



Introduction
The genus Monoplectanum Young 1969 is classified within the family Diplectanidae and is unique by having 1 squamodisc only, whereas other genera typically have 2 (Young, Reference Young1969). These monogeneans are ectoparasites found on the gills of fish and, to date, have been reported exclusively from sillaginid fishes. This genus currently includes only 2 previously described species: M. australe Young 1969 and M. youngi Hayward 1996.
Monoplectanum australe infects Sillago ciliata Cuvier, 1829, S. aeolus Jordan & Evermann, 1902, S. maculata Quoy & Gaimard, 1824, and S. burrus Richardson, 1842 from Moreton Bay, Shark Bay, Point Samson, Duyfken Point in Australia, and the upper Gulf of Thailand (Young, Reference Young1969; Hayward, Reference Hayward1996). However, Hayward (Reference Hayward1996), based on Young’s (Reference Young1969) report, noted that M. australe had been incorrectly recorded from Sillago ciliata, as no confirmed evidence of infection was found in this host species. Instead, the actual host was likely S. maculata (Hayward, Reference Hayward1996). Monoplectanum youngi has been reported from several sillaginid fish across a wide geographic range. According to Hayward (Reference Hayward1996), this species infects Sillago spp. from coastal and estuarine waters of northern and eastern Australia, including Queensland and the Northern Territory, as well as from the Indo-West Pacific region, encompassing Southeast Asia and parts of the western Pacific. Records from Thailand and Malaysia further support its distribution in tropical coastal waters of Southeast Asia.
In this study, we identified 3 parasite species based on the morphology of the sclerotized male copulatory organ, the haptor, and the number of rows in the squamodisc. These species include M. australe, which infects S. aeolus from both the upper and middle Gulfs of Thailand; M. youngi, which infects S. indica and S. sihama from the upper Gulf of Thailand; and a new species, M. sillaginis sp. nov., which infects S. aeolus, S. indica and S. sihama from the upper Gulf of Thailand. The infective status, morphological description, and distribution of these species are also provided.
Materials and methods
Study areas and fish sample collection
A total of 2096 sillaginid fish were collected from the following 3 regions of Thailand: the middle Gulf of Thailand, the upper Gulf of Thailand, and the Andaman Sea. These fish were purchased from fishermen who were able to record and specify the location of their capture. These included 9 localities: SS1 (13°20′45.0″N 100°12′46.0″E), SS2 (13°18′00.4″N 100°08′41.0″E), SS3 (13°12’09.4”N 100°06’21.7”E) in Samut Sakhon Province; ST1 (9°26’56.8”N 100°32’16.9”E), ST2 (10°03’07.7”N 100°00’03.8”E), and ST3 (9°58’50.9”N 100°13’46.5”E) in Surat Thani Province; and T1 (7°06’41.3”N 99°01’46.9”E), T2 (7°11’06.1”N 98°57’26.7”E), and T3 (7°15’23.4”N 98°58’23.1”E) in Trang Province, collected from May 2024 to January 2025. All fish samples were kept on ice and transferred to the laboratory at the Department of Zoology, Faculty of Science, Kasetsart University. Species identification was based on the morphological traits of their swim bladder, following the FAO Species catalogue Vol. 14, Sillaginid Fishes of the World (McKay, Reference McKay1992).
Examination of monogenean infection
To collect monogenean parasites, the gills of fish samples were dissected and placed in a Petri dish containing distilled water. The monogenean specimens were then carefully removed from the fish’s gill tissue using a needle under an Olympus SZ51 stereomicroscope (Olympus Corporation, Japan). The collected monogeneans were prepared using 2 methods for morphological investigation. They were mounted in ammonium picrate glycerin for semi-permanent slide preparation to study features such as the haptor, male copulatory organ (MCO), and squamodisc for species identification. For deposition at the museum, the monogeneans were also mounted in Canada balsam to prepare permanent slides. The specimens were photographed with Leica DMi8 and Olympus BX53 microscopes (Olympus Corporation, Japan). Drawings were made with the help of a drawing tube attached to an Olympus BX51 compound light microscope (Olympus Corporation, Japan). Morphological characteristics were measured from specimen images following Chaabane et al. (Reference Chaabane, Neifar and Justin2017) and Oliva (Reference Oliver1968) using ImageJ software (Schneider et al., Reference Schneider, Rasband and Eliceiri2012). The measurements are expressed in micrometers (µm) (Table 2). The monogenean was deposited in the Zoological Museum Kasetsart University as the holotype under accession number ZMKU-PM-2101, 2 paratypes under accession numbers ZMKU-PM-2102 and ZMKU-PM-2103 and 1 paratype was deposited in the Lee Kong Chian Natural History Museum (LKCNHM) at National University of Singapore (NUS) under the following accession number: ZRC.PLA.2258. Infection parameters, including prevalence and mean intensity (MI), were calculated following the definitions given by Margolis et al. (Reference Margolis, Esch, Holmes, Kuris and Schad1982) and Bush et al. (Reference Bush, Lafferty, Lotz and Shostak1997).
