


Non-technical Summary
This work describes and illustrates Ediacaran (latest Precambrian) and Cambrian body and trace fossils from Nevada and California and discusses their significance for understanding the early evolution of animals and for locating the Ediacaran–Cambrian boundary (about 539 million years ago) in sedimentary deposits of the southwestern Great Basin. The most distinctive body fossil is a new air-bed-like organism formed of tubular modules arranged lengthwise. At the growing ends, the tubular modules separated from each other and tapered to blunt points. How these organisms lived and functioned remain uncertain, partly because the fossils are found in deep water deposits far from where they lived. We also deal with another distinctive fossil, ‘Plagiogmus’, which was found during the 1960s, but largely ignored or misinterpreted since then. ‘Plagiogmus’ is the complex, often meandering burrow of a sizeable slug-like animal that first appeared in earliest Cambrian time. Its association in time with an important geochemical event suggests that the Ediacaran–Cambrian boundary in the Great Basin is older than previously thought.
Introduction
This work describes and illustrates Ediacaran (latest Precambrian) and Cambrian body and trace fossils from Nevada and California and discusses their significance for understanding the early evolution of animals and for locating the Ediacaran–Cambrian boundary in sedimentary deposits of the southwestern Great Basin. We first recount the discovery of Ediacaran body fossils in the craton margin succession of the Death Valley area, and then briefly discuss previous biostratigraphic work on the Precambrian–Cambrian boundary in both the Death Valley and White Inyo regions of California and Nevada and (Figs. 1–4). In following sections, we describe and illustrate the fossils (Figs. 4–12), summarize the results of this investigation, and conclude with a brief discussion of the significance of this work for Ediacaran paleobiology and the age of the eon boundary.
Locality maps. (1) Locations of fossiliferous sections of the middle (Esmeralda) member of the Deep Spring Formation (Mollie Gibson, Mount Dunfee), Wyman Formation (Hines Ridge), and the lower member of the Wood Canyon Formation (Desert Range, Johnnie, Spring Mountains, Chicago Pass) in California and Nevada. (2) Google Maps satellite image of the northern end of the Montgomery Mountains near Johnnie, Nevada, with UCLA localities 7333 and 7334 indicated by markers, near the bases of stratigraphic sections (Fig. 3) measured along the ridges extending to the ESE; RJH is Horodyski's discovery site (Fig. 4.3); 17130 refers to the location of LACNHM locality 17130 (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000, p. 359); WCF is Wood Canyon Formation and CF is Carrara Formation; as indicated, the investigated section has an average dip of about 40° SE. (3) Part of U.S.G.S. Geological Quadrangle Map Blanco Mountain (GQ-529; Nelson, Reference Nelson1966), republished with permission, showing UCLA fossil localities 6160 and 6153 in the Esmeralda (middle) Member (dm) of the Deep Spring Formation near the Mollie Gibson mines, White-Inyo Mountains. (4) Part of Nelson and Smith's (Reference Nelson and Smith2019, DR-fig. 1) geological map of Mount Dunfee, Nevada, republished with permission, showing localities/sections (white rectangles) in the Esmeralda Member (DSm) of the Deep Spring Formation that have yielded Psammichnites gigas arcuatus (Fig. 12). Ru (ru) = upper member of the Reed Dolomite; rh = Hines Tongue of the Reed Dolomite; du = upper (Gold Point) member of the Deep Springs Formation; dl = lower (Dunfee) member of the Deep Springs Formation; DSl = Dunfee Member of the Deep Springs Formation; Dsu = Gold Point Member of the Deep Springs Formation; Rht = Hines Tongue of the Reed Dolomite.

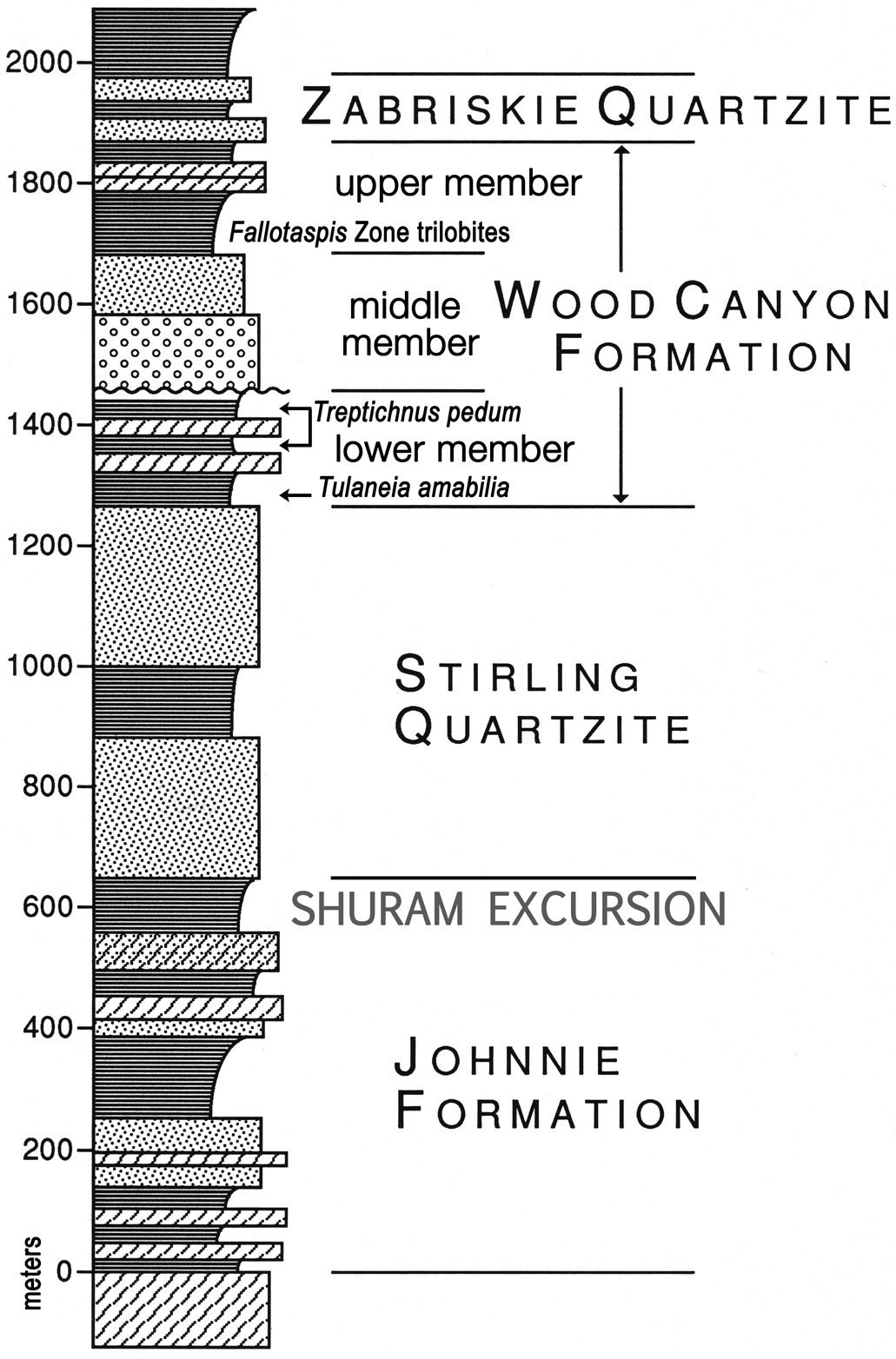
Summary of the stratigraphy enclosing the section of interest in the lower member of the Wood Canyon Formation, with the positions of the Shuram negative carbon isotope excursion and the oldest occurrence of Cambrian Stage 3 trilobites (Fallotaspis Zone trilobites) from Bergmann et al. (Reference Bergmann, Zentmyer and Fischer2011) and Hollingsworth (Reference Hollingsworth, Hollingsworth, Sundberg and Foster2011), respectively. The unconformity below the conglomeratic middle member of the Wood Canyon Formation is the base of the craton-wide Sauk megasequence (Sloss, Reference Sloss1963, Reference Sloss1988; Runnegar, Reference Runnegar1998; Keller et al., Reference Keller, Lehnert, Cooper, Derby, Fritz, Longacre, Morgan and Sternbach2012). Level of Tulaneia amabilia n. gen. n. sp. indicated in lower part of lower member, Wood Canyon Formation.
Cartoon representation of three sections measured by JGG through the lower member of the Wood Canyon Formation, simplified to emphasize major features of their lithostratigraphy and biostratigraphy and normalized to the top of the Stirling Quartzite; the three principal dolostone units are numbered 1, 2, and 3 and details are tabulated in Supplemental dataset 1; see text for explanation of ‘Treptichnus spectrum traces.’ Only some of the upward-fining parasequences are shown, but there are clearly more than the canonical ‘three parasequences’ (Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000). The occurrences of Tulaneia n. gen. and Treptichnus shown in gray need further verification.

Outcrops (1–4) and annulated tubular body fossils (5–8). (1) Lower part of measured section at UCLA 7333 (Fig. 1.2) with RJH in yellow jacket standing on the dip slope at the top of the Stirling Quartzite; arrow indicates the outcropping edge of the first major dolostone (1, Fig. 3); (2) middle of the measured section at Chicago Pass at UCLA 7329 (Fig. 1.2), with JGG and RJH standing at the base of the second major dolostone (2, Fig. 3); the third major dolostone is visible in the distance; (3) Bob Horodyski, sitting at his discovery site in the Montgomery Mountains (RJH, Fig. 1.2) in 1993; the piles of slabs beside the pack and behind his head are two of several cairns that were constructed from pieces of float searched for fossils and now serve as permanent markers for the discovery site; (4) fluvial pebbly sandstone of the middle member of the Wood Canyon Formation being examined by RJH in the Nopah Range, south of Chicago Pass. (5–8) Poorly preserved tubular body fossils that retain some traces of closely spaced annulations (5, 6) and show some evidence for wall flexibility or fragility (7, 8), from beneath the first major dolostone of the lower member of the Wood Canyon Formation in the Spring Mountains (5, LACMIP 43368.1) and Montgomery Mountains (6, LACMIP 43369.13; 7, LACMIP 43369.4; 8, LACMIP 43369.12). All scale bars = 1 cm.

Bob Horodyski (RJH) was prospecting for Precambrian fossils in the craton margin succession of the Death Valley area when he investigated the northern end of the Montgomery Mountains, near the derelict mining town of Johnnie, Nevada, early in 1991. He found a slab of sandstone (Fig. 5.2, B), which had two spectacular fossils—one on each side—that were novel to the succession and clearly not of Cambrian age. After returning to Tulane University, where he photographed the slab, he sent prints to BR and asked for advice as he was planning to submit an abstract for the annual meeting of Geological Society of America (GSA) in San Diego, California. BR enthusiastically (but erroneously) identified the fossils as Ernietta Pflug (Reference Pflug1966) in a fax sent to RJH on June 24, 1991, and drew his attention to Richard Jenkins’ re-analysis of Pflug's (Reference Pflug1972) material (Jenkins et al., Reference Jenkins, Plummer and Moriarty1981). The abstract was submitted and published (Horodyski, Reference Horodyski1991) and the discovery specimens and others collected during the summer of 1991 were displayed at a poster session in San Diego in October (McMenamin, Reference McMenamin1998, p. 30). The discovery slab had obviously not moved far from outcrop, so RJH began a systematic search of the site that took several days. He painstakingly examined each and every loose piece of rock in the vicinity of his discovery and built cairns (Fig. 4.3) from the pieces that lacked fossils. By August 1991, he had assembled the small collection of internal molds (parts) and external molds (counterparts) that now become the holotype and paratypes of the new genus and species Tulaneia amabilia Runnegar and Horodyski (Figs. 5–8).
Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp., lowest part of lower member of the Wood Canyon Formation, RJH discovery site (Figs. 1.2, 4.3), Montgomery Mountains, Nevada. (1) Distal region of one of two specimens occurring on the surfaces of three small slabs (A–C) that fit together (2); (2) views of the edges and surfaces of three small slabs (A–C), found separately, but fit together and preserve parts and counterparts of the two best specimens: the holotype (1, 2A, 2B; LACMIP 43369.1A, LACMIP 43369.1B) and paratype (2B, 2C, 5, 6; LACMIP 43369.2A, LACMIP 43369.2B); (3) distal and proximal ends of two specimens, the larger of which is the counterpart of paratype LACMIP 43369.3B; (4) paratype LACMIP 43369.3A (part); (5, 6) broken edge of paratype LACMIP 43369.2A (part) showing clean sandstone filling tubular modules. Scale bars = 1 cm (1), 3 cm (3–6); 3-cm-grid squares (2).

Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp.: (1–7) Lowest part of lower member of the Wood Canyon Formation, RJH discovery site (Figs. 1.2, 4.3), Montgomery Mountains, Nevada, and Pteridinium carolinaensis (St Jean, Reference St. Jean1973); (8–10), Nasep Member, Urusis Formation, UCLA 7322, Swartkloofberg farm, Namibia. (1) Front and side views of the holotype showing biplanar symmetry, clean quartzite filling, and absence of fill at the mid-height constriction, LACMIP 43369.1B (part); (2) two similar views of the distal end of paratype LACMIP 43369.2A (part) (also shown in Fig. 5.2B, 5.2C, 5.5, 5.6) at different focal planes to show the closely packed nature of the quartzite grains filling the modules; (3) oblique view of same specimen to show the parabolic cross section of the proximal end and the way the modules taper in thickness distally; (4, 5), two similar specimens on the same slab: (4) the larger specimen, paratype LACMIP 43369.3, is illustrated in proximal view in (5) to show the flat iron-like termination of one module (black arrow) and the planar cracks that follow the originally organic module walls (white arrows); (6, 7) part of paratype LACMIP 43369.3A (also shown in Fig. 5.4) showing views of the lateral surface and broken edge (part); (8–10) lateral and edgewise views of a quartz sandstone mold of a fragmentary specimen of Pteridinium carolinaensis, which preserves two of the three vanes, to illustrate a superficial similarity to Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp. and a fundamental difference, in that there are no transverse partitions between the two vanes of Pteridinium (9), GSN F 1997. All scale bars = 2 cm.

In the GSA abstract, the stratigraphic setting of RJH's discovery was described as “near the contact of the Wood Canyon Formation and the Stirling Quartzite” (Horodyski, Reference Horodyski1991, p. A163). Subsequent fieldwork allowed RJH to source the float specimens to the lowest 10–15 m of the Wood Canyon Formation, to measure a stratigraphic section along an adjacent ridge (Figs. 1.2, 4.1), and to find Cambrian trace fossils, including unquestionable specimens of Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955), in the upper part of the lower member of the Wood Canyon Formation (LMWCF; Fig. 10.2, Supplemental dataset 1). Follow-up fieldwork by RJH and the other authors in January and March–April 1993 and March 1995 gave rise to two other GSA abstracts (Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994; Runnegar, Reference Runnegar1998), but publication of the evidence for these statements is long overdue. This article attempts to remedy that situation.
Geological setting
Lithostratigraphy and geochronology
The stratigraphic nomenclature for the Proterozoic–Cambrian succession of the Death Valley region established by Stewart (Reference Stewart1966, Reference Stewart1970) has persisted with little change (Prave et al., Reference Prave, Fedo, Cooper, Walawender and Hanan1991; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023). The sections of interest lie between the top of the Stirling Quartzite and the regional unconformity at the base of the middle member of the Wood Canyon Formation near Johnnie in the Montgomery Mountains, Nye County, Nevada; at Chicago Pass in the Nopah Range, Inyo County, California; south of Wood Canyon in the Spring Mountains, Nye County, Nevada; and in the central Desert Range, Desert National Wildlife Range, Clark County, Nevada (Figs. 1.1, 1.2, 2–4, Appendix 1).
The Stirling Quartzite and the middle member of the Wood Canyon Formation were both deposited during sea level lowstands in shoreface and fluvial environments whereas the intervening LMWCF was formed in deeper marine environments that accumulated micaceous siltstones, hummocky cross-stratified sandstones, and subtidal to peritidal dolostones (Diehl, Reference Diehl1976; Fedo and Cooper, Reference Fedo and Cooper1990, Reference Fedo and Cooper2001; Muhlbauer et al., Reference Muhlbauer, Fedo and Moersch2020; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). The sequence stratigraphy of the ‘Death Valley facies’ (Nelson, Reference Nelson1978) has been extensively investigated and there is consensus that the disconformity at the base of the middle member of the Wood canyon Formation is the base of the Sauk megasequence (Sloss, Reference Sloss1963, Reference Sloss1988), at least where that impinges upon the craton (Fedo and Cooper, Reference Fedo and Cooper2001; Hogan et al., Reference Hogan, Fedo and Cooper2011; Keller et al., Reference Keller, Lehnert, Cooper, Derby, Fritz, Longacre, Morgan and Sternbach2012). The lower member sits on an older sequence surface at the top of the Stirling Quartzite and is composed of several upward-shallowing parasequences (Fig. 3), three of which are capped by dolostones and have been widely used for correlation within the member (Stewart, Reference Stewart1966; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). Our compilation of the biostratigraphy is consistent with this approach (Supplemental dataset 1; but see Hogan et al., Reference Hogan, Fedo and Cooper2011).
In the coeval succession to the north, the ‘White-Inyo facies’ (Nelson, Reference Nelson1978), the equivalent units are the lower (Dunfee), middle (Esmeralda), and upper (Gold Point) members of the Deep Spring Formation, plus the underlying Reed Dolomite (Fig. 1; Cloud and Nelson, Reference Cloud and Nelson1966; Alpert, Reference Alpert1974, Reference Alpert1975, Reference Alpert1976; Signor et al., Reference Signor, Mount and Onken1987; Ahn et al., Reference Ahn, Babcock and Hollingsworth2012; Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016). Stewart's (Reference Stewart1966) long-standing lithological correlation of the Deep Spring Formation with the LMWCF has been supported by more recent geochemical and paleontological evidence (Corsetti and Kaufman, Reference Corsetti and Kauffman1994; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023).
Biostratigraphy
Horodyski's (Reference Horodyski1991) report of the discovery of the Ediacaran fossil Ernietta near the contact of the Wood Canyon Formation and Stirling Quartzite placed the Precambrian–Cambrian boundary within the lower part of the Wood Canyon Formation, and the subsequent discovery of Treptichnus pedum in the upper part of the lower member (Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000; Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002; O'Neil et al., Reference O'Neil, Tackett and Meyer2022) confined that transition to at or below the dolomite capping the second major parasequence of the lower member (Fig. 3; Supplemental dataset 1; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002). Although our reassignment of Ernietta to the new genus Tulaneia weakens the argument for an Ediacaran age for the basal part of the lower member, Tulaneia Runnegar and Horodyski n. gen. occurs with tubular body fossils (Fig. 4.5–4.8) that elsewhere in the LMWCF and in the Deep Spring Formation indicate an Ediacaran rather than a Cambrian age (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000; Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016, Reference Smith, Nelson, Tweedt, Zeng and Workman2017; Selly et al., Reference Selly, Schiffbauerb, Jacquet, Smith and Nelson2020). Furthermore, Tulaneia n. gen. is clearly an erniettomorph and therefore, by default, should be regarded as Ediacaran in age.
Plagiogmus arcuatus Roedel, Reference Roedel1929, which now is considered a subjective junior synonym of Psammichnites gigas (Torell, Reference Torell1868), is a distinctive, complex burrow system that is found in Fortunian and younger Cambrian strata in many parts of the world (Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022). It was first reported as ‘cf. Pteridinium’ from the middle (Esmeralda) member of the Deep Spring Formation in the White-Inyo mountains (Fig. 12.3; Cloud and Nelson, Reference Cloud and Nelson1966, Reference Cloud and Nelson1967), where it had been found by a graduate student named Cyril Gallick, who mapped an area immediately northwest of the discovery site (Gallick, Reference Gallick1964). Soon afterwards, Glaessner (Reference Glaessner1969, p. 390) questioned the proposed similarity to Pteridinium and suggested that Gallick's fossil could be a specimen of Plagiogmus that was “preserved in a thinner-bedded and more compacted rock and hence lack[s] the depth of tracks preserved in more massive and less compressible sandstone.” Cloud and Bever (Reference Cloud and Bever1973) accepted this suggestion and re-illustrated Gallick's specimen together with examples of Plagiogmus from Wyoming and South Australia. Based on four additional specimens found nearby (Fig. 12.2) and some others collected by J. Wyatt Durham (Durham, Reference Durham1974), Alpert (Reference Alpert1974, p. 74) felt that “the resemblance of Cloud's [sic] and Nelson's specimen to Plagiogmus may be fortuitous”, a view shared by Nelson (Reference Nelson1978), Jensen and Grant (Reference Jensen and Grant1996), Runnegar (Reference Runnegar1998), Jensen et al. (Reference Jensen, Droser, Gehling, Xiao and Kaufman2006), and Smith et al. (Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023), but not by Durham (Reference Durham1974), Corsetti and Kaufman (Reference Corsetti and Kauffman1994), Kaufman and Corsetti (Reference Kaufman and Corsetti1996), or O'Neil et al. (Reference O'Neil, Tackett and Meyer2022). Nevertheless, Plagiogmus has largely disappeared from the ichnofossil inventory of the White-Inyo succession (e.g., Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022).
