


Introduction
Palmer amaranth (Amaranthus palmeri S. Watson) is a fast-growing, highly competitive, and dioecious annual weed that has emerged as a dominant and economically damaging species in U.S. agriculture (Van Wychen Reference Van Wychen2017). Amaranthus palmeri was ranked among the five most troublesome species in 2009 (Webster and Nichols Reference Webster and Nichols2012). It is native to the southwestern United States and northern Mexico (Sauer Reference Sauer1957). Currently, it has spread throughout the southern and midwestern United States, thriving in a wide range of agroecosystems. Its rapid expansion and persistence have been attributed to several biological and ecological factors, such as a C4 photosynthetic pathway, prolific seed production, genetic diversity, and widespread herbicide resistance (Chahal et al. Reference Chahal, Irmak, Jugulam and Jhala2018; Ward et al. Reference Ward, Webster and Steckel2013). It can severely limit crop performance by restricting the availability of soil nutrients, moisture, and sunlight (Berger et al. Reference Berger, Ferrell, Rowland and Webster2015).
Amaranthus palmeri is a major agronomic concern in row-cropping systems, including cotton (Gossypium hirsutum L.), corn (Zea mays L.), and soybean [Glycine max (L.) Merr.], in which it has been associated with yield losses of up to 54%, 91%, and 68%, respectively (Hall et al. Reference Hall, Urban, Wagentristl, Karrer, Winter, Czerny and Kaul2021; Liphadzi and Dille Reference Liphadzi and Dille2006; Massinga et al. Reference Massinga, Currie, Horak and Boyer2001; Ward et al. Reference Ward, Webster and Steckel2013). Female A. palmeri plants are prolific seed producers that can generate 200,000 to 600,000 seeds per plant under optimal conditions (Keeley et al. Reference Keeley, Carter and Thullen1987). Notably, high seed production is maintained even under competitive or stress-inducing environments (Massinga et al. Reference Massinga, Currie, Horak and Boyer2001; Sellers et al. Reference Sellers, Smeda, Johnson, Kendig and Ellersieck2003). Amaranthus palmeri’ s high reproductive capacity ensures that a single escape can lead to substantial seedbank replenishment, exacerbating future infestations and long-term management challenges (Norsworthy et al. Reference Norsworthy, Ward, Shaw, Llewellyn, Nichols, Webster, Bradley, Frisvold, Powles and Burgos2012).
One of the key traits contributing to the success of A. palmeri is its exceptional adaptability to environmental stress. In particular, its tolerance to drought, heat, and nutrient limitation makes it well suited to thrive in semiarid and arid agricultural environments, including the Texas High Plains (Cominelli and Patrignani Reference Cominelli and Patrignani2022; Matzrafi et al. Reference Matzrafi, Osipitan, Ohadi and Mesgaran2021). This region, historically reliant on irrigation from the Ogallala Aquifer, is increasingly facing water scarcity due to aquifer depletion and inconsistent rainfall patterns (Musick et al. Reference Musick, Jones, Stewart and Dusek1994; Nativ and Smith Reference Nativ and Smith1987). The Ogallala Aquifer in particular has been depleted by more than 50% of its maximum water storage capacity from 1935 to 2012 (Haacker et al. Reference Haacker, Kendall and Hyndman2016). As water availability declines, there has been a significant shift from irrigated to dryland cropping systems, which intensifies the importance of understanding plant responses to water-limited conditions.
Water is a critical resource in crop and weed physiology, influencing plant growth, development, and reproduction. While crops often suffer from yield penalties under drought stress, certain weed species, including A. palmeri, may exhibit considerable plasticity and maintain reproductive success under similar stress levels. This ability is supported by several physiological and morphological mechanisms, including efficient stomatal regulation, high water-use efficiency associated with C4 photosynthesis, and rapid root system development that enhances water acquisition under limited moisture conditions (Ehleringer Reference Ehleringer1983). In addition, A. palmeri demonstrates strong reproductive plasticity, allowing it to maintain or reallocate biomass toward seed production even under water-limited environments. These adaptive traits differ from many crop species, which are typically bred for yield optimization under favorable conditions and often exhibit reduced physiological flexibility under stress. As a result, A. palmeri can persist in dryland systems and outcompete moisture-stressed crops, further complicating weed management efforts (Berger et al. Reference Berger, Ferrell, Rowland and Webster2015; Sandler and Gibson Reference Sandler and Gibson2019). Despite this, the effects of water stress on weed species, particularly in relation to timing and severity, have been underexplored in previous studies.
