemergence herbicides was visually evaluated by determining the percentage of plant density reduction at 32 DAT. Percentage plant density reduction was calculated by counting the number of smooth pigweed plants with at least one true leaf per unit area, compared with untreated control plants using Equation 1 (Wortman Reference Wortman2014):
 $${\rm{Plant\;density\;reduction}}\;\left( \% \right) = \left( {1 - {{{PCEU}}\over{{PCUC}}}} \right) \times 100$$
$${\rm{Plant\;density\;reduction}}\;\left( \% \right) = \left( {1 - {{{PCEU}}\over{{PCUC}}}} \right) \times 100$$
where PCEU is the plant count of the experimental unit, and PCUC is the average plant count of the untreated controls for the respective populations. Herbicide products that provided <90% (± standard error) plant density reduction at 32 DAT were considered ineffective in controlling the smooth pigweed populations (Vennapusa et al. Reference Vennapusa, Faleco, Vieira, Samuelson, Kruger, Werle and Jugulam2018).
Experiment 2: Postemergence Herbicides Study
For the postemergence experiment, smooth pigweed seeds were sown in 50- by 30-cm seedling trays filled with pure sand (Silica Quartz, Delmas, South Africa) and coconut peat (Pelemix, South Africa) mixture in a 6:1 sand:coconut peat ratio by weight. Seedlings in the greenhouse were watered with tap water, and nutrition was sourced from a hygroponic N-P-K 1.6:1:5 mixture twice a week throughout the experiment. At the 4-leaf stage, seedlings were transplanted into 12.5-cm-diam seedling pots filled with a 6:1 mixture of pure sand and coconut peat. Two plants were transplanted into each seedling pot and kept under greenhouse conditions.
Herbicides were applied when seedlings were at the 6-leaf stage (8 to 12 cm in height) at five different herbicide rates (0×, 0.5×, 1×, 2×, and 4× the field-use rate) (Table 1). The experiment was a completely randomized design consisting of two factors: smooth pigweed populations and herbicide rates. Each herbicide rate was replicated five times. Spraying was conducted in an enclosed spraying room, as described in the preemergence experiment. The spray volume for postemergence herbicides was 300 L ha−1, and the sprayer was operated at 180 kPa.
Herbicide efficacy (phytotoxic effects) for postemergence herbicides was evaluated at 16 DAT as a percentage of visual control (PVC) on a scale of 0% to 100%, where 0% indicated no herbicide injury and 100% indicated complete plant death with no signs of recovery. At 35 DAT, the surviving plants were counted, and the counts were converted into a survival percentage (Equation 2), with dead plants scored as 0% (no green tissue) and alive plants scored as 100% (green plants with evidence of regrowth). Populations with ≥50% survival rates were classified as resistant to each mode of action (Schultz et al. Reference Schultz, Chatham, Riggins, Tranel and Bradley2015). Thereafter, all plants were clipped from the soil surface and dried at 80 C for 48 h to determine the percentage of biomass (biomass of treated plants expressed as a percentage of untreated control biomass) using Equation 3 (adapted and modified from Wortman Reference Wortman2014):
 $${\rm Survival\, percentage} = {{{PCEU}}\over{{PCUC}}}\times 100 $$
$${\rm Survival\, percentage} = {{{PCEU}}\over{{PCUC}}}\times 100 $$
 $$ {\rm Percentage\, biomass} ={{{BEU}}\over{{BUC}}} \times 100 $$
$$ {\rm Percentage\, biomass} ={{{BEU}}\over{{BUC}}} \times 100 $$
where BEU is the average biomass of the experimental unit, and BUC is the average biomass of the untreated controls.
Statistical Analysis
The experimental data used in this study came from two separate experiments conducted in 2023 and 2024. Data from both experimental runs were pooled to increase the number of replicates because the experimental runs had no significant effect on the response variables. Data analysis was performed using R software (version 4.4.3) (R Core Team 2024). The response variables (percentage plant density reduction, PVC, and percentage biomass) were checked for normality using Shapiro-Wilk tests, homogeneity of variance using Levene tests, and normal Q-Q plots (Arsenijevic et al. Reference Arsenijevic, Bernards, DeWerff, Arneson, Smith and Werle2025; McDonald et al. Reference McDonald, Striegel, Chahal, Jha, Rees, Proctor and Jhala2021).
A generalized linear model was used in this study. Generalized linear models have been applied in different disciplines, with notable use in herbicide resistance studies (Faleco et al. Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a, Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b; Godar et al. Reference Godar, Norsworthy and Barber2023; Larini et al. Reference Larini, Albrecht, Neuberger, Barroso and Albrecht2025). In this study, a generalized linear model with a beta distribution and a logit link function was used due to the nature of our data (i.e., skewed continuous positive response variables that are limited between 0% and 100%) (Bono et al. Reference Bono, Alarcón and Blanca2021). The model was implemented using the betareg package (Cribari-Neto and Zeileis Reference Cribari-Neto and Zeileis2010) and the lme4 package (McDonald et al. Reference McDonald, Striegel, Chahal, Jha, Rees, Proctor and Jhala2021). Herbicide rates and smooth pigweed populations were considered as main effects used to explain the response variables. Model 1 tested the main effects individually, and Model 2 tested their interaction. The best model was selected based on Akaike information criterion (Charles et al. Reference Charles, Sindel, Cowie and Knox2019; Priess et al. Reference Priess, Norsworthy, Godara, Mauromoustakos, Butts, Roberts and Barber2022; Schwarz Reference Schwarz1978) and Bayesian information criterion (Priess et al. Reference Priess, Norsworthy, Godara, Mauromoustakos, Butts, Roberts and Barber2022; Schwarz Reference Schwarz1978) model fit indicators. Multicollinearity was tested for Model 1, and the main effects were found to be uncorrelated. To ensure that we had robust standard errors and confidence intervals, we performed 1,000 bootstrap replications with a seed value of 2,025 (Efron and Tibshirani Reference Efron and Tibshirani1986).
