


Non-technical Summary
The geologic history of abalones, commercially important mollusks living on rocky shores and threatened by overfishing and climate change, is uclear due to a globally poor fossil record and an uncertain taxonomy. Fossils 3.5 million years old found at an exceptionally rich site in Italy are presented, and the European fossil record of abalones is revisited using a quantitative approach to morphological characters. A new Pliocene species is described, and the history of Mediterranean and eastern Atlantic abalones and their evolutionary relationships during the past 23 million years is tentatively reconstructed.
Introduction
The family Haliotidae Rafinesque, Reference Rafinesque1815 (Mollusca, Vetigastropoda), or abalone, includes 56 extant species and 18 subspecies within the genus Haliotis Linnaeus, Reference Linnaeus1758, distributed in fully marine environments at tropical and temperate latitudes, and many fossil species, often of uncertain status (Geiger, Reference Geiger1998, Reference Geiger2000; Geiger and Groves, Reference Geiger and Groves1999; Geiger and Owen, Reference Geiger and Owen2012). Abalones are grazing sea snails that commonly inhabit hard substrata at depths of 0–30 m (Geiger and Owen, Reference Geiger and Owen2012). Adults feed mostly on drift macroalgae, outcompeting sea urchins within the same guild of macroherbivores when food is not limiting (Jenkins, Reference Jenkins2004). They produce an aragonitic shell, with a size up to 120 mm in warm tropical waters (Geiger and Owen, Reference Geiger and Owen2012), more than doubling in temperate waters, where H. rufescens Swainson, Reference Swainson1822 reaches 313 mm in offshore Oregon (Estes et al., Reference Estes, Lindberg and Wray2005), confirming a latitude–size relationship encountered in other large gastropods (Dominici et al., Reference Dominici, Fornasiero and Giusberti2020). The large size of abalones has turned out to be a threat to the survival of natural populations because this shellfish is an important seafood item that has sustained historical commercial fisheries around the world. Overfishing, disease, ocean warming, and acidification recently led to stock collapse, and a few species are now threatened (Rogers-Bennet et al., Reference Rogers-Bennet, Haaker, Huff and Dayton2002; Neuman et al., Reference Neuman, Tissot and Vanblaricom2010; Kiyomoto et al., Reference Kiyomoto, Tagawa, Nakamura, Horii, Watanabe, Tozawa, Yatsuya, Yoshimura and Tamaki2013; Li et al., Reference Li, Mao, Jiang, Zhang, Fang and Bian2018; Rogers-Bennet and Catton, Reference Rogers-Bennett and Catton2019; Wells et al., Reference Wells, Keesing, Gagnon, Bessey, Spilsbury and Irvine2023), including the European green ormer, H. tuberculata (Peters, Reference Peters2021).
The roots of Haliotis modern diversity are not simple to trace. Its phylogenetic position within the basal clade Vetigastropoda is unclear (Bouchet et al., Reference Bouchet, Rocroi, Hausdorf, Kaim, Kano, Nützel, Parkhaev, Schrödl and Strong2017), notwithstanding recent efforts based on a comprehensive phylogenomic framework (Cunha and Giribet, Reference Cunha and Giribet2019). According to the latter, abalones are not closely related to any of the other Vetigastropoda superfamilies, justifying the elevation of the superfamily Haliotoidea to the status of order Haliotida Rafinesque, Reference Rafinesque1815 (Cunha et al., Reference Cunha, Reimer and Giribet2021). If abalones are the sister taxon to the Seguenziida, as still maintained by some (Uribe et al., Reference Uribe, Makiri and Harasewych2022), then their origin could date back to the Paleozoic. The oldest reported Haliotis is found in the Campanian and Maastrichian of California and the Caribbean (Upper Cretaceous; Sohl, Reference Sohl1992; Groves and Alderson, Reference Groves and Alderson2008). There are 42 recognized species of Haliotis, 38 of which are of Neogene age (<23 Ma; Geiger and Groves, Reference Geiger and Groves1999; Estes et al., Reference Estes, Lindberg and Wray2005). Molecular phylogenetic analyses and chromosome evidence suggest that the modern abalone biodiversity hotspot, centered in the tropical Indo-Pacific (Geiger, Reference Geiger2000), originated from a Tethyan ancestor (Geiger and Groves, Reference Geiger and Groves1999; Estes et al., Reference Estes, Lindberg and Wray2005; Bester-van der Merwe et al., Reference Merwe, D'Amato, Swart and and Roodt-Wilding2012), a hypothesis consistent with known global dynamics of molluscan diversity (Yasuhara et al., Reference Yasuhara, Huang, Reuter, Tian and Cybulski2022).
The reasons for a poor and confused fossil record are varied. The habitat of abalones is not particularly conducive to preservation of the shell after death (negative taphonomic bias; Geiger and Groves, Reference Geiger and Groves1999). Shallow-water rocky substrates are of limited extension and unfavorable to fossilization because of intense hydrodynamic processes that hinder sedimentation and burial and facilitate shell breakage, in addition to mollusk predators (Zuschin et al., Reference Zuschin, Stachowitsch and Stanton2003; Albano et al., Reference Albano, Hua, Kaufman and Zuschin2022), which include for abalones a variety of bony fishes, crabs, and cephalopods (Geiger and Owen, Reference Geiger and Owen2012; Aspe et al., Reference Aspe, Cabales, Sajorne and Creencia2019; see also Ponder and Lindberg, Reference Ponder and Lindberg2008). Lower sedimentation rates lead to long exposure to sea water, promoting colonization of the shell by microboring organisms and aragonite dissolution, other important factors of shell loss (Cherns et al., Reference Cherns, Wheeley, Wright, Allison and Bottjer2011).
The heterogeneity of microhabitats exploited by abalones adds to morphological plasticity of the shell, hinders species recognition, and facilitates taxonomic oversplitting, examples including past and present Mediterranean forms (see for example H. mykonosensis Owen, Hanavan, and Hall, Reference Owen, Hanavan and Hall2001; Chiappa et al. Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). With the preceding limitation in mind, the present paper presents new findings and a revision of the European record of Haliotis to shed further light on its geological history on the basis of a large dataset of fossil and Recent shells.
The Upper Cretaceous record from Europe, based on H. antiqua Binkhorst, Reference Binkhorst1861 and H. cretacea Lundgren, Reference Lundgren1894, has been dismissed due to misidentification (Kaunhowen, Reference Kaunhowen1897; Davies and Eames, Reference Davies and Eames1971). Although very sparse compared with that of other gastropod families, the European record ranges in age from the early Miocene to the Pleistocene (Eichwald, Reference Eichwald1829; Hörnes, Reference Hörnes1856; Cossmann, Reference Cossmann1896; Delhaes, Reference Delhaes1909 [fide Geiger and Groves, Reference Geiger and Groves1999]; Cossmann and Peyrot, Reference Cossmann and Peyrot1917; Glibert, Reference Glibert1949; Baluk, Reference Baluk1975; Krach, Reference Krach1981; Strausz, Reference Strausz1966; Lozouet et al., Reference Lozouet, Lesport and Renard2001b; Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003; Saint Martin et al., Reference Martin, Merle, Cornée, Filipescu, Saint Martin and and Bucur2007; Górka et al., Reference Górka, Studencka, Jasionovski, Hara, Wysocka and Poberezhskyy2012; Forli et al., Reference Forli, Stalennuy and Dell'Angelo2015; Owen and Berschauer, Reference Owen and Berschauer2017). New species recently described for the Spanish Pliocene are H. quinquecentenaris Lozano-Francisco and Vera-Peláez, Reference Lozano-Francisco and Vera-Peláez2002, H. iberica Landau et al., Reference Landau, Marquet and Grigis2003, and H. telescopica Vera-Peláez in Vera-Paláez and Lozano-Francisco, Reference Vera-Peláez and Lozano-Francisco2022. Outside Europe, the fossil record of Haliotidae is much poorer (Geiger and Groves, Reference Geiger and Groves1999).
Abalones have a low diversity in the Neogene of Italy. For a long time, Pliocene reports amounted only to H. lamellosoides Sacco, Reference Sacco1897 (often under the name H. tuberculata tuberculata Linnaeus, Reference Linnaeus1758), with the relatively recent addition of H. bertinii Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003. This paper reports on new findings from southern Tuscany, which allow a significant increase in the understanding of taxonomy and biogeography of Cenozoic to Recent Haliotidae. This report includes a large number of specimens, ranging from juveniles to fully adult shells, found in two outcrops in an area informally known as “Terre Rosse” (meaning “red lands”).
Available reviews of the European fossil record attributed most Neogene reports to the living H. tuberculata (Geiger and Groves, Reference Geiger and Groves1999; Estes et al., Reference Estes, Lindberg and Wray2005), other recently described species mentioned only in passing (Geiger and Owen, Reference Geiger and Owen2012). All Oligocene to Pliocene abalones have been grouped under the subspecies H. tuberculata volhynica, justified by the extreme plasticity in shell morphology of Atlantic and Mediterranean Recent populations (Geiger and Groves, Reference Geiger and Groves1999, p. 872, following Strausz, Reference Strausz1966).
The reappraisal of H. volhynica Eichwald, Reference Eichwald1829 (Forli et al., Reference Forli, Stalennuy and Dell'Angelo2015; Owen and Berschauer, Reference Owen and Berschauer2017) and the new data from Terre Rosse presented here allow us to clearly separate H. volhynica from the living H. tuberculata. For the first time, a multivariate statistical approach is applied to a large quantitative dataset of fossil and extant abalones, and the results are interpreted in the light of traditional taxonomic practice based on descriptive data. Geological relationships between past and present northwestern African and European abalones are re-evaluated.
Geologic and stratigraphic setting
The geographic name “Terre Rosse” informally refers to a small area near Castelnuovo Berardenga, in the province of Siena (Tuscany, Central Italy). It comprises the two localities Stroncoli, where Pliocene yellow sands with lenses of pebbly sand crop out (Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003), and Campino, characterized by yellow sands (Laghi, Reference Laghi1984; Forli et al., Reference Forli, Dell'Angelo, Montagna and Taviani2004, Reference Forli, Cresti and Corti2021; Cresti and Forli, Reference Cresti and Forli2021) (Fig. 1). The Pliocene of the Siena Basin, up to 600 m thick, is formed mainly by marine strata overlain by regressive sediments deposited during the uplift of southern Tuscany, before Pleistocene subaerial exposure (see “Siena sub-basin” in Martini and Aldinucci, Reference Martini and Aldinucci2017). The Terre Rosse succession belongs to nearshore sands and conglomerates of the San Vivaldo Sands Formation, passing southward (basinward) to offshore muds (informally known as “Blue Clays”; Martini and Aldinucci, Reference Martini and Aldinucci2017). The Stroncoli and Campino shallow-water pebbly and shelly sandstone unit from which abalones were collected is here tentatively referred to the informal chronostratigraphic units S3 and S4 recognized in the nearby Guistrigona area (Martini and Aldinucci, Reference Martini and Aldinucci2017; late Zanclean–early Piacenzian = ca. 4.0–3.0 Ma).
Stratigraphic log of the Pliocene of the Terre Rosse area (Siena basin; late Zanclean–early Piacenzian). TAD = terraced alluvial deposits (Quaternary); A = sands; B = pebbly sands; C = clays; D = silty sands; E = gravels. Top right, locality Campino; bottom right, locality Stroncoli (scale 1:55.000).
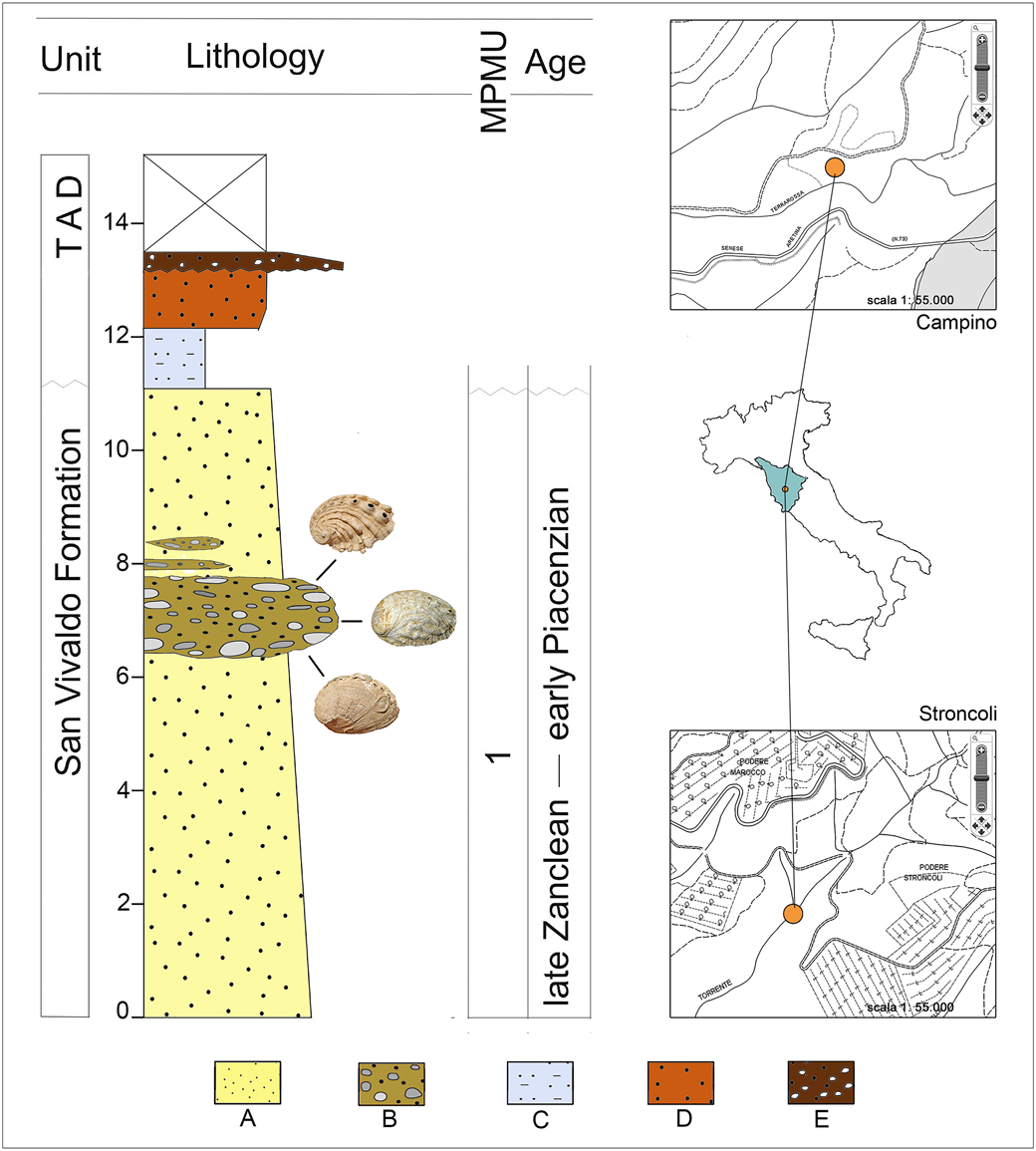
The Terre Rosse abalones were compared with available data from other Italian and European collections (see abbreviations that follow). Stratigraphy and provenance of specimens of the historical collection of Luigi Bellardi and Federico Sacco, collected in northwestern Italy and hosted in the Museum of Natural History of Turin, were taken from Ferrero Mortara et al. (Reference Ferrero Mortara, Montefameglio, Pavia and Tamperi1982). These fossils range in age from the middle Burdigalian (early Miocene, ca. 19.0–17.0; Zunino and Pavia, Reference Zunino and Pavia2009) to the early Pleistocene (= ca. 2.5–1.0 Ma; Sacco, Reference Sacco1897). Most studied specimens of H. volhynica were collected at Maksymivka (Ternopil, Ukraine; see Radwański et al., Reference Radwański, Górka and Wysocka2006), in middle Miocene fully marine deposits of eastern Europe that can be correlated with the Serravallian (= 14.0–12.0 Ma; Śliwiński et al., Reference Śliwiński, Bąbel, Nejbert, Olszewska-Nejbert, Gąsiewicz, Schreiber, Benowitz and Layer2012).
Materials and methods
Terre Rosse fossiliferous strata were bulk sampled, and additional shells were surface collected in the field. Specimens were isolated from the residue after wet-sieving sediment through a 1 mm-size screen. Shells (not casts or molds) in public institutions and in private collections, or their photographs in the literature, were measured. The initial dataset included 379 specimens (153 fossil, 226 Recent specimens; Supplementary Material, dataset 1).
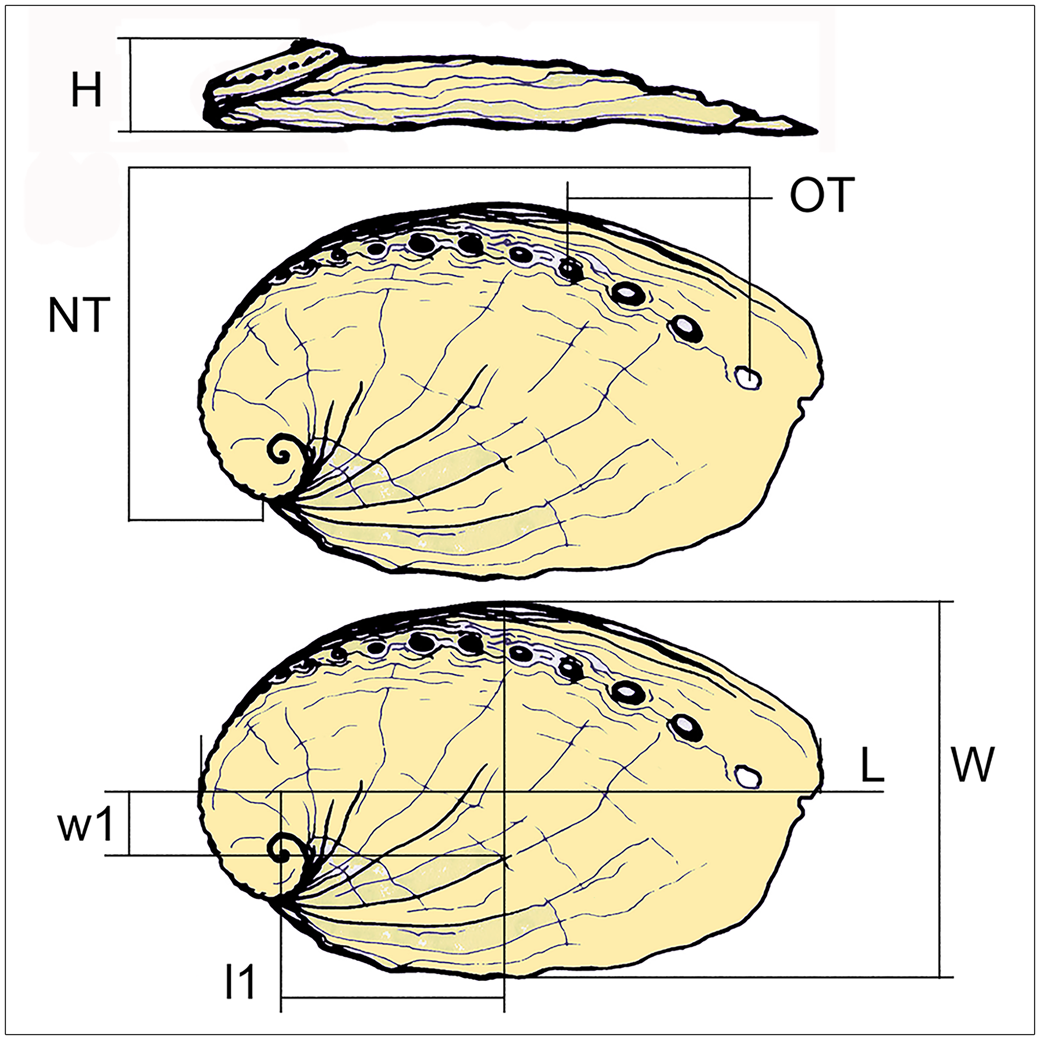
Morphological characters useful for the species- and subspecies-level taxonomy of abalones are general shape; position of apex; spire height; position, number, and shape of tremata; axial and spiral ornamentation; shape and position of the columnellar fold; and size (Geiger and Groves, Reference Geiger and Groves1999). We measured shell length (L), shell length from apex to L midline (l1), shell width (W), shell width from apex to W midline (w1), shell height (H), and number of tremata, separating total number (NT) from open tremata (OT; Fig. 2). Incomplete specimens were discarded, and statistical analysis was performed on 82 fossils and 124 Recent specimens belonging to 15 taxa recognized on traditional descriptive grounds. Secondary data to explore changes in shell shape among these a priori groups included height:length ratio (H/L), width:length ratio (W/L), height:width ratio (H/W), rate of length increase (l1/L), and rate of width increase (w1/W).
