

Fetal programming is a phenomenon characterised by adaptive responses of the fetus to specific environmental conditions (e.g. nutrition) during early life stages, which may alter gene expression and permanently affect the structure and function of several organs and tissues, most likely influencing the individual’s susceptibility to develop metabolic disorders over a lifetime, such as type 2 diabetes, CVD, obesity and hypertension( Reference Barker 1 , Reference Spencer 2 , Reference Bruce and Hanson 3 ).
Dietary fatty acids (FA) are one of the nutritional environmental factors that are capable of inducing changes in development and fetal programming( Reference Innis 4 , Reference Aoun, Feillet-Coudray and Fouret 5 , Reference Mennitti, Oliveira and Morais 6 ). As integral components of the phospholipid structure of biological membranes, FA play pivotal roles in cell function and tissue growth, modulation of intercellular and intracellular communication, control of gene expression and epigenetic regulation( Reference Chen and Bazan 7 , Reference Tontonoz and Spiegelman 8 , Reference Burdge and Lillycrop 9 ).
Studies on trans-unsaturated fatty acids (TFA), present in partially hydrogenated vegetable oil, have shown an association between oxidative stress, elevated pro-inflammatory cytokines, insulin resistance and metabolic programming( Reference Mennitti, Oliveira and Morais 6 , Reference Osso, Moreira and Teixeira 10 ), indicating that the consumption of these FA during pregnancy and lactation should be reduced( Reference Anderson, McDougald and Steiner-Asiedu 11 ). The use of several industrialised food products rich in TFA has led many countries to regulate the use of partially hydrogenated vegetable fat in processed foods and search for alternative sources to maintain product consistency and quality. Palm oil (PO), a natural vegetal source of saturated fat, has been used in particular because of its texture and stability( Reference Oguntibeju, Esterhuyse and Truter 12 , Reference Ong and Goh 13 ). Another alternative is the use of interesterified fat (IF), which results in a modification of the position of the FA in the glycerol molecule, forming a new TAG( Reference Berry 14 ). However, such substitution has been conducted without further studies on the effects of these types of fats on the metabolism and on long-term health.
Recently, our group demonstrated, in a fetal programming model for mice, that the maternal intake of PO or IF may predispose the adult offspring to obesity, promoting the accumulation of total body fat and increasing adipocyte number and area( Reference Magri, Fernandes and Souza 15 ). In addition, Misan et al. ( Reference Misan, Estato and de Velasco 16 ) showed that the maternal intake of TFA, PO or IF can trigger the initial steps of inflammatory pathways in the brain of the offspring in adult life, increasing Toll-like receptor 4 signalling and leucocyte–endothelial interactions in cerebral microcirculation. These results provide consistent evidence that the type of dietary FA consumed by dams during the perinatal period is an important dietary/environmental factor involved in the metabolic disturbances observed in later life.
Although the molecular mechanisms by which maternal lipid intake affects long-term health of the offspring are still not completely understood, it was demonstrated that alterations in mitochondrial function in different tissues play important roles in metabolic disturbances( Reference Putti, Sica and Migliaccio 17 ). FA are also of crucial importance in the composition of mitochondrial membranes, affecting mitochondria structure and function. The saturation of FA could generate disturbances in mitochondrial membrane fluidity, modifications in respiratory functions and the organisation of chain complexes, which may result in an increased production of reactive O2 species (ROS)( Reference Yu, Fink and Herlein 18 ) or mitochondrial permeability transition pore (MPTP) in which necrotic cell death may lead to pro-inflammatory states( Reference Khairallah, O’Shea and Brown 19 , Reference Ronchi, Vercesi and Castilho 20 ).
High-fat( Reference Kahle, Schafer and Seelig 21 ) and high-lard diets( Reference Lionetti, Mollica and Donizzetti 22 ) impaired mitochondrial functions by altering the lipid composition of liver mitochondria in mice. In addition, mice fed a high-fat diet presented a decrease in respiration coupled to ATP synthesis and a decrease in ATP content in liver mitochondria( Reference Putti, Sica and Migliaccio 17 , Reference Fink, Herlein and Almind 23 ). These effects seem to be mediated by an increase in ROS production. Moreover, altered mitochondrial fission and fusion( Reference Borengasser, Faske and Kang 24 ) and decreased FA oxidation( Reference Hellgren, Jensen and Waterstradt 25 ) are some possible mechanisms related to high-fat-diet-induced metabolic programming.
To the best of our knowledge, no studies have examined the effects of dietary palm and IF – products used to replace partially hydrogenated fats in processed foods – on mitochondrial bioenergetics, particularly during the critical developmental period of the liver. Thus, the purpose of the present study was to investigate whether normolipidic diets containing TFA-rich hydrogenated fat, or its industrial substitute lipid sources, during pregnancy and lactation can predispose the adult offspring to metabolic changes and mitochondrial dysfunction, by evaluating liver bioenergetics, FA profiles and parameters of oxidative stress.
Methods
Animals
C57BL/6 mice (20–25 g) were obtained from the animal breeding unit at the Institute of Nutrition of the Federal University of Rio de Janeiro (UFRJ) and were maintained under humidity- and temperature-controlled (24±1°C) conditions with a 12 h light–12 h dark cycle and ad libitum access to water and diets.
Experimental procedures followed the national guidelines for Animal Experimentation, and The Experimental Research Committee of the Federal University of Rio de Janeiro (Rio de Janeiro, Brazil) approved all the procedures involving animals, under the reference no. INJC011 (CEUA/UFRJ).