Post hoc analysis was conducted using the emmeans package in R to compute the estimated marginal means for the fixed effects. A Tukey HSD adjustment was used for multiple comparisons, with statistical significance set at P ≤ 0.05 (Souza et al. Reference Souza, Norsworthy, Carvalho-Moore, Godar, Fernandes and Butts2025). Comparisons were conducted among smooth pigweed populations at each herbicide rate and across herbicide rates within each population. ANOVA was performed on the fitted model with the Type III Wald chi-square test. Back-transformed means ± standard error (±SE) are presented to facilitate result interpretation. Survival percentage data exhibited a clear binary response pattern, with 100% survival in untreated controls and resistant populations, and 0% survival in susceptible populations. Due to complete separation and overdispersion, the data did not meet the generalized linear model assumptions. As a result, survival percentages were presented descriptively in a figure format to illustrate the observed patterns.
Results and Discussion
Efficacy of Preemergence Herbicides on Smooth Pigweed Populations
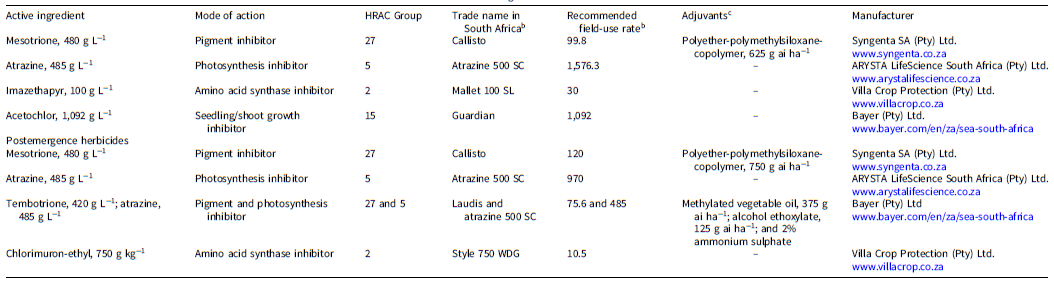
Mesotrione. The analysis revealed a significant main effect of herbicide rates (P < 0.001) and a significant interaction between herbicide rates and population (P < 0.001), indicating that smooth pigweed populations responded differently to increasing mesotrione rates. Mesotrione effectively controlled the GR populations from Bergville and Winterton districts and the GS population from Hendrina, as the plant density reduction was >95% from the 1× to 4× rates. However, at the 0.5× rate, the plant density Hendrina population was reduced by 77.8% (±2.3%), which was significantly lower than the Bergville and Winterton populations (Figure 1). The findings of this study agreed with those reported by Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a, Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b) and Papapanagiotou et al. (Reference Papapanagiotou, Alvanou, Giantsis, Kati and Eleftherohorinos2025) on waterhemp, Palmer amaranth, and redroot pigweed (Amaranthus retroflexus L.), respectively. In the study reported by Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a), application of mesotrione to waterhemp at rates from the recommended field-use rate to 3× the recommended rate resulted in a density reduction of >95% of populations that exhibited reduced sensitivity to atrazine and fomesafen (Group 14) applied preemergence. Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b) reported similar results in Palmer amaranth populations that were resistant to atrazine applied preemergence.
Plant density reduction (± standard error) of smooth pigweed populations in response to preemergence herbicides, illustrating the interaction between population and herbicide rate. Treatments with the same letters are not significantly different (Tukey HSD test, P ≤ 0.05). Herbicide rates are expressed relative to the recommended field-use rate for each herbicide product. Herbicide rates for each product resulting in <90% plant density reduction (red line) were considered ineffective. Recommended field-use rates (1×) are as follows: mesotrione, 99.8 g ai ha−1; atrazine, 1,576.3 g ai ha−1; imazethapyr, 30 g ai ha−1; acetochlor, 1,092 g ai ha−1.

Atrazine. Analysis showed significant effects of herbicide rate (P < 0.001) and the interaction between herbicide rate and population on plant density (P < 0.001), indicating that populations responded differently across atrazine rates. At the 0.5× rate, plant density reduction was 89% (±1.8%) and 77.2% (±2.3%) for the Hendrina and Winterton populations, respectively, whereas the Bergville population was completely controlled (100% ±0.5%). From the 1× to 4× rates, plant density reduction was 100% (±0.5%) (Figure 1), confirming that atrazine was highly effective against smooth pigweed populations. Vennapusa et al. (Reference Vennapusa, Faleco, Vieira, Samuelson, Kruger, Werle and Jugulam2018) reported a 100% plant density reduction in atrazine-resistant waterhemp populations when metribuzin (Group 5) was applied preemergence at 1,345 g ai ha−1. Similarly, Papapanagiotou et al. (Reference Papapanagiotou, Alvanou, Giantsis, Kati and Eleftherohorinos2025) demonstrated that a preemergence application of metribuzin effectively controlled redroot pigweed populations that evolved cross-resistance to inhibitors of acetolactate synthetase. In contrast, Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a) reported reduced efficacy of atrazine at the recommended field-use rate by some waterhemp populations, whereas in another report by Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b), a similar trend was observed in Palmer amaranth populations that were resistant to imazethapyr (Group 2), atrazine, and glyphosate applied postemergence.