Haliotis shell morphometrics: L = maximum length; l1 = length from apex to L midline; W = maximum width; w1 = width from apex to W midline; H = height from apex to aperture; NT = total number of tremata, from the contact of the whorl with the posterior labial margin to the anterior end of the row; OT = open tremata.
Extant taxa included in the large dataset (Supplementary Material, dataset 1) are H. tuberculata tuberculata Linnaeus, Reference Linnaeus1758, H. t. coccinea Reeve, Reference Reeve1846, H. t. fernandesi Owen and Afonso, Reference Owen, Afonso, Geiger and Owen2012, H. stomatiaeformis Reeve, Reference Reeve1846, H. tuberculata “Dakar”, H. geigeri Owen, Reference Owen2014, H. pustulata Reeve, Reference Reeve1846, and H. marmorata Linnaeus, Reference Linnaeus1758. All specimens for which not all of the morphometrics could be measured (mainly H) were removed, resulting in a first set of specimens upon which to perform a statistical analysis (Supplementary Material, dataset 2).
Fossil taxa are H. benoisti Cossmann, Reference Cossmann1896, H. volhynica Eichwald, Reference Eichwald1829, H. lamellosoides Sacco, Reference Sacco1897, H. bertinii Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003, H. plioetrusca n. sp., H. stalennuyi Owen and Berschauer, Reference Owen and Berschauer2017, H. ovata Michelotti, Reference Michelotti1847, H. monilifera Michelotti, Reference Michelotti1847, and H. torrei Ruggieri, Reference Ruggieri1990. The last four species, being scantily represented, were excluded from the statistical analysis applied to a third subset (Supplementary Material, dataset 3).
All multivariate analyses were performed using RStudio (version: 2023.12.1+402), with the stats (R Core Team, 2022) and vegan (Oksanen et al., Reference Oksanen, Simpson, Blanchet, Kindt and Legendre2022) packages.
Notwithstanding the advantage of exploring potentially covariate morphometrics, multivariate analysis of morphological traits has rarely been applied to the study of abalone shell, only at the population level (McShane et al., Reference McShane, Schiel, Mercer and Murray1994; Bachry et al., Reference Bachry, Solihin, Gustiano, Soewardi and Butet2019) and never on fossils. A recent multivariate approach applied to a conchological study of cowries (Southgate and Militz, Reference Southgate and Militz2023)—also a group of large gastropods with considerable infraspecific variation in shell form—offered a template for the ordination and statistical study of abalones. On the basis of that experience, non-metric multidimensional scaling (nMDS) was preferred to other techniques (e.g., principal component analysis) for resolving group differences when studying general shell form. While cowries have a smooth outer surface, the abalone shell offers characters such as ribs, tubercles, and lamellae as an additional means to diagnose taxa. These last characters were left unquantified but used nonetheless as diagnostic characters to define a priori groups, following traditional taxonomic practice (e.g., Geiger and Owen, Reference Geiger and Owen2012; Owen and Berschauer, Reference Owen and Berschauer2017).
For multivariate analysis, values resulting from both dimensionless and differently scaled measurements were transformed to Z-scores before testing (R function: scale) so that each morphometric was centered, with a mean of zero, and uniformly scaled, with values expressed in terms of deviation from the mean (i.e., |Z-score| > 3). We identified atypical specimens if at least one morphometric exceeded three standard deviations of the mean. A resemblance matrix was computed on the basis of Euclidean distances between the remaining specimens (R function: vegdist) and visualized in two dimensions through nMDS (R function: metaMDS). We measured the coefficient of determination (R2, or squared correlation) on the basis of morphometric variables and ordination scores (R function: envfit). The results of each morphometric were then overlayed on the existing nMDS ordination and visualized through morphometric clines (R function: ordisurf) to isolate the influence of each morphometric on the plot configuration.
To avoid the subjectivity of visual interpretations of multidimensional data after reducing dimensionality and thereby validate differences in shell form among a priori groups, a one-factor permutational analysis of variance (PERMANOVA) was used to assess whether within-group distances are smaller than among-group distances (R function: adonis2). We used the F-value to measure the ratio of within- and among-species variance in morphospace. Permutation-based tests for homogeneity of multivariate dispersions were used to assess whether dispersions (the distance of specimens from their group centroid) vary among groups (R function: permutest.betadisper; for other details on the multivariate analyses, see Southgate and Militz, Reference Southgate and Militz2023; the script adapted to the present dataset is available in the Supplementary Material). Boxplots were used to visualize differences in mean and median tendencies, range, and quantiles of morphometrics among groups.
Biogeographic provinces discussed in the study, following Geiger (Reference Geiger2000, p. 58), are Northeast Atlantic (western Atlantic from Normandy, France, to western Morocco, 30°N); Mediterranean (Mediterranean Sea); West African (central and southwestern Atlantic, from 30°N, including Canary Islands, to Cape Town, Republic of South Africa); Red Sea (North of Djibouti).
Most French fossil sites can be located following Lozouet et al. (Reference Lozouet, Maestrati, Dolin and Favia2001a), to the exclusion of Nouvelle-Aquitaine, Vienne, Moulin Pochard, with coordinates 48.053°N, 0.670°E. Italian sites include Petralia Sottana, Palermo (37.804°N, 14.082°E); Stroncoli, Siena (43.19594°N, 11.35428°E); Campino, Siena (43.20176° N, 11.34463°E); and Quercecchio, Siena (43.030°N, 11.413°. Ukranian sites include Ternopil, Maksymivka quarry (49.361°N, 25.543°E).
Suprageneric systematics follows the World Register of Marine Species (WoRMS, 2023). Abbreviations: H = height from apex to horizontal plane; L = maximum length; l1 = length from apex to L midline; W = maximum width; w1 = width from apex to W midline; NT = total number of tremata from the contact of the whorl with the posterior labial margin to the end of row in anterior part; OT = number of open tremata. Size is expressed in millimeters throughout the paper.
Repositories and institutional abbreviations
Figured specimens and other material examined in this study are deposited in the following institutions: British Museum of Natural History (BMNH), London, United Kingdom; Linnean Society of London (LSL), London, UK; Museo di Storia Naturale, Università di Firenze (MSNF IGF), Firenze, Italy; Museo di Storia Naturale di Milano, (MSNM), Milano, Italy; Museo Regionale di Scienze Naturali (MRSN), Torino, Italy; Museo di Zoologia, Università di Bologna (MZB), Bologna, Italy; National Museum of New Zealand (NMNZ), Wellington, New Zealand; Natuurhistorisch Museum Rotterdam (NHMR), Rotterdam, Netherland; Muséum d’Histoire naturelle de Bordeaux (MHNBx), Bordeaux, France; Muséum national d'Histoire naturelle (MNHN), Paris, France; Wiener Naturhistorische Museum (NHMW), Wien, Austria.
Private collections temporarily hosting specimens are: CLZ, Alain Cluzaud (France); JLC, Jean-François Lesport (France); RMQ, Robert Marquet (Belgium); BDA, Bruno dell'Angelo (Italy); CBC, Cesare Bogi (Italy); FCC, Fabio Ciappelli (Italy); MCC, Massimo Cresti (Italy); MFC, Maurizio Forli (Italy); MRC, Massimo Rocca (Italy); MBB, Mauro M. Brunetti (Spain).
Others collections are: BOC, Buzz Owen, California, USA; DDC, Dwayne Dinucci, California, USA; FFC, Franck Frydman, Paris, France; FRC, Ramiro Fladeiro, Valhascos, Portugal; PRC, Peter Ryall, Austria; RKC, Robert Kershaw, NSW, Australia; (Owen et al., Reference Owen, Ryall and Pan2015).
Results
Haliotis at Terre Rosse.—Haliotis lamellosoides is everywhere abundant in the area (16 specimens at Campino, 79 at Stroncoli), whereas H. bertinii (14 specimens) and Haliotis plioetrusca n. sp. (13 specimens) were found only at Stroncoli, a site with a total of 106 abalone specimens. The abundant biometric data available for H. lamellosoides, including both juvenile and adult individuals, testify to a wide range of variability of this species and suggest including in its synonymy recently described species at Estepona, Spain (Lozano-Francisco and Vera-Peláez, Reference Lozano-Francisco and Vera-Peláez2002; Landau et al., Reference Landau, Marquet and Grigis2003; Vera-Peláez in Vera-Paláez and Lozano-Francisco, Reference Vera-Peláez and Lozano-Francisco2022).
The Terre Rosse fossils greatly contributed to the significance of the multivariate analysis of fossil abalone, with 50 out of 76 fossils of the quantitative subset (Supplementary Material, dataset 3).
Multivariate analysis was performed on successive subsets of the whole, until two of them were selected to build meaningful and more easily interpretable matrices. The first set includes Recent specimens of Mediterranean and Atlantic species and subspecies of Europe and northwestern Africa. All specimens of the larger dataset that did not allow us to measure morphometric H were excluded. The a priori groups in this set are H. stomatiaeformis Reeve, Reference Reeve1846 (N = 27), H. tuberculata tuberculata Linnaeus, Reference Linnaeus1758 (N = 32), H. t. coccinea Reeve, Reference Reeve1846 (N = 29), H. t. fernandesi Owen and Afonso, Reference Owen, Afonso, Geiger and Owen2012 (N = 4), and a population of H. tuberculata from Dakar (Senegal), formerly attributed to H. speciosa Reeve, Reference Reeve1846 (N = 20) (Supplementary Material, dataset 2). The first three taxa were recently validated by molecular data (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022) to offer reliable a priori groups to test the multivariate approach.
The second matrix included species-level data collected from Neogene fossils of Europe and previously recognized by taxonomists. This set includes H. benoisti Cossmann, Reference Cossmann1896 (N = 11), H. volhynica Eichwald, Reference Eichwald1829 (N = 11), H. lamellosoides (Sacco, Reference Sacco1897) (N = 37), H. bertinii Forli et al. Reference Forli, Dell'Angelo, Ciappelli and Taviani2003 (N = 8), and the Stroncoli paleo-population of H. plioetrusca (N = 11). Two further taxa were considered during a preliminary trial, namely H. ovata Michelotti, Reference Michelotti1847 and H. torrei Ruggieri, Reference Ruggieri1990, but discarded because of insufficient data. All specimens of H. tuberculata in the strict sense (excluding subspecies from Macaronesia and Senegal) of the first dataset were included in the second matrix to compare fossil abalones with the two extant European abalones (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022).
The a priori assigned groups of the first dataset (i.e., H. stomatiaeformis, H. tuberculata tuberculata, H. t. coccinea, H. t. fernandesi, and the Dakar population of H. tuberculata) were capable of explaining a significant amount (R2 = 0.42, F = 18.762, p < 0.0001) of the variation in shell form (Fig. 3.1). Differences of shell form among species were highly significant judging from F, or the ratio of within- and among-species variance in morphospace (Table 1) and particularly when confronted with molecular data (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022), allowing us to distinguish H. stomatiaeformis from H. tuberculata, particularly from the other Mediterranean abalone H. t. tuberculata (F = 52.23, p = 0.0001: if p is insignificant, then the significant F from PERMANOVA, like in this case, indicates that the differences are driven by differences in centroids). Subspecies H. t. tuberculata and H. t. coccinea show some overlap (F = 16.21, p = 0.0001), confirming a limited gene flow suggested by molecular studies (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). The overlap increases between H. t. fernandesi and H. t. coccinea (F = 4.82, p = 0.0013). Shell form for the Dakar population of H. tuberculata, formerly attributed to H. speciosa, was distinct from H. stomatiaeformis (F = 33.80, p = 0.0001), overlapping with H. t. coccinea (F = 7.37, p = 0.0001) and H. t. fernandesi (F = 4.13, p = 0.0035) and strictly comparable to that of H. t. tuberculata (F = 2.71, p = 0.0283; Fig. 3.1; Table 1).
(1) nMDS ordination (stress = 0.16) of the resemblance matrix for extant abalone of Europe and West Africa, where shaded ellipses indicate the 95% confidence interval of group (species, subspecies, or population) centroids, and plot characters indicate data source. (2–8) Associations between ordination structure and morphometrics influencing this structure, where the thin black lines illustrate: (2) length; (3) height:length ratio; (4) width:length ratio; (5) height:width ratio; (6) relative spire position; (7) total tremata; (8) open tremata.
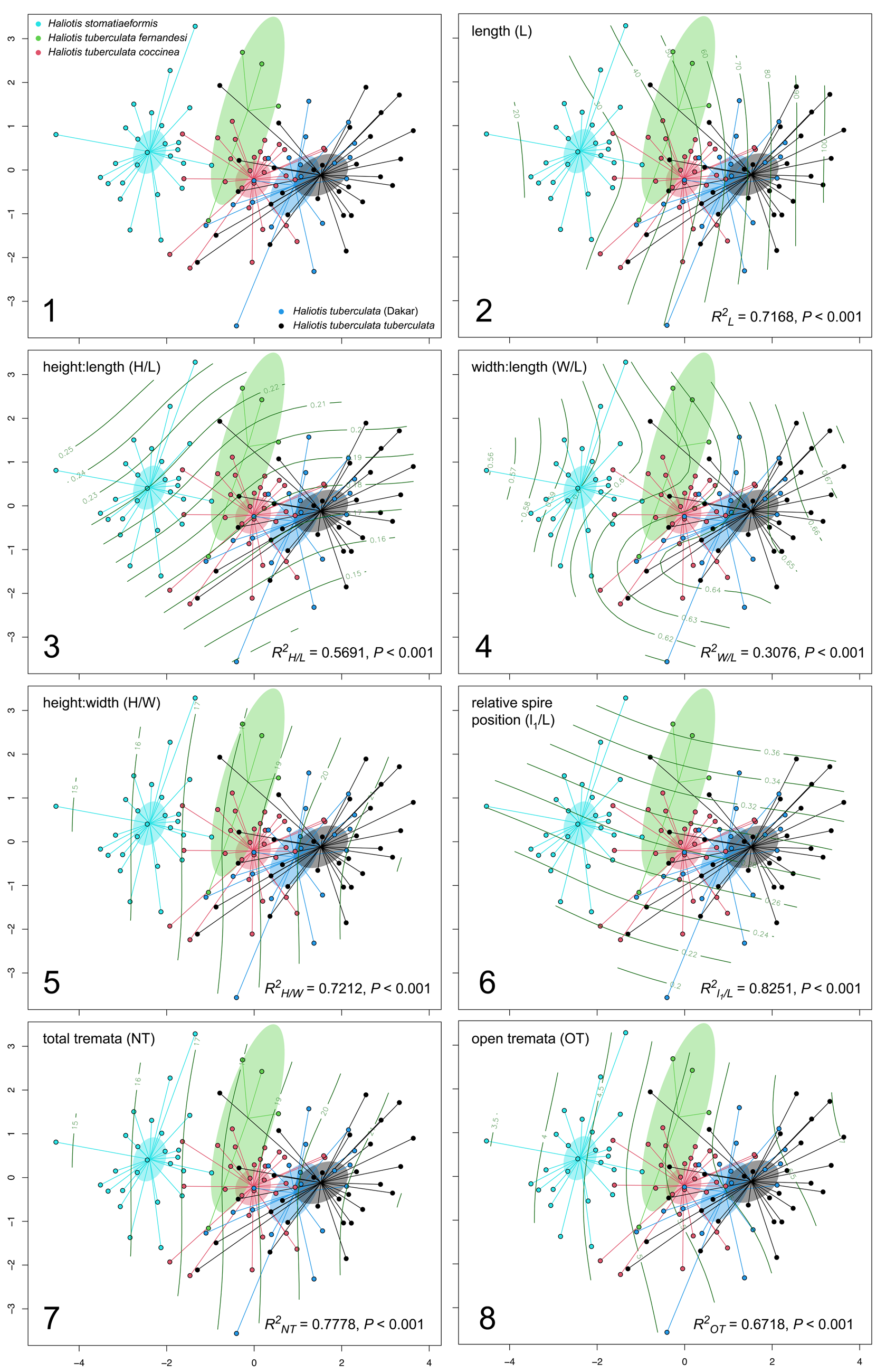
Results of pairwise comparisons testing the hypotheses that there were no differences in central tendency (i.e., centroid) of shell form among the studied extant abalone groups (species, subspecies, or population of extant northeastern Atlantic and Mediterranean abalone). Statistic F is the ratio of the amount of variation between versus within groups, with the numerator and denominator each weighted by their degrees of freedom. It is 0 or positive, with larger values corresponding to larger proportional importance of the grouping factor. Holm-adjusted probability that the distance between centroids arose by random chance (p) is presented.