Results
Infection rate of Monoplectanum spp. in sillaginid fish
All 2096 sillaginid fish were examined and classified into 5 species (S. aeolus, S. asiatica, S. indica, S. ingenuua and S. sihama) (Table 1). Of these, 235 fish belonging to 3 species (Figure 1) of S. aeolus (N = 105, total length [TL]: 11–26.5 cm), S. indica (N = 55, TL: 16–28 cm) and S. sihama (N = 75, TL: 11.7–25 cm) were found to be infected with 3 species of Monoplectanum monogeneans, with an overall prevalence of 11.21% and a mean intensity of 1.72 monogeneans per infected fish. The 3 monogenean species were morphologically identified as M. australe, M. youngi and M. sillaginis sp. nov., the last being described as a new species. Table 1 shows the prevalence and mean intensity of these 3 Monoplectanum species infecting the sillaginid fish. Monoplectanum sillaginis sp. nov. exhibited the broadest host range, infecting 3 fish species (S. aeolus, S. indica and S. sihama), followed by M. youngi, which was found in 2 species (S. indica and S. sihama), and M. australe, found only in S. aeolus. Among the infected sillaginid fish, S. indica showed the highest prevalence of monogenean infection at 29.26%, while S. sihama had the lowest at 9.18%. Regarding the mean intensity of infection, S. sihama displayed the highest at 2.80 (1–7), followed by S. indica and S. aeolus with 1.63 (1–4) and 1.00, respectively. Among the 3 monogenean species examined, M. australe exhibited the highest infective prevalence at 6.79%, while M. youngi was the least prevalent at 3.82%. Concerning the mean intensity, M. youngi exhibited the highest (2.62).
Three sillaginid fish species infected with Monoplectanum spp., (A) S. aeolus, (B) S. indica and (C) S. sihama. Scale bars: (A–C) 1 cm.

Figure 1 Long description
The image shows three sillaginid fish species in lateral view, labeled A, B and C. Each fish is displayed horizontally with visible fins and scales. The fish labeled A has a streamlined body with distinct dorsal and pectoral fins. The fish labeled B appears slightly elongated with visible dorsal and anal fins. The fish labeled C has a prominent dorsal fin and a tapered tail. Scale bars are present beneath each fish, indicating size reference.
Prevalence and mean intensity of the Monoplectanum spp. Infections in sillaginid fish

Table 1 Long description
The table measures the prevalence and mean intensity of Monoplectanum spp. infections in various sillaginid fish species. S. indica has the highest prevalence at 29.26%, with a mean intensity of 1.63, indicating frequent infections. S. aeolus and S. sihama also show notable infection rates, with prevalences of 10.46% and 9.18%, respectively. S. asistica and S. ingenuua have no recorded infections, suggesting possible resistance or absence of the parasite. The data highlights that M. australe primarily infects S. aeolus, while M. youngi and M. sillaginis sp. nov. are more prevalent in S. indica and S. sihama. Overall, the total prevalence across all species is 11.21%, with varying mean intensities, indicating differences in infection severity among species.