Imazethapyr. The rates of imazethapyr applied preemergence had a significant effect (P < 0.001) on the plant density reduction of smooth pigweed populations compared with untreated control plants. The density reduction in all the populations ranged from 93.3% to 99.6% at imazethapyr rates that ranged from 0.5× to 4×, with no significant difference between herbicide rates (Figure 1). Imazethapyr was effective in controlling all smooth pigweed, with a plant density reduction of more than 90%. The density reduction in populations from Hendrina and Winterton from the 0.5× to 4× rates ranged from 96% to 99.6%. However, the density reduction of the Bergville population ranged from 93.3% to 93.9% between the 0.5× and 2× rates (Figure 1). Spiny amaranth (Amaranthus spinosus L.) was identified as a susceptible biotype in lettuce (Lactuca sativa L.) fields, achieving more than 95% control when imazethapyr was applied preemergence to soils with 80% organic matter (Odero and Wright Reference Odero and Wright2022).
Acetochlor. Acetochlor applied preemergence effectively controlled all smooth pigweed populations, with no significant difference observed between herbicide rates and populations (P > 0.05). The density reduction in all smooth pigweed populations across all herbicide rates was 100% (Figure 1). The findings of this study correspond with those reported by Reinhardt et al. (Reference Reinhardt, Vorster, Küpper, Peter, Simelane, Friis, Magson and Aradhya2022) on a GR Palmer amaranth population from the Northern Cape Province in South Africa. In that study, acetochlor and pyroxasulfone provided 100% and 95% control of Palmer amaranth, respectively, when they were applied at the recommended field-use rates. Symington et al. (Reference Symington, Soltani, Kaastra, Hooker, Robinson and Sikkema2024) also reported that the density of susceptible waterhemp population was reduced by 99% when acetochlor (2,950 g ai ha−1) was applied preemergence. However, in a study conducted by Strom et al. (Reference Strom, Gonzini, Mitsdarfer, Davis, Riechers and Hager2019), when an encapsulated formulation of acetochlor (2,700 g ai ha−1) was applied, the percentage control was 75%, whereas the efficacy of S-metolachlor was remarkably reduced to 60% in the same population. Our findings also support those of Symington et al. (Reference Symington, Soltani, Kaastra, Hooker, Robinson and Sikkema2024), who reported that the density of a waterhemp population was reduced by 98% when treated with acetochlor.
Efficacy of Postemergence Herbicides on Smooth Pigweed Populations
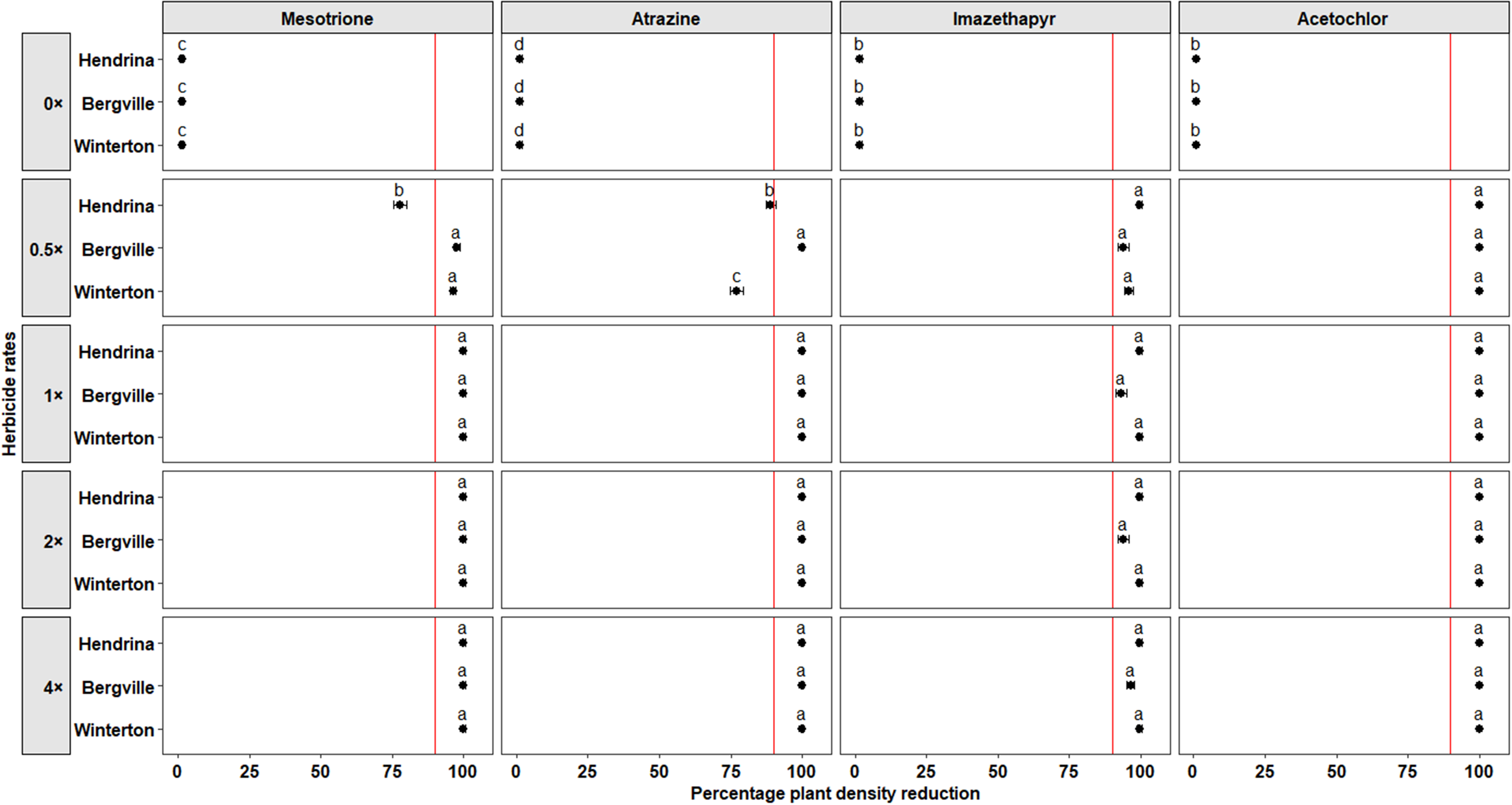
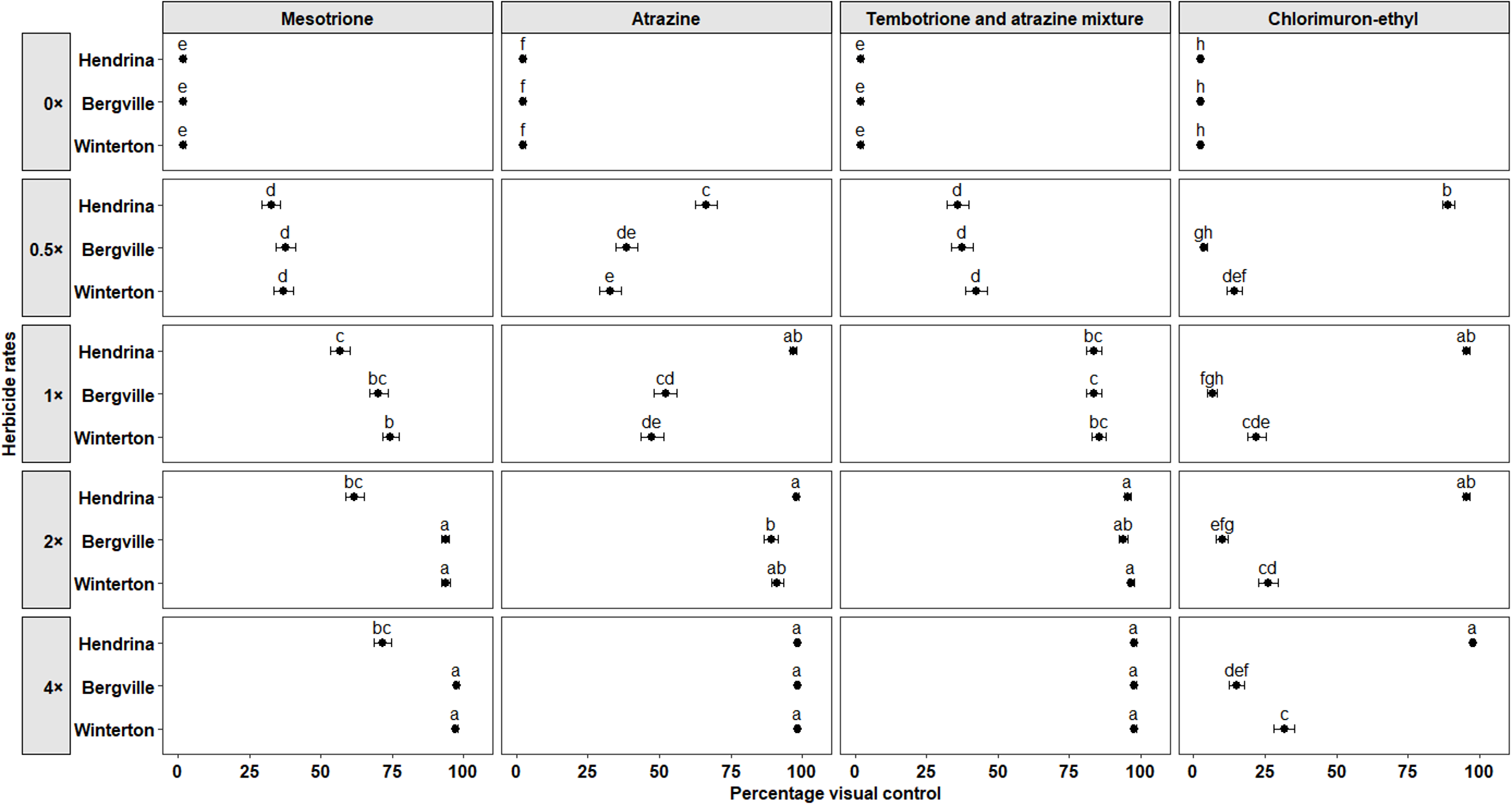
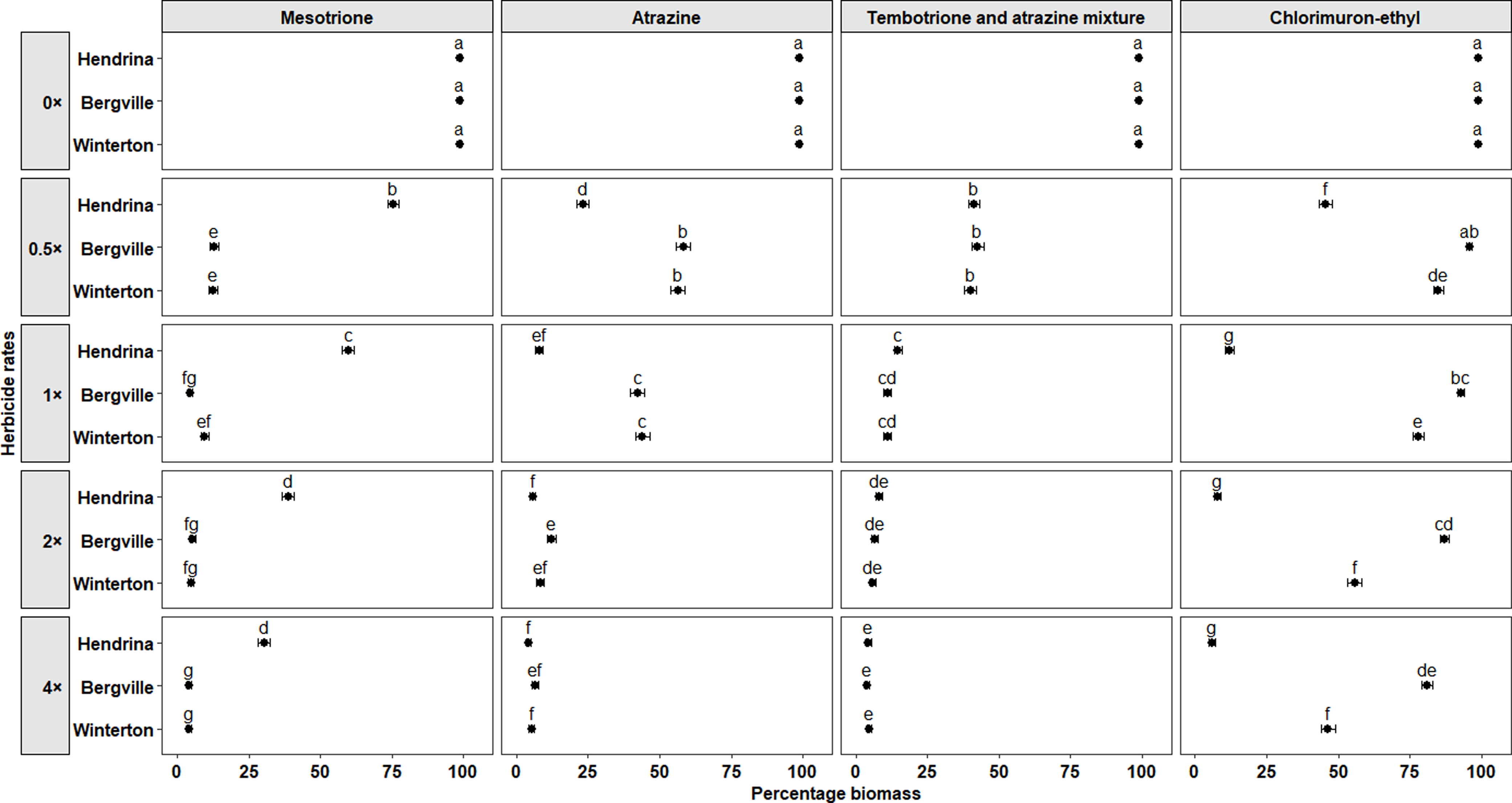