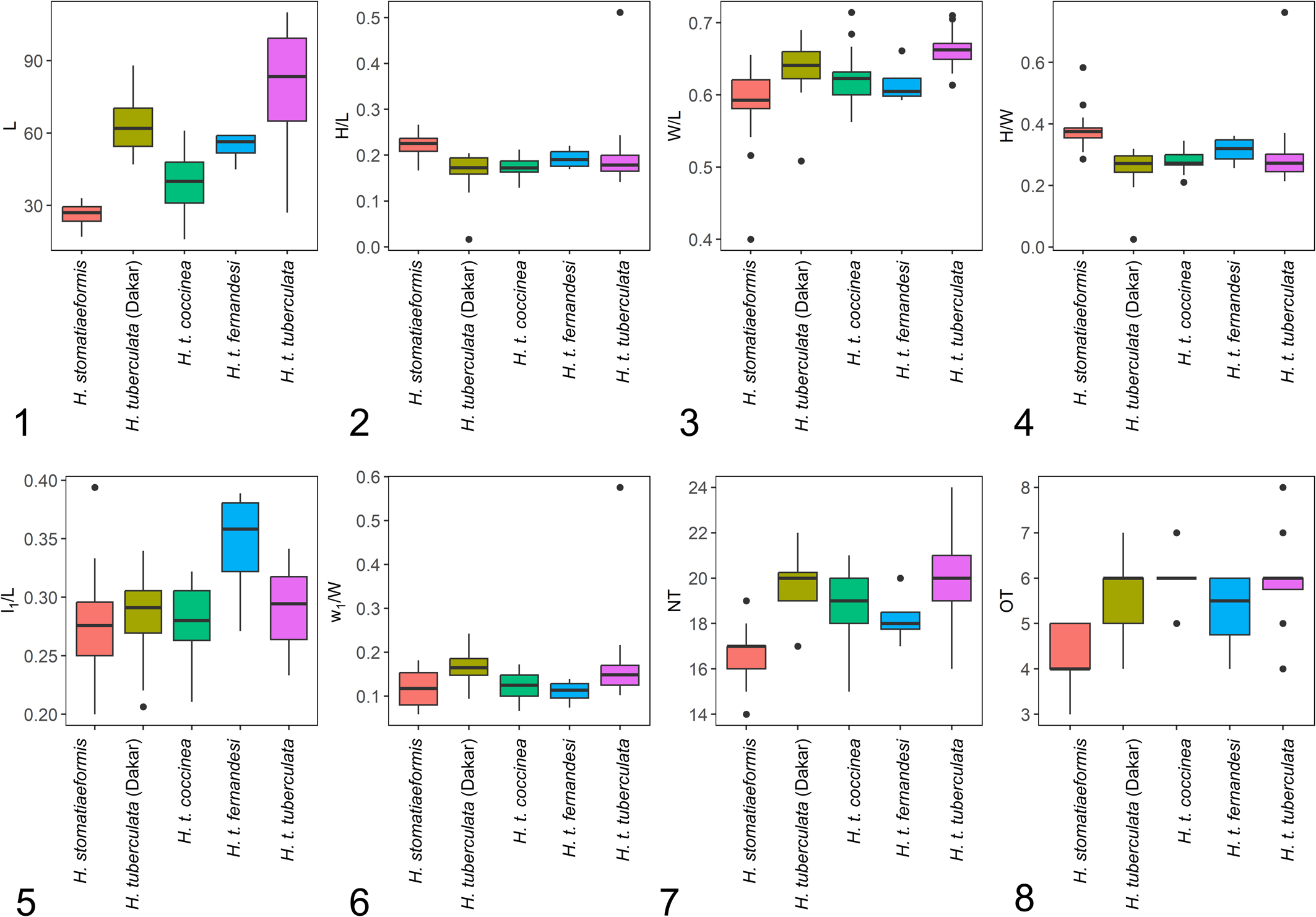
All morphometrics considered representative of shell form (i.e., L, H/L, W/L, H/W, l1/L, w1/W, NT, OT) significantly influenced the ordination structure of the abalone groups visualized in Fig. 3. The most important morphometric was l1/L (R2 = 0.82, p < 0.001), followed by NT (R2 = 0.78, p < 0.001), H/W (R2 = 0.72, p < 0.001), L (R2 = 0.72, p < 0.001), OT (R2 = 0.67, p < 0.001), H/L (R2 = 0.57, p < 0.001), W/L (R2 = 0.31, p < 0.001), and last w1/W (R2 = 0.31, p < 0.001; not shown in Fig. 3). Most univariate methods allow us to separate H. stomatiaeformis from H. tuberculata, with the exception of w1/W (Figs 3.2–3.8, 4).
Box plots showing univariate comparisons among extant European and West African abalone species and subspecies. (1) Shell length. (2) Height:length ratio. (3) Width:length ratio. (4) Height:width ratio. (5) Relative spire position with respect to L. (6) Relative spire position with respect to W. (7) Total number of tremata. (8) Open tremata. Boxes illustrate first and third quartile as box edges and median as central line.
Since the results of the nonmetric MDS of the first dataset are consistent with an approach to the taxonomy of the family that includes genetic data (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022), the same multivariate statistics can be used to separate species known only from fossils, whose taxonomic status relies only on shell morphology. The a priori groups in the second dataset, including both extinct and extant taxa (i.e., H. benoisti, H. lamellosoides, H. volhynica, H. plioetrusca, H. tuberculata, and H. stomatiaeformis), were able to explain a highly significant amount (R2 = 0.56, F = 25.832, p < 0.001) of the variation in shell form (Fig. 5.1). The multivariate approach was particularly significant (Table 2), with H. plioetrusca dissimilar to H. stomatiaeformis (F = 57.03, p = 0.0001), H. benoisti (F = 27.42, p = 0.0001), and H. volhynica (F = 26.20, p = 0.0001), and H. benoisti and H. volhynica the most similar (F = 3.63, p = 0.0023; Fig. 5). Similar pairs include H. tuberculata and H. plioetrusca (F = 7.92, p = 0.0001) and H. lamellosoides and H. plioetrusca (F = 11.38, p = 0.0001). H. stomatiaeformis is distinct from H. benoisti (F = 18.56, p = 0.0001) and H. volhynica (F = 20.37, p = 0.0001), and particularly from H. tuberculata (F = 57.16, p = 0.0001). Finally, multivariate analysis separates H. lamellosoides from H. tuberculata (F = 20.43; p = 0.0001; Fig. 5.1; Table 2).
(1) nMDS ordination (stress = 0.16) of the resemblance matrix for fossil and extant abalone of Europe and West Africa, where shaded ellipses indicate the 95% confidence interval of group (species) centroids, and plot characters indicate data source. (2–8) Associations between ordination structure and morphometrics influencing this structure, where the thin black lines illustrate: (2) length; (3) height:length ratio; (4) width:length ratio; (5) height:width ratio; (6) relative spire position; (7) total tremata; (8) open tremata.
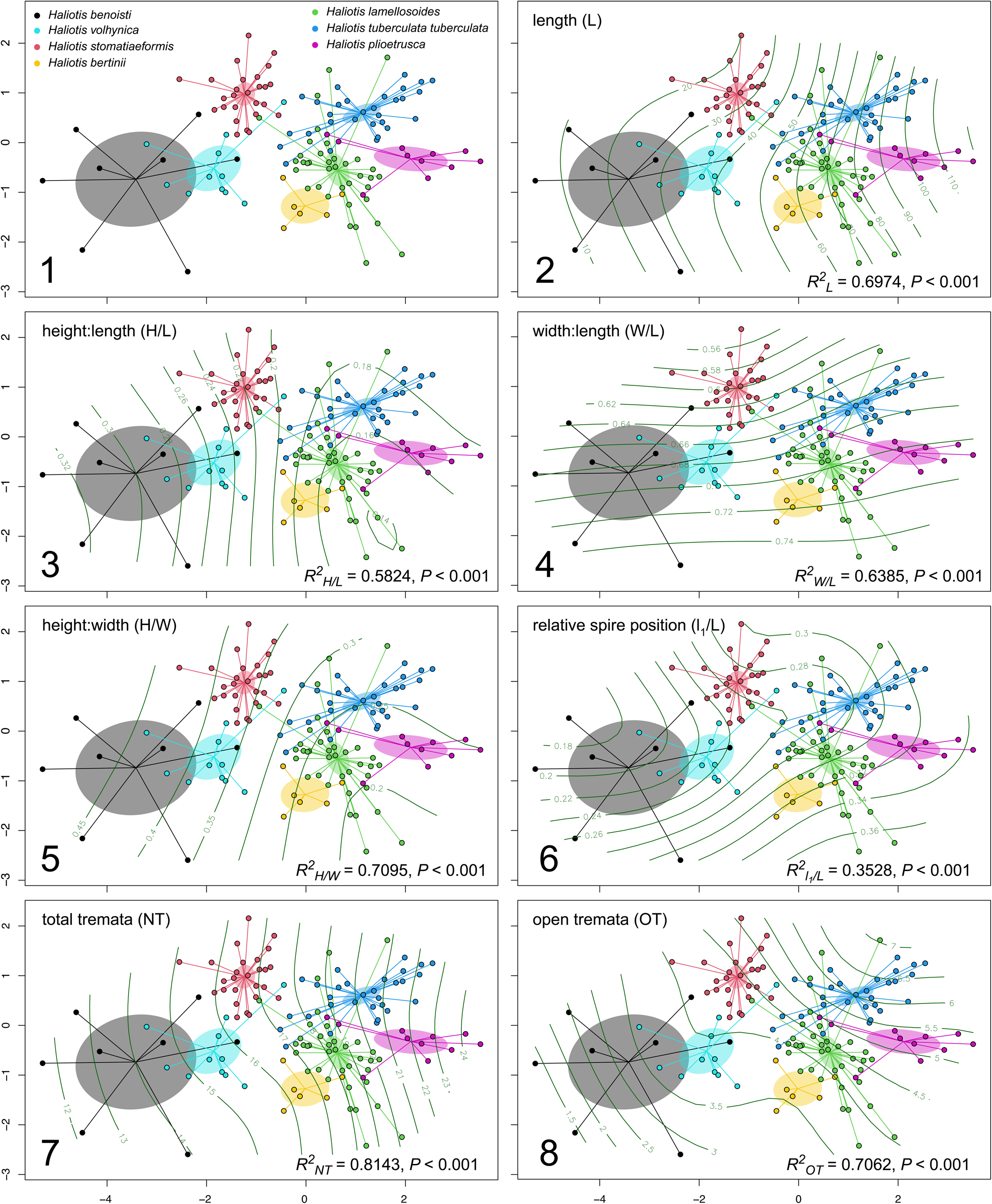
Results of pairwise comparisons testing the hypotheses that there were no differences in central tendency (i.e., centroid) of shell form among the studied extant abalone groups (species or subspecies of fossil and extant European abalone). Statistic F is the ratio of the amount of variation between versus within groups, with the numerator and denominator each weighted by their degrees of freedom. It is 0 or positive, with larger values corresponding to larger proportional importance of the grouping factor. Holm-adjusted probability that the distance between centroids arose by random chance (p) is presented.
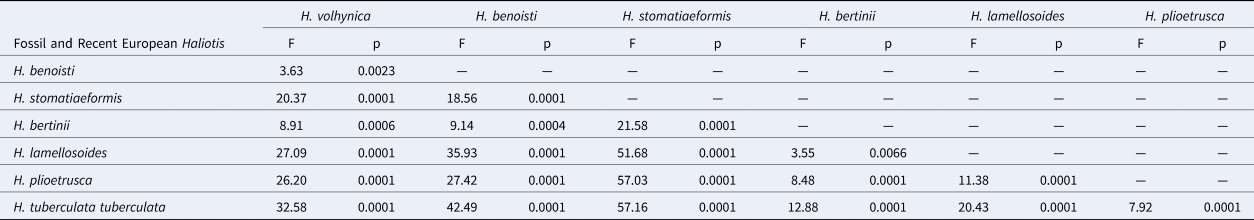
Similarly to the results obtained from the first dataset, also working with fossil taxa morphometrics considered representative of shell form significantly influenced the ordination structure (Fig. 5.2–5.8), with some differences in rank. The most important morphometric was NT (R2 = 0.81, p < 0.001), followed by H/W (R2 = 0.70, p < 0.001), OT (R2 = 0.70, p < 0.001), L (R2 = 0.70, p < 0.001), W/L (R2 = 0.64, p < 0.001), H/L (R2 = 0.58, p < 0.001), l1/L (R2 = 0.35, p < 0.001), and w1/W (R2 = 0.12, p < 0.001; not shown in Fig. 5). Univariate analysis expressed through bloxplots (Fig. 6) underline that the relative position of the spire (morphometrics l1/L and w1/W) and the number of open tremata (OT) do not allow us to differentiate among abalone species (Fig. 6.5–6.8), whereas all other morphometrics clearly separate Miocene and Pliocene European abalones, with extant H. stomatiaeformis being more similar to the first, and H. t. tuberculata to the second.
Box plots showing univariate comparisons among European known fossil and extant abalone species. (1) Shell length. (2) Height:length ratio. (3) Width:length ratio. (4) Height:width ratio. (5) Relative spire position with respect to L. (6) Relative spire position with respect to W. (7) Total number of tremata. (8) Open tremata. Boxes illustrate first and third quartile as box edges and median as central line.
Ultimately, from the associations between each morphometric and nMDS plot configurations (Figs 3.2–3.8, 5.2–5.8), relative differences in shell form could be inferred, analogously to results obtained by applying the same technique to cowries (Southgate and Militz, Reference Southgate and Militz2023), confirming the general utility of multivariate morphometric methods for statistical comparison of shell form between gastropod species (and to a lesser extent to subspecies). The two sets shared NT, H/W, OT, and L among the most informative morphometrics.
Systematic paleontology
Family Haliotidae Rafinesque, Reference Geiger1815
Genus Haliotis Linnaeus, Reference Linnaeus1758
Type species
Haliotis asinina Linnaeus, Reference Linnaeus1758 Recent, Eastern Indian Ocean to the Central Pacific, by subsequent designation Montfort, Reference Monfort1810.
Remarks
The supraspecific taxonomy of haliotids is problematic, and the use of a single genus-level taxon Haliotis is warranted (Geiger and Owen, Reference Geiger and Owen2012).
Haliotis benoisti Cossmann, Reference Cossmann1896
Figures 7, 8.1–8.4
- Reference Cossmann1896
Haliotis benoisti Cossmann, p. 22, pl. 5, figs. 14, 15.
- Reference Bial de Bellerade1903
Haliotis neuvillii Bial de Bellerade, p. 196.
- Reference Cossmann and Peyrot1917
Haliotis benoisti; Cossmann and Peyrot, p. 224, pl. 3, figs. 7–9, pl. 10, fig. 36.
- Reference Cossmann1918
Haliotis benoisti; Cossmann, p. 314, pl. 10, figs. 38, 39.
- Reference Geiger and Groves1999
Haliotis tuberculata volhynica Eichwald; Geiger and Groves, p. 875.
- Reference Lozouet, Lesport and Renard2001b
Haliotis (Sulculus) benoisti; Lozouet et al., p. 16, pl. 3, fig. 4.
(1–5) Haliotis benoisti Cossmann, Reference Cossmann1896. (1–3) France, Gironde, Martillac, Miocene, Burdigalian. (1–3) MHNBx 2014.10.3910, L 25 mm, W 16 mm. (4, 5) Original labels. Scale bars = 5 mm (left) and 10 mm (right).