Ironically, the most convincing example is one of several similar specimens described by Nelson and Smith (Reference Nelson and Smith2019, p. 909) as “microbially induced sedimentary structures” (MISS, Noffke, Reference Noffke2009). Nelson and Smith (Reference Nelson and Smith2019, fig. 2A) illustrated two aligned, annulated burrows, one of which has the characteristic ladder-like ridges of Plagiogmus on its presumed original floor (Fig. 12.1). Several other specimens (Nelson and Smith, Reference Nelson and Smith2019, fig. 2; Reference Nelson and Smith2020, fig. 1) are far less like typical Plagiogmus, but may be interpreted as more or less intact burrows that have been deformed by soft-sediment movement after they were produced. As Glaessner (Reference Glaessner1969) had noted, most examples of Plagiogmus are preserved in medium- to coarse-grained sandstones and therefore resist compaction and other kinds of deformation. Apparently, this was not the case for the structures found at Mount Dunfee (Nelson and Smith, Reference Nelson and Smith2019), nor for the one from Cloud and Nelson's original locality (Glaessner, Reference Glaessner1969; Alpert, Reference Alpert1974, pl. 5, fig. 11), both of which are found in finer-grained sediment. The preservation of the burrows is also complicated by superposition during dewatering (Fig. 12.4), which has led to interference patterns that apparently obscure the meandrine morphology that is typical of Psammichnites gigas arcuatus (Roedel, Reference Roedel1929), the type species of Plagiogmus. Additional confidence in the identification of the White-Inyo and Mount Dunfee structures as ‘Plagiogmus’ comes from their almost exact stratigraphic equivalence in the middle part of the Esmeralda Member of the Deep Spring Formation (Cloud and Nelson, Reference Cloud and Nelson1966; Nelson and Smith, Reference Nelson and Smith2019; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023), and from the report of abundant Plagiogmus by Oliver (Reference Oliver1990) from about the same stratigraphic position in the Mount Dunfee section as Nelson and Smith's (Reference Nelson and Smith2019) MISS structures. There are, however, some significant differences between the ‘Plagiogmus’ from the Deep Spring Formation and other early Cambrian examples of this ichnofossil; these discrepancies are discussed in the Systematic Paleontology section.
In the Mount Dunfee section, ‘Plagiogmus’ occurs ~435 m above the top of the Reed Dolomite (Nelson and Smith, Reference Nelson and Smith2019, fig. 1) at a level that is at or very near the peak (nadir) of the BACE negative carbon isotope excursion (Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016, Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). ‘Plagiogmus’ is an iconic Cambrian ichnofossil and is not known from the Ediacaran, which means that the first appearance datum (FAD) for Treptichnus pedum—which has been used as a local standard for identifying the Ediacaran–Cambrian boundary in the Great Basin succession (Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023)—may be just the start of the local stratigraphic range of T. pedum rather than the beginning of its biozone, and that the criterion for identifying the boundary in this region should now be supplanted by the FAD of ‘Plagiogmus’ (i.e., Psammichnites gigas arcuatus). If that is done, the need to reduce the age of the Ediacaran–Cambrian boundary from ca. 539 to ca. 533 Ma (Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023) goes away, because the nadir of the BACE excursion is ca. 539 Ma old in Sonora, Mexico (Hodgin et al., Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2021; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023).
Chemostratigraphy and geochronology
In the White-Inyo successions, Deep Spring Formation carbonates yield the outlines of a significant negative carbon isotope excursion that has been identified as the basal Cambrian excursion (BACE; Corsetti and Kauffman, Reference Corsetti and Kauffman1994; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2003; Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016; Hodgin et al., Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2021; Topper et al., Reference Topper, Betts, Dorjnamjaa, Li, Li, Altanshagai, Enkhbaatar and Skovsted2022). The limited availability of carbonates in the Death Valley succession makes recognition of this event more difficult there, but the available data do support the traditional correlation of the Deep Spring Formation with the LMWCF (Corsetti and Hagadorn, Reference Corsetti and Hagadorn2003; Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023).
The passive margin succession of the Great Basin has few volcanic flows or ash beds, but Nelson et al. (Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023) found and dated three successive ash beds in the upper part of the LMWCF in the Spring Mountains (Fig. 1.1) using the U–Pb ID-TIMS method. The ages range from 532.83 ± 0.98 to 531.32 ± 0.66 Ma and they embrace the section that, in the Montgomery Mountains and Nopah Range, contains the FAD of Treptichnus pedum (Supplemental dataset 1). For this reason, Nelson et al. (Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023) advocated elevating the age of the base of the Cambrian by ca. 6 Ma. However, for the reasons already discussed, this is not necessary; instead, the FAD of T. pedum in the LMWCF should be regarded as late Fortunian in age, according to recent estimates (533–530 Ma) of the age of the base of Cambrian Stage 2 (Bowyer et al., Reference Bowyer, Zhuravlev, Wood and Shields2022, Reference Bowyer, Zhuravlev, Wood, Zhao, Sukhov, Alexander, Poulton and Zhu2023; Hamilton et al., Reference Hamilton, Álvaro, Barr, Jensen, Johnson, Palacios, van Rooyen and White2024).
Stratigraphic information
See Figures 2–4, Appendix 1, Supplementary dataset 1, and the discussion under Materials and methods.
Locality information
RJH used his own numbering system based on his initials and date of collection (e.g., RH-91-8-19-1) and recorded site information in his field notebooks: https://sova.si.edu/record/sia.fa16-098. We used the UCLA locality numbering system (e.g., UCLA 7333) for sites, occasionally at the same place in stratigraphic order, and numbered each piece of rock collected accordingly. Important fossils were then given decimal numbers (e.g., 7333.1, 7333.2, etc.). Pieces of an individual fossil and parts and counterparts were given the same decimal number and then distinguished by letters (e.g., 7333.1A, 7333.1B, etc.). Those UCLA numbers for individual fossils were replaced by Los Angeles County Museum Invertebrate Paleontology (LACMIP) locality numbers (e.g., LACMIP 43369), specimen numbers (e.g., LACMIP 43369.1), and type numbers (e.g., LACMIP Type 15019) after the collection was donated to the Museum, but the UCLA locality numbers (e.g., UCLA 7307) still pertain and are used in the figures, Appendix 1, and Supplemental dataset 1. All collection numbers associated with type specimens are listed for clarity and in order to maintain specimen provenance; locality information is tabulated in Supplemental dataset 1 and the localities are described in Appendix 1. Because this work was carried out before GPS became widely available, the geographic positions of localities were plotted on road and topographic maps of various sources and scales. These map records were used recently to find the exact locations of the sites on Google Maps and to obtain their decimal longitudes and latitudes (Fig. 1; Appendix 1); Google Maps uses the WGS84 standard.
Materials and methods
All of the material used for this study was obtained during field work carried out in 1993 and 1995. Stratigraphic sections were measured using a Jacob staff or a measuring tape. There are, however, some discrepancies between our measurements and those of others who have studied the same sections (Supplemental dataset 1), presumably because of differences in estimates of average dip, which varies significantly from place to place.
Preparation of the fossils has been minimal. Field photographs and images of specimens taken during the 1990s were made with a Minolta X700 35 mm FSLR camera equipped with Minolta MC Macro Rokkor-QF 50 mm lens using Kodak Ektachrome Professional film. Color slides and negatives were digitized using an Epson Perfection V700 Photo scanner. Digital images, taken more recently, were made with a Nikon D3100 DSLR camera equipped with Nikon AF-S Micro Nikkor 40 mm lens. Preparation of the figures was carried out with Adobe Photoshop, Adobe Illustrator, and Aldus Super3D.
Repositories and institutional abbreviations
Figured and other specimens examined or mentioned in this study are deposited in the following institutions: Natural History Museum of Los Angeles County, Invertebrate Paleontology, Los Angeles (LACMIP); South Australian Museum, Adelaide, Australia (SAM); Swedish Museum of Natural History, Stockholm, Sweden (SMNH); Faculty of Geology Museum, University of Oveido, Spain (AMC); Department of Earth and Planetary Sciences, University of California, Riverside, California (UCR). Locality information is registered with the Department of Earth, Planetary, and Space Sciences, University of California, Los Angeles (UCLA), Los Angeles, U.S.A., the Natural History Museum of Los Angeles County, Invertebrate Paleontology, Los Angeles (LACMIP or LACNHM), the Department of Earth and Planetary Sciences, Johns Hopkins University, Baltimore (JHU) and the School of Environmental and Rural Science, University of New England, Armidale, Australia (UNE).
Systematic paleontology
Class Erniettomorpha Pflug, Reference Pflug1972
Family Erniettidae Pflug, Reference Pflug1972
Genus Tulaneia Runnegar and Horodyski new genus
Type species
Tulaneia amabilia n. gen n. sp. from the lower member of the Wood Canyon Formation, Montgomery Mountains, Nevada.
Diagnosis
As for the type species by monotypy.
Occurrence
Lower member, Wood Canyon Formation, Nevada, U.S.A. (Horodyski, Reference Horodyski1991; Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994; Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017, Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023).
Etymology
Named for Tulane University, New Orleans (formerly the University of Louisiana) for its role in encouraging and supporting research in paleontology and other areas of natural history as exemplified by the distinguished careers of Robert J. Horodyski, Alfred R. Loeblich and Helen Tappan Loeblich, Ronald L. Parsley, Harold E. and Emily H. Vokes, and Shuhai Xiao.
Remarks
Tulaneia n. gen. resembles Ernietta Pflug (Reference Pflug1966) in being constructed from a stockade of tubular modules but differs in having only a single, fan-shaped array, rather than being shaped like a sack. Because there are two sets of growing tips on one of the paratype specimens (Figs. 7.1–7.6, 8), it was originally thought to have a sack-shaped body that was flattened during burial and compaction. Closer inspection of the paratype revealed that the modules in this specimen bifurcate near their growing tips (Fig. 7.4) and do not, therefore, represent two sides of a sack-shaped organism. This terminal bifurcation is thought to be a property of the organism rather than a developmental abnormality, although the latter possibility cannot be excluded.
Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp. (1–9) and two possible taphomorphs of T. amabilia Runnegar and Horodyski n. gen. n. sp. (10–12), all except (8) from the lowest part of the lower member of the Wood Canyon Formation at the RJH discovery site (Figs. 1.2, 4.3), Montgomery Mountains, Nevada; (8) is from ~14 m above the base of the Wood Canyon Formation on the opposite (south) side of the ridge used to measure section 7333 (Fig. 1.2). (1–6) Several images of three small pieces (A–C), found separately by RJH (B and C) and KJP (A), but which proved to fit together as counterpart A (1, left), part B (4, right), counterpart C (2, left), and part D (1, right; 2, right; 3) to form paratype LACMIP 43369.6; three pieces (B–D) of paratype LACMIP 43369.6 are assembled in (4), (5), and (7) and their original biological continuity is best illustrated by the smooth curved external surface that connects part A with part C (4, arrow). (1) Small slab that has cast three equal-sized tubular modules (a–c) and one of double width (d–e) that is divided on the opposite side of the organism, paratype LACMIP 43369.6A (counterpart); insert shows a view of part C that has been whitened with ammonium chloride for comparison with (3); (2) all three fragments juxtaposed, with part B and counterpart C lying in place on counterpart A, and part D inverted on counterpart A to the right; the end of the b module of part B is just visible (arrow), paratype LACMIP 43369.6A–D; (3) unwhitened view of part C for comparison with the insert in (1), paratype LACMIP 43369.6C; (4) three views of the assembled parts B through D showing two tubular modules (b and c) and one of double width (d + e) on the counterpart A side of the organism, paratype LACMIP 43369.6B–D; (5) view of distal end of the assembled parts B through D with the b module of part B indicated by the arrow, paratype LACMIP 43369.6B–D; (6) proximal view of the broken edge of part D showing cross-sections of four modules and the positions of the three intervening walls (arrows), paratype LACMIP 43369.6D; (7) view of the proximal end of the assembled parts B through D showing a wide flat surface that represents the unseparated d + e modules of part B, paratype LACMIP 43369.6B–D; (8, 9) fragments of poorly preserved specimens of Tulaneia n. gen. that have more stratigraphic than biological significance, having been found at approximately the same level on opposite sides of the ridge section UCLA 7333 (Fig. 1.2) LACMIP 43369.5 and LACMIP 43369.11, respectively; (10–12) two similar sized concretion-like structures that may represent inflated taphomorphs of the outer wall of an erniettomorph such as Tulaneia n. gen.; the one that has been sectioned has a volume of quartzite that may reflect an earlier filled shape, LACMIP 43369.10 (10) LACMIP 43369.9 (11, 12). Scale bars = 5 mm (6), 1 cm (1–5, 7–9), and 2 cm (10–12).