Earlier studies reported general growth responses of A. palmeri to moisture stress, including reduction in germination, biomass, and seed production under severe drought conditions (Chahal et al. Reference Chahal, Irmak, Jugulam and Jhala2018; Chandi et al. Reference Chandi, Jordan, York, Burton, Milla-Lewis, Spears, Whitaker and Wells2013; Paudel et al. Reference Paudel, Grantz, Vu and Shrestha2016). However, there is limited understanding of how the timing of water stress, specifically, the growth stage at which it is imposed, affects the plant’s physiological and reproductive outcomes. In many crop species, developmental stage plays a critical role in drought sensitivity, with early-season stress often affecting establishment and late-season stress influencing yield and reproduction (Ma et al. Reference Ma, Li, Wang, Li, Wang, Zhang, Zhen, Duan, Yan and Li2017). It is likely that similar patterns exist in weeds, but data specific to A. palmeri are lacking.
Understanding how A. palmeri responds to water stress at different growth stages is essential for several reasons. First, it can help identify windows of vulnerability that may be exploited through early-season weed control measures. Second, it provides insight into how environmental stressors may influence long-term weed population dynamics by affecting fecundity. Finally, it informs integrated weed management strategies, particularly in dryland production systems where nonchemical control methods and optimal timing of intervention are increasingly important due to herbicide resistance and resource constraints. Therefore, the objective of this study was to determine the effect of varying degrees of water stress applied at different growth stages on the growth and fecundity of A. palmeri under controlled greenhouse conditions.
Materials and Methods
Study Site
The pot experiment was conducted twice (two repetitions) in 2024 at the Horticultural Greenhouse of Texas Tech University, Lubbock, TX (33.584°N, 101.887°W). The average air temperature inside the greenhouse was maintained at 25 C during the day and 20 C at night during both the experiments. No additional lighting was introduced.
Plant Material and Experimental Design
The A. palmeri seeds used in this study were collected from Quaker Research Farm, Texas Tech University, Lubbock, TX, in 2022 and 2023. The seeds were stored at room temperature in brown paper bags before use. The seed germination percentage was determined before the experiment in Plant and Soil Science Department laboratory. Petri dishes were thoroughly cleaned before starting the experiment, and 20 A. palmeri seeds were placed in each petri dish, with three replications. The seeds were watered as needed. Germination percentage was recorded daily, starting from 4 d after seed placement and continuing until 15 d.
After the seed germination percentage was calculated, the seeds for the first and second experiments were planted on March 31, 2024, and May 6, 2024, respectively. Small black plastic pots (5- to 7-cm diameter and 8-cm height) were filled with soil, and 10 seeds were planted in each pot. When a couple of seedlings reached a height of 7 to 10 cm; they were transplanted to larger free-draining 11.4-L black plastic pot containing 7.5 kg of finely sieved soil collected from Quaker Research Farm, Texas Tech University, in February 2024. The soil was classified as Amarillo sandy clay loam (fine-loamy, mixed, super active, thermic Aridic Paleustoll) with 23% field capacity (FC) (Singh et al. Reference Singh, Singh, Parkash, Ritchie, Wallace and Deb2022) and particle size distribution consisted of 73.2% sand, 10.1% silt, and 16.7% clay (Parkash et al. Reference Parkash, Singh, Singh, Deb, Ritchie and Wallace2021). Pebbles were placed underneath each pot to facilitate drainage. To minimize transplant shock, the pots were watered for the next 7 d, and thinning was carried out before initiating water stress treatments. The experiment followed a split-plot design with five replications. The main factor was the growth stage at which stress was imposed, including the vegetative-stage stress (4- to 5-leaf stage) and reproductive stages stress (inflorescence emergence), while the subfactor was water stress levels: 100%, 80%, 60%, and 40% FC. For the vegetative-stage stress, water stress was initiated 7 d after transplanting (DATr) to allow plants to establish and recover from transplant shock, ensuring uniform growth and minimizing variability before stress imposition. Water stress treatments were maintained until approximately 50% of the plants reached the inflorescence stage, with irrigation applied according to the assigned FC levels. However, for reproductive-stage treatment, water stress was initiated when half of the plants reached inflorescence and continued until maturity. Plants not assigned to water stress treatments were maintained at 100% FC throughout the experiment. The experiment was repeated over time under the same greenhouse conditions previously described.