Experimental design
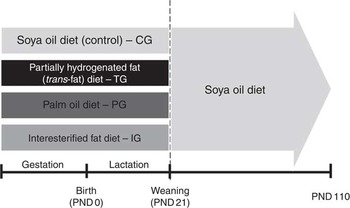
Female mice, 3-month-old, were left overnight to mate, and copulation was verified by the presence of sperm on the vaginal smears, as previously described by Misan et al. ( Reference Misan, Estato and de Velasco 16 ). On the 1st day of pregnancy, dams were isolated in individual cages and randomly divided into four groups. The control group (CG; 6–8/group) received a diet containing 7 % of soyabean oil. The partially hydrogenated vegetable oil rich in trans-fatty acid group (TG; 6–8/group) received 6 % partially hydrogenated vegetable oil plus 1 % soyabean oil. The palm oil group (PG; 6–8/group) received 5 % PO plus 2 % soyabean oil, and the interesterified fat group (IG; 6–8/group) received 5 % IF plus 2 % soyabean oil. Dams were fed their respective group diets throughout pregnancy and lactation. On the day of birth, considered day 0 of lactation, each mother was kept with her six to eight pups. At postnatal day 21, litters were weaned and male pups from each group had their diet changed to the control diet (soyabean oil) until the 110th day of life (Fig. 1).
Summary of experimental design. On the 1st day of pregnancy, dams were divided in groups and received diets with different sources of fat: soyabean oil (control diet – CG), partially hydrogenated vegetable oil rich in trans-fatty acids (TG), palm oil (PG) and interesterified fat (IG). Dams were fed each diet during pregnancy, and after delivery litters received diets through lactation. After weaning (postnatal day 21 – PND21), litters from all experimental groups were fed control diet until PND110 (adulthood), when analyses were performed.
Diets
All diets were isoenergetic (providing 17·2 kJ/g of dry diet) and differed from each other only in the nature of lipid source. The diet compositions followed the recommendations of the American Institute of Nutrition (AIN-93)( Reference Reeves, Nielsen and Fahey 26 ), as described in Misan et al.( Reference Misan, Estato and de Velasco 16 ). The composition of the experimental diets is shown in Table 1.
Composition of diets (g/kg dry diet)
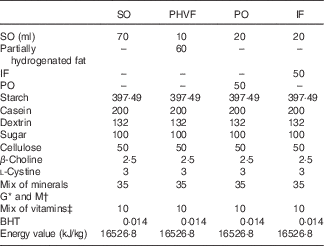
SO, soyabean oil; PHVF, partially hydrogenated vegetable fat; PO, palm oil; IF, interesterified fat; BHT, butylhydroquinone.
* Salt mix G (mg/kg diet) for growth, pregnancy and lactation periods: Ca, 5000·0; P, 1561·0; K, 3600·0; S, 300·0; Na, 1019·0; Cl, 1571·0; Mg, 507·0; Fe, 35·0; Zn, 30·0; Mn, 10·0; Cu, 6·0; I, 0·2; Mo, 0·15; Se, 0·15; Si, 5·0; Cr, 1·0; F, 1·0; Ni, 0·5; B, 0·5; Li, 0·1; V, 0·1.
† Salt mix M (mg/kg diet) for maintenance periods: Ca, 5000·0; P, 1992·0; K, 3600·0; S, 300·0; Na, 1019·0; Cl, 1571·0; Mg, 507·0; Fe, 35·0; Zn, 30·0; Mn, 10·0; Cu, 6·0; I, 0·2; Mo, 0·15; Se, 0·15; Si, 5·0; Cr, 1·0; F, 1·0; Ni, 0·5; B, 0·5; Li, 0·1; V, 0·1.
‡ Vitamin mix (mg/kg diet): retinyl palmitate, 2·4; cholecalciferol, 0·025; menadione sodium bisulfite, 0·8; biotin, 0·22; cyanocobalamin, 0·01; riboflavin, 6·6; thiamin hydrochloride, 6·6; tocopheryl acetate, 100.
Soyabean oil (ml) was added to each diet to adjust the essential FA requirements in each diet. Diets were freshly prepared every 3 d as pellets, gassed with N2 and stored at −20°C until use. All fats used in the diets were donated by INDÚSTRIA TRIÂNGULO ALIMENTOS LTDA. According to the company, soyabean oil is used for the formulation of partially hydrogenated vegetable oil. Four different types of lipids, palm kernel oil (2·5 %), palm stearin (45 %), soyabean oil (45 %) and fully hydrogenated fat (7·5 %), were used to prepare the IF.
The FA composition of dietary fats was determined by GC and is presented in Table 2. The methylated FA were identified based on a comparison with the relative retention time of the standard peaks. The characterisation of each FA was performed by area normalisation, expressed as a percentage of the total FA (mean and sem/ n 6). The trans-fat experimental diet had increased levels of trans-fatty acids when compared with all the diets offered. The PO, trans-fat and IF diets had more SFA when compared with the control diet. In addition, all experimental diets had lower levels of total PUFA, mostly from the n-3 and n-6 families( Reference Magri, Fernandes and Souza 15 ).
Fatty acid profile (% of total) of the experimental diets (Mean values with their standard errors)
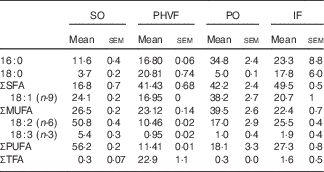
SO, soyabean oil; PHVF, partially hydrogenated vegetable fat; PO, palm oil; IF, interesterified fat.