Infection summary and morphological description of Monoplectanum spp
Monoplectanum australe Young 1969 (Figure 2)
Morphological characters of Monoplectanum australe under light microscope. (A) whole body; (B) MCO with posterior prostatic reservoir with fork-shaped; (C) rows of minute rodlets of squamodisc; (D) haptor with hamuli, bar and marginal hooks. Abbreviations as follows: DH: dorsal hamuli; DLB: dorsal (lateral) bar; E: egg; ES: eye spot; H: haptor; HG: Head organ; MCO: male copulatory organ; MH: marginal hook; P: pharynx; PPS: posterior prostatic reservoir; VB: ventral bar; VH: ventral hamuli. Scale bars: (A) 500 µm, (B–C) 10 µm and (D) 50 µm.

Figure 2 Long description
Image A shows the whole body of Monoplectanum australe with labeled parts: eye spot, pharynx, male copulatory organ and haptor. Image B displays the male copulatory organ with a fork-shaped posterior prostatic reservoir. Image C presents rows of minute rodlets of the squamodisc. Image D illustrates the haptor with hamuli, bar and marginal hooks, including dorsal hamuli, ventral bar, ventral hamuli and marginal hook. Scale bars are visible in each image, indicating magnification levels.
Type host: Sillago ciliata Cuvier, 1829
Present Host: Sillago aeolus Jordan & Evermann, 1902 (Figure 1A)
Locality: the upper Gulf of Thailand (13°20’45.0”N 100°12′46.0″E)
Site of infection: Gill filaments
Other records: S. ciliata, S. aeolus, S. maculata and S. burrus from Moreton Bay, Shark Bay, Point Samson, Duyfken Point, Australia and the upper Gulf of Thailand, (Young, Reference Young1969; Hayward, Reference Hayward1996; Niyom and Purivirojkul, Reference Niyom and Purivirojkul2020)
The morphological description
The morphological features of the M. australe (Table 2) are based on 10 specimens. The body elongate and slender, 2 pairs of lateral head organs connected to cephalic glands on each side of the pharynx. Two pairs of eye spots: anterior pair smaller than posterior pair. Pharynx spherical. Intestine bifurcates immediately posterior to the pharynx; the caeca and vitellarium terminate well anterior to squamodiscs. Haptor consists of 2 pairs of lateral hamuli, comprised of ventral and dorsal hamuli, 3 bars-1 ventral bar and 2 dorsal (lateral) bars and small squamodisc, along with 14 marginal hooks of the same morphotype.
Measurement characteristics of 3 species of Monoplectanum from the current study and the original description. All measurements are given in micrometers (µm) and abbreviations are as follows: L: Length and W: Width

Table 2 Long description
The table compares measurement characteristics of three Monoplectanum species across different studies and regions, focusing on body and haptor dimensions, hamulus lengths, and other anatomical features. M. australe from the present study has the largest body length and width compared to previous studies by Young and Hayward. M. youngi shows a wide range of body sizes, with the present study reporting larger dimensions than Hayward's 1996 study. M. sillaginis, a new species, has measurements similar to M. youngi. The dorsal and ventral hamulus lengths are consistent across species, with slight variations. Fish hosts and study locations vary, with M. australe and M. sillaginis found in Thailand, while M. youngi is distributed across multiple countries. These variations highlight the influence of geographical and environmental factors on species morphology.
Body elongates, total length (Figure 2A), measuring 1920–2108 (2089) × 341–434 (393). Haptor (Figure 2A, 2B) measures 123–144 (138) × 125–152 (146). Ventral hamuli (Figure 2A, D) have 2 distinct guards, and the tip is recurved, with outer length 40–48 (43) and inner length 12–19 (15). Dorsal hamuli (Figure 2A, D) have an indistinct guard, and the tip is recurved, with outer length 43–57 (49) and inner length 18–26 (22). The single ventral bar bends in a similar V-shape (Figure 2A, D), measuring 25–49 (36) × 6–9 (7). Two dorsal (lateral) bars (Figure 2A, D) are similar in shape, straight, with the initial part flattened, medial end, and articulate laterally with the hamuli (Figure 2A, D), measuring 20–34 (26) × 10–17 (14). Squamodisc consists of rows of minute rodlets, inner row forming closed oral structure (Figure 2C), measuring 25–31 (29) × 26–29 (27), with 14–16 (15) rows connected.