The FieldScout TDR-350 (Spectrum Technologies, Aurora, IL, USA) was used to monitor soil volumetric water content daily. The volumetric FC of the soil was 23%, and irrigation was applied to maintain soil moisture at target levels corresponding to 100% (23%), 80% (18.4%), 60% (13.8%), and 40% (9.2%) FC.
Data Collection
Growth parameters such as A. palmeri height and plant diameter were collected at biweekly intervals, starting from 2 wk after transplanting (WATr) until harvest. Plant height was measured using a standard measuring scale, and a vernier caliper was used to determine plant diameter. Stomatal conductance (GSW) and leaf temperature (T leaf) were recorded at 4 WATr and 8 WATr using a LI-600 Porometer/Fluorometer (LI-COR Biosciences, Lincoln, NE, USA). The aboveground biomass was recorded at the end of each experiment. Amaranthus palmeri plants were harvested when they reached 90% maturity and dried in an oven at 60 C for 48 h. The oven-dried plants were weighed using a calibrated weighing balance. After harvest, root samples from all the water stress levels were collected using a stainless-steel split-core sampler (diameter 5 cm), with one core taken per pot. The collected samples were analyzed for root parameters, such as root biomass and root length density (RLD) using the WinRHIZO Pro software version 2016a (Regent Instruments Inc., Quebec, Canada). Following harvest, seed germination percentage for water-stressed plants was determined by planting 20 seeds from female plants of each stress level in 200-cell germination trays (approximately 53 by 27 by 5 to 6 cm) filled with potting soil and maintained in the greenhouse. These trays were maintained at 25 C and watered daily to assess germination.
The germination percentage was calculated by using Equation 1:
 $\eqalign{\rm Germination\;percentage\;(\% ) = & {\rm \;Final\;number\;of\;seeds\;germinated} \; \cr & {\rm Total \; number \; of \; seeds}}$
$\eqalign{\rm Germination\;percentage\;(\% ) = & {\rm \;Final\;number\;of\;seeds\;germinated} \; \cr & {\rm Total \; number \; of \; seeds}}$
Statistical Analysis
The data from both pot experiments were analyzed using split-plot design ANOVA in R v. 3.5.2 (https://www.r-project.org), with the Agricolae package (v. 1.2-8). Data analysis was performed separately for each experiment due to a significant interaction between them. The parameter values presented in the tables and figures represent the mean values across A. palmeri growth stages and irrigation levels. Treatment means were compared using the LSD test at a 5% significance level. Figures were generated using SigmaPlot (v. 14; Systat Software, San Jose, CA, USA).
Results and Discussion
Effect of Water Stress on Plant Height and Stem Diameter
The experiment demonstrated how varying degrees of water stress affected the growth of A. palmeri over a period of 12 WATr. There were no significant differences in plant height among stressed plants at the vegetative and reproductive stages of development in either experiment (Figure 1A and 1C). Although plants subjected to reproductive-stage stress might be expected to be taller at early time intervals due to absence of initial water stress, similar absolute height values were observed across both treatments, with differences generally within 5% to 10% across observation times. The absence of growth stage–specific differences indicates that A. palmeri maintains comparable height responses when exposed to varying levels of water stress and recovers rapidly following early stress (Chahal et al. Reference Chahal, Irmak, Jugulam and Jhala2018). The consistent height response across treatments indicates a strong capacity for growth maintenance (physiological tolerance) rather than stage-specific plasticity. As a C4 species adapted to arid and semiarid environments, A. palmeri can sustain carbon assimilation and cellular expansion under water-limited conditions, resulting in similar height responses across developmental stages (Ward et al. Reference Ward, Webster and Steckel2013). In contrast, long-term field studies in corn have shown that water stress imposed at different developmental stages can differentially affect plant growth and yield (Cakir Reference Cakir2004).
Effect of growth stage–specific water stress on height of Amaranthus palmeri. (A) First and (C) second experiments comparing plant height during vegetative and reproductive stages. (B) First and (D) second experiments showing the effect of different irrigation levels (100%, 80%, 60%, and 40% field capacity [FC]) on plant height. Asterisks (*) indicate significant differences (P < 0.05) among treatments at respective time points. WATr, weeks after transplanting. Vertical arrows indicate the timing of the shift from vegetative-stage to reproductive-stage water stress.