Determination of plasma glucose and TAG
Blood was obtained from male mice, after an 8-h fast, anaesthetised with ketamine and xilazine by cardiac puncturing. Blood was then distributed in tubes (1 ml) with EDTA and centrifuged at 3000 rpm for 15 min at 4°C and the plasma was collected and stored at −80°C. For determining the plasma glucose and TAG levels, we used enzymatic colorimetric kits (BIOCLIN). All samples (10 µl) were well homogenised with the specific reagents, put in a water bath at 37°C for 10 min and then placed into microplates for absorbance reading. The absorbance of samples and standard was performed at 500 nm.The plasma glucose and TAG were calculated by multiplying the sample absorbance per 100 and dividing by standard absorbance. The data were expressed in mmol/l.
Determination of liver fatty acids profile
The relative content of FA from the total lipids from male offspring livers was determined by GC, using an Agilent Technologies 7890A GC System, equipped with a flame ionisation detector, coupled to EZChrom Elite CDS software (Agilent Technologies Inc.). The lipid extraction, saponification and methylation of FA were performed by direct methylation, according to the official method of the American Oil Chemists’ Society, and implemented in the laboratory. The methyl esters were separated using a fused silica capillary column, SP-2560 bis-cyanopropylpolysiloxane (Supelco Inc.) with an internal diameter of 100 m×0·25 mm×0·2 µm, whose programming column temperature was set to 100°C with a gradual increase of 3°C/min until it reached 140°C. The temperature increased at a rate of 0·5°C/min until it reached 170°C, and then 3·2°C/min until it reached 220°C and was then kept at this temperature for 35 min. The injector and detector temperatures were 250°C. The flow of gases (Linde Gases) was 0·75 ml/min for the carrier gas (H2), 25 ml/min for the auxiliary gas (N2) and 30 ml/min and 400 ml/min for H2 and synthetic air to the flame, respectively. The splitter reason (split) was 1/100. The injections were performed with an automatic injector and the volume was 1 μl.
FA methyl esters were identified on the basis of a comparison of the relative retention time of the peaks of standards from Nu-Chek Prep Inc., a blend of methyl esters 463, and the quantification of each FA was carried out by normalisation of areas, expressed as a percentage of total FA.
Isolation of mitochondria
Animal and liver weights were collected. After that, liver samples were placed on ice during the procedures. About 1 g of tissue was placed in an isolation buffer that contained mannitol 225 mm, sucrose 75 mm, EGTA (ethylene glycol-bis(2-aminoethylether)-N,N,N′,N′-tetraacetic acid) 1 mm, bovine serum albumin (BSA) 0·1 % and HEPES 10 mm (pH 7·2) at 4°C. The mitochondria from mouse livers were isolated by differential centrifugation with modifications( Reference Frezza, Cipolat and Scorrano 27 , Reference Meyer, Machado and Santiago 28 ). Tissues were sliced and washed in the isolation buffer. After four washes, livers were homogenised by hand in a glass potter with a Teflon piston rotated for six cycles. The homogenates were centrifuged (Hitachi) at 200 g for 5 min. The supernatants were carefully separated and then centrifuged at 10 000 g for 8 min. After that, the supernatants were discarded and the precipitants were resuspended in the isolation buffer and were considered as the mitochondrial fractions. Protein concentration was determined using BSA as a standard( Reference Lowry, Rosebrough and Farr 29 )and the mitochondrial fractions ranged from 19 to 23 mg/ml.
VO2 measurements and mitochondrial bioenergetics parameters
VO2 was measured with the polarography technique using high-resolution respirometry (Oxygraph-O2K; Oroboros Instruments®). Measurements were performed in a chamber with controlled temperature (37°C±0·001°C) and the electrode was calibrated for 0–100 % of saturation (220 μM O2) and atmospheric pressure (101·3 kPa). Mouse liver mitochondrial fractions (0·5 mg/ml) were incubated with 2·0 ml of respiratory buffer MIR05 containing sucrose 110 mm, EGTA 0·5 mm, MgCl2 3 mm, MES 60 mm, Taurine 20 mm, KH2PO4 10 mm, HEPES 20 mM and BSA 0·1 % (pH 7·2), under constant shaking( Reference Gnaiger, Kuznetsov and Schneeberger 30 ). VO2 rates were monitored according to the protocol described in Pesta & Gnaiger with modifications( Reference Pesta and Gnaiger 31 ), using a sequential addition of substrates from complex I (pyruvate/malate/glutamate (PMG), 2 mm each) or complex II (succinate (SUCC) 2 mm), followed by ADP 100 µm, ADP 1 mm, oligomycin 2 µg/ml, carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) 0·25 µm, FCCP 0·25 µm, rotenone 0·5 µm or malonate 5 mm, and potassium cyanide (KCN) 2 mm. Results were expressed in pmol O2/mg of protein×s( Reference Ramos-Filho, Chicaybam and de-Souza-Ferreira 32 ).
Citrate synthase enzyme activity
Citrate synthase (CS) activity was measured in isolated liver mitochondria samples, as previously described, with minor modifications( Reference Ramos-Filho, Chicaybam and de-Souza-Ferreira 32 , Reference Teodoro, Palmeira and Rolo 33 ). The reaction medium contained 20 mm TRIS-HCl, pH 8, 0·3 mm 5,5'-dithio-bis (2-nitrobenzoic acid) (DTNB), 2 mm EDTA, 0·3 mm acetyl coenzyme A and 100 μg/ml protein. The medium was incubated for 5 min with continuous shaking at 37°C and the reaction was initiated with the addition of 5 mm oxaloacetate. The reactions were performed in a 96-well plate, and the results were read at 420 nm in a Victor-Perkin Elmer plate reader. The specific activity was calculated based on a curve of CoA in the presence of 0·3 mm DTNB and was expressed as μmol citrate/min per mg protein.