(1–4) Haliotis benoisti Cossmann, Reference Cossmann1896. (1, 2) France, Vienne, southwest of Ambèrre, Moulin-Pochard, alongside D24, Miocene, Langhian, NHMR 6914, L 6 mm, W 4 mm. (3, 4) France, Gironde, Mérignac, Miocene, Aquitanian, holotype, MNHN.F.J04614, L 4 mm, W 2 mm. (5–16) Haliotis volhynica Eichwald, Reference Eichwald1829. (5, 6) Austria, Horn District, Gauderndorf, Miocene, Badenian, NHMW A 629, L 5 mm, W 4 mm. (7, 8) Ex Haliotis monilifera Michelotti, Reference Michelotti1847. Italy, Piedmont, Torino Hills, Miocene, Burdigalian. (7) MRSN BS.082.01.008, L 21 mm, W 15 mm. (8) MRSN BS.082.01.011, L 19 mm, W 15 mm. (9–11) Ex Haliotis tuberculata var. tauroparva Sacco, Reference Sacco1897. Torino Hills, Miocene, Burdigalian. (9, 10) MRSN BS.082.01.002, L 21 mm, W 14 mm. (11) MRSN BS.082.01.001, L 26 mm, W 20 mm. (12) Ukraine, Ternopil, Maksymivka, Miocene, Badenian, IGF 105317, L 23 mm, W 17 mm. (13) IGF 105318, L 27 mm, W 20 mm. (14, 15) IGF 105319, L 26 mm, W 19 mm, H 6 mm. (16) IGF 105320, L 52 mm, W 36 mm, H 17 mm. Unless otherwise indicated, scale bars = 10 mm.
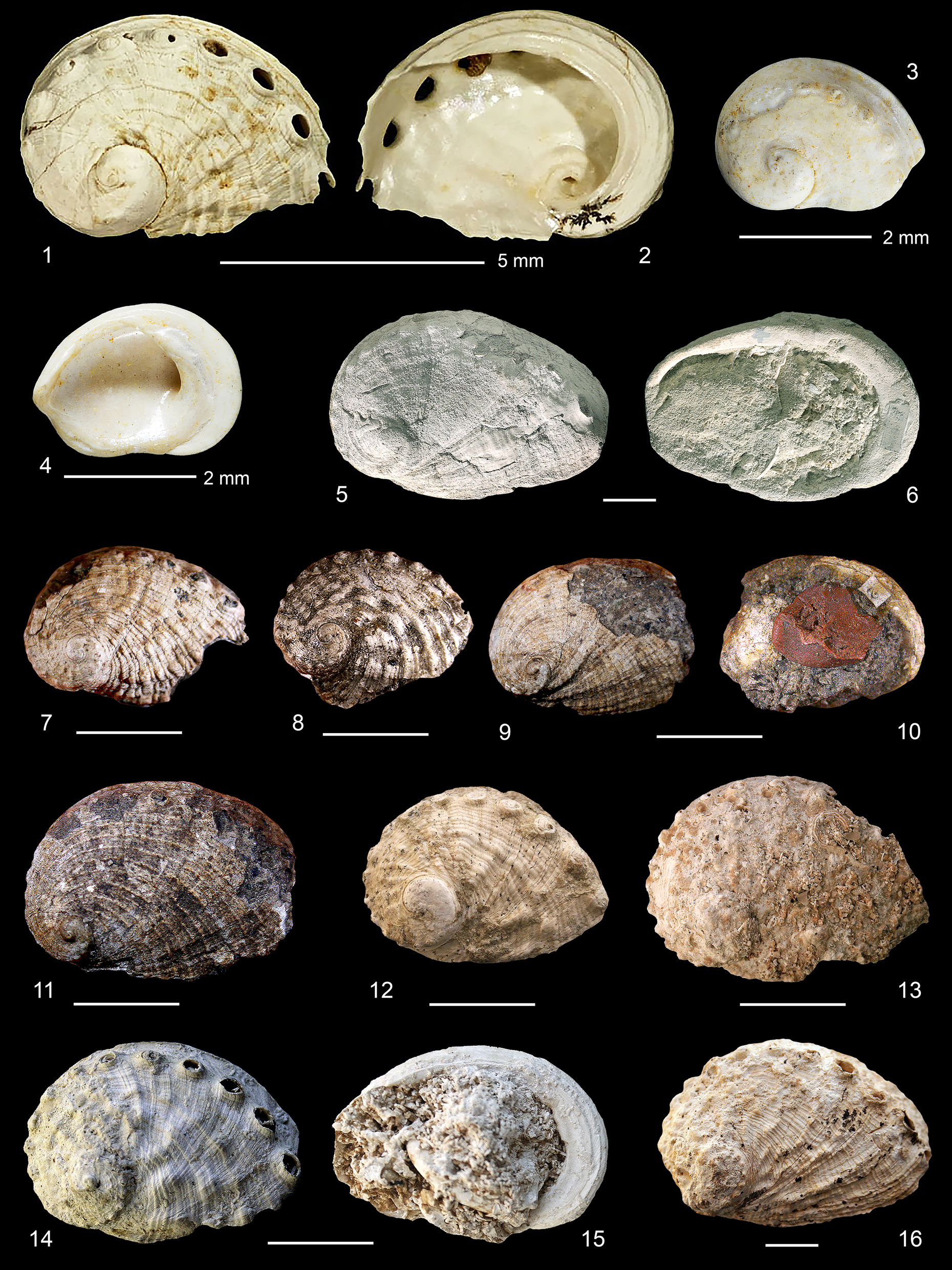
Type material
Holotype, MNHN. F. J04614, L 4.0 mm (Fig. 8.3– 8.4). Paratypes (four specimens): MNHN.F.J04615. Type locality: France, Gironde, Mérignac, Aquitanian Basin, early Miocene, Aquitanian.
Occurrence
France: from late Oligocene to early Miocene (Aquitanian–Burdigalian), Lariey, Léognan, Martillac, Mérignac, Western Atlantic France, Aquitanian Basin.
Description
Small, ear-shaped shell (Lmax = 25 mm) with rapidly increasing whorls. Spire not elevated, apex in subcentral position located 30–35% from the posterior margin. Dorsal surface ornamented with seven to eight well-defined spiral threads in early teleoconch, doubling in number in adult shell by the addition of secondary threads; spiral elements intersected by growth marks in early teleoconch, by closely spaced imbricate scales in late teleoconch. Elongated and slightly elevated tremata: five to six open, up to 19 total in larger shells. Thin, smooth spiral cord between the row of tremata and the peripheral carina. Flattened or slightly convex base with two to three fine parallel cords to columella. Columellar callus narrow, flattened.
Additional specimens
France, Miocene: ACC, three specimens, Landes, Meilhan carrière Vives (Burdigalian); ACC, two specimens, Landes, Campagne carrière Vives (Burdigalian); ACC, three specimens, Gironde, Pessac Lorient (Aquitanian); ACC, one specimen, Gironde, Martillac le Breyra (Burdigalian); MHNBx, one specimen, Gironde, Martillac (Burdigalian) (figured in Cossmann and Peyrot, Reference Cossmann and Peyrot1917, pl. 10, fig. 3); NHMR, one specimen, Nouvelle-Aquitaine, Vienne, Moulin Pochard (Langhian); JLC, one incomplete specimen, Gironde, Lariey (Aquitanian) (Lozouet et al., Reference Lozouet, Lesport and Renard2001b, p. 16, pl. 3, fig. 4).
Remarks
The species was introduced on the basis of five abraded specimens that are probably only fragments of a much larger original shell (see Cossmann, Reference Cossmann1896). In a second paper, an adult specimen was figured (Cossmann and Peyrot, Reference Cossmann and Peyrot1917). Geiger and Groves (Reference Geiger and Groves1999) attributed Oligocene and early Miocene abalones of Aquitaine to H. volhynica Eichwald, Reference Eichwald1853, in turn considered subspecies of H. tuberculata (Linnaeus, Reference Linnaeus1758). The multivariate analysis of shell forms based on type material and additional shells from several French localities confirms the separation from H. volhynica. Following Lozouet et al. (Reference Lozouet, Lesport and Renard2001b), Haliotis benoisti is considered valid and applied to Haliotis from the late Oligocene and early Miocene of Aquitaine. The Oligocene record is from St.-Paul-Les-Dax, France (Geiger and Groves, Reference Geiger and Groves1999, and references therein).
Haliotis volhynica Eichwald, Reference Eichwald1829
Figures 8.5–8.16, 9.1–9.5
- Reference Owen and Berschauer1829
Haliotis volhynica Eichwald, p. 294, pl. 5, fig. 18.
- Reference Michelotti1847
Haliotis monilifera Michelotti, p. 167, pl. 6, figs. 12, 12a.
- Reference Hörnes1856
Haliotis volhynica; Hörnes, p. 510, pl. 46, fig. 26.
- Reference Sacco1897
Haliotis tuberculata var. tauroparva Sacco, p. 5, pl. 1, figs. 1–3.
- Reference Sacco1897
Haliotis monilifera; Sacco, p. 7, pl. 1, figs. 9–14.
- Reference Sacco1897
Haliotis ? anomiaeformis Sacco, p. 7, pl. 1, fig. 15 (nomen dubium).
- Reference Friedberg1928
Haliotis volhynica; Friedberg, p. 530, pl. 34, figs. 8, 9.
- Reference Davidaschvili1937
Haliotis volhynica; Davidaschvili, p. 540, pl. 1, fig. 5.
- Reference Glibert1949
Haliotis sp. Glibert, p. 12, pl. 1, fig. 1.
- Reference Csepreghy-Meznerics1954
Haliotis tuberculata lamellosoides; Csepreghy-Meznerics, p. 10, pl. 1, fig. 24.
- Reference Korobkov1955
Haliotis (Haliotis) volhynica; Korobkov, pl. 2, fig. 3.
- Reference Kojumdgieva and Strachimirov1960
Haliotis (Haliotis) tuberculata var. lamellosoides; Kojumdgieva and Strachimirov, p. 84, pl. 28, fig. 9.
- Reference Steininger1963
Haliotis sp. Steininger, p. 37, pl. 12, fig. 1.
- Reference Strausz1966
Haliotis tuberculata volhynica; Strausz, p. 26, fig. 16c.
- Reference Bielecka1967
Haliotis volhynica; Bielecka, p. 132, pl. 8, figs. 3, 4 (fide Bałuk, 1975).
- Reference Zelinskaya, Kulichenko, Makarenki and Sorochan1968
Haliotis volhynica; Zelinskaya et al., p. 95, pl. 27, fig. 1.
- Reference Baluk1975
Haliotis (Sulculus) sp. Baluk, p. 22, pl. 1, figs. 4, 5.
- Reference Krach1981
Haliotis tuberculata; Krach, p. 39, pl. 11, figs. 1–3.
- Reference Jakubowski and Musiał1979
Haliotis (Sulculus) volhynica; Jakubowski and Musiał, p. 61, pl. 5, fig. 5.
- Reference Martin, Merle, Cornée, Filipescu, Saint Martin and and Bucur2007
Haliotis tuberculata; Saint Martin et al., p. 43, fig. 5.
- Reference Górka, Studencka, Jasionovski, Hara, Wysocka and Poberezhskyy2012
Haliotis tubercolata; Górka et al., p. 163, figs. 7a, 15a, b.
- Reference Forli, Stalennuy and Dell'Angelo2015
Haliotis volhynica; Forli et al., p. 89, figs. 2–14.
- Reference Owen and Berschauer2017
Haliotis volhynica; Owen and Berschauer, p. 40, figs. 1–17; 1–15; 7; 10; 13; 16.
(1–4) Haliotis volhynica Eichwald, Reference Eichwald1829. Ukraine, Ternopil, Maksymivka, Miocene, Badenian, IGF 105321, L 57 mm, W 38 mm, H 15 mm. (5–8) Haliotis stalennuyi Owen and Berschauer, Reference Owen and Berschauer2017. Ukraine, Maksymivka, Miocene, Badenian. (5) IGF 105361, L. 60 mm, W 33 mm. (6, 7) IGF 105322, L 46 mm, W 27 mm. (8) Austria, NHMW, Miocene, Badenian, L 47 mm, W 34 mm. (9–16) Haliotis ovata Michelotti, Reference Michelotti1847. Italy, Torino Hills, Miocene, Burdigalian. (9–11) MRSN BS.082.01.007, L 28 mm, W 20 mm, H 9 mm. (12, 13) Ex Haliotis tuberculata ? var. perspirata Sacco, Reference Sacco1897. Italy, Torino Hills, Miocene, Burdigalian. MRSN BS.082.01.003, L 14 mm, W 9 mm, H 4 mm. (14–16) Ex Haliotis tuberculata var. tauroplanata Sacco, Reference Sacco1897. Italy, Torino Hills, Miocene, Burdigalian. MRSN BS.082.01.004, L 41 mm, W 24 mm, H 7 mm. Scale bars = 10 mm.