As the water stress levels increased, the plant’s ability to grow was significantly reduced (Figure 1B and 1D). In the first experiment, at 2, 4, 6, and 8 WATr, plants grown under 100% FC consistently showed significantly greater plant height compared with those under 80%, 60%, and 40% FC (Figure 1B). At 10 WATr, plants at 100% FC were approximately 17% taller than those at 40% FC and about 8% to 10% taller than those at 60% and 80% FC, respectively. Although water was not limited, plant height under 100% FC showed no significant increase from 8 until 10 WATr (harvest), indicating maximum height was attained before harvest (Figure 1B). This response suggests that after the plants reach maximum height, further water availability does not promote additional shoot elongation, because assimilates are preferentially diverted to reproductive tissues rather than continued stem growth under favorable moisture conditions (Bazzaz et al. Reference Bazzaz, Ackerly, Reekie and Fenner2000). In the second experiment, plants subjected to 100% FC showed significantly greater plant height compared with those under 40% FC at various time intervals (Figure 1D). Also, plants subjected to 80% and 60% FC had significantly greater plant height compared with 40% FC at 6, 8, and 10 WATr. This significant difference is attributed to reduced water availability, which ultimately impacts overall plant growth by disrupting key metabolic processes. Similarly, increasing water stress has been shown to significantly reduce the growth potential and height of waterhemp [Amaranthus tuberculatus (Moq.) Sauer], a closely related C4 weed species (Chauhan Reference Chauhan2013; Sarangi et al. Reference Sarangi, Irmak, Lindquist, Knezevic and Jhala2016). Overall, these results demonstrate that A. palmeri growth is progressively limited by increasing water stress, with more significant effects under severe (40% FC) deficit conditions.
The effects of varying water stress levels on stem diameter were evaluated across two experiments. In the first experiment, growth stage did not significantly affect stem diameter (Figure 2A), with differences between treatments remaining within ∼25% at early stages (e.g., 2 WATr: 3.8 vs. 3.0 mm) and diminishing to less than 5% by 10 WAT (7.0 to 7.3 mm). In contrast, irrigation level significantly influenced stem diameter, with plants at 100% FC exhibiting greater stem diameter than those at 80%, 60%, and 40% FC at 2 WATr (Figure 2B). At this stage, plants at 100% FC (∼4.2 mm) had stem diameters approximately 70% greater than those at 40% FC (∼2.5 mm). Plant growth is mostly affected by water limitation, and changes in stem diameter have been found to be a sensitive indicator of water stress (Fernández and Cuevas Reference Fernández and Cuevas2010). However, by 4 and 10 WATr, the stem diameters increased across all treatments despite reductions in irrigation levels, and stem diameter was comparable across all treatments by harvest (Figure 2B). This finding shows the early growth sensitivity of A. palmeri to water deficit and reveals that it was able to compensate for early water stress later in development, which might be due to physiological adaptation or resource reallocation under stress. Similar compensatory mechanisms have been reported in drought-tolerant species that can rapidly adjust osmotic balance and maintain growth once stress increased (Ward et al. Reference Ward, Webster and Steckel2013). In the second experiment, similar trends were observed with respect to growth-stage stress, where no significant differences were detected between vegetative and reproductive stages at any time point (Figure 2C). Both stages followed a comparable pattern of increasing from 2 WATr to 10 WATr. Conversely, at 2 WATr, plants grown under 100% FC exhibited significantly greater stem diameter than those subjected to reduced irrigation (Figure 2D). However, from 4 WATr until harvest, stem diameter was significantly greater under moderate water stress (60% and 80% FC) compared with fully irrigated plants. This response reflects morphological plasticity in A. palmeri, whereby moderate water limitation promotes greater allocation to stem thickening rather than continued vertical growth, enhancing mechanical support under fluctuating soil moisture conditions (Bazzaz et al. Reference Bazzaz, Ackerly, Reekie and Fenner2000; Poorter et al. Reference Poorter, Niklas, Reich, Oleksyn, Poot and Mommer2012). Amaranthus palmeri, a C4 species adapted to arid and semiarid environments, can maintain cell turgor and carbon assimilation under moderate water stress through osmotic adjustment, allowing continued cell expansion and secondary growth even when soil moisture is reduced. This physiological capacity likely contributed to increased stem diameter under water-limited conditions relative to fully irrigated plants (Ward et al. Reference Ward, Webster and Steckel2013; Watts and Tenhumberg Reference Watts and Tenhumberg2021).