Hydrogen peroxide production in liver mitochondria
Mitochondrial release of hydrogen peroxide (H2O2) was determined using the Amplex Red® (N-acetyl-3, 7-dihydroxyphenoxazine) oxidation method( Reference Starkov 34 ). Liver mitochondrial fractions from mice (0·5 mg protein/ml) were incubated in the MIR05 (pH 7·2) buffer, supplemented with 5 μm Amplex Red and 4 units/ml horseradish peroxidase. The fluorescence was monitored with a Spectrofluorimeter (Varian Cary Eclipse®; Agilent Technologies) at 563-nm excitation (slit 5 nm) and 587-nm emission wavelengths (slit 5 nm) under constant shaking at 37°C( Reference Meyer, Machado and Santiago 28 , Reference Ramos-Filho, Chicaybam and de-Souza-Ferreira 32 ). The rate of mitochondrial H2O2 production was assessed through a standard curve with known amounts of H2O2. Results were expressed in pmol H2O2/mg of protein×min. The experiments were done with sequential addition of substrates from complex I (PMG, 2 mm each) or complex II (SUCC 2 mm), followed by 0·5 mM ADP, 2 µg/ml oligomycin, 0·5 µm FCCP and 5 µm antimycin A.
Determination of electron leakage
We estimated the fraction of electrons that leaked out of the respiratory chain by dividing the rate of H2O2 formation by the rate of VO2 ( Reference Ramos-Filho, Chicaybam and de-Souza-Ferreira 32 , Reference Santiago, Chaves and Oliveira 35 ). To calculate electron leakage, these measurements were expressed using the same units (pmol/min×mg) at the same respiratory state. The electron leakage related to a specific substrate was calculated as the ratio of H2O2 formed by a specific substrate (for example, SUCC) to VO2 with that substrate.
Determination of calcium retention capacity
The capacity to retain Ca2+ was determined using the method proposed by Wong et al.( Reference Wong, Steenbergen and Murphy 36 ). Mitochondrial fractions of mouse liver were incubated in MIR05 buffer without EGTA (pH 7·2) at 37°C, with the addition of Calcium Green 0·2 µm (Invitrogen®), a fluorescent marker that indicates the presence of Ca2+ in the medium. After that, the respective substrates from Complex I and II were added, followed by successive additions of 30 μm calcium chloride (CaCl2). Experiments were conducted in a Spectrofluorometer (Varian Cary Eclipse®) at 506-nm excitation (slit 10 nm) and 532-nm emission (slit 10 nm) wavelengths under constant shaking at 37°C.
Statistical analysis
VO2 measurements were analysed using DatLab 5 software. The results of H2O2 production and Ca2+ retention capacity were analysed using Origin 8.0 software. For statistical analysis of the data obtained in each of the groups, we used GraphPad Prism 6.0 software, with which we performed an ANOVA. Values were expressed as means with their standard errors. To compare the average results obtained between groups, we applied post-test Newman–Keuls or Dunnett’s tests for multiple comparisons, adopting a significance level of P<0·05, as a parameter for determining the differences between the groups. Sample size was calculated to provide 5 % error and 95 % confidence.
Results
Differences in total body weight, liver weight and plasma glucose and TAG levels in adult male mouse offspring
At 110 d of life, the groups did not present differences in absolute weight or in liver weight (Table 3). On the other hand, when we analysed the liver/body weight ratio, we observed increased ratios in the groups in which mothers were fed TG or IG diets.
Total body weight, liver weight and plasma glucose and TAG levels of male offspring at 110 d of life (Mean values with their standard errors, n 7–8)
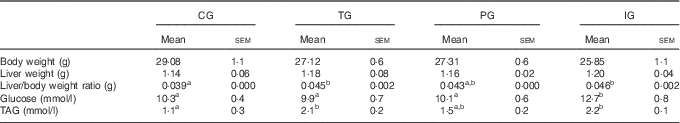
CG, control group; TG, trans-fat group; PG, palm oil group; IG, interesterified fat group.
a,b Mean values with unlike superscript letters were significantly different between experimental groups (P<0·05 ANOVA, with Newman–Keuls post hoc test).
We also evaluated the plasma glucose and TAG concentrations of male offspring. The results, shown in Table 3, demonstrate that the IG had plasma glucose levels increased by 1·2-fold in relation to the CG. Moreover, TAG plasma concentration was 1·8-fold in the TG and 1·9-fold higher in the IG when compared with the CG.
Maternal consumption of trans-fat and its substitutes during pregnancy and lactation decreased MUFA and PUFA in offspring livers
The data presented in Table 4 indicate that the composition of ΣSFA in the livers of adult offspring did not differ among groups. However, palmitic acid (C16 : 0) was significantly lower in the TG and IG compared with the CG. The content of ΣMUFA decreased by 20, 16·9 and 15·3 % in the TG, PG and IG, respectively, when compared with the CG. The decrease in oleic acid (C18 : 1n-9) contributed significantly to these differences.
Liver fatty acid profile (% of total) from male offspring at 110 d of life (Mean values with their standard errors, n 7–8)

CG, control group; TG, trans-fat group; PG, palm oil group; IG, interesterified fat group; TFA, trans-fatty acids; LA, linoleic acid; ALA, α-linolenic acid; AA, arachidonic acid.
a,b,c Mean values with unlike superscript letters were significantly different between experimental groups (P<0·05 ANOVA, with Newman–Keuls post hoc test).
Regarding the total amount of PUFA in the liver, ΣPUFA in the TG and IG was significantly lower when compared with the CG and PG. In addition, ΣPUFA n-6 in the TG and IG was also reduced. The essential n-6 precursor linoleic acid (LA) was lower in the TG and IG, even though arachidonic acid (AA), an important LA product, did not differ among groups. Moreover, ΣPUFA n-3 did not vary significantly among the groups. On the other hand, essential n-3 α-linolenic acid (ALA) was considerably lower in the TG (by31·9 %), PG (by 19·4 %) and the IG (by 20·1 %) when compared with the CG. The decrease in ALA was associated with a decrease in its main n-3 derivatives, EPA content, in the TG and IG compared with the CG.