Male copulatory organ (MCO) consists of tube and posterior prostatic reservoir (Figures 2B, 6A). MCO’s tube curved and consists of 2 nested tubes: outer tube surrounding thin-walled inner tube that recurves and thickened proximally, gradually narrowing towards distal end to 60–70 (65) × 4–6 (5). Terminal part of posterior prostatic reservoir fork-shaped (Figures 2B, 6A), measuring 24–39 (30), and posterior prostatic reservoir located at end of tube. Vagina not observed. Vitelline follicles form numerous transversely elongated lobes. Arranged longitudinally along lateral fields of the body, from posterior margin of pharynx to well before position of squamodisc (Figure 2A).
Remarks: The species was originally described as M. australis by Young (Reference Young1969), however, Hayward (Reference Hayward1996) later referred to it as M. australe, which is most likely an erroneous spelling. The specimens examined in this study correspond morphologically to the original description of M. australe, particularly in the shape of MCO and haptor. However, this species is very similar to M. youngi, such as the number of squamodisc rows, it can be distinguished by differences especially in the shape of MCO and the morphology of the dorsal (lateral) bar.
Morphological characters of Monoplectanum youngi under light microscope. (A) whole body; (B) MCO with posterior prostatic reservoir with wrench-shaped; (C) position of squamodisc; (D) rows of minute rodlets of squamodisc; (E) haptor with hamuli, bar and marginal hooks. Abbreviations used: DH: dorsal hamuli; DLB: dorsal (lateral) bar; E: egg; ES: eye spot; H: haptor; HG: Head organ; MCO: male copulatory organ; MH; marginal hook; P: pharynx; PPS: posterior prostatic reservoir; SQ: squamodisc; VB: ventral bar; VH: ventral hamuli. Scale bars: (A) 500 µm; (B–C, E) 50 µm and (D) 10 µm.

Figure 3 Long description
Image A shows the whole body of Monoplectanum youngi with labeled parts: eye spot, head organ, male copulatory organ, egg, pharynx, haptor and squamodisc. Image B displays the male copulatory organ with a posterior prostatic reservoir, wrench-shaped. Image C illustrates the position of the squamodisc. Image D shows rows of minute rodlets of the squamodisc. Image E depicts the haptor with dorsal hamuli, ventral hamuli, dorsal lateral bar, ventral bar and marginal hooks. Scale bars are visible in each image, indicating magnification levels.
Monoplectanum youngi Hayward 1996 (Figure 3)
Type hosts: Sillago analis Whitley, 1943
Present host: Sillago indica McKay, Dutt & Sujatha, 1985 and S. sihama (Forsskål, 1775) (Figure 1B, C)
Locality: The upper Gulf of Thailand (13°20′45.0″N 100°12’46.0”E) and the middle Gulf of Thailand (9°26’56.8”N 100°32’16.9”E)
Site of infection: Gill Filaments
Other records: Gills of S. analis (type host), S. attenuata, S. ciliata, S. lutea, S. parvisquamis and S. sihama from Burdekin River, Point Samson, Princess Charlotte Bay, Torres Strait, Queensland; Moreton Bay, Charles Point, Darwin and Duyfken Point, Australia; Upper Bay Zaal Island, Saudi Arabia; Wan Chai Market, Sai Kung, Hong Kong; Phuket, Bang Saen, Thailand; Kuala Terengganu; and Kuala Lumpur Market, Malaysia (Hayward, Reference Hayward1996).
The morphological description
The morphological features of the M. youngi (Figure 3) are based on 10 specimens. The body elongate and slender, 2 pairs of lateral head organs are connected to cephalic glands on each side of the pharynx. Two pairs of eyespots are present: normally, the anterior pair is smaller than the posterior pair. Pharynx spherical. Intestine bifurcates immediately posterior to pharynx; the caeca and vitellarium terminate well anterior to squamodiscs. Haptor consists of 2 pairs of lateral hamuli, comprising ventral and dorsal hamuli, 3 bars, including 1 ventral bar and 2 dorsal (lateral) bars, and small single squamodisc. There are 14 marginal hooks, all the same morphotype.