Type material
Holotype, NHMW A629. Type locality: Austria, north of Eggenburg, Gauderndorf, early Miocene (Mandic and Steininger, Reference Mandic and Steininger2003) (Fig. 8.5–8.6).
Occurrence
Haliotis volhynica Eichwald, Reference Eichwald1829 extends from the Western (Italy) to the Central and Eastern Paratethys (Austria, Romania, Bulgaria, Poland, Ukraine), ranging from the Burdigalian (Torino Hill, Italy) to the early Tortonian (Korytnica, Poland). It is particularly abundant in infralitoral organogenic limestones of Ukraine.
Description
Medium-sized, moderately convex oval shell (Lmax = 70 mm). Spire moderately elevated, apex in subcentral position located 30–35% from the posterior margin. Grainy dorsal surface with spiral cords more or less broken up by radial folded ridges. Oval tremata: four to five open, up to 17 total in adult shell. Concave peripheral area between row of tremata and a prominent peripheral carina running parallel to columella. Columellar callus narrow, flattened.
Additional specimens
RMQ, one specimen, Poland, Korytnica, late Miocene, early Tortonian; 30 specimens, Ukraine, Ternopil, Maksymivka quarry, middle Miocene, Badenian.
Other fossil material
MRSN BS.082.01.001 (Fig. 8.11), 002 (Fig. 8.9, 8.10), Italy, Piedmont, Torino Hills, early Miocene, Burdigalian; MRSN BS.082.01.008 (Fig. 8.7), 009, 010, 011 (Fig. 8.8), Italy, Piedmont, Torino Hills, early Miocene, Burdigalian; MZB, one fragmentary specimen, Italy, Piedmont, Torino Hills, early Miocene, Burdigalian.
(1–13) Haliotis lamellosoides Sacco, Reference Sacco1897. (1–3) Italy, Piedmont, Asti, Colli Astesi, Pliocene, Piacenzian. (1, 2) Lectotype, MRSN BS.082.01.005, L 37.2 mm, W 25.3 mm. MRSN Original label. (4, 6–8, 10–13) Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (4) IGF 105323, L 48 mm, W 34 mm. (6) IGF 105324, L 44 mm, W 31 mm. (7) IGF 105325, L 54 mm, W 36 mm. (8) IGF 105326, L 50 mm, W 35 mm. (10) IGF 105327, L 66 mm, W 46 mm. (11) IGF 105328, L 72 mm, W 49 mm. (12, 13) IGF 105329, L 72 mm, W 49 mm. (5, 9) Italy, Siena, Tuscany, Castelnuovo Berardenga, Il Campino, Pliocene (Piacenzian). (5) IGF 105330, L 39 mm, W 26 mm. (9) IGF 105331, L 50 mm, W 35 mm. Scale bars = 10 mm.

Remarks
Some authors considered H. volhynica Eichwald, Reference Eichwald1829 a synonym of the widespread extant Mediterranean and Atlantic H. tuberculata, either as a distinct subspecies (H. tuberculata volhynica; Strausz, Reference Strausz1966; Geiger and Groves, Reference Geiger and Groves1999) or even as H. tuberculata tuberculata (Górka et al., Reference Górka, Studencka, Jasionovski, Hara, Wysocka and Poberezhskyy2012). General shell form allows us to clearly separate the two species (Figs. 5, 6). The morphological characteristics of H. volhynica are fairly constant and distinct: the apex moved toward the geometric center (Fig. 6.2; studied specimens allowed us to cover a wide range of sizes, enabling us to exclude the possibility that this difference is due to anisometric growth), the more rounded and tighter early teleoconch, and the ornamentation, consisting of marked tuberculate spiral cords (Csepreghy-Meznerics, Reference Csepreghy-Meznerics1954; Zelinskaya et al., Reference Zelinskaya, Kulichenko, Makarenki and Sorochan1968; Krach, Reference Krach1981; Górka et al., Reference Górka, Studencka, Jasionovski, Hara, Wysocka and Poberezhskyy2012) differentiate it also from H. lamellosoides. The same diagnostic features are seen in molds (e.g., Friedberg, Reference Friedberg1928; Strausz, Reference Strausz1966). Haliotis tuberculata var. tauroplanata Sacco, Reference Sacco1897 (Fig. 9.14–9.16) and H. tuberculata var. lamellosoides Sacco, Reference Sacco1897 (Fig. 10.1–10.3), from the Miocene and the Pliocene of northern Italy, respectively, possess slender and subequal spiral cords with small tubercles, whereas H. volhynica is more rounded and usually bears four to five open tremata, versus six to seven in H. tuberculata. H. volhynica found at Maksymivka (Ukraine) are well characterized and show a small degree of variability (Owen and Berschauer, Reference Owen and Berschauer2017). Haliotis monilifera Michelotti, Reference Michelotti1847, with more marked spiral ornaments and slightly larger tubercles that cross the growth lines (Fig. 8.7, 8.8), shares with Haliotis volhynica the general shape, the flattened columellar side, and the analogous number of respiratory holes. The same characters are seen in Haliotis tuberculata var. tauroparva Sacco, Reference Sacco1897 (Fig. 8.9–8.11) and the isolated specimen of Haliotis anomiaeformis Sacco, Reference Sacco1897 (p. 7, pl. 1, fig. 15; specimen now lost [D. Ormezzano, MRSN, personal communication, 2019]; the poor material upon which H. anomiaeformis was established does not help in establishing its validity, so it is here regarded as nomen dubium). The east African Haliotis rugosa pustulata Reeve, Reference Reeve1846 (Owen, Reference Owen2013), recently introduced in the eastern Mediterranean via the Red Sea (Talmadge, Reference Talmadge1971 Geiger, Reference Geiger1998, Reference Geiger2000; Zenetos et al., Reference Zenetos, Gofas, Russo and Templado2004), and the Indo-Pacific Haliotis varia Linnaeus, Reference Linnaeus1758, share with H. volhynica the general shell outline, the pattern of spiral growth, the ornamentation with pustules and elongated tubercles, and the shape of the columellar callus.
(1–7) Haliotis lamellosoides Sacco, Reference Sacco1897. Italy, Tuscany, Siena, Castelnuovo Berardenga, Il Campino, Pliocene (Piacenzian). (1, 2) Pullus, IGF 105356, L 2.5 mm, W 1.72 mm. (3, 4) Juvenile specimen, IGF 105357, L 7 mm, W 5.2 mm. (5) Juvenile specimen with little red flammae, IGF 105358, L 9.3 mm, W 5 mm. (6) Juvenile specimen, IGF 105359, L 10.8 mm, W 8.1 mm. (7) Juvenile specimen, IGF 105360, L 15 mm, W 10.5 mm. Unless specified, scale bars = 10 mm.

Haliotis stalennuyi Owen and Berschauer, Reference Owen and Berschauer2017
Figure 9.6–9.8
- Reference Krach1981
Haliotis tuberculata tauroplanata; Krach, p. 40, pl. 11, figs. 4–7.
- Reference Forli, Stalennuy and Dell'Angelo2015
Haliotis sp.; Forli et al., p. 92, figs. 15–18.
- Reference Owen and Berschauer2017
Haliotis stalennuyi Owen and Berschauer, p. 40, figs. 1–6; 8, 9; 11, 12; 14, 15.
Type material
Holotype: NMNZ, M.321138. Type locality: Ukraine, Ternopil, Maksymivka quarry.
Occurrence
Paratethys: middle Miocene, Badenian; Ukraine, Ternopil, Maksymivka (Forli et al., Reference Forli, Stalennuy and Dell'Angelo2015; Owen and Berschauer, Reference Owen and Berschauer2017); East Poland, Węglinek (Krach, Reference Krach1981).
Description
Oblong shell of medium size (Lmax = 60 mm), somewhat flattened and barely arched. Spire moderately flat to slightly elevated, apex in marginal position located 15–20% from the posterior margin. Smooth dorsal surface with spiral ribs almost entirely absent, with the exception of early teleoconch, where thin ribs and small tubercles are present. Shallow oval tremata, four to five open. Slightly rounded area, with three to four smooth cords, from the row of tremata and the peripheral carina.
Additional specimens
Type locality, middle Miocene, Badenian, one specimen, ASC, and one specimen, MSNF; middle Miocene, Badenian, one specimen, NHMW.
Remarks
Haliotis stalennuy is considered to differ from H. vohlynica by its different L/W ratio (1.79 and 1.46 respectively; Owen and Berschauer, Reference Owen and Berschauer2017), giving rise to a wider early teleoconch, and by the different ornamentation, without radial folds and evident spirals cords. Differences in L/W ratio were not validated by the multivariate analysis due to lack of data (measures from only one specimen). In H. stalennuyi, the earliest teleoconch shows some slender spiral cords. Small tubercles are present in both species, but in H. stalennuyi they are present only in the first part of the teleoconch, whereas in H. volhynica they are present throughout life.
Haliotis ovata Michelotti, Reference Michelotti1847
Figure 9.9–9.16
- Reference Michelotti1847
Haliotis ovata Michelotti, p. 166.
- Reference Sacco1897
Haliotis tuberculata? var. perspirata Sacco, p. 5, pl. 1, fig. 4.
- Reference Sacco1897
Haliotis tuberculata var. tauroplanata Sacco, p. 6, pl. 1, fig. 5.
- Reference Sacco1897
Haliotis ovata; Sacco, p. 6, pl. 1, fig. 8.
- Reference Ferrero Mortara, Montefameglio, Pavia and Tamperi1984
Haliotis ovata; Ferrero Mortara et al., p. 276, pl. 51, fig. 6a, b.
Type material
MRSN, BS.082.01.007. Type locality: Italy, Piedmont, Torino Hills, Miocene, Burdigalian.
Occurrence
Central Mediterranean, Italy, Piedmont, known only for the early Miocene (Burdigalian) of the Torinese Hills.
Description
Small shell (Lmax = 40 mm), oval to oblong, moderately convex. Spire depressed, apex in peripheral position, located 10–15% from the posterior margin. Smooth dorsal surface with fine spiral striation. Tremata oval, not elevated, four open. Rounded peripheral area with four to five smooth cords, crossed by fine growth lines. Columellar callus large, flat.
Additional material
Haliotis tuberculata? var. perspirata Sacco, Reference Sacco1897, MRSN BS.082.01.003. Italy, Piedmont, Torino Hills, Miocene, Burdigalian. Haliotis tuberculata var. tauroplanata Sacco, Reference Sacco1897, MRSN BS.082.01.004. Italy, Piedmont, Torino Hills, Miocene, Burdigalian. NHMW, one specimen, Italy, Piedmont, Torino Hills, Miocene, Burdigalian.
Remarks
Michelotti (Reference Michelotti1847) wrongly figured a specimen of H. monilifera, not fitting with his description of H. ovata, which remains a valid species. The right type material was figured by Sacco (Reference Sacco1897) (Fig. 9.9–9.11). H. ovata is distinguished from H. tuberculata by its flattened profile and the larger, flat columella.
Haliotis torrei Ruggieri, Reference Ruggieri1990
1990 Haliotis (Sulculus) torrei Ruggieri, p. 351, figs. 2, 3.
Type material
Holotype not found in the Ruggieri collection hosted at MGG in Palermo (C. D'Arpa, personal communication, 2024). Type locality: Italy, Sicily, Palermo, Petralia Sottana, late Miocene, Tortonian/Messinian.
Occurrence
The age determination is dubious. The sedimentary facies associated with the fossil is a calcareous marl with the colonial coral Tarbellastrea sp. The molluscan association suggests a late Tortonian age (Ruggieri, Reference Ruggieri1990, p. 351), but Petralia reefal carbonates of the Terravecchia Formation have also been attributed to the early Messinian (Grasso and Pedley, Reference Grasso and Pedley1988; similarly to other analogous settings; Dominici et al., Reference Dominici, Benvenuti, Forli, Bogi and Guerrini2019).
Description
Small ear-shaped shell (Lmax = 33 mm). Spire elevated, apex in peripheral position located about 20% from the posterior margin. Sloping dorsal surface, first a little convex after becoming flat near the carina, tessellated by nine large, flat, spiral cords crossed with deep growth lines. Oval tremata, elevated, four open. From the row of tremata and the peripheral carina, concave area with four to five small cords crossed by fine growth lines. Sinuous arched base with columellar callus narrow, rounded.
Remarks
The species is known only from the now lost holotype, and since its published image does not allow definitive taxonomic statements, the species should be considered a nomen inquirendum.
Haliotis lamellosoides Sacco, Reference Sacco1897
Figures 10–15
- Reference Sacco1897
Haliotis tuberculata var. lamellosoides Sacco, p. 6, pl. 1, figs. 6, 7.
- Reference Inzani1983
Haliotis tuberculata lamellosoides; Inzani, p. 12, figs. 1–8.
- Reference Cavallo and Repetto1992
Haliotis tuberculata lamellosa; Cavallo and Repetto, p. 36, fig. 17.
- Reference Borghi and Vecchi1998
Haliotis tuberculata lamellosa; Borghi and Vecchi, p. 83, pl. 1, figs. 1, 2.
- Reference Lozano-Francisco and Vera-Peláez2002
Haliotis (Sulculus) quinquecentenaris Lozano-Francisco and Vera-Peláez, p. 159, figs. A–D.
- Reference Landau, Marquet and Grigis2003
Haliotis iberica Landau, Marquet, and Grigis, p. 11, pl. 2, figs. 3, 4.
- Reference Landau, Marquet and Grigis2003
Haliotis tuberculata; Landau, Marquet, and Grigis, p. 12, pl. 2, figs. 1, 2.
- Reference Chirli2004
Haliotis tuberculata lamellosa; Chirli, p. 35, pl. 11, fig. 11.
- Reference Vera-Peláez and Lozano-Francisco2022
Haliotis (Sulculus) quinquecentenaris Vera-Peláez in Vera-Paláez and Lozano-Franisco, p. 26, pl. 3, figs. 10–12.
- Reference Vera-Peláez and Lozano-Francisco2022
Haliotis tuberculata; Vera-Peláez, p. 26, pl. 3, fig. 13.
- Reference Vera-Peláez and Lozano-Francisco2022
Haliotis telescopica Vera-Peláez, p. 27, figs. 14, 15.
Type material
MRSN BS.082.01.005, Italy, Piedmont, Asti, Colli Astesi, Pliocene, Piacenzian; BS.082.01.006, fragmentary, same data.
Emended diagnosis
Large Haliotis with a flat and large, slightly concave columellar callus and 17–19 total tremata, the last four to five of which are open.
Occurrence
West and Central Mediterranean. Pliocene, Zanclean: Italy, Siena, Montalcino, Quercecchio. Piacenzian: Italy, Piedmont, Asti, Colli Astesi; Emilia Romagna, Piacenza, Castell'Arquato; Tuscany, Pisa, Balconevisi; Siena, Bibbiano, Borgatello, Il Campino, Melograni, Pietrafitta, Podere Sant'Uliviere, Poggio alla Fame, Ponte a’ Mattoni, Stroncoli. Spain: Malaga, Estepona.
Description
Ear-shaped to oval, large shell (Lmax = 130 mm). Spire moderately flat to slightly elevated, apex in peripheral position located about 15–20% from the posterior margin. Moderately convex dorsal surface with highly variable ornamentation, from very fine spiral striation to 30–44 prominent and regularly spaced spiral cords, more or less broken up by radially folded ridges. Up to 21 oval tremata of medium size, round, somewhat elevated; last four to five tremata open. Sloping area from the row of tremata to the peripheral carina, with four to five cords crossed by growth lines. Base of whorl flat or gently arched, with large, flat, columellar callus.
Additional material
Italy, Tuscany: Siena, Montalcino, Quercecchio, Pliocene, Zanclean: two specimens. Siena, Castelnuovo Berardenga, Stroncoli, Pliocene, Piacenzian: 80 specimens. Siena, Castelnuovo Berardenga, Il Campino, Pliocene, Piacenzian: 15 specimens. Siena, San Gimignano, Podere Sant'Uliviere, Pliocene, Piacenzian: three specimens. Spain: Malaga, Estepona, Pliocene, Piacenzian: three specimens.
Remarks
Sacco (Reference Sacco1897) proposed Haliotis lamellosoides as a variety of H. tuberculata and in connection with H. lamellosa Lamarck, Reference Lamarck1822. Inzani (Reference Inzani1983) separated the first two at the subspecies level. H. lamellosoides is here considered distinct from H. tuberculata Linnaeus, Reference Linnaeus1758 due to the flatter and larger, slightly concave columellar callus, the less-distinctly auriform outline, and the wider variety of ornaments. H. lamellosoides has a lower total number of tremata (17–19 versus 19–21 in H. tuberculata; Fig. 6.7) and lower number of open tremata (four on average versus six to seven in most H. tuberculata; Fig. 6.8). Specimens collected at Stroncoli show a wide range of variability (see juvenile, colored, and teratological specimens in Figs. 11.1–11.7, 15.16–15.19), with no solutions of continuity. This range includes forms described in the Piacenzian (Pliocene) of Estepona (Spain) and recently introduced as separated species. Larger and more rounded Terre Rosse specimens with wavy and interrupted spiral ornaments match Haliotis iberica Landau, Marquet, and Grigis, Reference Landau, Marquet and Grigis2003 (specimens with prominent spiral cords widely spaced toward the periphery, interrupted at irregular intervals by radial fold, making them irregular and knobby; Figs. 12.11–12.13. 13.1–13.6. 14.1–14.5). Other specimens match with Haliotis quinquecentenaris Lozano-Francisco and Vera-Peláez, Reference Lozano-Francisco and Vera-Peláez2002 (specimens with very fine spiral cords, nearly smooth; Fig. 15.10–15.15) and others with Haliotis telescopica Vera-Peláez in Vera-Paláez and Lozano-Francisco, Reference Vera-Peláez and Lozano-Francisco2022, both also from Estepona. All the preceding reports from Estepona, southern Spain—here referred to H. lamellosoides—and its abundance in Tuscany, confirm that the species was widespread in the Mediterranean during the Piacenzian.
(1–13) Haliotis lamellosoides Sacco, Reference Sacco1897. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1) IGF 105362, L 42 mm, W 31 mm. (2) IGF 105363, L 30 mm, W 21 mm. (3) IGF 105364, L 40 mm, W 31 mm. (4, 5) IGF 105365, L 46 mm, W 30 mm. (6) IGF 105366, L 52 mm, W 36 mm. (7, 8) GF 105367, L 53 mm, W 35 mm. (9) IGF 105368, L 44 mm, W 30 mm. (10) IGF 105369, L 68 mm, W 46 mm. (11) IGF 105370, L 79 mm, W 54 mm. (12, 13) IGF 105371, L 80 mm, W 56 mm. Scale bars = 10 mm.