Mesotrione. The PVC varied when mesotrione was applied postemergence at 16 DAT with a significant effect of herbicide rate (P < 0.001), and a significant interaction between herbicide rate and population (P < 0.001), suggesting that the populations responded differently to mesotrione rates. At the 1× rate, PVC for the Bergville and Winterton populations was 70.3% (±3.1%) and 74.5% (±3%), respectively, compared with 56.8% (±3.4%) for the Hendrina population. At the 2× and 4× rates, the PVC for the Bergville and Winterton populations was greater than 93%, while the PVC for the Hendrina population was 61.9% (±3.4%) and 71.6% (±3.1%), respectively (Figure 2). At 35 DAT, the Bergville and Winterton populations were effectively controlled, even at the 0.5× rate, while the Hendrina population exhibited 100% survival at all rates of mesotrione (Figure 3). Herbicide rate and its interaction with population also had significant effects (P < 0.001) on the percentage of biomass. At the 1× rate, the percentage of biomass for the Bergville and Winterton populations was reduced to 4.5% (±0.9%) and 9.6% (±1.3%), respectively, compared with 59.7% (±2.2%) for the Hendrina population. At the 2× and 4× rates, the percentage of biomass for the Bergville and Winterton populations was <6% while for the Hendrina population, it was >30% (Figure 4).