Effect of growth stage–specific water stress on stem diameter of Amaranthus palmeri. (A) First and (C) second experiments comparing stem diameter during vegetative and reproductive stages. (B) First and (D) second experiments showing the effect of different irrigation levels (100%, 80%, 60%, and 40% field capacity [FC]) on stem diameter. Asterisks (*) indicate significant differences (P < 0.05) among treatments at respective time points. WATr, weeks after transplanting. Vertical arrows indicate the timing of the shift from vegetative-stage to reproductive-stage water stress.

Effect of Water Stress on Stomatal Conductance and Leaf Temperature
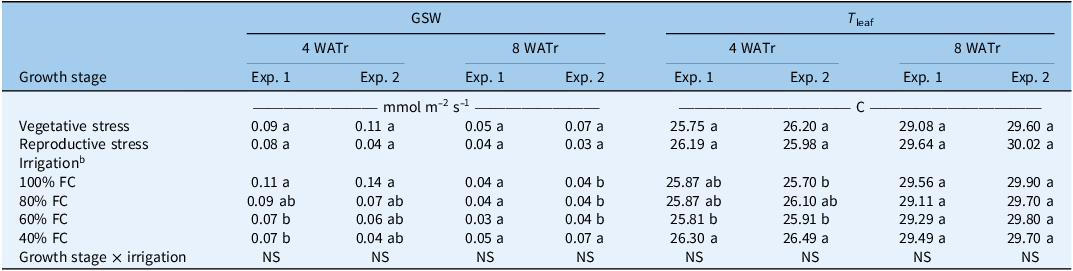
The effects of growth stage and irrigation levels on GSW and T leaf are presented in (Table 1). There were no significant differences in GSW and T leaf between vegetative and reproductive stress treatments at 4 and 8 WATr in either experiment. These results may suggest that A. palmeri maintained physiological stability across developmental stages, potentially due to its inherent low-water or drought tolerance. Cominelli and Patrignani (Reference Cominelli and Patrignani2022) also reported that A. palmeri sustained gas exchange and photosynthetic activity through osmotic adjustment and regulated stomatal behavior, even under declining soil moisture conditions.
Effect of water stress on stomatal conductance (GSW) and leaf temperature (T leaf) of Amaranthus palmeri in a greenhouse experiment conducted in Lubbock, TX. a

a Means followed by the same letter within each column and factor (growth stage or irrigation) are not significantly different according to Least Significant Difference (LSD) test at P ≤ 0.05. WATr, weeks after transplanting.
b FC, field capacity.
At 4 WATr, GSW was the highest under 100% field capacity (FC) in both experiments, whereas the lowest GSW values were observed under 40% FC, demonstrating a clear decline in stomatal conductance with increasing water deficit. By 8 WATr, GSW values were generally lower than those observed at 4 WATr across all treatments. In both experiments, relatively higher GSW values compared with other treatments were observed at 40% FC at 8 WATr, suggesting physiological acclimation to prolonged water stress, potentially through osmotic adjustment and partial stomatal reopening rather than improved plant water status. In the second experiment, GSW at 40% FC (0.07) was approximately 75% higher than at 100% FC (0.04) at 8 WATr. Similar stomatal regulation responses under extended water stress, including partial stomatal closure and acclimation of photosynthetic processes, have been reported in drought-tolerant weed species, including A. palmeri (Cominelli and Patrignani Reference Cominelli and Patrignani2022; Flexas et al. Reference Flexas, Bota, Cifre, Mariano Escalona, Galmés, Gulías, Martínez-Cañellas, Moreno and Ribas-Carbó2004; Ward et al. Reference Ward, Webster and Steckel2013).
T leaf did not exhibit a consistent response to irrigation levels across sampling times. At 4 WATr, T leaf values were approximately 26 C across treatments in both experiments, with plants under 40% FC exhibiting slightly higher leaf temperatures, likely due to reduced transpirational cooling under water-limited conditions with differences of approximately 0.4 to 0.6 C (∼1.5% to 2.3%) higher compared with well-watered treatments. At 8 WATr, T leaf values were relatively uniform across irrigation treatments, between 29 to 30 C in both experiments, indicating effective thermal regulation at later growth stages. Collectively, these results demonstrate that leaf temperature in A. palmeri is more sensitive to water stress at early growth stages than at later stages, reflecting increased physiological adaptability to water-limited environments as plants mature.