In addition, the ratios C18 : 1/C18 : 0 and C18 : 3n-6/C18 : 2n-6, which indicate Δ-9 and Δ-6 desaturation indexes, respectively, decreased significantly in all experimental groups (TG, PG and IG) when compared with the CG.
The source of dietary lipids during pregnancy and lactation caused mitochondrial changes in VO2 and respiratory states in the livers of adult offspring
To evaluate whether the maternal intake of fat substitutes for trans-fats could influence mitochondrial physiology, we used high-resolution respirometry to analyse samples of isolated liver mitochondria from male adult offspring to evaluate VO2 following the addition of respiratory substrates for Complex I (PMG) or Complex II (SUCC).
Fig. 2(a) shows representative VO2 rates of each experimental group in the presence of PMG. The values shown in Fig. 2(b) indicate that the TG exhibited lower rates of VO2 of all respiratory substrates studied, with significant differences in the presence of ADP and FCCP, in relation to the CG.
Oxygen flux through complex I in high-resolution respirometry. (a) Oxygen flux per mass after multi-substrate experiments from control group (CG) (![]() ), trans-fat group (TG) (
), trans-fat group (TG) (![]() ), palm oil group (PG) (
), palm oil group (PG) (![]() ) and interesterified fat group (IG) (
) and interesterified fat group (IG) (![]() ). (b) Quantitative analyses indicate that TG presented lower oxygen rates after sequential addition in relation to all experimental groups. Significant reduced values were seen in TG after ADP and carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) when compared with CG. Pyruvate/malate/glutamate (PMG, 2 mm each), ADP (100 µm), ADP (1 mm), oligomycin (Oligo, 2 µg/ml), FCCP (0·5 µm), rotenone (Rot, 0·5 μm) and potassium cyanide (KCN, 2 mm). Results were expressed in pmol oxygen/mg protein×s. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. * P<0·05.
). (b) Quantitative analyses indicate that TG presented lower oxygen rates after sequential addition in relation to all experimental groups. Significant reduced values were seen in TG after ADP and carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) when compared with CG. Pyruvate/malate/glutamate (PMG, 2 mm each), ADP (100 µm), ADP (1 mm), oligomycin (Oligo, 2 µg/ml), FCCP (0·5 µm), rotenone (Rot, 0·5 μm) and potassium cyanide (KCN, 2 mm). Results were expressed in pmol oxygen/mg protein×s. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. * P<0·05.

Moreover, when we analysed the O2 flux through complex II after SUCC addition, we also detected lower rates of VO2 in liver mitochondria from the TG (Fig. 3(a)). Quantitative results confirmed the observations, with the TG displaying the lowest rates throughout the additions, with significantly reduced values after FCCP, when compared with the PG (Fig. 3(b)).
Oxygen flux through complex II in high-resolution respirometry. (a) Oxygen flux per mass after multi-substrates experiments from control group (CG) (![]() ), trans-fat group (TG) (
), trans-fat group (TG) (![]() ), palm oil group (PG) (
), palm oil group (PG) (![]() ) and interesterified fat group (IG) (
) and interesterified fat group (IG) (![]() ). (b) Quantitative analyses indicate that after complex II stimulation with succinate (SUCC), TG also displayed reduced oxygen rates in relation to all groups. Statistically different values were seen in TG after carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) when compared with PG. SUCC (2 mm), ADP (100 µm), ADP (1 mm), oligomycin (Oligo, 2 µg/ml), FCCP (0·5 µm), malonate (Malo, 5 mM) and potassium cyanide (KCN, 2 mm). Results were expressed in pmol oxygen/mg of protein×s. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. * P<0·05.
). (b) Quantitative analyses indicate that after complex II stimulation with succinate (SUCC), TG also displayed reduced oxygen rates in relation to all groups. Statistically different values were seen in TG after carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) when compared with PG. SUCC (2 mm), ADP (100 µm), ADP (1 mm), oligomycin (Oligo, 2 µg/ml), FCCP (0·5 µm), malonate (Malo, 5 mM) and potassium cyanide (KCN, 2 mm). Results were expressed in pmol oxygen/mg of protein×s. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. * P<0·05.

Average respiratory rates and respiratory parameters are presented in the online Supplementary Tables S1 and S2 after PMG and SUCC addition. It can be observed that when PMG were the respiratory substrates, the TG presented a significant (56 %) reduction in the electron transfer system (ETS) capacity, which corresponded to maximal respiration recorded after FCCP titration, when compared with the CG (P=0·0354).
The results of respiratory parameters after complex II substrate addition indicated that the TG indeed presented important alterations in mitochondrial bioenergetics (online Supplementary Table S2). A significant (50·0 %) decrease in ETS capacity was observed in the TG in relation to the PG. In addition, the IG also displayed a slight reduction when compared with the CG and PG. On the other hand, contrary to what was observed after PMG addition, the PG presented similar respiratory rates to the CG. These findings suggest that the metabolic programming induced by trans-fat involved alterations in the control of coupled respiration by the phosphorylation system. Leak flux control ratio, which reflects the extent of intrinsic uncoupling, was not different among the groups (online Supplementary Table S2).
In an attempt to better understand the association between oxidative phosphorylation and ETS capacities of liver mitochondria in the context of metabolic programming, we subtracted the oxidative phosphorylation capacity rate from the ETS capacity rate (Fig. 4). By doing so, respiratory reserve capacity is represented as an absolute respiratory rate, as opposed to a fraction of maximal respiration. It is clear that the TG presented no reserve capacity when compared with all other groups (P=0·0207), when PMG were the respiratory substrates.