Visual control (± standard error) of smooth pigweed populations in response to postemergence herbicides, illustrating the interaction between population and herbicide rate. Treatments with the same letters are not significantly different (Tukey HSD test, P ≤ 0.05). Herbicide rates are expressed relative to the recommended field-use rate for each herbicide product. Recommended field-use rates (1×) are as follows: mesotrione, 120 g ai ha−1; atrazine, 970 g ai ha−1; tembotrione and atrazine tank mixture, 75.6 and 485 g ai ha−1; chlorimuron-ethyl, 10.5 g ai ha−1.

Survival of smooth pigweed populations in response to postemergence herbicides, illustrating the interaction between population and herbicide rate. Herbicide rates are expressed relative to the recommended field-use rate for each herbicide product. Herbicide rates for each herbicide product resulting in >50% survival rate (red line) were considered ineffective. Recommended field-use rates (1×) are as follows: mesotrione, 120 g ai ha−1; atrazine, 970 g ai ha−1; tembotrione and atrazine tank mixture, 75.6 and 485 g ai ha−1; chlorimuron-ethyl, 10.5 g ai ha−1.

Biomass (± standard error) of smooth pigweed populations in response to postemergence herbicides, illustrating the interaction between population and herbicide rate. Treatments with the same letters are not significantly different (Tukey HSD test, P ≤ 0.05). Herbicide rates are expressed relative to the recommended field-use rate for each herbicide product. Recommended field-use rates (1×) are as follows: mesotrione, 120 g ai ha−1; atrazine, 970 g ai ha−1; tembotrione and atrazine tank mixture, 75.6 and 485 g ai ha−1; chlorimuron-ethyl, 10.5 g ai ha−1.

This study demonstrates that the Bergville and Winterton populations were susceptible to mesotrione. In contrast, the Hendrina population exhibited reduced sensitivity to mesotrione, suggesting putative resistance to an inhibitor of 4-hydroxphenylpyruvate dioxygenase (HPPD). The findings on the Bergville and Winterton populations are consistent with those reported by Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a; Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b), that it was possible to achieve effective control of waterhemp and Palmer amaranth populations with mesotrione. Hausman et al. (Reference Hausman, Tranel, Riechers and Hager2016) reported <40% control of waterhemp populations when treated with sole applications of mesotrione, tembotrione, or topramezone. Reinhardt et al. (Reference Reinhardt, Vorster, Küpper, Peter, Simelane, Friis, Magson and Aradhya2022) further documented resistance to mesotrione in Palmer amaranth from South Africa’s North Cape Province. Although the Hendrina population exhibited reduced sensitivity to mesotrione in this study, it cannot yet be classified as being resistant to mesotrione. Further research is needed to determine whether this reduced sensitivity represents resistance to Group 27 herbicides.
Atrazine. The analysis revealed a significant main effect of herbicide rate (P < 0.001) and a significant interaction between herbicide rate and population (P < 0.001) on PVC, indicating differential responses among smooth pigweed populations. At the 0.5× rate, the PVC was 66.5% (±3.8%) for the Hendrina population, 38.6% (±3.9%) for the Bergville population, and 32.8% (±3.7%) for the Winterton population. At the 1× rate, the Hendrina population was effectively controlled with a PVC of 96.8% (±1.1%), while the Bergville and Winterton populations exhibited partial control, with PVC values of 52.2% (±4%) and 47.3% (±4%), respectively (Figure 2). At the 2× and 4× rates, the PVC of the Bergville and Winterton populations increased significantly, indicating improved control at higher rates (Figure 2). Survival percentage at 35 DAT supported these findings: at the 0.5× rate, 60% of the Hendrina population survived, 100% of the Bergville population survived, as did 80% of the Winterton population (Figure 3). At the 1× rate, the Bergville and Winterton populations were not effectively controlled, with survival percentages of 70% and 50%, respectively. The Winterton population was effectively controlled at the 2× and 4× rates. Forty percent of the Bergville population survived applications of atrazine at the 2× rate, whereas complete control was achieved at the 4× rate (Figure 3). Percentage biomass was also a statistically significant main effect of herbicide rate (P < 0.001) and a significant interaction with population (P < 0.001). These results suggest differential sensitivity to atrazine, with biomass decreasing more substantially in susceptible populations as the herbicide rate is increased. The percentage biomass for the Hendrina, Bergville, and Winterton populations at the 1× rate was 7.9% (±1.3%), 42.2% (±3.2%), and 44% (±2.5%), respectively; however, at the 2× and 4× rates, the biomass was <13% for all the populations, suggesting that atrazine was effective at higher rates (Figure 4).
Our findings suggest that smooth pigweed populations may survive the 1× rate of atrazine when sprayed after the 4-leaf stage (Braz and Takano Reference Braz and Takano2022; Kaundun Reference Kaundun2021). In other studies, Faleco et al. (Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022a, Reference Faleco, Oliveira, Arneson, Renz, Stoltenberg and Werle2022b) reported reduced efficacy of atrazine at the recommended field-use rate on Amaranthus species when applied at the 4- to 6-leaf stage. However, the Amaranthus species were effectively controlled when atrazine was applied at 3× the recommended field-use rate. Ma et al. (Reference Ma, Evans and Riechers2016) also reported that waterhemp populations survived atrazine applications when applied at the 8- to 10-leaf stage. In South Africa, smooth pigweed populations have been reported to survive even at 10× the recommended field-use rate of atrazine (Sereda et al. Reference Sereda, Erasmus and Coetzer1996). Recently, Reinhardt et al. (Reference Reinhardt, Vorster, Küpper, Peter, Simelane, Friis, Magson and Aradhya2022) reported reduced efficacy of atrazine in a GR Palmer amaranth population from the North Cape Province.