Three-dimensional model of several mature modules of Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp. based on the holotype and paratypes, some artistic license and the assumption that the two sets of growing tips seen in paratype LACMIP 43369.6 (Fig. 7.1–7.6) are a feature of the organism, not an individual aberration such as a congenital abnormality. (1) Front view of model at right angles to plane of bilateral symmetry; (2) side view of model showing most of one septum (yellow/light area); (3) oblique view of back of model.

Tulaneia amabilia Runnegar and Horodyski new species
Figures 5, 6.1–6.7, 7.1–7.9, 7.11, 7.12?, 8
- Reference Horodyski1991
Ernietta Pflug; Horodyski, p. A163.
- Reference Horodyski, Gehling, Jensen and Runnegar1994
Ernietta plateauensis Pflug; Horodyski et al., p. 60.
- Reference McMenamin1998
erniettid Ediacaran; McMenamin, p. 30, fig. 2.16.
- Reference Smith, Nelson, Tweedt, Zeng and Workman2017
Ernietta Pflug; Smith et al., p. 4, fig. 3a–h.
- Reference Hall, Smith, Tamura, Fakra and Bosak2020
erniettomorph; Hall et al., p. 2, fig. 1b.
- Reference Runnegar2022
Ernietta plateauensis Pflug; Runnegar, p. 1104, fig. 3a.
Holotype
Part (RH-91-8-19-1/UCLA 7331.1B/LACMIP 43369.1A LACMIP Type 15019) and counterpart (RH-91-8-19-1/UCLA 7331.1A/LACMIP 43369.1B LACMIP 15019) preserved on two of three stratigraphically contiguous thin sandstone slabs (Fig. 5.2) from the lowest 10–20 m of the lower member of the Wood Canyon Formation at the discovery site (RJH, Fig. 1.2), northwest of Johnnie, Nevada; the part preserves a clean quartzite filling of the modules (Fig. 6.1).
Paratypes
Paratype, part (RH-91-8-19-1/UCLA 7333.1C/LACMIP 43369.2A LACMIP Type 15020) and counterpart (RH-91-8-19-1/UCLA 7333.1B/LACMIP 43369.2B Type 15020) preserved on the second and third slabs of the three stratigraphically contiguous slabs that also preserve the holotype (Fig. 5.2); paratype, larger of two individuals on another slab (RH-92-1-12-1/UCLA 7333.2/LACMIP 43369.3 LACMIP Type 15021; Fig. 6.4), also filled with quartzite (Fig. 6.5); paratype, part (UCLA 7333.3B/LACMIP 43369.4A LACMIP Type 15022) and counterpart (UCLA 7333.3A/LACMIP 43369.4B LACMIP Type 15022) of the distal half of a specimen that is partially filled with clean quartzite (Fig. 6.7); paratype, three small separate pieces that fit together and preserve the counterpart (UCLA 7333.5A/LACMIP 43369.6A LACMIP Type 15023; Fig. 7.1), part and counterpart (RH-91-8-17-1/UCLA 7333.5B/LACMIP 43369.6B–C LACMIP Type 15023; Fig. 7.2), and part (RH-91-8-16-1/UCLA 7333.5C/LACMIP 43369.6D LACMIP Type 15023; Fig. 7.3) of the tips of two sets of quartzite-filled modules (Fig. 7.1–7.6).
Diagnosis
Decimeter-scale lyre-shaped organism composed of unmineralized tubular modules arranged side-by-side like a palisade and separated by planar partitions; modules are wide near the proximal edge of the body, diminish in size gradually in the distal direction, and then separate laterally and taper to blunt points at their distal terminations; duplication of the tips of the modules has been observed in one case.
Description
Body lyre-shaped, composed of as many as ~30 tubular modules in complete specimens, arranged side-by-side in a palisade fashion, and which join proximally in an acutely rounded margin; body cross section is bilaterally symmetrical with maximum width near the proximal margin and minimum width at tapered distal terminations; profile at right angles to the proximal margin is lyre-shaped and parallel to the margin; it is aerodynamic (streamlined); modules are largest at the midline, up to ~5 mm width near the proximal margin and up to ~9 mm in depth perpendicular to the margin, and they diminish gradually in size in both directions away from the axis; lateral limits of body not well preserved/exposed; one three-dimensional specimen has modules with bifurcated distal tips, which may or may not be a general property of the organism.
Etymology
From amabilis, Latin, lovely, in reference to the exquisite preservation of the type material.
Taphonomy
All of the type specimens of Tulaneia n. gen. were completely or partially filled with clean quartz sand prior to burial (Figs. 5.5, 5.6, 6.1–6.3, 6.5, 6.7, 7.5–7.7), as was one of the associated ‘taphomorphs’ (Fig. 7.11). The obvious contrast between the clean fillings and their surrounding silty matrices is an indication that the internal cavities of Tulaneia n. gen. were loaded with suspended quartz grains, either in a high-energy shoreface environment like that of the underlying Stirling Quartzite (Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017), or during onshore to offshore transport via storm surge activity. As a result, the bodies of Tulaneia n. gen. seem to have behaved like sandbags during transport and were deposited with cobbles (Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017) in deeper water environments. As in Namibia and South Australia, erniettomorphs (Ernietta, Pteridinium, and Tulaneia n. gen.) are rarely if ever preserved in the environments in which they lived (Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024).
Remarks
A cluster of distorted specimens of Tulaneia amabilia n. gen. n. sp. discovered by Smith et al. (Reference Smith, Nelson, Tweedt, Zeng and Workman2017) in the vicinity of LACNHM locality 17130 (Fig. 1.2) are similar in shape, size, and preservation to the material from the Horodyski site (RJH, Fig. 1.2). However, one of their specimens (Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017, fig. 3a, b, e, f) was thought to show a suture of the kind found in Ernietta, which has a zig-zag seam between opposite sides of the body (Ivantsov et al., Reference Ivantsov, Narbonne, Trusler, Greentree and Vickers-Rich2016). Close inspection of the published images suggests that the ‘suture’ is due to a fortuitous juxtaposition of two individuals, with the proximal margin of one nearly aligned with the proximal margin of the other. In this interpretation, the proximal margin of the larger of the two individuals is fully exposed whereas the proximal margin of the smaller individual abuts the wall of the larger one at some distance from its proximal margin. Although this configuration could possibly be explained by budding (asexual reproduction), sedimentary stacking during deposition is our preferred hypothesis.
Several puck-shaped structures, one of which is partially filled with clean quartzite, were also found at RJH's discovery site (Fig. 7.10–7.12) and contributed to the erroneous identification of Tulaneia n. gen. as Ernietta. Circumstantial evidence based on their co-occurrence suggests that these and similar objects found by Smith et al. (Reference Smith, Nelson, Tweedt, Zeng and Workman2017) may be the distended exterior walls of individuals of Tulaneia n. gen. or perhaps another eriettomorph. Some of the structures reported by Smith et al. (Reference Smith, Nelson, Tweedt, Zeng and Workman2017) showed traces of fine or coarse ribbing and were regarded as ‘taphomorphs’ of the better-preserved body fossils (Hall et al., Reference Hall, Smith, Tamura, Fakra and Bosak2020). We concur with that suggestion.
Ichnofossils
Remarks
Ichnofossils have been instrumental in attempts to place the Ediacaran–Cambrian boundary in both the White-Inyo and Death Valley regions of the Great Basin for almost 60 years (Cloud and Nelson, Reference Cloud and Nelson1966; Glaessner, Reference Glaessner1969; Cloud and Bever, Reference Cloud and Bever1973; Langille, Reference Langille1973, Reference Langille1974; Alpert, Reference Alpert1974, Reference Alpert1975, Reference Alpert1976; Durham, Reference Durham1974; Nelson, Reference Nelson1978; Oliver, Reference Oliver1990; Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994; Runnegar, Reference Runnegar1998; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000, Reference Corsetti and Hagadorn2003; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002; Oliver and Rowland, Reference Oliver, Rowland and Corsetti2002; Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017, Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023; Tarhan et al., Reference Tarhan, Myrow, Smith, Nelson and Sadler2020; O'Neil et al., Reference O'Neil, Tackett and Meyer2022). Here we focus on three kinds of ichnofossils, selected for their paleoenvironmental significance and biostratigraphic importance: (1) looped structures (Fig. 9.1, 9.2; Langille, Reference Langille1973, Reference Langille1974), either body or trace fossils, from the Wyman Formation at Hines Ridge (Fig. 1.1) in the White-Inyo facies; (2) traces of the Treptichnus spectrum (Figs 9.5–9.8, 10.1, 10.3, 10.7, 11.5, 11.6), including bona fide examples of Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.2, 10.4; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002) and Didymaulichnus miettensis Young, Reference Young1972 (Fig. 11.1–11.3; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002), from the lower member of the Wood Canyon Formation in the Death Valley facies; (3) Psammichnites gigas arcuatus (Roedel, Reference Roedel1929) (Fig. 12.1–12.3, 12.5; Cloud and Nelson, Reference Cloud and Nelson1966; Alpert, Reference Alpert1974; Nelson and Smith, Reference Nelson and Smith2019) from the Esmeralda Member of the Deep Spring Formation in the White-Inyo facies.