Effect of Water Stress on Plant Root Parameters
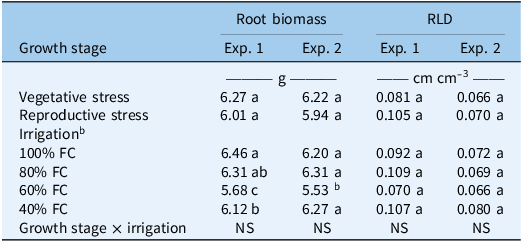
The water stress treatments did not have a significant effect on the root biomass and root length density at either vegetative or reproductive stage in both experiments (Table 2). However, irrigation level significantly influenced root biomass, with root biomass significantly reduced at 60% FC compared with other irrigation levels. In the first experiment, root biomass at 60% FC (5.68 g) was approximately 12% lower than at 100% FC (6.46 g), and in the second experiment, root biomass at 60% FC (5.53 g) was about 11% lower than at 100% FC (6.20 g). This enhanced root biomass under optimal moisture conditions is likely due to active cell division and elongation. Similar results were reported by Vaughn et al. (Reference Vaughn, Bernards, Arkebauer and Lindquist2016), who observed that the root biomass of corn and velvetleaf (Abutilon theophrasti Medik.) was maximized under conditions of maximum water availability. Higher root biomass under 100% and 80% FC suggests that adequate soil moisture supports root growth by facilitating cell expansion, carbon allocation, and nutrient uptake (Lynch Reference Lynch2007). In contrast, the reduction in root biomass at 60% FC indicates that moderate water stress restricted carbon assimilation and root development, likely as a result of reduced photosynthetic activity and limited assimilate availability for belowground growth (Farooq et al. Reference Farooq, Wahid, Kobayashi, Fujita and Basra2009). Interestingly, root biomass under 40% FC was comparable to that observed under well-watered conditions, particularly in the second experiment. This pattern may reflect an adaptive response to severe water limitation, whereby plants maintain or prioritize root investment to enhance soil exploration and water acquisition. Similar adaptive responses have been documented in both crops and weed species, where drought stress promotes root persistence or changes in root architecture rather than a uniform reduction in root biomass (Comas et al. Reference Comas, Becker, Cruz, Byrne and Dierig2013).
Effect of water stress on root biomass and root length density (RLD) of Amaranthus palmeri in a greenhouse experiment conducted in Lubbock, TX. a

a Means followed by the same letter within each column and factor (growth stage or irrigation) are not significantly different according to Least Significant Difference (LSD) test at P ≤ 0.05.
b FC, field capacity.
RLD did not show any significant difference among the various water stress treatments in both experiments (Table 2). In the first experiment, RLD values were numerically greater at 80% FC (0.109 cm cm⁻3), which was approximately 33% higher than at 60% FC (0.070), followed by 40% FC. Similarly, in the second experiment, the highest RLD values were observed at 40% FC (0.080), which was approximately 21% higher than at 60% FC (0.066). However, these differences were not statistically significant. This trend could be a response to water limitation that enables the plant to increase its soil exploration and improve water uptake efficiency. Such adaptive responses are consistent with the findings of Kalra et al. (Reference Kalra, Goel and Elias2024), who reported that roots commonly modify their architecture under drought to ensure maximum resource capture at reduced metabolic cost. As drought progresses, plants often allocate greater biomass or elongation capacity to deeper root zones, facilitating water uptake from subsurface soil layers. Similarly, in the second experiment, the highest RLD values were observed at 40% FC, further suggesting a tendency toward increased root elongation under severe water limitation. However, these trends were not statistically significant. In contrary, Sampathkumar et al. (Reference Sampathkumar, Pandian and Mahimairaja2012) showed that mild water stress promotes lateral root and biomass, whereas severe stress limits root spread and reduces root mass. Their observation of uneven soil moisture under deficit irrigation emphasizes the importance of root plasticity to access water resources, consistent with the root elongation observed here under severe stress (40% FC).