Tembotrione and atrazine tank mixture. This mixture, applied postemergence, had additive effects in controlling smooth pigweed populations. The increased phytotoxicity was attributed to the main effect of herbicide rate, which was statistically significant (P < 0.001). At the 1× rate of tembotrione and atrazine, PVC was 83.7% (±2.7%), 83.5% (±2.7%), and 85.5% (±2.5%) for the Hendrina, Bergville, and Winterton populations, respectively. At the 2× and 4× rates, PVC was >90% across all populations, indicating enhanced control (Figure 2). Survival assessment at 35 DAT showed complete control, with 0% survival even at the 0.5× rates of tembotrione and atrazine tank mixture (Figure 3). Percentage biomass analysis also indicated a significant reduction due to the main effect of herbicide rate (P < 0.001). From the 1× to 4× rates of tembotrione and atrazine tank mixture, the percentage of biomass remained below 15% for all populations, which was significantly less than that of the untreated controls and the 0.5× rate (Figure 4).
The inhibition of carotenoid biosynthesis by HPPD inhibitors increases the binding efficacy and efficiency of photosystem II (PS II) inhibitors by reducing the reformation of the D1 protein after photoinhibition has been initiated (Armel et al. Reference Armel, Rardon, McComrick and Ferry2007). This process explains why the tembotrione and atrazine tank mixture provided excellent control of GR smooth pigweed populations compared to the sole application of mesotrione or atrazine in our experiments. Preemergence and postemergence studies of tank mixtures of HPPD-inhibiting herbicides with other PS II inhibitors (non-atrazine) have also demonstrated an additive effect in controlling dicot weeds. Other studies found that a mesotrione and atrazine tank mixture effectively controlled several annual weeds compared to a sole applications of HPPD inhibitors such as mesotrione, topramezone, tolpyralate, isoxaflutole, and tembotrione (Armel et al. Reference Armel, Hall, Wilson and Cullen2005; Osipitan et al. Reference Osipitan, Scott and Knezevic2018; Willemse et al. Reference Willemse, Soltani, Benoit, Jhala, Hooker, Robinson and Sikkema2021).
Chlorimuron-ethyl. This herbicide, applied postemergence, exhibited a significant main effect of herbicide rate (P < 0.001) and a significant interaction between herbicide rate and population (P < 0.001) on PVC, demonstrating that the smooth pigweed populations responded differently when various chlorimuron-ethyl rates were applied. At the 1× rate, the PVC values for the Hendrina, Bergville, and Winterton populations were 89.2% (±2.2%), 6.5% (±1.6%) and 22.1% (±3.2%), respectively. The Bergville population exhibited a high level of resistance to chlorimuron-ethyl, with PVC values of 10.1% (±2.1%) and 15.1% (±2.7%) at the 2× and 4× rates, respectively (Figure 2). The Winterton population exhibited significantly greater phytotoxicity than the Bergville population at all rates (Figure 2). At 35 DAT, the survival percentage of the Bergville and Winterton populations remained at 100% at all chlorimuron-ethyl rates, confirming resistance. In contrast, the Hendrina population exhibited 20% survival at the 0.5× rate, and 0% survival at the 1× to 4× rates, indicating susceptibility (Figure 3). A significant main effect of herbicide rate (P < 0.001) and a significant interaction between herbicide rate and population (P < 0.001) were also observed for biomass percentage. At the 1× rate, the Hendrina population had a biomass of 10.5% (±1.55), which was significantly lower than that of the Bergville and Winterton populations, at 93% (±1.2%) and 78% (±2.0%), respectively (Figure 4). At the 2× rate, the biomass of the Hendrina, Bergville, and Winterton populations was 7.7% (±1.2%), 46.5% (±2.4%), and 81% (±1.9%), respectively (Figure 4).
This is the first scientific report on the resistance of smooth pigweed populations to chlorimuron-ethyl (Group 2) in the Republic of South Africa; however, Palmer amaranth has been reported in the country to have evolved resistance to chlorimuron-ethyl (Reinhardt et al. Reference Reinhardt, Vorster, Küpper, Peter, Simelane, Friis, Magson and Aradhya2022). Aulakh et al. (Reference Aulakh, Chahal, Kumar, Price and Guillard2021) also reported some differences in the response of Palmer amaranth populations to imazaquin, with the highest rate (2,196 g ai ha−1) controlling 18% of plants in some populations. Milani et al. (Reference Milani, Scarabel and Sattin2020) reported a smooth pigweed population resistant to imazamox with survival ranging from 45% to 95%. A smooth pigweed population from Argentina, reported by Larran et al. (Reference Larran, Lorenzetti, Tuesca, Perotti and Permingeat2018), had a survival of more than 80%, even at 10 times the recommended field-use rates of imazethapyr and chlorimuron-ethyl. The mechanism of resistance to chlorimuron-ethyl was not established in this study; however, differences in the responses of the Bergville and Winterton populations could be attributed to the involvement of different resistance alleles (Milani et al. Reference Milani, Scarabel and Sattin2020). These alleles carry mutations that alter the binding site of herbicides, thereby influencing the resistance levels of weeds. The presence of other herbicide resistance mechanisms such as overexpression of the ALS gene or metabolic resistance, has also been reported in herbicide resistance studies (Han et al. Reference Han, Gao, Sun, Wang, Yan, Ma, Liu and Huang2024; Sen et al. Reference Sen, Hamouzová, Mikulka, Bharati, Košnarová, Hamouz, Roy and Soukup2021; Wang et al. Reference Wang, Sun, Lan, Wei, Huang, Li and Huang2023).