Looped structures (body fossils or trace fossils) that have constant widths/diameters and are preserved in convex hyporelief from the Wyman Formation at Hines Ridge, California (1, 2), the Ediacara Member of the Rawnsley Quartzite at Ediacara, South Australia (3), and the lower member of the Wood Canyon formation in the UCLA 7334 section, Montgomery Mountains, Nevada (4), as well as bed-base traces of the Treptichnus spectrum, all from the lower member of the Wood Canyon Formation (5–8). (1) Backyard photograph of a specimen collected prior to November 1970 by Langille (Reference Langille1974) and reported anecdotally in the literature since then; (2) a specimen found by BR in 1988 that is similar in size and topology (arrows) to Langille's tentatively identified as the ichnotaxon ?Gordia isp. (1), LACMIP 43374.1; (3) a comparable looped hyporelief structure from an Ediacara Member flagstone, previously illustrated as the bilaterian trace fossil Gordia isp. (Runnegar and Fedonkin, Reference Runnegar, Fedonkin, Schopf and Klein1992, fig. 7.6.3C), SAM P27977; (4) a float specimen from UCLA 7334 that, perhaps, is more confidently identified as the trace fossil Gordia isp. rather than a body fossil, LACMIP 43372.1; (5), a specimen found by RJH in 1991 that, according to him, came from ~10 m above the Tulaneia n. gen. horizon in the 7333 section (Supplemental dataset 1), LACMIP 43369.15; (6) another specimen from the RJH collection at an unrecorded level in the 7333 section that is similar to (5) but may be more confidently be referred to Treptichnus isp., LACMIP 43369.16; (7, 8) unwhitened and whitened images of a trace from the 203 m level in the Chicago Pass section, UCLA 7329, showing behavior that is approaching that of bona fide Treptichnus pedum (Fig. 10.8), LACMIP 43370.1. Scale bars = 5 cm (1, 2) and 2 cm (3–8).

Treptichnus isp. cf. T. pedum (Seilacher) from the lower member of the Wood Canyon Formation in the Desert Range and Montgomery Mountains (1, 3) and Treptichnus pedum from the lower member of the Wood Canyon Formation in the Montgomery Mountains (2, 4, 6), Chicago Pass (7), and the Nomtsas Formation, Sonntagsbrunn farm, southern Namibia (8). (1) Bed-base traces from a level with numerous similar structures that collectively can be identified as Treptichnus spectrum (Fig. 3), LACMIP 43371.1; (2) RJH sample from an unrecorded level within the lower member of the Wood Canyon Formation but probably from the same horizon as UCR 10703/1 (Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002, fig. 4B), LACMIP 43369.17; (3) one of several structures (Figs 8.5, 8.6, 8.7, 8.8, 10.5, 10.6) that grade between Treptichnus and Gordia in continuity of the trace and geometry of its trajectory, LACMIP 43369.20; (4) iconic example of T. pedum found by MJV in float between the second and third major dolostones, LACMIP 43372.5; (5) enlarged view of a segmented trace that falls well within the Treptichnus spectrum shown in Figure 11.7, LACMIP 43369.18; (6) another iconic example of T. pedum with better stratigraphic control (Supplemental dataset 1), LACMIP 43369.19; (7, 8) comparable specimens of T. pedum from Nevada and Namibia, LACMIP 43370.2 and GSN F 1952, respectively. All scale bars = 2 cm.

Didymaulichnus miettensis Young (1–3) from the lower member of the Wood Canyon Formation, Chicago Pass and Montgomery Mountains, California (1, 3) and Member IV, Arumbera Formation, Ross River, Northern Territory, Australia (2), plus looped trace fossils (4, 5) and Treptichnus spectrum traces (6, 7) from the lower member of the Wood Canyon Formation, Nevada. (1) Small piece, found in situ at ~200 m (Supplemental dataset 1), showing parts of three curved traces, LACMIP 43370.3; (2) longer, similar traces from the Arumbera Formation, Australia, also preserved in convex hyporelief, LACMIP 43373.1; (3) sandstone joint block with several hyporelief traces, previously illustrated by Jensen et al. (Reference Jensen, Droser, Heim and Corsetti2002, fig. 5A), LACMIP 43369.20; (4) two looped traces that show features of both Gordia and Didymaulichnus, bed base whitened with ammonium chloride, LACMIP 43369.2; (5) looped trace on upper bed surface at 95 m, closely comparable to Psammichnites gigas circularis, LACMIP 43372.3; (6) looped trace that is intermediate between Treptichnus and Gordia, found in situ 6–7 m below third major dolostone (Supplemental dataset 1), LACMIP 43372.4; (7) two Treptichnus spectrum traces on a small slab, whitened with ammonium chloride from ~150 m, LACMIP 43369.18. All scale bars = 2 cm.

Psammichnites gigas arcuatus (Roedel), Esmeralda Member, Deep Spring Formation at Mount Dunfee, Nevada (1, 5) and Mollie Gibson, White-Inyo Mountains, California (2, 3); Member IV, Arumbera Sandstone, Ross River, Northern Territory, Australia (4); and File Haidar Formation, Sweden (6). (1) Least-deformed and therefore most recognizable of ‘problematic corrugated tubular structures’ illustrated by Nelson and Smith (Reference Nelson and Smith2019, p. 911, fig. 2A, republished with permission); arrow indicates presumed wall of burrow for comparison with (2); (2) two of four additional specimens of ‘cf. Plagiogmus’ UCLA 49569 and UCLA 49570, now LACMIP 26153.1 and LACMIP 26160.1, respectively from Alpert (Reference Alpert1974, pl. 5, figs 3, 4, republished with permission) UCLA localities 6153 and 6150 (Fig. 1.3), showing corrugation of both upper (arrow) and lower surfaces of the burrow; (3) specimen found by Cyril Gallick ~600 m SE of the Mollie Gibson mines (Fig. 1.3; Cloud and Nelson, Reference Cloud and Nelson1966, fig. 3D), LACMIP 16338.1 ex UCLA 45726; (4) sandstone cast of lower surface of a complete burrow with a characteristic sinuous groove on the upper surface (not shown), LACMIP 43373.2; (5) upper surface of sizeable slab described as “bifurcating ripples with corrugations” by Nelson and Smith (Reference Nelson and Smith2019, p. 912, fig. 4A, republished with permission), with one superimposed crossover indicated by arrow; (6) inverted image of a photograph provided by Anders Martinsson of the bases of ‘empty’ burrows, cast in sandstone, with regularly spaced transverse ridges caused by peristaltic contractions of the producers, SMNH X2074 (Jaeger and Martinsson, Reference Jaeger and Martinsson1980, fig. 2). All scale bars = 3 cm.

Looped structures (trace or body fossils)
Figure 9.1–9.4
Description
Poorly preserved, apparently cylindrical structure at least ~50 cm in length and 5 mm in width that makes one complete loop of ~5 cm diameter.
Material
LACMIP 43374.1, slab of slate from the upper Wyman Formation, Hines Ridge, California (Fig. 1.1).
Remarks
In 1970, Gerry Langille, then a graduate student at SUNY Binghamton (now Binghamton University), sent Clem Nelson a field photograph of a similar trace that he found in the Wyman Formation; Nelson passed that photograph on to BR in ca. 1988 and it is reproduced here (Fig. 9.1). Our specimen closely resembles Langille's in dimensions and topology, with similar crossings (indicated by arrows in Fig. 1.1 and 1.2). A third, possibly similar specimen, was illustrated by Corsetti and Hagadorn (Reference Corsetti and Hagadorn2003, fig. 5A) as Helminthoidichnites but with the proviso that “we cannot falsify the hypothesis that some of these are body fossils” (Corsetti and Hagadorn, Reference Corsetti and Hagadorn2003, p. 6).
Runnegar and Fedonkin (Reference Runnegar, Fedonkin, Schopf and Klein1992, fig. 7.6.3C) illustrated a comparable structure from the base of a bed at Ediacara, South Australia, which they confidently identified as the trace fossil Gordia isp. We refigure that specimen (Fig. 9.3) in order to illustrate its similarity to the Wyman structures but note that it is no longer possible to be confident that it is a trace fossil rather than a body fossil. The new understanding that followed discoveries of numerous tubular body fossils in the Ediacaran of Namibia (Budd and Jackson, Reference Budd and Jackson2015; Schiffbauer et al., Reference Schiffbauer, Huntley, O'Neil, Darroch, Laflamme and Cai2016; Darroch et al., Reference Darroch, Smith, Laflamme and Erwin2018; Chai et al., Reference Chai, Wu and Hua2021; Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024) and elsewhere, make it equally likely that the Wyman structures, and the one from Ediacara, are body fossils rather than traces, although the half-meter length of the former would be extraordinary for a tubular body fossil (but see Bobrovskiy et al., Reference Bobrovskiy, Nagovitsyn, Hope, Luzhnaya and Brocks2022, fig. S1). Another similar structure from the LMWCF (Fig. 9.4) is more confidently identified as the ichnofossil Gordia isp. because of the complexity of the loops and the ways they are tied together. None of these structures, whether they be trace or body fossils, has much biostratigraphic utility, but they include the only known fossils from the Wyman Formation and are compatible with an Ediacaran age for that unit.
Ichnogenus Didymaulichnus Young, Reference Young1972
Type ichnospecies
Fraena lyelli Rouault, Reference Rouault1850; partim, from Ordovician sandstones of the Armorican Massif of western France, by original designation (Young, Reference Young1972).
Didymaulichnus miettensis Young, Reference Young1972
Figure 11.1–11.3
Holotype
GSC 27802, ostensibly from the upper part of the Miette Group, but possibly from Cambrian float (Buatois and Mángano, Reference Buatois and Mángano2016, p. 38), at GSC locality C-5466 near the headwaters of the McGregor River, eastern British Columbia, Canada, by original designation.
Material
LACMIP 43370.3 and LACMIP 43369.20 from the LMWCF, Chicago Pass, California (Fig. 11.1) and Montgomery Mountains, Nevada, plus LACMIP 43373.1, Member IV, Arumbera Formation, Ross River, Northern Territory, Australia (Fig. 11.1).
Remarks
At the type locality, D. miettensis is preserved as sandstone casts of bilobed trails that have symmetrically inclined planar side zones (Young, Reference Young1972, fig. 10). That profile is visible in the larger of the two curved traces from the upper part of the LMWCF at Chicago Pass (Fig. 11.1) but the smaller one on the same slab is perhaps more similar to the hypichnial trace (Fig. 11.2) from Member 4 of the Arumbera Formation, central Australia, that was identified by Walter et al. (Reference Walter, Elphinstone and Heys1989) as Didymaulichnus lyelli (Rouault, Reference Rouault1850). Jensen et al. (Reference Jensen, Droser, Heim and Corsetti2002, fig. 5A) illustrated several specimens of D. miettensis on the base of a thick sandstone bed from near the top of the LMWCF in section 7333 (Fig. 1.2).
Ichnogenus Gordia Emmons, Reference Emmons1844
Type ichnospecies
Gordia marina Emmons, Reference Emmons1844, from an Ordovician ‘fine flagging stone’ (calcareous turbidite) of the Giddings Brook slice, Taconic Allochthon (Landing, Reference Landing, Derby, Fritz, Longacre, Morgan and Sternbach2012), at Mr. M‘Arthur's quarry, Jackson, New York, by monotypy.
Gordia isp.
Figures 9.4, 11.4, 11.6
Material
LACMIP 43372.1 (Fig. 9.4), LACMIP 43369.20 (Fig. 11.4), and LACMIP 43372.4 (Fig. 11.6), all from the LMWCF, Montgomery Mountains, Nevada.