Reserve capacity from liver mitochondria of adult offspring. Quantitative results from VO2 rate (the difference of the electron transfer system capacity and oxidative phosphorylation capacity) showed that offspring that received, during pregnancy and lactation, trans-fat group (TG) presented no reserve capacity, suggesting that liver mitochondria from TG work under its maximum capacitance. Results were expressed in pmol oxygen/mg of protein×s. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. CG, control group; PG, palm oil group; IG, interesterified fat group. * P<0·05.

CS activity was measured in livers of mouse offspring as a marker of mitochondrial content. The results showed no differences among all experimental groups where the mean results were 51·40 (sem 3·19), 45·39 (sem 2·09), 46·81 (sem 4·45) and 43·29 (sem 1·8) nmol×mg/protein×min for the CG, TG, PG and IG, respectively. These results indicate that the effects on mitochondrial respiratory parameters were not related to the decreased mitochondrial contents in any of the experimental groups.
Hydrogen peroxide production rates from oxidative and non-oxidative phosphorylation in isolated liver mitochondria from adult offspring
Next, we evaluated H2O2 production in intact liver mitochondria. When complex II was activated by SUCC, H2O2 rates were significantly different (P=0·0013) in the IG, increasing 1·4-fold when compared with the CG (Fig. 5). In addition, the TG showed reduced levels after the sequential additions, which supports that trans-fat presented low production of H2O2 in liver mitochondria.
Hydrogen peroxide (H2O2) production using multiple titrations. (a) The representative graph of H2O2 rates measured after complex II stimulation with succinate (SUCC). Interesterified fat group (IG, ![]() ) increased H2O2 production when compared with all experimental groups. Trans-fat group (TG,
) increased H2O2 production when compared with all experimental groups. Trans-fat group (TG, ![]() ) presented reduced H2O2 production throughout multiple additions. (b) The quantitative results indicate that levels of H2O2 increased after substrate stimulation in IG. The experiments were done with sequential addition of substrates from SUCC (2 mm), followed by 0·5 mm ADP, 2 µg/ml oligomycin (Oligo), 0·5 µm carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) and 5 µm antimycin A (Ant A). Results were expressed in pmol H2O2/mg of protein×min. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars.
) presented reduced H2O2 production throughout multiple additions. (b) The quantitative results indicate that levels of H2O2 increased after substrate stimulation in IG. The experiments were done with sequential addition of substrates from SUCC (2 mm), followed by 0·5 mm ADP, 2 µg/ml oligomycin (Oligo), 0·5 µm carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP) and 5 µm antimycin A (Ant A). Results were expressed in pmol H2O2/mg of protein×min. Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. ![]() , control group (CG);
, control group (CG); ![]() , palm oil group (PG). * P<0·05.
, palm oil group (PG). * P<0·05.

H2O2 generation through mitochondria induced by complex I was undetectable, as we observed that the H2O2 flux under basal conditions and after PMG addition changed slightly (online Supplementary Fig. S1). This result suggests that the H2O2 flux detected could be related to production by a non-oxidative phosphorylation, which responds differently to that generally observed in functional mitochondria. Therefore, H2O2 generation by extra-mitochondrial fractions tended to increase in the IG and PG, compared with the CG, whereas the TG clearly showed reduced H2O2 levels. The data indicate that dietary fat induced H2O2 production by other factors, possibly related to pro-inflammatory mechanisms.
Interesterified fat induced an increase in electron leakage ratio using succinate as substrate
As we observed altered patterns in mitochondrial VO2 and H2O2 production rates after complex II activation, we next sought to evaluate the impact of those changes on mitochondrial electron leakage. After SUCC addition, we found a significant increase (P=0·0156) in the electron leakage rate in the IG when compared with the CG, confirming the highest levels in H2O2 mitochondrial production compared with the other groups (Table 5).
Electron leakage for hydrogen peroxide (H2O2) formation in liver mitochondria with succinate (SUCC) (Mean values with their standard errors, n 7–8)
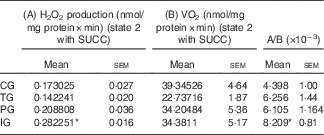
CG, control group; TG, trans-fat group; PG, palm oil group; IG, interesterified fat group.
* Significantly different in relation to control (P<0·05 ANOVA, with Dunnett’s post hoc test).
Dietary intake of trans-fat or interesterified fat during pregnancy and lactation reduced Ca2+ retention capacity in liver mitochondria
We investigated whether maternal dietary intake of trans-fat, PO or IF could affect MPTP susceptibility in isolated liver mitochondria from mouse offspring. The mitochondrial Ca2+ retention capacities are shown in Fig. 6. A representative graph showing all groups is demonstrated in Fig. 6(A) and (B), in response to sequential additions of CaCl2 (30 µmol every minute) until mitochondrial swelling and Ca2+ release.
Determination of calcium retention capacity in liver mitochondria. (A) and (B) Representation of calcium uptake through liver mitochondria from all groups after sequential additions of calcium chloride (30 µm). The range of the inset is 50 AUF. Quantitative results confirmed that trans-fat group (TG) and interesterified fat group (IG) presented less retention capacity after pyruvate/malate/glutamate (C) and succinate (D). In addition, TG and IG differ in calcium levels remaining outside the mitochondria (E) and (F). Values are expressed as means (n 7–8), with their standard errors represented by vertical bars. CG, control group; PG, palm oil group; A: ![]() , CG;
, CG; ![]() , TG;
, TG; ![]() , PG;
, PG; ![]() , IG; E and F:
, IG; E and F: ![]() , CG;
, CG; ![]() , TG;
, TG; ![]() , PG;
, PG; ![]() , IG. a,b,c Mean values with unlike letters were significantly different between experimental groups (P<0·05 ANOVA, with Newman–Keuls post hoc test).