(1–7) Haliotis lamellosoides Sacco, Reference Sacco1897. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1) IGF 105372, L 95 mm, W 72 mm. (2) IGF 105373, L 67 mm, W 47 mm. (3) IGF 105332, L 73 mm, W 53 mm. (4) IGF 105374, L 97 mm, W 72 mm. (5) IGF 105333, L 77 mm, W 59 mm. (6) IGF 105375, L 102 mm, W 72 mm. (7) IGF 105334, L 54 mm, W 36 mm. Scale bars = 10 mm.

(1–6) Haliotis lamellosoides Sacco, Reference Sacco1897. (1, 3–4, 6) Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1) IGF 105335, L 94 mm, W 67 mm. (3) IGF 105336, L 111 mm, W 86 mm. (4) IGF 105376, L 52 mm, W 37 mm. (6) IGF 103388, L 88 mm, W 65 mm. (2) Italy, Tuscany, Siena, Castelnuovo Berardenga, Il Campino, Pliocene (Piacenzian), IGF 105337, L 58 mm, W 42 mm. Italy, Tuscany, Siena. Montalcino, Quercecchio, Pliocene (Zanclean). (5) IGF 105338, L 76 mm, W 59 mm. Scale bars = 10 mm.

(1–19) Haliotis lamellosoides Sacco, Reference Sacco1897. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1, 2) IGF 105377, L 44 mm, W 29 mm. (3) IGF 105339, L 53 mm, W 35 mm. (4) IGF 105340, L 45 mm, W 30 mm. (5, 6) IGF 105341, L 23 mm, W 16 mm. (7) MFC F036B 2901.53, L 54 mm, W 36 mm. (8) IGF 105378, L 44 mm, W 29 mm. (10) IGF 105379, L 44 mm, W 30 mm. (11, 12) IGF 105342, L 46 mm, W 31 mm. (14) IGF 105343, L 42 mm, W 24 mm. (15) IGF 105344, L 31 mm, W 21 mm. (16) Colored specimen, IGF 105380, L 37 mm, W 27 mm. (17) Teratological specimen, IGF 105345, L 63 mm, W 41 mm. (18) Teratological specimen, IGF 105381, L 53 mm, W 35 mm. (19) Colored specimen (highlighted color), IGF 105382, L 34 mm, W 23 mm. (13) Italy, Tuscany, Siena, Castelnuovo Berardenga, Il Campino, Pliocene (Piacenzian). IGF 105346, L 38 mm, W 24 mm. (9) Spain, Malaga, Estepona, Pliocene (Zanclean-Piacenzian), IGF 105387, L 51 mm, W 31 mm. Scale bars = 10 mm.

More convex, slightly smaller Terre Rosse specimens, bearing uniform spiral cords and lacking wavy plicae (Fig. 15.1–15.9), resemble Haliotis tuberculata coccinea Reeve, Reference Reeve1846, presently living in the Canary Islands and the Albora Sea. Molecular data indicate, however, that the latter diverged from a parental stock of H. tuberculata about 1.5 Ma (Van Wormhoudt et al., Reference Van Wormhoudt, Gaume, Le Bras, Roussel and Huchette2011), in the early Pleistocene. This datum is consistent with the Late Pleistocene report of H. tuberculata coccinea from Santa Maria Island, Azores (Avila et al., Reference Avila, Amen, Azevedo, Cachão and Garcia-Talavera2002), but not with the Tortonian, tentative identification of Haliotis from St-Clément-de-la-Place, Maine-et-Loire (northwestern France) (Landau et al., Reference Landau, Van Dingenen and Ceulemans2017, pl. 11, fig. 1).
Haliotis bertinii Forli, Dell'Angelo, Ciappelli, and Taviani, Reference Forli, Dell'Angelo, Ciappelli and Taviani2003
Figures 16.14, 16.15, 17.1–17.5
- Reference Michelotti1952
Haliotis tubercolata var. monilifera Michelotti in Lecointre, p. 88, pl. 28, fig. 14.
- Reference Forli, Dell'Angelo, Ciappelli and Taviani2003
Haliotis bertinii Forli et al., p. 149, pl. 1, figs. 1–6, pl. 2, figs. 1–10.
- Reference Chirli and Linse2011
Haliotis bertinii; Chirli and Linse, p. 30, pl. 2, fig. 5a–c.
- Reference Juárez-Ruiz and Mas2022
Haliotis bertinii; Juárez-Ruiz and Mas, p. 243, fig. 10A,B.
(1–13) Haliotis tuberculata tuberculata Linnaeus, Reference Linnaeus1758. (1–6) Italy, Lombardia, Milano, San Colombano, early Pleistocene (Gelasian). Ex Haliotis prisca De Cristofori and Jan, Reference De Cristofori and Jan1832. (1) Holotype, MSNM i 4288, L 57 mm, W 37 mm. (2, 3) MSNF IGF 105221, L 46 mm, W 30 mm. (4, 5) MSNF IGF 105222, L 40 mm, W 31 mm. (6) IGF 102117A, L 35 mm, W 26 mm. (7) Italy, Tuscany, Livorno, Rosignano Marittimo, Early Pleistocene (Calabrian), IGF 105348, internal cast, L 37 mm, W 23 mm. (8) Italy, Latina, Foce Verde, Late Pleistocene (Euthyrrenian). IGF 105347, L 19 mm, W 11 mm. (9–12) Italy, Calabria, Reggio Calabria, Bovetto, Late Pleistocene (Euthyrrenian). (9, 10) IGF 105349, L 30 mm, W 20 mm. (11, 12) IGF 105350, L 32 mm, W 20 mm. (13) Italy, Tuscany, Livorno, Lazzeretto, Middle Pleistocene. CBC, L 43 mm, W 27 mm. (14, 15) Haliotis bertinii Forli, Dell'Angelo, Ciappelli, and Taviani, Reference Forli, Dell'Angelo, Ciappelli and Taviani2003. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). IGF 105383, incomplete specimen, L 34 mm, W 29 mm. Scale bars = 10 mm.

(1–5) Haliotis bertinii Forli, Dell'Angelo, Ciappelli, and Taviani, Reference Forli, Dell'Angelo, Ciappelli and Taviani2003. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1, 2) IGF 105351, L 104 mm, W 68 mm. (3) Juvenile specimen, IGF 105384, L 12 mm, W 8 mm. (4) Juvenile specimen, IGF 105352, L 44 mm, W 30 mm. (5) IGF 105353, incomplete specimen, L 86 mm, W 67 mm. (6, 7) Haliotis plioetrusca n. sp. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (6) Paratype 1, IGF105219, L 115 mm, W 80 mm. (7) Paratype 6, IGF 105234, L 57 mm, W 39 mm. Scale bars = 10 mm.

Type material
Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene, Piacenzian. Holotype, MZB 25046, L 81.2 mm, W 59 mm, H 17 mm. Paratype 1, MZB 25041, L 65 mm, W 46 mm, H 12 mm. Paratype 2, MZB 25048, incomplete specimen. Paratype 3, FCC, L 111 mm, W 82 mm, H 18.4 mm. Paratype 4, FCC, L 107 mm, W 78.8 mm, H 21 mm. Paratype 5, FCC, L 66.2 mm, W 46.4 mm, H 10.5 mm. Paratype 6, MRC, L 105 mm, W 87 mm.
Occurrence
Atlanto-Mediterranean species reported at Morocco, Agadir, Pliocene (Lecointre, Reference Lecointre1952); Spain, Baleares Islands, Pliocene, late Zanclean–Piacenzian, (Juárez-Ruiz and Mas, Reference Juárez-Ruiz and Mas2022); Italy, Siena, Stroncoli, Pliocene, Piacenzian (Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003); Greece, Rhodes Island, Pleistocene, Calabrian (Chirli and Linse, Reference Chirli and Linse2011).
Description
Ear-shaped, medium-large, and robust shell (Lmax = 107 mm). Spire not much elevated, apex in peripheral position located about 20% from the posterior margin. Dorsal surface slightly convex at first, then gently sloping, nearly flat, crossed by small, regularly spaced and slightly tuberculated spiral cords. Last cord with oval holes, four open, form strong carina with protruding edge. The space between this last and the lower carina is slightly inclined, concave, and smooth, crossed only by growth lines. Flat and widened columellar edge fusing with the outer lip. The lip margin is narrow, arched, and convex.
Additional material
Five specimens, from type locality.
Remarks
The most distinctive character of this species is the presence of very strong, separated, and scaly spiral cords, with very little intraspecific variability across the geographic distribution (Rhodos Island, Balearic Islands, Morocco) and in time (Pliocene–Pleistocene; Chirli and Linse, Reference Chirli and Linse2011; Juárez-Ruiz and Mas, Reference Juárez-Ruiz and Mas2022; Lecointre, Reference Lecointre1952) compared with that of H. tuberculata.
Haliotis plioetrusca new species
Figures 17.6, 17.7, 18, 19.1–19.4.
(1–8) Haliotis plioetrusca n. sp. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1, 2) Paratype 2, MSNF IGF 105220, L 130 mm, W 90 mm. (3) Holotype, MSNF IGF 105218, L 86 mm, W 57 mm. (4) Paratype 3, MSNF IGF 105231, L 70 mm, W 52 mm. (5) Paratype 4, MSNF IGF 105232, L 41 mm, W 28 mm. (6) Paratype 5, MSNF IGF 105233, L 46 mm, W 34 mm. (7–8) Haliotis marmorata Linnaeus, Reference Linnaeus1758. Ghana, Terna, Recent. MMK 4586, L 37 mm, W 25 mm. Scale bars = 10 mm.

(1–4) Haliotis plioetrusca n. sp. (1–3) Italy, Siena, Castelnuovo Berardenga, Stroncoli, Pliocene (Piacenzian). (1) Paratype 7, MSNF IGF 105235, L 128 mm, W 93 mm. (2) Paratype 8, MSNF IGF 105236, L 79 mm, W 56 mm. (3) Paratype 9, MSNF IGF 105237, L 135 mm, W 92 mm. Italy, Tuscany, Siena, Montalcino, Quercecchio, Pliocene (Zanclean). (4) Incomplete specimen, IGF 105385, L 73 mm, W 64 mm. (5–8) Haliotis marmorata Linnaeus, Reference Linnaeus1758. Ghana, Busua Island, Recent. (5, 6) IGF 105354, L 40 mm, W 26 mm. (7, 8) IGF 105355, L 25 mm, W 17 mm. Scale bars = 10 mm.