Remarks
Specimens from near the top of the LMWCF in section 7333 (Fig. 11.4) and as float in section 7334 (Fig. 9.4) are illustrated, but Gordia isp. was also found in situ below the third dolostone in that section (Supplemental dataset 1). The complexity of the loops, with apparent self-level crossing, is the only feature that unites these three examples, so this is a poorly characterized form taxon at this stage. The specimens in Figure 11.4 have a poorly preserved bilobed base, which aligns them with Didymaulichnus, but the strongly concentrated circling is unlike the behavior recorded by that ichnogenus.
Ichnogenus Psammichnites Torell, Reference Torell1870
Type ichnospecies
Psammichnites gigas (Torell, Reference Torell1868) from the early Cambrian Hardeberga Formation, Scania, Sweden, by subsequent designation (Fischer and Paulus, Reference Fischer and Paulus1969).
Remarks
Mángano et al. (Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022) subsumed three widely recognized Cambrian ichnospecies, Psammichnites gigas (Torell, Reference Torell1868), Plagiogmus arcuatus Roedel, Reference Roedel1929, and Taphrhelminthopsis circularis Crimes et al., Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977, into three subspecies of Psammichnites gigas: P. gigas gigas, P. gigas circularis, and P. gigas arcuatus. This unusual procedure was carried out for sound paleobiological reasons, but it had the unfortunate consequence of the replacement of a useful keyword ‘Plagiogmus’ with the cumbersome trinomial term, Psammichnites gigas arcuatus. Psammichnites gigas circularis and P. gigas arcuatus differ from the type subspecies, P. gigas gigas, in having the tendency to form extensive loops or ‘scribbles’ rather than just a generally unidirectional trace, and P. gigas circularis lacks the transverse bars that are so characteristic of P. gigas arcuatus (Fig. 12.1; Jaeger and Martinsson, Reference Jaeger and Martinsson1980; Walter et al., Reference Walter, Elphinstone and Heys1989; McIlroy and Heys, Reference McIlroy and Heys1997; Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022). All three subspecies are said to co-occur in Member 4 of the Arumbera Formation, central Australia (Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022, tables 1–3), based on data presented by Walter et al. (Reference Walter, Elphinstone and Heys1989) and McIlroy and Heys (Reference McIlroy and Heys1997), although those authors preferred to include all examples from the Arumbera Formation in Plagiogmus arcuatus.
Psammichnites gigas circularis (Crimes et al., Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977)
Figure 11.5
Holotype
AMC 1219 from the Cambrian Stage 3 Cayetano Beds of the Cándana Formation, Punta del Cerrón, Asturias, Spain (Crimes et al., Reference Crimes, Legg, Marcos, Arboleya, Crimes and Harper1977, pl. 8c; Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022, fig. 3) by original designation.
Material
LACMIP 43372.3 from the LMWCF, Montgomery Mountains, Nevada (Fig. 11.5).
Remarks
Psammichnites gigas gigas co-occurs with P. gigas circularis at the type locality of P. gigas circularis, but followed less-curved paths. A hand specimen from near the top of the LMWCF in section 7334 (Fig. 1.2) was identified as ‘Taphrhelminthopsis’ circularis by Jensen et al. (Reference Jensen, Droser, Heim and Corsetti2002, fig. 5B) and a complex bilobed scribbling trace from the younger early Cambrian Poleta Formation in the White-Inyo Mountains region was referred to a new species of Taphrhelminthopsis, T. nelsoni, by Hagadorn, Schellenberg, and Bottjer (Reference Hagadorn, Schellenberg and Bottjer2000); Mángano et al. (Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022) recommended synonymizing T. nelsoni with P. gigas circularis. A small specimen outlining an elegant pince-nez shape (Fig. 11.5) may be another example of P. gigas circularis.
Psammichnites gigas arcuatus (Roedel, Reference Roedel1929)
Figure 12
Holotype
Specimen figured by Häntzschel (Reference Häntzschel and Moore1962, fig. 168.6) from an early Cambrian, glacial erratic, sandstone block, found in northern Germany, but probably sourced from southern Sweden (Roedel, Reference Roedel1929; Jaeger and Martinsson, Reference Jaeger and Martinsson1980), by monotypy.
Material
Specimens previously illustrated by Alpert (Reference Alpert1974) and Nelson et al. (Reference Nelson and Smith2019) from the Esmeralda Member, Deep Spring Formation, California and Nevada (Fig. 12.1–12.3, 12.5) plus a small collection from Member IV, Arumbera Sandstone, Amadeus Basin, Australia (Fig. 12.4); LACMIP 26153.1 (= UCLA 49659 from UCLA locality 6153), LACMIP 26160.1 (= UCLA 49670 from UCLA locality 6160), LACMIP 26153.2 (= UCLA 49571 from UCLA locality 6153), LACMIP 26153.3 (= UCLA 49572 from UCLA locality 6153).
Remarks
Although only short segments of the structures are preserved in the small slabs so far recovered from California and Nevada (Fig. 12.1–12.3, 12.5), they are similar to specimens of P. gigas arcuatus from the Arumbera Sandstone (Fig. 12.4; McIlroy and Heys, Reference McIlroy and Heys1997) and early Cambrian glacial erratics in Sweden (Fig. 12.6; Jaeger and Martinsson, Reference Jaeger and Martinsson1980), when viewed from their presumed lower side. The U-turn (shown by an arrow in Fig. 12.5) resembles one at the bottom of a Swedish specimen (Fig. 12.6) and surfaces of the burrows from Mount Dunfee (Fig. 12.1) are similar to those of Cloud and Nelson's (Reference Cloud and Nelson1966) specimen and other examples found subsequently in the White-Inyo Mountains (Fig. 1.2, 1.3). A characteristic feature of Psammichnites gigas arcuatus and other subspecies of the ichnogenus is a median furrow on the top surface. The lack of this groove in the Deep Spring material may stem from their preservation in less cohesive sediments, which were retaining interstitial fluid during compaction and deformation. As a result, the median furrow, which McIlroy and Heys (Reference McIlroy and Heys1997) attributed to the collapse of an originally cylindrical burrow, did not develop. A possibly comparable morphology is shown by uncompacted burrows of Psammichnites gigas preserved in very fine-grained quartz arenite from the early Cambrian Backbone Ranges Formation of the Mackenzie Mountains, Canada. One sizeable block has 2- to 3-cm-wide burrows with variably developed, transverse corrugations on their upper surfaces and inconspicuous median grooves (MacNaughton et al., Reference MacNaughton, Fallas and Finley2021, fig. 4A).
The first appearance of P. gigas circularis is considered to be a defining feature of the Fortunian Rusophycus avalonensis Zone of Avalonian Newfoundland (Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022), but P. gigas arcuatus also appears during the early Terraneuvian in central and South Australia (Glaessner, Reference Glaessner1969; McIlroy and Heys, Reference McIlroy and Heys1997) and during the Fortunian in Russia (Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022). Its Great Basin occurrences could be even older, and they encourage the speculation that there is a closer similarity to Parapsammichnites pretzeliformis Buatois et al., Reference Buatois, Almond, Mángano, Jensen and Germs2018, from the latest Ediacaran of Namibia than was accepted when that large, complex, looped, and bilobed ichnofossil was first described.
Mángano et al. (Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022) differentiated Parapsammichnites from Psammichnites because the former followed a more diverse course, has plentiful self-overcrossings, pad-like back-fill structures, and lacks an axial furrow. A larger population of ‘Plagiogmus’ from the Deep Spring Formation is needed before a more detailed comparison with Parapsammichnites can be made. Comparison with the Nama Group material is also made difficult by the contrast in preservation; Parapsammichnites is mostly preserved in full relief on sandy bed bases at its only known locality, whereas the Deep Spring ‘Plagiogmus’ specimens seem to be largely intrastratal. In summary, the Deep Spring ‘Plagiogmus’ material is best interpreted as deformed and unusually preserved examples of Psammichnites gigas arcuatus, but more work is needed to confirm this assessment and to explore possible affinities with Parapsammichnites.
Ichnogenus Treptichnus Miller, Reference Miller1889
Type ichnospecies
Treptichnus bifurcus Miller, Reference Miller1889, from the Carboniferous, Gzhelian, Mansfield Formation of Indiana (Maples and Archer, Reference Maples and Archer1987), by original designation and monotypy.
Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955)
Figure 10.2, 10.4, 10.6–10.8
Holotype
Specimen figured by Seilacher (Reference Seilacher, Schindewolf and Seilacher1955, p. 387, fig. 4a) from the early Cambrian Neobulus shale (Khussak Formation), Salt Range, Pakistan (Buatois, Reference Buatois2018).
Material
LACMIP 43372.5 and LACMIP 43369.19 from the LMWCF, Montgomery Mountains, Nevada (Fig. 10.2, 10.4, 10.6) and LACMIP 43370.2 from the LMWCF, Chicago Pass, California (Fig. 10.7).
Remarks
Wilson et al. (Reference Wilson, Grotzinger, Fischer, Hand and Jensen2012) described and illustrated great variability in traces attributed to Treptichnus pedum from incised valley fillings of the Nomtsas Formation, southern Namibia. Those with outwardly projecting probe marks (‘segments’; Fig. 10.2, 10.4, 10.6) are the types most commonly considered characteristic of T. pedum, but pathways with more subdued segments are also common in the Nomtsas Formation (Fig. 10.8). Both types of traces are present in the LMWCF (e.g., Fig. 10.4 and 10.7). As discussed elsewhere, recent U–Pb ages from within the stratigraphic range of T. pedum in the LMWCF (Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023) make it clear that these occurrences are not earliest Fortunian in age.
Material
LACMIP 43369.15, LACMIP 43369.16, LACMIP 43369.17, LACMIP 43369.18, and LACMIP 43372.4 from the LMWCF, Montgomery Mountains, Nevada (Figs. 9.5, 9.6, 10.3, 11.6); LACMIP 43370.1 from the LMWCF Chicago Pass, California (Figs. 9.7, 9.8, 10.5, 11.7); LACMIP 43371.1 from the LMWCF, Desert Range, Nevada (Fig. 10.1).
Remarks
Most of the trace fossils from the LMWCF were collected from float, consequently no samples of populations are available to assess within-population variability. Furthermore, some traces vary significantly along their length (Fig. 9.7, 9.8) or express differently levees produced by motion parallel to the substrate versus probes made at right angles to the sediment–water interface. In order to avoid applying too many names to structures that may have been produced by the activities of a single sort of animal, we refer to these traces as the Treptichnus spectrum.
Traces of this kind include a specimen found by RJH (Fig. 9.5) at ~10 m above the Tulaneia n. gen. level in the 7333 section (Figs. 1.2, 3; Supplemental dataset 1). It occurs with circular structures of the Nimbia kind (Fig. 9.5; Fedonkin, Reference Fedonkin1980), similar to those previously reported from the Stirling Quartzite (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000). Other traces at this end of the spectrum occur higher in the LMWCF in the Montgomery Mountains (Figs. 9.6, 10.3) and in the Desert Range (Fig. 10.1). We identify some as Treptichnus isp. cf. T. pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) in the figure explanations, Appendix 1, and Supplemental dataset 1.