Reduced Sensitivity of Smooth Pigweed Populations to Atrazine and Mesotrione Rates
The Bergville and Winterton populations survived at the recommended field-use rate of atrazine but were effectively controlled at higher rates. In contrast, both populations were susceptible to mesotrione, with effective control achieved even at the 0.5× rate. These findings contrast with observations of cross-resistance when resistance to HPPD inhibitors is accompanied by resistance to PS II inhibitors. This pattern is primarily attributed to non-target site resistance mechanisms, particularly enhanced herbicide metabolism. In Amaranthus species, metabolic resistance to HPPD inhibitors is commonly facilitated by overexpression of the cytochrome P450 enzymes (Concepcion et al. Reference Concepcion, Kaundun, Morris, Brandenburg and Riechers2024; Kaundun et al. Reference Kaundun, Hutchings, Dale, Howell, Morris, Kramer, Shivrain and Mcindoe2017; Küpper et al. Reference Küpper, Peter, Zöllner, Lorentz, Tranel, Beffa and Gaines2018; Nakka et al. Reference Nakka, Godar, Wani, Thompson, Peterson, Roelofs and Jugulam2017b), while resistance to PS II inhibitors is mostly associated with glutathione S-transferases (GST) activity (Evans et al. Reference Evans, O’Brien, Ma, Hager, Riggins, Lambert and Riechers2017; Nakka et al. Reference Nakka, Godar, Thompson, Peterson and Jugulam2017a; Vennapusa et al. Reference Vennapusa, Faleco, Vieira, Samuelson, Kruger, Werle and Jugulam2018). Therefore, the resistance to atrazine observed in the Bergville and Winterton populations may similarly be due to GST-mediated detoxification. Target-site resistance involving a nonsynonymous mutation in the psbA gene encoding the D1 protein could also explain the reduced sensitivity in these populations. The most common target-site resistance mechanism known to confer resistance to PS II inhibitors in Amaranthus species is the Ser-264-Gly mutation in the psbA gene, which reduces the binding affinity of PS II inhibitors at the QB site (Nakka et al. Reference Nakka, Godar, Thompson, Peterson and Jugulam2017a).
The reduced sensitivity to mesotrione exhibited by the Hendrina smooth pigweed population raises concerns about the potential evolution of resistance to HPPD inhibitors. Additionally, the survival of the Bergville and Winterton populations at the recommended field-use rate of atrazine is a considerable concern in managing smooth pigweed populations. Herbicide tank mixtures, such as the combination of atrazine and HPPD inhibitors, are widely used to delay the evolution of herbicide resistance and improve weed control. However, the effectiveness of such tank mixtures can be significantly compromised when resistance evolves to one mode of action.
Practical Implications
Management of GR smooth pigweed remains a major challenge for maize and soybean farmers in South Africa. This issue is particularly severe in soybeans due to the limited number of SOAs compared with maize. Mesotrione, atrazine, and acetochlor, applied preemergence are recommended for use in maize production to manage GR populations of smooth pigweed. However, atrazine has long-lasting residual effects when applied preemergence, compelling farmers to wait up to 24 mo before growing susceptible crops such as soybeans and dry beans (Phaseolus vulgaris L.). Therefore, atrazine is best suited for monoculture systems or rotation systems involving crops that are tolerant to it. For preemergence applications to soybeans, imazethapyr is recommended; however, a follow-up with an early postemergence application with fomesafen is recommended to manage surviving plants. When applied to maize, a postemergence application of tembotrione and atrazine tank mixture was more effective than either mesotrione or atrazine applied alone. However, the sole application of mesotrione was effective only in controlling the GR populations. Atrazine should be applied at the 2- to 4-leaf stage, and delayed application reduces the herbicide’s effectiveness, likely due to increased plant size and metabolic capacity. Chlorimuron-ethyl was not effective in controlling the GR populations, highlighting the need to evaluate additional herbicide products with other SOAs for use in soybean production.
To reduce the risk of herbicide resistance, farmers should adopt integrated weed management strategies that include herbicide rotation with different SOAs, use of herbicide mixtures with multiple SOAs, sequential herbicides of different SOAs at each application, and integration of nonchemical management techniques. Nonchemical management techniques may include the use of fast-growing crop cultivars, cover crops to suppress the germination of Amaranthus species, increasing crop density, weed mowing, and preventing seed production from flowering plants.
Acknowledgments
We thank Mr. Jacques Marneweck and the Innovation Africa personnel at the University of Pretoria for their technical assistance.
Funding
This work was funded through the South African Herbicide Resistance Initiative (SAHRI), which is supported by Bayer SA, Syngenta SA, and The Maize Trust. The National Research Foundation of South Africa and the Oil and Protein Seeds Development Trust provided support for postgraduate students.
Competing Interests
The authors declare they have no competing interests.

 Open access
Open access