Type material and type locality
Holotype, MSNF IGF 105218, L 86 mm, W 57 mm. Paratype 1, MSNF IGF105219, L 115 mm, W 80 mm. Paratype 2, MSNF IGF 105220, L 130 mm, W 90 mm. Paratype 3, MSNF IGF 105231, L 70 mm, W 52 mm. Paratype 4, MSNF IGF 105232, L 41 mm, W 28 mm. Paratype 5, MSNF IGF 105233, L 46 mm, W 34 mm. Paratype 6, MSNF IGF 105234, L 57 mm, W 39 mm. Paratype 7, MSNF IGF 105235, L 128 mm, W 93 mm. Paratype 8, MSNF IGF 105236, L 79 mm, W 56 mm. Paratype 9, MSNF IGF 105237, L 135 mm, W 92 mm. Italy, Tuscany, Siena, Castelnuovo Berardenga, Stroncoli. Pliocene, Piacenzian.
Diagnosis
Large Haliotis with 20–25 tremata and fine, closely spaced spiral threads crossed by minute imbricate scales.
Occurrence
Known only for the Tuscan Pliocene, at the type locality (Piacenzian), Siena, and near Montalcino, Siena, at Quercecchio (Zanclean).
Description
Large (up to 140 mm) auriform shell with a small, not elevated spire. Outer surface slightly convex, crossed by many spiral threads, subequal, equidistant. Tightly spaced growth increments cross spiral ornaments, forming minute, forward-leaning imbricate scales. Last five tremata open, with edge slightly elevated. Columellar callus wide, expanded, flat, connecting with convex, thin, crenulated outer lip. Internal surface smooth, without muscle scar.
Etymology
Named after the Pliocene Epoch and Latin adjective etruscus, derived from Etruria, an ancient Latin name for Tuscany.
Additional material
Three specimens, same locality as the holotype, including MZB, two incomplete specimens; BDA, one specimen. MCC, two incomplete specimens from Siena, Montalcino, Quercecchio, Pliocene, Zanclean.
Recent comparison material
H. marmorata Linnaeus, Reference Linnaeus1758: Ghana, Busua Island, MSNF IGF, two specimens. Ghana, Tema, MMK, two specimens. Gabon, Capo Esterias, MMK, two specimens (Fig. 1.5–1.8).
Remarks
H. plioetrusca n. sp. differs from all other Pliocene abalones by its larger size (Figs. 5.2, 6.1) and higher number of tremata (Figs. 5.7, 6.7). Superficially similar to H. marmorata from the tropical coasts of West Africa. The latter, however, is nearly half as large, with more spiral ornaments and a larger columellar callosity. Some incomplete specimens of H. plioetrusca have been found associated with H. lamellosoides also in a shoreface assemblage indicating a vegetated bottom (Quercecchio, Val d'Orcia basin, Zanclean, Pliocene; Dominici and Forli, Reference Dominici and Forli2021) (Fig. 19.4).
Haliotis stomatiaeformis Reeve, Reference Reeve1846
- Reference Reeve1846
Haliotis stomatiaeformis; Reeve, p. 73, fig. 74.
- Reference Philippi1848
Haliotis neglecta Philippi, p. 16.
- Reference Geiger and Owen2001
Haliotis stomatiaeformis; Geiger and Owen, p. 77, figs. 1–4.
- Reference Owen2003
Haliotis stomatiaeformis; Owen, p. 287, pl. 1, 2.
- Reference Geiger and Owen2012
Haliotis stomatiaeformis; Geiger and Owen, p. 130, pl. 65, figs. 1–14.
- Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022
Haliotis stomatiaeformis; Chiappa et al., fig. 1E 2024 Haliotis stomatiaeformis; Reitano et al., figs 7–12.
Type material
Lectotype (BMNH 1950.3.16.22-24) and paralectotypes (BMNH: Geiger and Owen, Reference Geiger and Owen2001).
Occurrence
The species is rare in the Mediterranean; shells are found only along the rocky coasts of Sicily and Malta, and living specimens were collected under stones 2–3 m deep. Fossil near Grammichele, Catania, Italy, Late Pleistocene (Reitano et al., Reference Reitano, Di Franco and Scuderi2024).
Description
Small to medium, oblong to ovate shell (Lmax = 45 mm), very convex. 20–37 smooth spiral cords between suture and row of tremata, stronger cords intercalated by one to three intermediary ones. Radially finely plicated, spire nearly terminal, elevated; three to five open tremata.
Remarks
One paratype is 45 mm long, but most specimens range 20–30 mm, with four open tremata on average (Geiger and Owen, Reference Geiger and Owen2001). Lives in sympatry with H. tuberculata, from which it can be distinguished: more elongated, more arched, strong spiral sculpture, no lamellae, smaller size (Geiger and Owen, Reference Geiger and Owen2012; Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). Erroneously reported by Reeve (Reference Reeve1846) from New Zealand, lives only in Malta and Sicily (rare).
Haliotis tuberculata Linnaeus, Reference Linnaeus1758
Figure 16.1–16.13
- Reference Linnaeus1758
Haliotis tuberculata; Linnaeus, p. 780.
- Reference Lamarck1822
Haliotis lamellosa Lamarck, p. 217.
- Reference Eichwald1829
Haliotis tuberculata; Eichwald, p. 294.
- Reference De Cristofori and Jan1832
Haliotis prisca De Cristofori and Jan, p. 3.
- Reference Reeve1846
Haliotis lamellosa; Reeve, pl. 14, figs. 14, 15.
- Reference Reeve1846
Haliotis speciosa Reeve, pl. 14, figs. 14, 15.
- Reference Reeve1846
Haliotis tuberculata; Reeve, pl. 39, fig. 34.
- Reference Reeve2001
Haliotis mykonosensis Owen et al., p. 301, figs. 1–4.
Type material
Six syntypes (LSL 576; Geiger and Owen, 20012).
Occurrence
H. tuberculata ranges the eastern Atlantic from the Channel, northwestern France, to Senegal, Gulf of Guinea, including Macaronesia, and the whole Mediterranean (Mgaya and Mercer, Reference Mgaya and Mercer1994; Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). Fossil reports range from the early (Gelasian, San Colombano, northern Italy) to the Late Pleistocene (129–11.7 ka; Avila et al., Reference Avila, Amen, Azevedo, Cachão and Garcia-Talavera2002).
Description
Large shell (Lmax = 123 mm; up to 102 mm in the Mediterranean; Geiger and Owen, Reference Geiger and Owen2012), from oval to ear-shaped, somewhat elongated, convexly depressed. Spire depressed to strongly elevated. Tremata medium sized, round, three to nine open, on average six. Deep spiral cords crossed by radial tubercles or lamellae, but highly variable from smooth with faint radial folds to sharp radial lamellae. Columella narrower than medium width.
Remarks
Haliotis tuberculata differs from H. lamellosoides by its narrower and somehow convex columella and by a higher number of tremata. Three subspecies are recognized on morphological grounds (Owen et al., Reference Owen, Ryall and Pan2015): H. tuberculata tuberculata (much larger than the other subspecies but also larger than H. lamellosoides; Fig. 4.1), H. tuberculata coccinea (Reeve, Reference Reeve1846), and H. tuberculata fernandesi Owen and Afonso, Reference Owen, Afonso, Geiger and Owen2012. A separation of the first two is confirmed on molecular grounds (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). The status of H. tuberculata fernandesi, however—largely overlapping in shell form with H. t. coccinea (Fig. 3), the two differing only in ornamentation (Owen et al., Reference Owen, Ryall and Pan2015)—needs a molecular validation. H. speciosa Reeve, Reference Reeve1846 from Senegal has been placed in synonymy with H. tuberculata (Owen, Reference Owen2006), an interpretation confirmed by the multivariate approach to shell form (Fig. 3).
Discussion
Adaptations in Mediterranean Haliotis
The site of Stroncoli provided the largest abundance of abalones among all known European fossil localities and the evidence of the coexistence of three species at one time and space, evidenced by pristine shells sampled in a laterally limited outcrop of an individual event bed. The co-occurrence here of well-preserved representatives in the families Patellidae and Haliotidae testifies to the proximity of the shoreline and the presence of a rocky substrate (Forli et al., Reference Forli, Dell'Angelo, Ciappelli and Taviani2003, Reference Forli, Dell'Angelo, Montagna and Taviani2004). Three species of Haliotis co-occurring in just one single shell bed is an exceptional feature even with respect to syntopic abalones in the Recent. Modern occurrences of Haliotis in the Mediterranean and eastern Atlantic indicate that, although two species may occur sympatrically in Sicily, Lampedusa and Malta, where the distribution of H. tuberculata overlaps with that of H. stomatiaeformis (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022), only one species is reported at any one site. The morphotype “H. mykonosensis,” with its thin, oblong to ovate, from flat to slightly convex shell, generally with five to six open tremata and an average adult size of about 40–48 mm (Crocetta and Rismondo, Reference Crocetta and Rismondo2009; Geiger and Owen, Reference Geiger and Owen2012), does not correspond to any of the Terre Rosse morphologies herein described, neither is its validity corroborated by molecular data (Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). More than one species may coexist at many other places, for example, in West Africa, where the area inhabited by H. tuberculata overlaps that of H. marmorata (Fig. 20), and at Rottnest Island, off northwestern Australia, Indian Ocean. where temperate seagrass and reef-building corals co-occur, contributing to a variety of microhabitats and where at least three species of abalones are reported (Geiger, Reference Geiger2000; Invert E Base, 2023; Wells et al., Reference Wells, Keesing, Gagnon, Bessey, Spilsbury and Irvine2023). At Port Alfred, along the coast of South Africa, up to five species of abalones can coexist in sympatry (Bester-van der Merwe et al., Reference Merwe, D'Amato, Swart and and Roodt-Wilding2012). Other discrete regions with endemic species, in the North Pacific (seven species) and New Zealand (three species), are similarly associated with temperate climates (Geiger, Reference Geiger2000; Estes et al., Reference Estes, Lindberg and Wray2005). Camouflage and cryptic behavior typical of abalones (Searcy-Bernal and Gorrostieta-Hurtado, Reference Searcy-Bernal and Gorrostieta-Hurtado2007; Cenni et al., Reference Cenni, Parisi, Scapini and Gherardi2010; Kim et al., Reference Kim, Lee and Yoo2020) in closely coexisting microhabitats of the indented and rugged late Pliocene coastline of Tuscany (Nalin et al., Reference Nalin, Ghinassi, Foresi and Dallanave2016) may thus explain the exceptionally high diversity encountered at Stroncoli. This is not unexpected, for high species richness of Mediterranean herbivore gastropods is associated with the mid-Piacenzian Warm Period (Dominici and Danise, Reference Dominici, Danise, Nawrot, Dominici, Tomašových and Zuschin2023).
Distribution of species of Haliotis in the Mediterranean Pliocene and on the modern coasts of southern Europe, the Middle East, the Red Sea, and West Africa. H. tuberculata coccinea is very common also in the Azores, not included in the map. H. tuberculata fernandesi is restricted to Cape Verde, also not included.
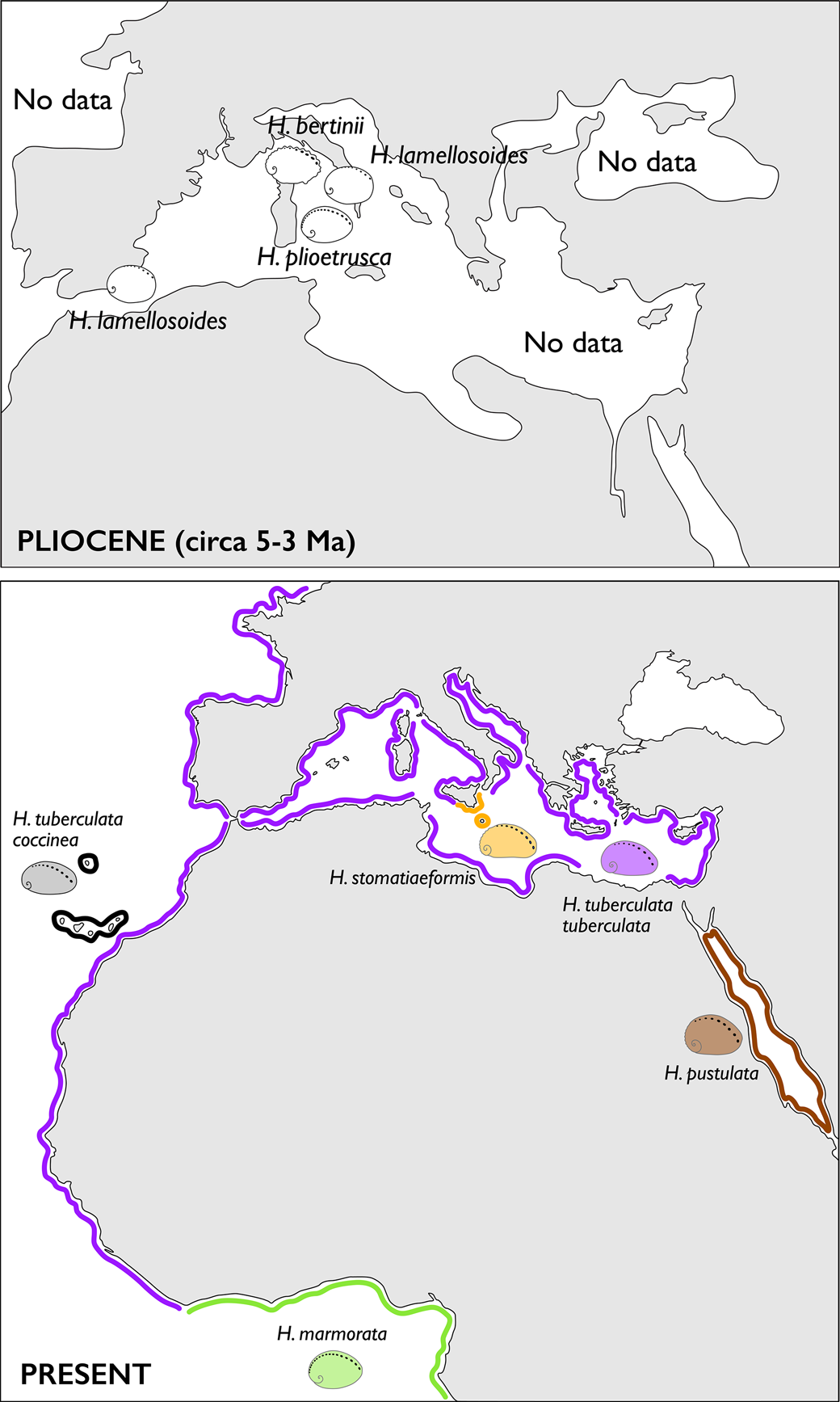
Relationships between Mediterranean and East Atlantic Haliotis.—The multivariate analysis of shell form and the systematic and stratigraphic review highlight the presence of two main groups of European fossil and Recent Haliotis. These data, based on a large number of specimens from many localities, coupled with known genetic relationships between extant forms, allow hypotheses on species-level evolutionary relationships. One group comprises specimens distributed in the Oligocene and Miocene of western and central Europe (H. benoisti and H. volhynica). The other connects Pliocene, Pleistocene, and Recent Mediterranean and West African species (H. lamellosoides, H. bertinii, H. plioetrusca and H. tuberculata; Fig. 5). The multivariate statistical comparison of shell form confirms that H. volhynica and H. lamellosoides represent two distinct species and that both can be clearly separated also from H. tuberculata tuberculata on the basis of the number of tremata and general size, larger in the second (Fig. 6). Both H. volhynica and H. lamellosoides average four open tremata (Fig. 6.8), but H. lamellosoides can be distinguished on a higher total number of tremata (Fig. 6.7).