, IG. a,b,c Mean values with unlike letters were significantly different between experimental groups (P<0·05 ANOVA, with Newman–Keuls post hoc test).

The quantitative results indicate that after activation of complex I (Fig. 6(C)) with PMG, the TG and IG displayed a 44 % (P=0·0163) and 58 % (P=0·0018) reduction in Ca2+ retention, respectively, in relation to the CG. The PG did not differ from the CG. When complex II was activated with SUCC (Fig. 6(D)), we observed a similar pattern, as the TG and IG also presented a 34 % (P=0·0215) and 55 % (P=0·0002) reduction in Ca2+ retention, respectively, when compared with the CG.
Thereafter, we analysed the differences among groups in relation to the remaining Ca2+ levels outside the mitochondria before MPTP opening – that is, the Ca2+dose-dependence for MPTP opening after PMG (Fig. 6(E)) and SUCC (Fig. 6(F)). The data indicate that different Ca2+ retention dynamics in the TG and IG, after introducing PMG, presented less cumulative Ca2+ pulses and Ca2+ releases out of mitochondria faster than the PG and CG. After SUCC, we could also infer that cumulative pulses before MPTP opening were reduced in the TG and IG, suggesting that the liver mitochondria of offspring from these groups do not tolerate elevated Ca2+ concentrations.
Discussion
In this study, we were able to demonstrate, for the first time, that the intake of normolipidic diets containing trans-fat and its substitute, IF, during pregnancy and lactation predisposes the adult offspring to liver mitochondrial dysfunction, related to alterations in the redox state. Modifications in mitochondrial bioenergetics included impairment in VO2 in the TG and increased H2O2 production in the IG. In both groups, mitochondrial membrane permeability was compromised, as evidenced by a decreased capacity for Ca2+ uptake. In addition, our results demonstrate that liver mitochondrial dysfunction was accompanied by impaired glucose homoeostasis and alterations in plasma and liver lipids profile, favouring the onset of steatosis and CVD (Table 3). These results suggest an association between early life exposure to trans-unsaturated and interesterified FA and an increased risk for developing chronic metabolic diseases in adulthood( Reference Magri, Fernandes and Souza 15 , Reference Misan, Estato and de Velasco 16 ).
Fat accumulation in the liver can be associated with local oxidative stress, impaired mitochondrial FA oxidation and inflammation, which may damage the liver( Reference Machado, Stefano and Oliveira 37 , Reference Burgueno, Cabrerizo and Gonzales Mansilla 38 ). At 110 days of life, the liver/body weight ratio was significantly higher in the TG and IG when compared with the CG and PG (Table 3). Indeed, our findings show that the IG presented higher levels of plasma glucose and the IG and TG presented increased TAG plasma concentration. These results may indicate a higher content of fat in the offspring’s livers from these groups, which may contribute to an imbalance in lipid metabolism in the liver and other tissues, although additional analysis is necessary to confirm fat accumulation in the liver.
The composition of liver FA showed that the TG and IG offspring presented a lower amount of total MUFA, total PUFA, LA and ALA (Table 4). In addition, the desaturation indexes, using the ratios C18 : 1/C18 : 0 and C18 : 3n-6/C18 : 2n-6, were significantly decreased in the TG and IG, despite the lack of differences in AA (20 : 4n-6) and DHA (22 : 6n-3) contents. This alteration in the metabolism of unsaturated FA could have consequences on membrane fluidity, phospholipid composition and the activity of channels, proteins and transporters in the liver( Reference Fu, Watkins and Hotamisligil 39 ). In addition, modifications in phospholipids composition induced by maternal diet in mitochondria-associated membranes (MAM) could interfere with the communication between mitochondria and endoplasmic reticulum (ER)( Reference Rutter and Pinton 40 ). Here we addressed the question whether mitochondrial bioenergetics plays a role in liver function and metabolic abnormalities induced by the maternal intake of different FA. Nevertheless, changes of FA in the mitochondria per se induced by the maternal intake of different FA cannot be completely excluded.
Mitochondrial phospholipids play crucial roles in mitochondrial function, as they regulate organelle fission and fusion, mitophagy and apoptosis( Reference Zhang, Tamura and Roy 41 ). Furthermore, obesity is associated with impaired mitochondrial function( Reference Putti, Sica and Migliaccio 17 ). In the present study, we observed that the TG displayed the lowest VO2 rates in liver mitochondria among all the groups of offspring (Fig. 2 and 3). In addition, ETS capacity (VO2 rate induced by FCCP) was lower in the TG compared with the CG (online Supplementary Data). This reduction in ETS capacity also caused a decrease in respiratory reserve capacity (Fig. 4), indicating that liver mitochondria from TG offspring is already working near their bioenergetics limit, possibly because of alterations in mitochondrial architecture( Reference Genova and Lenaz 42 ) and/or respiratory complex activities( Reference Chen and Bazan 7 ). The respiratory parameters in the IG were not significantly affected when compared with the CG and PG (Fig. 2 and 3), despite the same alterations in unsaturated FA composition and glucose homoeostasis observed in the TG (Tables 3 and 4). Thus, the IG demonstrated an essential change in phospholipids of the inner mitochondrial membrane, but we were not able to detect changes in oxygen consumption. These findings provide additional support for efforts to reduce industrial TFA in foods in general. In agreement with our results, it has been shown that a diet containing fish oil, rich in n-3 long-chain PUFA, compared with lard fat, rich in SFA, improved mitochondrial function, whereas lard fat decreased the mitochondrial respiratory control ratio and respiration associated with complex I and complex II( Reference Aoun, Feillet-Coudray and Fouret 5 ). In addition, high-fat diets, rich in SFA, prompted hepatic fat accumulation and insulin resistance, which were associated with impaired mitochondrial function, increased ROS production and a dysregulated expression profile of mitochondrial dynamic proteins( Reference Lionetti, Mollica and Donizzetti 22 ).