Body elongated and slender (Figure 3A), total length, 1543–2802 (1917) × 227–384 (311). Haptor (Figure 3H) measures 105–157 (131) × 110–207 (150). Ventral hamuli (Figure 3E) have 2 distinct guards, and the tip is recurved, with an outer length of 36–49 (42) and an inner length of 12–19 (15). The dorsal hamuli (Figure 3E) have an indistinct guard, and the tip is recurved, with an outer length of 45–67 (49) and an inner length of 20–32 (23). The single ventral bar bends in a similar V-shape (Figure 3H), measuring 27–45 (34) × 5–9 (7). Two dorsal (lateral) bars (Figure 3E) are similar in shape, not overlapping; they are straight and curve at the initial part, expanding wider than the lateral and terminal parts, and articulate with the hamuli laterally. Squamodisc consists of rows of minute rodlets, inner row forming closed oral (Figure 3C, D), measuring 27–30 (29) × 27–33 (29), with 14–15 (15) rows connected.
MCO comprises tube and posterior prostatic reservoir (Figure 3B, 6B). The tube of the MCO curved and consists of 2 nested tubes: outer tube enclosing thin-walled inner tube that recurves, thickened proximally, and gradually narrows along its length towards distal end 57–69 (65) × 4–5 (4). Terminal part of the posterior prostatic reservoir wrench shape (Figures 3B, 6B), 27–41 (33), and the position of the posterior prostatic reservoir is found at the end of the tube. Vagina not observed. Vitelline follicles form numerous transversely elongated lobes. Arranged longitudinally along lateral fields of the body, extending from posterior margin of pharynx to well before position of squamodisc (Figure 3A).
Remarks: This species appears to be very closely related to M. australe; however, the terminal part of the posterior prostatic reservoir is wrench-shaped, unlike the fork-shaped posterior prostatic reservoir of M. australe, and the shape of the dorsal (lateral) bar also differs. It also occurs on various hosts and has a relatively larger distribution than M. australe.
Monoplectanum sillaginis sp. nov. (Figures 4, 5)
The morphology of Monoplectanum sillaginis sp. nov. under light microscope. (A) whole body; (B–E) MCO with different views of posterior prostatic reservoir with shuttle-shaped; (F) haptor with hamuli, bar, marginal hooks and squamodisc. Abbreviations used: DH: dorsal hamuli; DLB: dorsal (lateral) bar; E: egg; ES: eye spot; H: haptor; HG: Head organ; MCO: male copulatory organ; MH: marginal hook; P: pharynx; PPS: posterior prostatic reservoir; SQ: squamodisc; VB: ventral bar; VH: ventral hamuli. Scale bars: (A) 500 µm, (B) 50 µm, (C–E) 20 µm and (F) 50 µm.

Figure 4 Long description
The image A shows the whole body of Monoplectanum sillaginis sp. nov. labeled with HG subscript a, HG, P, MCO, E and SQ. The image B to E show different views of the posterior prostatic reservoir labeled as PPS. The image F shows the haptor with various structures labeled as MH, VH, DH, DLB, VB and SQ. The haptor includes hamuli, bar, marginal hooks and squamodisc. Scale bars are visible in each image, indicating the magnification level.
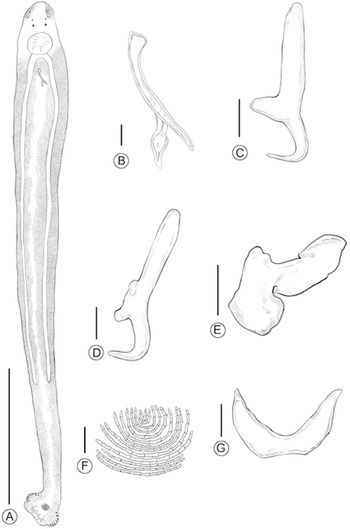
Schematic drawings of Monoplectanum sillaginis sp. nov. (A) whole body; (B) MCO; (C) ventral hamuli; (D) dorsal hamuli; (E) dorsal (lateral) bar; (F) squamodisc; (G) ventral bar. Scale bars: (A) 500 µm and (B–G) 10 µm.