The Miocene group includes (1) the possible ancestral form H. benoisti Cossmann, Reference Cossmann1896 from the Oligocene–early Miocene of the northeastern Atlantic coast, (2) the younger H. volhynica from the early Miocene of western Paratethys, expanding its range eastward in the middle Miocene, joined by (3) H. stalennuyi Owen and Berschauer, Reference Owen and Berschauer2017 (Figs. 21, 22). The multivariate analysis of shell form indicates that extant Mediterranean H. stomatiaeformis is more similar to Miocene H. benoisti and H. volhynica than to the sympatric H. tuberculata tuberculata (Fig. 5; Table 2; the latter with a record not older than the early Pleistocene). This could mean that the direct ancestor of H. stomatiaeformis is not H. tuberculata but an older form closer to the H. benoisti–H. volhynica lineage. The fact that in molecular phylogenies H. stomatiaeformis clusters with H. tuberculata and the Senegalese H. marmorata Linnaeus, Reference Linnaeus1758 (Van Wormhoudt et al., Reference Van Wormhoudt, Le Bras and Huscette2009; Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022) reveals only that the latter three extant species are ultimately rooted in the H. benoisti–H. volhynica lineage. The proximity in shell form of H. lamellosoides and H. tuberculata (Fig. 5) suggests that the first could be the direct ancestor of the second. A candidate ancestral form of all the three species found at Terre Rosse is H. ovata Michelotti, Reference Michelotti1847, found in the early Miocene of western Paratethys (Fig. 22). The high morphological disparity recorded in H. lamellosoides and its association with H. bertinii and H. plioetrusca roughly coincide with an interval of high overall Mediterranean species richness recorded around the Zanclean–Piacenzian passage and in the first part of the Piacenzian (Aguirre et al., Reference Aguirre, Cachão, Domènech, Lozano-Francisco Ma, Martinell, Mayoral, Santos, Vera- Peláez and Da Silva2005; Dominici and Danise, Reference Dominici, Danise, Nawrot, Dominici, Tomašových and Zuschin2023). H. bertinii and H. plioetrusca are now extinct. Tropical H. marmorata living in West Africa at subtropical and tropical latitudes (Fig. 20), limited southward by the Angola–Benguela Front, a major biogeographic boundary since the Miocene (Sessa et al., Reference Sessa, Callapez, Dinis and Hendy2013, shows superficial similarities with H. plioetrusca (Fig. 19), pointing to possible phylogenetic relationships. The smaller size of the extant species would confirm the tendency for ectotherms to be smaller in warmer habitats (Dominici et al., Reference Dominici, Fornasiero and Giusberti2020).
Oligocene–Miocene distribution of species of Haliotis in the Atlantic, Mediterranean, and Paratethys.
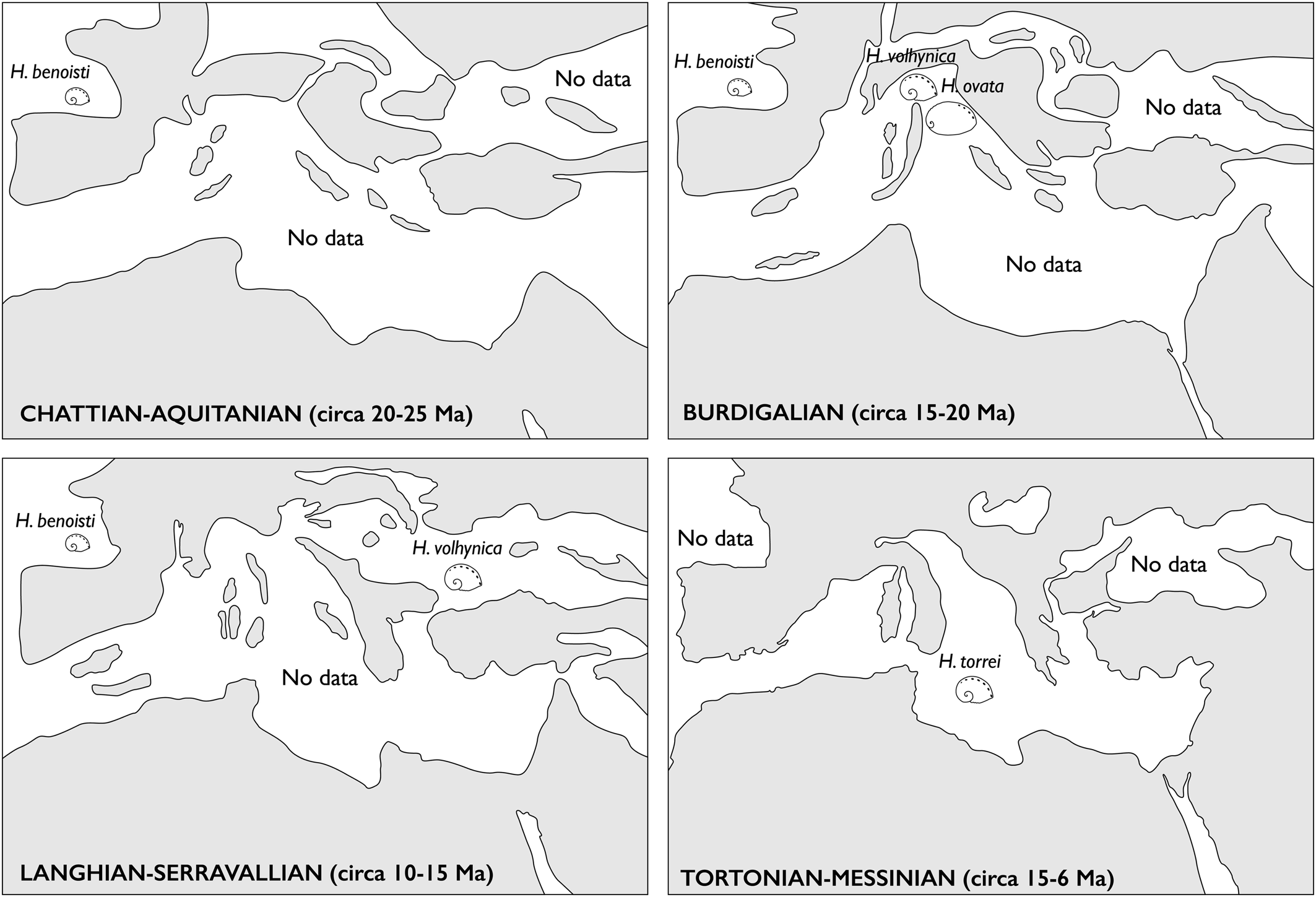
Species- and subspecies-level range of Haliotis in Europe during the Neogene. Tropical and subtropical species in red; temperate species in blue. Dashed lines indicate possible evolutionary relationships.
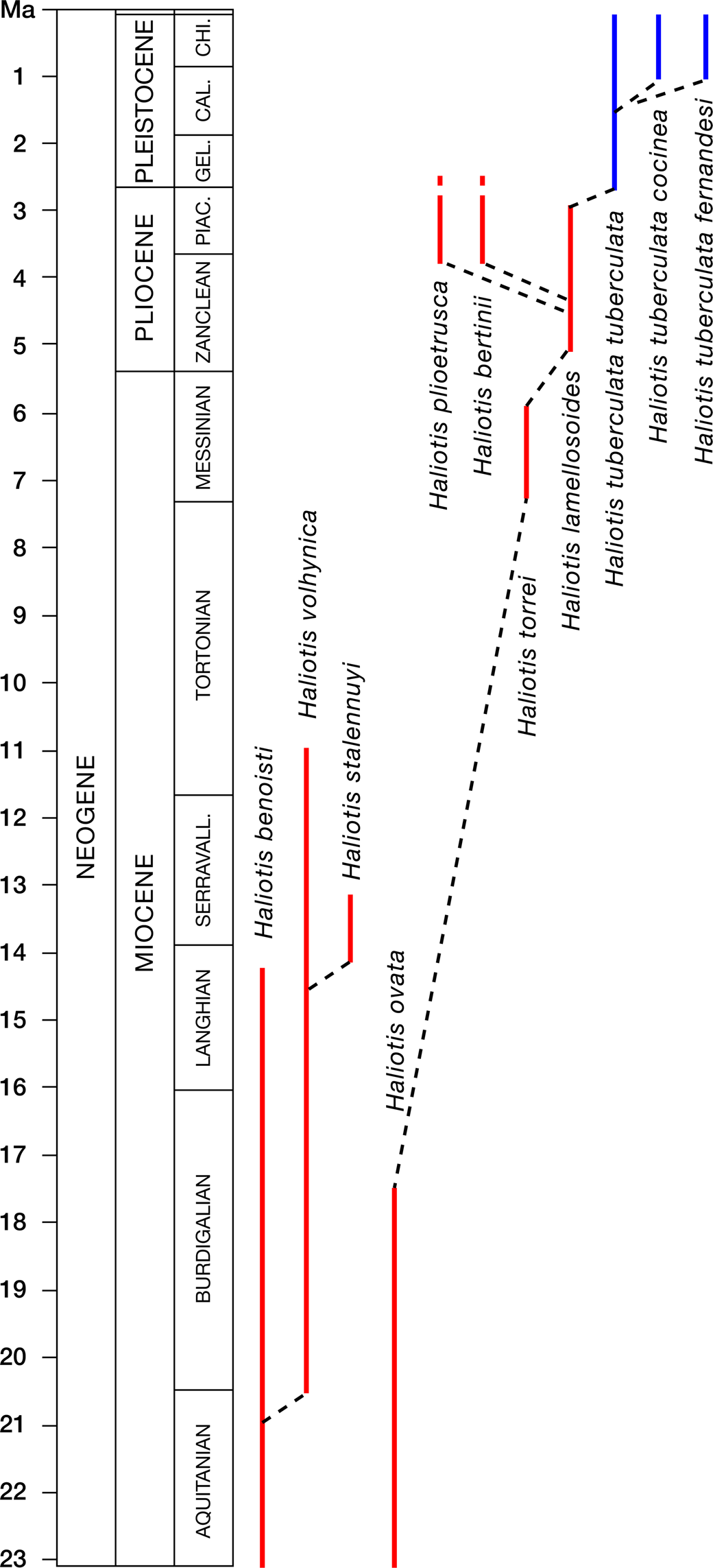
The contiguous, non-overlapping record of H. lamellosoides and H. tuberculata suggests that the second originated by anagenesis (Fig. 21), when Mediterranean climate shifted from tropical to temperate at the onset of the Northern Hemisphere Glaciation. H. tuberculata expanded its range from temperate regions southward along the Atlantic coast to its modern distribution. The present-day occurrence of endemic forms at Atlantic and Mediterranean islands (Fig. 20) indicates that species of Haliotis—like all stenotopic species confined to the shallow zone (Holland and Christie, Reference Holland and Christie2013; Dominici and Danise, Reference Dominici, Danise, Nawrot, Dominici, Tomašových and Zuschin2023)—are sensitive to successive eustatic cycles.
Morphological data retrieved from fossil shells thus show that a possible order of divergence of Mediterranean and eastern Atlantic abalones, from a longstanding, original stock of H. lamellosoides, is lamellosoides–plioetrusca by splitting, lamellosoides–tuberculata tuberculata– by anagenesis, and tuberculata tuberculata–tuberculata coccinea by splitting, a hypothesis corroborated by molecular data collected in living abalones (Van Wormhoudt et al., Reference Van Wormhoudt, Le Bras and Huscette2009, Reference Van Wormhoudt, Gaume, Le Bras, Roussel and Huchette2011; Chiappa et al., Reference Chiappa, Fassio, Corso, Crocetta, Modica and Oliverio2022). The fossil record indicates that Pleistocene climate change, as it did with other benthic mollusks (Mondanaro et al., Reference Mondanaro, Dominici and Danise2024), played a major role in the geographic distribution of European abalones (Roussel and Van Wormhoudt, Reference Roussel and Van Wormhoudt2017).
The eastern connection
Molecular phylogenetic analyses and chromosome evidence suggest that the modern biodiversity abalone hotspot, centered in the tropical Indo-Pacific (Geiger, Reference Geiger2000), originated from Tethyan ancestors (Geiger and Groves, Reference Geiger and Groves1999; Estes et al., Reference Estes, Lindberg and Wray2005; Bester-van der Merwe et al., Reference Merwe, D'Amato, Swart and and Roodt-Wilding2012). The oldest group of European fossils herein discussed hints at an established eastern connection between the Mediterranean and the Indo-Pacific province through H. volhynica. The youngest report of H. volhynica is from the early Tortonian of the Paratethys, well after the closure of the Tethyan seaway during the early Miocene, at around 19 Ma (Yasuhara et al., Reference Yasuhara, Huang, Reuter, Tian and Cybulski2022), and no evidence exists of extant Mediterranean descendants. Morphological affinities with recent immigrants from the Red Sea (small size, tight spiral growth originating shells with an oval outline, spiral ornaments with pustules or tubercles) are encountered with the East African and Indo-Pacific H. rugosa pustulata Reeve, Reference Reeve1846 and H. varia Linnaeus, Reference Linnaeus1758, presently ranging from Israel to East Africa, Persian Gulf, Red Sea, Madagascar, and Sri Lanka to Tonga, southern Japan, Philippines, and northwestern and western Australia (Geiger and Owen, Reference Geiger and Owen2012). Their fossil record is relatively young (Pleistocene of Egypt, Sudan, Zanzibar, and Tanzania for H. rugosa pustulata; Pliocene of Sri Lanka for H. varia; Geiger and Owen, Reference Geiger and Owen2012), and a phylogenetic relationship with H. volhynica can be hypothesized.
The middle Miocene (c. 16 Ma) Haliotis amabilis (Itoigawa and Tomida, Reference Itoigawa and Tomida1982) from Japan instead shares morphological characters, including general form, dimensions, ornaments, and the presence of a flat columellar callous, with H. ovata and H. lamellosoides (compare with Tomida et al., Reference Tomida, Okumura and Kaede2006, fig. 2.1–2.3), hinting at an older Miocene center of diffusion. The fossil record of other large herbivore gastropods (Dominici et al., Reference Dominici, Fornasiero and Giusberti2020) and large benthic species in general (Yasuhara et al., Reference Yasuhara, Huang, Reuter, Tian and Cybulski2022) corroborates the hypothesis of a Miocene Tethyan center of origin of the modern diversity hotspot. The oldest fossil record of the family (two Late Cretaceous species from California) points to an even older, north-eastern Pacific origin of the clade (Groves and Alderson, Reference Groves and Alderson2008). Expansion possibly took place during the Paleogene in a greenhouse world, via the Atlantic at the time connected to the Pacific (Barnet et al., Reference Barnet, Harper, LeVay, Edgar and Henehan2020).
Conclusions
New fossil finds of abalones in Tuscany add to the known Pliocene richness of Haliotis in the Mediterranean, peaking at a single site to the three species Haliotis lamellosoides, H. bertinii, and H. plioetrusca. A multivariate analysis of shell form of the European fossil and modern record of abalones, including morphometrics of 379 specimens from 17 countries in Europe and Africa, allows us to reassess the species- and subspecies-level taxonomy of Haliotis during the Neogene and the Quaternary. Merging new data with the known chronostratigraphic and paleogeographic distribution of fossil and extant species, two lines of descent are proposed. The first connects H. benoisti (Oligocene–early Miocene, northwestern Atlantic), H. volhynica (early–middle Miocene, Paratethys), H. stalennuyi (middle Miocene, Paratethys), H. torrei (late Miocene, Sicily), and H. stomatiaeformis (Recent, Sicily, Lampedusa, and Malta). The second possibly connects H. ovata (early Miocene, Paratethys), H. torrei (late Miocene), H. lamellosoides (Pliocene, Mediterranean), H. bertinii (late Pliocene, Mediterranean), H. plioetrusca (late Pliocene), H. marmorata (Recent, western Atlantic), H. tuberculata tuberculata (Gelasian–Recent, Mediterranean and western Atlantic), and H. tuberculata coccinea (late Pleistocene–Recent, western Atlantic). The available record justifies a hypothesis of diversification in the Mediterranean during a late Pliocene warmer interval, leading from H. lamellosoides to H. bertinii and H. plioetrusca, then from H. lamellosoides to H. tuberculata at the onset of climatic cooling, at the Pliocene–Pleistocene boundary.
Acknowledgments
Special thanks to A. Cluzaud (Pessac, Gironde, France), the personnel of the Muséum d'Histoire Naturelles de Bordeaux (France), J.-F. Lesport (Sainte-Hélène, Gironde, France), J.-M.l Pacaud (Muséum National d'Histoire Naturelle Paris, France), M. Harzhauser (Natural History Museum of Wien, Austria), A. Bonfitto, B. Sabelli (Museo dell'Evoluzione di Bologna, Italy), D. Ormezzano (Museo Regionale di Scienze Naturali di Torino, Italy), C. D'Arpa (Museo G.G. Gemmellaro, Palermo, Italy), G. Bini, and B. Santucci (Associazione Malakos, Città di Castello, Perugia, Italy) for their assistance with collections hosted in their facilities. We are indebted to D. Bertini (Firenze), C. Bogi (Livorno), C. Chirli (Tavarnelle, Firenze), F. Ciappelli (Calenzano, Firenze), M. Cresti (San Casciano, Firenze), B. Dell'Angelo (Genova), S. Gori (Livorno), M. Rocca (Torino), C. Sbrana (Livorno), and A. Ventura (Colle val d'Elsa, Firenze) for sharing the results of their fieldwork on Neogene outcrops in Italy, and M. McCulloch (Sidney), J. Trotter (Sidney), and B. Gualandi (Bologna) for the trip at Rottnest Island, Australia. A. Mondanaro (Florence) and A. Tomašových (Bratislava) helped with statistical analyses. Finally, our sincere thanks to an anonymous reviewer and D. Geiger, whose detailed reviews helped to improve the quality of the article. This is Ismar-CNR, Bologna, scientific contribution n. 2094.
Competing interests
The authors declare none.
Data Availability Statement
Data available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.4b8gthtnp.

























 Open access
Open access