Effect of Water Stress on Plant Biomass, Seed Production, and Seed Germination
Plant Biomass
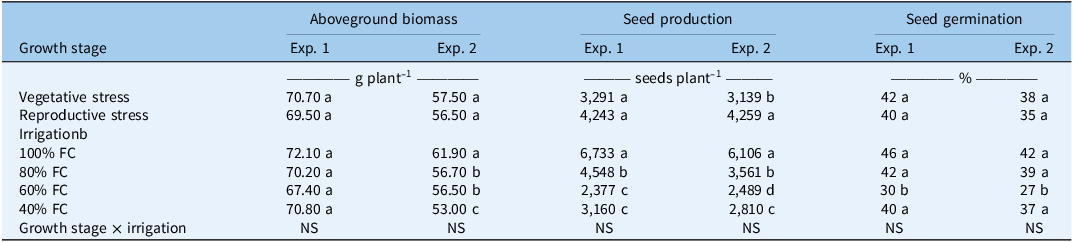
The effects of water stress on the final aboveground biomass, seed count, and seed germination of A. palmeri were evaluated in two greenhouse experiments under varying degrees of water stress levels at various growth stages (Table 3). Biomass is an important indicator of plant growth that showed no significant differences at vegetative and reproductive growth stage under water stress conditions in both experiments (Table 3). In the first experiment, the final aboveground biomass at vegetative stress was 70.7 g, which was approximately 2% higher compared with 69.5 g at reproductive stress. This finding demonstrated that A. palmeri may have the ability to sustain biomass production across different growth stages even under stress. It may be due to its inherent C4 drought-tolerance mechanisms (Chandi et al. Reference Chandi, Jordan, York, Burton, Milla-Lewis, Spears, Whitaker and Wells2013). Similarly, in the second experiment, the difference between vegetative and reproductive stress was less than 2% (57.5 vs. 56.5 g), indicating no significant effect of growth stage. This lack of significant differences in biomass under varying growth stages may indicate a stable physiological response to stress, although slight fluctuations between the two experiments could be attributed to inherent experimental variability or environmental conditions in the greenhouse.
Effect of water stress on the final aboveground biomass, seed production, and seed germination of Amaranthus palmeri in a greenhouse experiment conducted in Lubbock, TX.a

a Means followed by the same letter within each column and factor (growth stage or irrigation) are not significantly different according to Least Significant Difference (LSD) test at P ≤ 0.05.
b FC, field capacity.
In the first experiment, biomass did not differ significantly among irrigation treatments; however, values were numerically greater at 100% FC and appeared to decrease at 80%, 60%, and 40% FC. In the second experiment, biomass followed a declining trend with increasing water stress. The highest biomass was recorded at 100% FC, followed by a notable decline at 80% and 60% FC, dropping further at 40% FC, representing an approximate 14% decrease under severe water stress. This trend indicates greater sensitivity to reduced irrigation levels compared with the first experiment. These results revealed that A. palmeri can tolerate a moderate decrease in water availability, but the reduction in biomass becomes more distinct under severe stress (Chahal et al. Reference Chahal, Irmak, Jugulam and Jhala2018).
Seed Production
Seed count, which is a direct measure of reproductive success, demonstrated significant variation in response to both growth-stage stress and irrigation levels (Table 3). In both experiments, when stress was imposed on the plants during the reproductive growth stage, they produced ∼29% higher numbers of seeds than plants who were given stress during vegetative growth stage. Amaranthus palmeri produced the highest number of seeds at 100% FC, followed by 80%, 40%, and 60% FC in both experiments. Seed production at 100% FC was approximately 48% higher than at 80% FC and nearly three times greater than at 60% FC in the first experiment, and 72% higher than at 80% FC and more than two times greater than at 60% FC in the second experiment. This reduction is likely due to fewer flowers and seed sets resulting from limited water availability for growth and development under water stress (Höfer et al. Reference Höfer, Lindner, Ayasse and Kuppler2023). Plants under 60% FC produced significantly lower number of seeds in both experiments compared with 40% FC. Even under severe water stress or drought conditions, A. palmeri produced a considerable number of seeds in both experiments. This highlights that water stress alone is insufficient to limit its reproductive potential and emphasizes the importance of effective early-season management to minimize weed pressure, reduce seedbank replenishment, and prevent crop growth and yield losses at harvest (Legleiter and Johnson Reference Legleiter and Johnson2013). Similarly, Mausbach et al. (Reference Mausbach, Irmak, Chahal, Sarangi and Jhala2022) reported that A. theophrasti produced more seeds under 75% FC than under 100% FC, indicating greater reproductive success under moderate water limitation and demonstrating relatively high resilience to drought conditions. In contrast, other weed species, including prickly lettuce (Lactuca serriola L.), silverleaf nightshade (Solanum elaegnifolium Cav.), A. tuberculatus, and A. palmeri, exhibit reduced seed production as water stress levels increase (Singh, et al. Reference Singh, Thapa, Kukal, Irmak, Mirsky and Jhala2022).