Results and Discussion
Ediacaran–Cambrian transition
The proposal by Nelson et al. (Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023) to reduce the age of the Ediacaran–Cambrian boundary by ca. 6 Ma was based on the understanding that the FAD of Treptichnus pedum in the correlative Montgomery Mountains and Chicago Pass sections corresponded, at least approximately, to the times of its first appearance at the Cambrian GSSP site in Newfoundland (Gehling et al., Reference Gehling, Jensen, Droser, Myrow and Narbonne2001; Gougeon et al., Reference Gougeon, Mángano, Buatois, Narbonne, Laing and Paza2023), Namibia (Jensen et al., Reference Jensen, Saylor, Gehling and Germs2000; Linnemann et al., Reference Linnemann, Ovtcharova, Schaltegger, Gärtner and Hautmann2019; Darroch et al., Reference Darroch, Cribb, Buatois, Germs and Kenchington2021; Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024), Mexico (Hodgin et al., Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2021), South Australia (Jensen et al., Reference Jensen, Gehling and Droser1998), and Norway (Högström et al., Reference Högström, Jensen, Palacios and Ebbestad2013), and at similar levels elsewhere. Precise U–Pb ages from the upper half of LMWCF of the Spring Mountains section range from 532.83 ± 0.98 to 531.32 ± 0.66 Ma (Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023). These ages overlap the earlier of two possible estimates for the Fortunian–Stage 2 boundary (531–532 Ma) in New Brunswick, Canada (Bowyer et al., Reference Bowyer, Zhuravlev, Wood and Shields2022; Hamilton et al., Reference Hamilton, Álvaro, Barr, Jensen, Johnson, Palacios, van Rooyen and White2024). If these ages are correct, the null hypothesis should be the FAD of T. pedum in the LMWCF is late, not earliest Fortunian in age.
At Hines Ridge (Corsetti and Kauffman, Reference Corsetti and Kauffman1994; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023), Molly Gibson (Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023), and Mount Dunfee (Corsetti and Hagadorn, Reference Corsetti and Hagadorn2003; Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016), a negative carbon isotope excursion within the Deep Spring Formation has been identified as the BACE, the basal Cambrian negative carbon isotope excursion (also known as ‘N’ or ‘1n’), which is well expressed in Siberia (e.g., Bartley et al., Reference Bartley, Pope, Knoll, Semikhatov and Petrov1998; Kouchinsky et al., Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007), and has been proposed as the principal criterion for an auxiliary stratotype section and point (ASSP) for the eon boundary (Topper et al., Reference Topper, Betts, Dorjnamjaa, Li, Li, Altanshagai, Enkhbaatar and Skovsted2022). The trough (nadir) of this excursion is precisely dated as 539.40 ± 0.23 Ma in Mexico (Hodgin et al., Reference Hodgin, Nelson, Wall, Barrón-Díaz, Webb, Schmitz, Fike, Hagadorn and Smith2021) and, as mentioned previously, it coincides with the occurrences of ‘Plagiogmus’ (Psammichnites gigas arcuatus) at Hines Ridge (extrapolated from Mollie Gibson; Corsetti and Kaufman, Reference Corsetti and Kauffman1994, fig. 2), Mollie Gibson (Fig. 1.3; Smith et al., Reference Smith, Nelson, O'Connell, Eyster and Lonsdale2023, fig. 7, MGM), and Mount Dunfee (Oliver, Reference Oliver1990, fig. 3; Oliver and Rowland, Reference Oliver, Rowland and Corsetti2002; Smith et al., Reference Smith, Nelson, Strange, Eyster, Rowland, Schrag and Macdonald2016, fig. 1; Nelson and Smith, Reference Nelson and Smith2019, fig. 1).
Tracing this negative excursion southward into the Death Valley facies is not straightforward because the section thins appreciably and there are far fewer carbonates. Topper et al. (Reference Topper, Betts, Dorjnamjaa, Li, Li, Altanshagai, Enkhbaatar and Skovsted2022, fig. 17) followed earlier suggestions (Corsetti and Kaufman, Reference Corsetti and Kauffman1994; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2000, Reference Corsetti and Hagadorn2003; Corsetti et al., Reference Corsetti, Awramik, Pierce and Kaufman2000) in allocating the nadir to the middle dolostone and attributing falling and rising limbs to the lower and upper dolostones, respectively. However, Nelson and Smith (Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023, fig. 1) placed the peak at (or just above?) the top of the first major dolostone in the Boundary Canyon and Montgomery Mountains sections, presumably on the assumption that this is a disconformable surface. In this interpretation, the interval of the LMWCF yielding Tulaneia amabilia n. gen. n. sp. and Saarina hagadorni Selly et al., Reference Selly, Schiffbauerb, Jacquet, Smith and Nelson2020, predates the nadir and is latest Ediacaran in age; the remainder of the LMWCF postdates the nadir and is therefore Cambrian in age, for reasons already given. However, in order to reconcile the Spring and Montgomery Mountains sections of the LMWCF, it was necessary for Nelson et al. (Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023, fig. 1) to postulate that the lowest dolostone in the Montgomery Mountains preserves only the declining limb of the BACE excursion, whereas the same dolostone in the adjacent (~20 km away) Spring Mountains section records only part of its post-nadir rise.
If this correlation between the White-Inyo and Death Valley facies is correct, then in the Spring Mountains and nearby exposures of the LMWCF, ca. 6.6 Ma of time must be compressed into 30–50 m of stratigraphic section (Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023, fig. 1). Given the complexity of the Fortunian carbon isotope record in Siberia, with three positive (2p–4p) and three negative (2n–4n) events before the positive excursion (5p) near the Fortunian–Stage 2 boundary (Kouchinsky et al., Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007; Zhuravlev et al., Reference Zhuravlev, Wood and Bowyer2023), it is unlikely that any particular named excursion could be recognized in the sparsely sampled and probably incomplete carbon isotope record of the LMWCF. The suggestion (Nelson et al., Reference Nelson, Crowley, Smith, Schwartz, Hodgin and Schmitz2023, fig. 1) that the whole of the Cambrian part of the LMWCF represents merely the 1n to 2p secular increase in δ13C, as seen, for example, in the Sukharihka section of the Siberian platform (Kouchinsky et al., Reference Kouchinsky, Bengtson, Pavlov, Runnegar, Torssander, Young and Ziegler2007, fig. 2), is unrealistic in light of the U–Pb age constraints. Given these uncertainties, we prefer to defer to lithological and paleontological criteria to correlate the White-Inyo and Death Valley successions, but note that the ca. 539 Ma age for the BACE nadir in Mexico corresponds to the middle (Esmeralda Member) of the Deep Spring Formation in California and Nevada, and that the nadir of the BACE excursion is unlikely to be older than the top of the first major dolostone of the LMWCF, according to traditional lithostratigraphic evidence (Stewart, Reference Stewart1966, Reference Stewart1970). There are also two paleontological matters that are worthy of consideration in this context. First, could Psammichnites gigas arcuatus have appeared during the Ediacaran? And second, could Tulaneia amabilia n. gen. n. sp. be Cambrian in age?
Mángano et al. (Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022) noted that the first appearance of Psammichnites gigas circularis is one of the diagnostic elements of the [early Fortunian] Rusophycus avalonensis Zone. Psammichnites gigas arcuatus, which only differs from P. gigas circularis in having the ladder-like burrow base (Fig. 12.1), occurs in the Fortunian of Siberia (Mángano et al., Reference Mángano, Buatois, MacNaughton, Jensen, Gougeon, Marcos, Meek, Piñuela and García-Ramos2022, table 3) and possibly in the earliest Fortunian (parasequence 3 of the Ingta Formation) in Canada (Carbone and Narbonne, Reference Carbone and Narbonne2014, fig. 4.1). An allied ichnofossil, Parapsammichites pretzeliformis, has been found in the undisputedly Ediacaran part of the Spitskop Member of the Schwarzrand Subgroup of Namibia (Buatois et al., Reference Buatois, Almond, Mángano, Jensen and Germs2018), so this degree of behavioral complexity began well before the Cambrian. Thus, we cannot rule out an Ediacaran age for the Deep Spring Formation ‘Plagiogmus’, but the default hypothesis must be that it is Cambrian or younger until proven otherwise.
There are also suggestions that some erniettomorphs may have survived into the Cambrian (Jensen et al., Reference Jensen, Gehling and Droser1998; Hagadorn et al., Reference Hagadorn, Schellenberg and Bottjer2000; Hoyal Cuthill, Reference Hoyal Cuthill2022; Mussini and Dunn, Reference Mussini and Dunn2024; but see Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024), but if Tulaneia amabilia n. gen. n. sp. is restricted to the interval below the first major dolostone of the LMWCF, as seems likely, the nature of the associated body and trace fossils makes an earliest Cambrian age both implausible and unnecessary. In summary, all available evidence suggests that Tulaneia amabilia n. gen. n. sp. is latest Ediacaran in age, the Ediacaran–Cambrian boundary in the Death Valley succession coincides with the top of the first major dolostone of the LMWCF, and that the remainder of the LMWCF spans most of the early Cambrian Fortunian Stage.
Great Unconformity
Peters and Gaines (Reference Peters and Gaines2012) showcased the ‘Great Unconformity’ at the base of the Sauk megasequence as a possible trigger for the Cambrian explosion. They noted that the base of the Sauk is diachronous (Sloss, Reference Sloss1963, Reference Sloss1988), but argued that it is a unique, globally occurring stratigraphic surface that, through unprecedented chemical weathering and erosion, may have modified ocean water chemistry enough to have influenced the evolution of biomineralization. The basis for this hypothesis was reviewed by Shahkarami et al. (Reference Shahkarami, Buatois, Mángano and Hagadorn2020), who concluded that the unconformable ‘surface’ has multiple local origins and should not be used for global correlation. We mention the hypothesis here because of the new, late Fortunian age, for the top of the LMWCF and, therefore, the base of the Sauk megasequence at the craton margin, which was well separated in time from the Ediacaran–Cambrian boundary in this area.
Erniettomorphs
Because it has long been confused with Ernietta, there seems little doubt that Tulaneia amabilia n. gen. n. sp. is an erniettomorph, as defined in Runnegar et al. (Reference Runnegar, Gehling, Jensen and Saltzman2024). Diagnostic features are all related to the modular, tubular construction, seen also in Ernietta (Ivantsov et al., Reference Ivantsov, Narbonne, Trusler, Greentree and Vickers-Rich2016), Pteridinium (Elliott et al., Reference Elliott, Trusler, Narbonne, Vickers-Rich, Morton, Hall, Hoffmann and Schneider2016; Darroch et al., Reference Darroch, Gibson, Syversen, Rahman, Racicot, Dunn, Gutarra, Schindler, Wehrmann and Laflamme2022; Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024), and Phyllozoon (Gehling and Runnegar, Reference Gehling and Runnegar2022), each of which is readily distinguished by its unique geometry. Unfortunately, the addition of Tulaneia n. gen. to the clade provides little enlightenment as to the nature of their bodies, their modes of life, or their place in the tree of life. Our best guess is that Tulaneia n. gen. housed facultative or obligate bacterial symbionts that used sunlight and/or reduced compounds plus oxygen to grow, reproduce, and to supply nutrients to the host organism. We speculate that Tulaneia n. gen. was epifaunal, attached somehow at its basal seam, and inhabited nearshore sandy bottoms well within the photic zone. As to its place in the tree of life, the simplicity of its morphology, even when compared with Ernietta, may indicate a position lower in the metazoan tree than the crown group, if these organisms are indeed metazoans.