Figure 5 Long description
The image shows schematic drawings of Monoplectanum sillaginis sp. nov. A depicts the whole body with a scale bar of 500 micrometers. B shows the MCO with a scale bar of 10 micrometers. C illustrates the ventral hamuli, also with a 10 micrometer scale bar. D presents the dorsal hamuli, accompanied by a 10 micrometer scale bar. E displays the dorsal (lateral) bar with a 10 micrometer scale bar. F features the squamodisc and G shows the ventral bar, both with 10 micrometer scale bars.
Type host: Sillago sihama (Fabricius, 1775) (Figure 1C)
Other hosts: Sillago aeolus Jordan & Evermann, 1902 and S. indica McKay, Dutt & Sujatha, 1985
Type locality: Samut Sakhon Province, the upper Gulf of Thailand (13°20′45.0″N 100°12’46.0”E)
Type specimens: Holotype, ZMKU-PM-2101; 3 paratypes, ZMKU-PM-2102 and 2103 and ZRC.PLA.2258
Species LSID: urn:lsid:zoobank.org:act:47CA1C77-C639-470E-BB2C-F6C8393EA0C0
Site of infection: Gill filaments
The morphological description
The morphological features based on 10 specimens. Slender and elongated body with 2 pairs of lateral head organs connected to cephalic glands on each side of pharynx. There are 2 pairs of eyespots: anterior pair smaller than the posterior pair. Pharynx spherical. Intestine bifurcates immediately posterior to the pharynx; caeca and vitellarium terminate well before the squamodiscs. Haptor consists of 2 pairs of lateral hamuli, including ventral and dorsal hamuli; 3 bars 1 ventral bar and2 dorsal (lateral) bars; small squamodisc; and 14 marginal hooks of the same morphotype.
Body elongated and slender (Figures 4A, 5A), total length 1511–2101 (1922) × 200–431 (299). Haptor (Figure 4A, F) measures 118–145 (133) × 128–207 (168). Ventral hamuli (Figures 4F, 5C) have 2 distinct guards, with the tip recurved; outer length is 38–47 (42), and inner length is 13–20 (15). Dorsal hamuli (Figures 4F, 5D) have an indistinct guard, with the tip recurved; outer length is 43–57 (49), and inner length is 20–28 (23). The single ventral bar is bent into a V-shape (Figures 4F, 5G), measuring 29–46 (34) × 5–8 (6). Two dorsal (lateral) bars (Figures 4F, 5E) are similar in shape, not overlapping; they are straight and curve at the initial part, widening more than the lateral and terminal parts, and articulate with the hamuli laterally. The squamodisc consists of rows of minute rodlets and an inner row forming a closed oral structure (Figures 4A, F, 5F), measuring 27–33 (28) × 26–33 (28), with 12–14 (13) rows connected.
MCO comprises the tube and posterior prostatic reservoir (Figures 4B–E, 5B, 6A–E). Tube of the MCO is curved and consists of two nested tubes: an outer tube encases a thin-walled inner tube that is recurved and thickened proximally, gradually narrowing along its length toward the distal end 60–72 (66) × 4–6 (5). Terminal part of posterior prostatic reservoir shuttle-shaped (Figures 4B–E, 5B, 6C), measuring 23–29 (26) × 7–10 (8). Posterior prostatic reservoir positioned two-thirds along the length of the tube. Vagina not observed. Vitelline follicles form numerous transversely elongated lobes. Arranged longitudinally along lateral fields of the body from posterior margin of pharynx to just before squamodisc (Figure 4A, 5A).
Differential diagnosis: Monoplectanum sillaginis sp. nov. is clearly distinguished from other Monoplectanum species by the morphology of the MCO and the number of rodlet rows in the squamodisc. In M. sillaginis sp. nov., the terminal part of the posterior prostatic reservoir of the MCO is shuttle-shaped, and the squamodisc comprises 12–13 (13) rows of rodlets. In contrast, M. australe possesses a fork-shaped terminal part of posterior prostatic reservoir and squamodisc with 14–16 (15) rows, whereas M. youngi is characterized by a wrench-shaped terminal part of posterior prostatic reservoir and squamodisc bearing 14–15 (15) rows.