Mitochondria can be an important source of ROS/RNS production and mitochondrial bioenergetics is directly involved in this characteristic( Reference Nathan and Cunningham-Bussel 43 ). Even though the TG presented significant alterations in mitochondrial respiratory parameters, the H2O2 production rate was the lowest among all the groups. Nevertheless, the IG demonstrated an increase in H2O2 production after complex II stimulation (Fig. 5). As H2O2 production was correlated to electron leakage (Table 5), several aspects related to redox signalling are worth discussing. The decrease in H2O2 production observed in the TG might reflect a decrease in mitochondrial O2 flux (Fig. 2 and 3), which could be the result of decreased activities of ETS proteins, which, in turn, decreased the escape of electrons and ROS production. On the other hand, the significant increase in H2O2 production and electron leakage in the IG strongly suggest that specific alterations in spatial arrangements of ETS proteins are involved in mitochondrial dysfunction, possibly associated with the opening of the MPTP( Reference Zorov, Juhaszova and Sollott 44 ).
The results presented in Fig. 6 show that liver mitochondria of the TG and IG adult offspring are more susceptible to MPTP opening, as the mitochondrial capacity for handling Ca2+ loads is significantly decreased in both groups when compared with the CG and PG. Arruda et al.( Reference Arruda, Pers and Parlakgul 45 ) has demonstrated that in cases of obesity the liver is susceptible to a marked reorganisation by physical interaction between ER and mitochondria, through MAM( Reference Lynes, Raturi and Shenkman 46 ), which was shown to be dependent on the palmitoylation state in cysteine residues( Reference Arruda, Pers and Parlakgul 45 ). Our data indicate that the TG and IG presented reduced levels of palmitic acid (Table 4), which can result in a different state of palmitoylation and regulation of Ca2+ homoeostasis. Further studies are necessary to confirm this interaction between diet and MAM.
Alterations in mitochondrial membrane permeability and function are strongly related to the composition of the phospholipid cardiolipin (CL). The main fatty acyl moiety in CL is LA, with 60–80 % of CL being tetralinoleoyl CL and it has been proposed that high amounts of tetralinoleoyl CL are essential for optimal mitochondrial function( Reference Khairallah, Kim and O’Shea 47 ). Indeed, the adult offspring from dams fed TFA or and interesterified FA presented decreased contents of LA. In addition, the desaturation index is lower in the TG and IG. Therefore, it is plausible to assume that alterations in mitochondrial cardiolipin metabolism induced by the maternal diet might be involved in liver mitochondrial dysfunction. An increase in LA content in liver mitochondria induced by MitoQ has been associated with improved mitochondrial function and the prevention of some factors of metabolic syndrome in mice fed an obesogenic diet( Reference Fouret, Tolika and Lecomte 48 ).
Mitochondrial dysfunction as a consequence of alterations in FA metabolism in the liver is possibly mediated by epigenetic events. Accordingly, the decrease in Δ6 desaturase index observed in all three experimental groups compared with control animals (Table 4) could be related to a decreased expression of Fads2 gene mRNA expression, encoding for Δ6 desaturase, the rate-limiting enzyme in PUFA synthesis, as it has been shown that a high-fat diet during pregnancy and lactation increased the methylation of this gene in the adult offspring, regulating PUFA metabolism in the liver( Reference Hoile, Irvine and Kelsall 49 ).
The implications of the types of FA consumed during pregnancy and lactation can lead to the development of non-communicable diseases, as well as affecting fetus maturation and postnatal development. Altogether, our results imply that the consumption of trans-fat and its substitutes during critical stages of life could lead to impairments in the metabolism and development of offspring and to a ‘programming’ of long-term modifications in mitochondrial bioenergetics that might negatively impact metabolic outcomes in adult life. These observations suggest that we should be concerned with the use of PO, partially hydrogenated vegetable fat and IF by the food industry. We recommend monitoring the dietary consumption of trans-fat and SFA of pregnant and lactating women.
Acknowledgements
The authors thank the Brazilian Company Triângulo Alimentos Ltda, Itápolis, SP, Brazil (http://www.trianguloalimentos.com.br) for supplying the fats.
This work was supported by grants from Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro and ConselhoNacional de DesenvolvimentoCientífico e Tecnológico.
P. C. d. V., G. C., D. M. R.-F., R. M. A. R. d. S., T. E.-B., A. G. and M. d. G. T.d.-C.: contributed to the assembly, analysis and interpretation and manuscript drafting; P. C. d. V., G. C., D. M. R.-F., R. M. A. R. d. S., T. E.-B., A. G., C. M. and M. d. G. T.d.-C.: contributed to the data collection; P. C. d. V., G. C., D. M. R.-F., R. M. A. R. d. S., T. E.-B., A. G. and M. d. G. T.d.-C., C. M. and F. L. C. S. approval of the final version of the manuscript; P. C. d. V., G. C., D. M. R.-F., R. M. A. R. d. S., T. E.-B., A. G. and M. d. G. T.d.-C. and F. L. C. S.: contributed to the study conception and design.
None of the authors has any conflicts of interest to declare.
Supplementary material
For supplementary material/s referred to in this article, please visit https://doi.org/10.1017/S0007114517001817