Seed Germination
Seed germination is a key indicator of seed viability and successful plant establishment. Across both experiments, germination rates did not differ between growth stages, yet they showed clear and significant differences under varying irrigation treatments. The highest germination was observed at 100% FC, with 46% in the first experiment and 42% in the second experiment. The results showed that optimal soil moisture is crucial for successful seedling emergence, which is supported by the evidence that seed germination is influenced by environmental factors such as soil moisture, temperature, and so on (Sidhu et al. Reference Sidhu, Dhaliwal and Gaikwad2024). At 80% FC, seed germination was 42% and 39% in the first and second experiments, respectively. Although these values were numerically lower than those observed at 100% FC, the differences were not statistically significant, suggesting that germination under moderate water availability was comparable to that at full field capacity. However, the lowest germination rates were recorded at 60% FC, with 30% and 27% germination in the first and second experiments, respectively. This significant reduction in seed germination under 60% FC suggests that moderate water stress adversely affects seed quality. Water stress during seed filling has been shown to disrupt assimilate translocation, hormonal balance, and seed reserve deposition, ultimately reducing germination percentage (Abdelhamied et al. Reference Abdelhamied, Kandil, El-Badry and Taha2021; Khaeim et al. Reference Khaeim, Kende, Balla, Gyuricza, Eser and Tarnawa2022). Similar declines in germination under reduced water availability have been documented in rice (Oryza sativa L.) and wheat (Triticum aestivum L.), where limited water supply impaired seed imbibition and early metabolic activity (Ali et al. Reference Ali, Perveen, Saeed, Manzoor, Ali, Hussain and Ahmad2024).
Interestingly, seed germination under 40% FC was comparable to higher-irrigation treatments, suggesting a potential stress-threshold response. Severe water limitation may reduce total seed production but allow the remaining seeds to complete development with relatively preserved viability. Such maternal stress effects, wherein drought alters seed traits and germination behavior, have been widely reported across crop and weed species (Matzrafi et al. Reference Matzrafi, Osipitan, Ohadi and Mesgaran2021). These responses may represent an adaptive strategy enabling seeds to maintain germination capacity under adverse environmental conditions.
Therefore, this study provides important information on the growth and reproductive responses of A. palmeri under varying degrees of water stress during different growth stages, offering valuable insights for weed management in water-limited agricultural systems. The findings signify the weed’s ability to maintain high seed production and germination rates even under severe drought conditions, emphasizing the importance of early-season control measures to prevent seedbank replenishment and future infestations. Also, targeting A. palmeri during its vegetative growth stage, when it is more vulnerable to water stress, could enhance the effectiveness of management strategies. Moreover, the study underlines the need for integrated weed management approaches that combine chemical, mechanical, and cultural practices to address the challenges posed by A. palmeri’s drought tolerance and herbicide resistance. In dryland cropping systems, timely interventions during critical growth stages offer an effective approach to suppress weed growth while maximizing water-use efficiency. Furthermore, understanding the weed’s adaptive mechanisms, such as root elongation under drought, can inform the development of strategies to limit its competitive advantage over crops. For growers in semiarid and arid regions, our research shows the importance of monitoring soil moisture levels and implementing proactive weed control measures to minimize crop yield losses. By utilizing the knowledge from our study, agricultural stakeholders can develop sustainable practices to mitigate the economic and ecological impacts of A. palmeri in water-scarce environments.
Acknowledgments
The authors gratefully acknowledge the Davis College of Agricultural Sciences & Natural Resources for providing the financial support necessary to conduct this research. Appreciation is extended to the Department of Plant and Soil Science at Texas Tech University for their continued support. Finally, we sincerely thank Vikram Baliga, Daniel Mayer, and the entire greenhouse team for their dedicated support and assistance throughout the study.
Funding statement
This study was supported by Catalyst Grant, Davis College of Agricultural Sciences & Natural Resources, Texas Tech University, USA.
Competing interests
The authors declare no conflicts of interest.



 Open access
Open access