Discussion
Sillaginid fish are widely distributed in the Indo-Pacific region, including Thailand, especially the Andaman Sea and the Gulf of Thailand (McKay, Reference McKay1992; Froese and Pauly, Reference Froese and Pauly2025). However, the investigation of their parasitic monogenean infections in Thailand is still limited. Among 5 sillaginid fish species collected in the current study, 3 of them were observed to harbour Monoplectanum spp. Among 3 species of monogeneans currently observed, M. sillaginis sp. nov. shows distinct morphological differences from other species in the genus Monoplectanum, especially in the structure of the MCO and the number of rodlet rows in the squamodisc.
The general morphology of the 2 Monoplectanum species, M. australe and M. youngi, as observed in our study, agrees with the original description by Hayward (Reference Hayward1996) regarding body shape, organ appearance, and the position and arrangement of organs. However, the measurements of M. australe in our study showed that at least 2 organs, the ventral hamulus and squamodisc, were smaller than those reported in the original description (Table 2). For M. youngi, an organ of the ventral hamulus was found to be smaller than previously reported (Table 2). These differences in morphological features based on measurements may be related to specimen preservation and preparation processes, as well as the measurement methods used (Justine, Reference Justine2005; Fankoua et al., Reference Fankoua, Nyom, Bahanak, Bilong Bilong and Pariselle2017).
Our results show the prevalence and mean intensity of infection by 3 Monoplectanum species in 3 sillaginid fishes (Table 1). The overall prevalence and mean intensity of the infection were 11.21% and 1.72 monogeneans per infected fish, respectively. Monoplectanum australe was found only in S. aeolus, with a prevalence and mean intensity of 6.97% and 1, respectively, meanwhile no infection was found in the remaining fish species. Compared to a previous study (Niyom and Purivirojkul, Reference Niyom and Purivirojkul2020) that they reported M. australe infecting S. aeolus fish with 2.82% and 3 of prevalence and mean intensity of the infection, the present study shows a higher prevalence and lower mean intensity that reported M. australe in 2 fish species, including S. maculata with high infection (20%, 2) in comparison to S. aeolus with a low infection (2.82%, 3), the present study shows a higher prevalence and lower mean intensity. Variations in sampling years, environmental conditions, or host-parasite interactions may cause this difference. Infection of the M. sillaginis sp. nov. observed in 3 fish species might suggest high infectability of them compared to the remaining examined monogeneans. Moreover, infection of different hosts can reduce the risk of extinction due to fluctuations in host abundance (Tavares-Dias et al., Reference Tavares-Dias, Silva and Oliveira2022). However, some previous studies did not provide detailed data about the infection rates, making it to be difficult to compare the infective prevalence and mean intensity with earlier findings.
Distribution of the Monoplectanum has been primarily reported in the Oriental and Australian faunal regions (Figure 7), from 1969 to the presence (Young, Reference Young1969; Hayward, Reference Hayward1996; Niyom and Purivirojkul, Reference Niyom and Purivirojkul2020). Species of Monoplectanum are distributed according to the dispersion of sillaginid fish, with several species serving as hosts for Monoplectanum spp. (Hayward, Reference Hayward1996; Niyom and Purivirojkul, Reference Niyom and Purivirojkul2020). Different species of Monoplectanum have varying host ranges, despite being distributed in the same region, especially in Australia. Between the 2 species, the distribution range of M. youngi is broader than that of M. australe. Monoplectanum youngi has been recorded in several areas of Australia, Malaysia, Thailand, Hong Kong, and even Saudi Arabia, while M. australe is known to be distributed in Australia and Thailand. The difference in the distribution ranges of these 2 monogeneans may be due to the taxon spectrum and distribution ability of their fish hosts. Seven species of the following sillaginid fish have been documented as hosts of M. youngi: S. analis, S. attenuata, S. ciliata, S. indica, S. lutea, S. parvisquamis and S. sihama, while 4 sillaginid species (S. aeolus, S. burru, S. ciliata and S. maculata) have harboured M. australe. This broader spectrum of host species undoubtedly contributes to the enhancement of parasite distribution. One of these sillaginid species, S. sihama (host of M. youngi), exhibits a distribution range throughout the Indo-West Pacific region (from South Africa to Japan) (McKay, Reference McKay1992; Froese and Pauly, Reference Froese and Pauly2025), demonstrating that its wider distribution could also expand the range of M. youngi. Furthermore, the distribution ability of M. youngi may relate to its size, particularly its body size. Compared to M. australe, M. youngi has a smaller body size, based on both this study’s comparative data and previous research. The smaller size of the monogenean may reduce the chance of dislodging from the gill filament in water currents, leading to a broader distribution following its fish host. Regarding the new species, M. sillaginis sp. nov., it is currently reported to be distributed from the upper Gulf of Thailand and infects 3 sillaginid species, 2 of which (S. aeolus and S. sihama) have wider distributions across the oriental and Australian regions (McKay, Reference McKay1992; Froese and Pauly, Reference Froese and Pauly2025). Consequently, the distribution of M. sillaginis sp. nov. could potentially expand similarly to M. youngi due to their shared fish hosts (S. sihama and S. indica), especially S. sihama, which is distributed throughout the Indo-West Pacific region (McKay, Reference McKay1992; Froese and Pauly, Reference Froese and Pauly2025). Focusing on Thailand, the upper Gulf region currently shows the distribution of all 3 Monoplectanum species. Meanwhile, the middle Gulf of Thailand, also documented for M. youngi in this study.