Acknowledgments
Funding was provided by the U.S. National Science Foundation (EAR-9627924), the Division of Physical Sciences, UCLA, the Leverhulme Trust, and the U.K. National Environment Research Council. We thank J. Hook, Los Angeles County Museum, Invertebrate Paleontology, and H. Mocke, National Earth Sciences Museum, Windhoek, Namibia, for assistance with the donation and curation of the illustrated material. M. Laflamme and G. Narbonne are thanked for their thorough, thoughtful, and helpful reviews.
Competing interests
The authors declare none.
Author contributions
Field work, guided initially by RJH, was carried out in January 1993 by JGG, RJH, and BR; in March and April 1993 by SB, JGG, RJH, SJ, KJP, BR, and MRS; and in March 1995 by KJP, BR, MRS, and MJV; stratigraphic sections were measured by JGG and SJ. The fossils were studied at UCLA by JGG, SJ, and BR; BR carried out nearly all of the photographic work, prepared the illustrations, and wrote the manuscript; all authors reviewed the manuscript and contributed to its final form.
Data availability statement
Supplemental dataset 1.—Tabulated stratigraphic measurements and occurrences. The columns filled with numbers represent measured stratigraphic sections, which are labeled and referenced accordingly; the gray-shaded cells refer to specimens that are illustrated in this work or in the literature, as shown in order from left to right; the stratigraphic heights of the three main dolostones of the LMWCF are highlighted in orange to enable visual correlation of the measured sections; heights in sections are in meters and are taken from field records and/or the cited references. Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.f1vhhmh4s.
Appendix 1: descriptions of fossil localities used for this study
Mollie Gibson, Inyo County, California
UCLA 6153/LACMIP 26153 (Fig. 1.3; Alpert, Reference Alpert1974).—Esmeralda Member, Deep Spring Formation at intersection of mining road and stream bed, ~230 m E and ~190 m N of SW corner sec. 15, T7S, R35E, ~1.4 km SE of the Mollie Gibson mines (37.344538°N, 118.142230°W), Blanco Mountain Quadrangle, California; Cambrian (Fortunian).
Psammichnites gigas arcuatus (Roedel, Reference Roedel1929) (Fig. 12.2; Alpert, Reference Alpert1974, pl. 5, figs 3, 11, 12).
UCLA 6160/LACMIP 26160 (Fig. 1.3; locality 6 of Cloud and Nelson, Reference Cloud and Nelson1966, fig. 3; Alpert, Reference Alpert1974).—Esmeralda Member, Deep Spring Formation, NE¼, NW¼, SE¼, sec. 16, T7S, R35E, ~600 m SE of Mollie Gibson mines (37.344538°N, 118.142230°W), Blanco Mountain Quadrangle, California; Cambrian (Fortunian).
Psammichnites gigas arcuatus (Roedel, Reference Roedel1929) (Fig. 12.2 right, 12.3; Cloud and Nelson, Reference Cloud and Nelson1966, fig. 1B; Cloud and Bever, Reference Cloud and Bever1973, pl. 1, fig. 5; Alpert, Reference Alpert1974, pl. 5, fig. 4).
Hines Ridge, Inyo County, California
UCLA 6156/LACMIP 43374 (Fig. 1.3).—Wyman Formation and Hines member of the Reed Dolomite, S of Waucoba Road and W of Hines Road (37.1020°N, 118.0900°W), sec. 6, T10S, R36E, Waucoba Mountain Quadrangle, California; Ediacaran.
?Gordia isp. (Fig. 9.1, 9.2; Langille, Reference Langille1973, Reference Langille1974; Corsetti and Hagadorn, Reference Corsetti and Hagadorn2003, fig. 5A).
Mount Dunfee, Esmeralda County, Nevada
JHU 1645, 1648, 1649 (Fig. 1.4; Nelson and Smith, Reference Nelson and Smith2019, fig. DR1).—Esmeralda Member, Deep Spring Formation, near Mount Dunfee (37.3429°N, 117.3278°W), Esmeralda County, Nevada; Cambrian (Fortunian).
Psammichnites gigas arcuatus (Roedel, Reference Roedel1929) (Fig. 12.1, 12.5; Nelson and Smith, Reference Nelson and Smith2019, fig. 2A–F, fig. 4A, B; Nelson and Smith, Reference Nelson and Smith2020, fig. 1).
Chicago Pass, Inyo County, California
UCLA 7329/LACMIP 43370 (Figs 1.1, 4, Supplemental dataset 1).—Lower member of the Wood Canyon Formation, Chicago Pass (36.1213°N, 116.1788°W) on the Charles Brown Highway (CA route 178), northern end of Nopah Range, Inyo County, California; Ediacaran–Cambrian (Fortunian).
Didymaulichnus miettensis Young, Reference Young1972 (Fig. 11.1).
Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.7; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002, fig. 4A).
Treptichnus isp. cf. T. pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955).
Spring Mountains, Nye County, Nevada
UCLA 7330/LACMIP 43368 (Figs. 1.1, 4, Supplemental dataset 1).—Lower member of Wood Canyon Formation, first wash S of Wood Canyon (36.3852°N, 115.9210°W), Spring Mountains, Nye County, Nevada; Ediacaran–Cambrian (Fortunian).
Annulated tubular fossils (Fig. 4.5).
Desert Range, Nye County, Nevada
UCLA 7331/LACMIP 43371 (Fig. 1.1, 4, Supplemental dataset 1; Gillett and Van Alstine, Reference Gillett and Van Alstine1979).—Lower member, Wood Canyon Formation, ~800 m WSW of National Geodetic Survey benchmark GR0872 (Q 313 1941), Desert National Wildlife Range (36.8039°N, 115.3222°W), Clark County, Nevada, Ediacaran–Cambrian (Fortunian).
Annulated tubular fossils.
Treptichnus isp. cf. T. pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.1).
Montgomery Mountains, Nye County, Nevada
UCLA 7333/LACMIP 43369 (Figs. 1.2, 4, Supplemental dataset 1; Horodyski, Reference Horodyski1991; Horodyski et al., Reference Horodyski, Gehling, Jensen and Runnegar1994).—Lower member of the Wood Canyon Formation, 2.8 km WNW of Johnnie (36.419967°N, 116.072604°W), Nye County, Nevada, Ediacaran–Cambrian (Fortunian).
Tulaneia amabilia Runnegar and Horodyski n. gen. n. sp. (Figs. 5, 6.1–6.7, 7.1–7.9).
Annulated tubular fossils (Fig. 4.6–4.8).
Didymaulichnus miettensis Young (Reference Young1972) (Fig. 11.3; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002, fig. 5A).
Gordia isp. (Fig. 9.4).
Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.2, 10.6).
Treptichnus isp. cf. T. pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.3).
Treptichnus spectrum traces (Figs. 9.5, 9.6, 10.3, 11.6).
UCLA 7334/LACMIP 43372 (Fig. 1.2, Supplemental dataset 1; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002).—Lower member of the Wood Canyon Formation, 3.1 km W of Johnnie (36.419967°N, 116.072604°W), Nye County, Nevada, Ediacaran–Cambrian (Fortunian).
Rusophycus isp. (Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002, fig. 4C, 4D).
Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.2, 10.6; Jensen et al., Reference Jensen, Droser, Heim and Corsetti2002, fig. 4B).
Treptichnus spectrum traces (Fig. 9.6).
NHMLAC 17130/LACMIP 17130 (Fig. 1.1; Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000; Smith et al., Reference Smith, Nelson, Tweedt, Zeng and Workman2017; Selly et al., Reference Selly, Schiffbauerb, Jacquet, Smith and Nelson2020).—Lower member of the Wood Canyon Formation, SW¼, NW¼, SW¼, sec. 11, T18S, R52E, Mt. Schader Quadrangle, 3.4 km SW of Johnnie, Nye County, Nevada; Ediacaran–Cambrian (Fortunian).
Saarina hagadorni Selley et al., Reference Selly, Schiffbauerb, Jacquet, Smith and Nelson2020 (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000, fig. 3.7–3.9; Selley et al., Reference Selly, Schiffbauerb, Jacquet, Smith and Nelson2020, figs. 2B, 2E, 3E)
?Swartpuntia germsi Narbonne, Saylor, and Grotzinger, Reference Narbonne, Saylor and Grotzinger1997 (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000, fig. 4.1–4.3).
?Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Hagadorn and Waggoner, Reference Hagadorn and Waggoner2000, fig. 5.8, 5.9).
Ross River, Northern Territory, Australia
UCLA 7346/LACMIP 43373.—Glauconitic sandstones of Member IV of the Arumbera Formation (23.5956°S, 134.4718°E), ~1 km W of the Ross River resort, ~40 km E of Alice Springs, Northern Territory, Australia; Cambrian (Terraneuvian).
Didymaulichnus miettensis Young, Reference Young1972 (Fig. 11.2).
Psammichnites gigas arcuatus (Roedel, Reference Roedel1929) (Fig. 12.4; McIlroy and Heys, 1997).
Ediacara, South Australia
UNE L1765.—Thinly bedded quartz sandstones near the top of the Ediacara Member, Rawnsley Quartzite on ridge (30.7800°S, 138.1468°E) S of track from Sundown bore to Gap well, Ediacara Hills, W of Beltana, South Australia; Ediacaran.
Gordia isp. (Fig. 9.3).
Nama Basin, Namibia
UCLA 7322 Swartkloofberg 1 (Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024).—Thin sandstones, Nasep Member, Urusis Formation, Schwarzrand Subgroup, Nama Group on dip slope immediately N of Swartkloofberg homestead (27.485491°S, 16.523883°E), Swartkloofberg farm, Rekvlakte 1:50,000 map sheet (2716BC), southern Namibia; Ediacaran.
Pteridinium carolinaensis (St. Jean, Reference St. Jean1973) (Fig. 6.8–6.10).
UCLA 7324 Sonntagsbrunn (Runnegar et al., Reference Runnegar, Gehling, Jensen and Saltzman2024).—Thin event beds in siltstones of the Kreyrivier Member, Nomtsas Formation, Schwarzrand Subgroup, Nama Group on E and W sides of hill 850 m on E side of road D463 (27.368114°S, 17.509523°E) ~1.5 km S of Koedoeslaagte homestead, W of the Fish River Canyon, Koedoeslaagte 1:50,000 map sheet (2717DA), southern Namibia; Cambrian (Fortunian).
Treptichnus pedum (Seilacher, Reference Seilacher, Schindewolf and Seilacher1955) (Fig. 10.8).













 Open access
Open access