Schematic drawings of MCO differences of 3 Monoplectanum species, (A) M. australe (fork-shaped); (B) M. youngi (wrench-shaped); (C) M. sillaginis sp. nov. (shuttle-shaped). Scale bar: 20 µm.

Figure 6 Long description
The image shows three schematic drawings labeled A, B and C, depicting male copulatory organs with different shapes. Drawing A illustrates a fork-shaped organ, characterized by a bifurcated structure at the base. Drawing B presents a wrench-shaped organ, featuring a curved design with a prominent hook-like element. Drawing C shows a shuttle-shaped organ, distinguished by a streamlined form with a rounded end. A scale bar is visible next to the drawings, indicating size reference.
Distribution map of Monoplectanum spp. reported by previous and current studies. Different colours indicate different Monoplectanum species, and different symbol shapes denote different sources of study. This map was created using QGIS version 3.34 based on the following coordinate reference system: ESPG:32647–WGS48/UTM Zone 47N (QGIS Development Team, 2021).

Figure 7 Long description
The map illustrates the distribution of Monoplectanum species across various regions, including Australia, Thailand, Malaysia and Saudi Arabia. Different symbols represent sources of study: triangles for Young (1969), squares for Hayward (1996), diamonds for Niyom and Purivirojkul (2020) and circles for the present study. The species M. australe, M. youngi and M. sillaginis are marked with different colors. An inset map highlights the upper Gulf of Thailand, showing detailed distribution points. The map uses the coordinate reference system ESPG:32647–WGS48/UTM Zone 47N.
In conclusion, our investigation identified 3 species of Monoplectanum from the upper and the middle Gulf of Thailand, including M. australe, M. youngi and M. sillaginis sp. nov., the latter of which was described as a new species. Key characteristics distinguishing these 3 species were clearly defined and demonstrated. The infection data revealed that M. sillaginis sp. nov. was the most prevalent species infecting sillaginid fish. Additionally, the study provided information on the distribution of monogeneans. The data obtained may contribute to a better understanding of the biodiversity of monogeneans in marine ecosystems. While this study primarily focused on morphological characteristics, future research should consider incorporating ecological approaches and molecular character to clarify infection patterns or dynamics of the monogeneans and their fish host related to environmental factors.
Acknowledgements
The authors would like to thank the Department of Zoology, Faculty of Science, Kasetsart University, for their facility and equipment support. We also thank Miss Orathai Uoolek for her assistance with the specimen drawings.
Author contributions
Conceptualization: NN and WP; Data Curation: NN; Methodology and Investigation: NN; Analysis: NN and PW; Resources: NN and WP; Visualization: NN and PW; Writing – Original Draft Preparation: NN and PW; Writing – Review & Editing: PW and WP; Supervision: WP. All authors have read and agreed to the manuscript that has been submitted.
Financial support
This research received no specific grant from any funding agency, commercial or not-for-profit sectors.
Competing interests
The authors declare there are no conflicts of interest.
Ethical standards
This study was approved by the ethics committee of Kasetsart University (approval no.: ACKU61-SCI-019).









 Open access
Open access