


Introduction
Nonmarine molluscs are highly imperilled (Lydeard et al., Reference Lydeard, Cowie, Ponder, Bogan, Bouchet and Clark2004) and have been subject to the highest number of documented extinctions in modern historical times for any major group of organisms (IUCN, 2024). The actual number of extinctions is uncertain because of a lack of data for many species of molluscs, and the IUCN figures may be underestimates (Régnier et al., Reference Régnier, Fontaine and Bouchet2009; Cowie et al., Reference Cowie, Rėgnier, Fontaine and Bouchet2017). Land snail faunas on oceanic islands are particularly vulnerable to various threats, including introduced predators and habitat destruction; this is exemplified on Pacific islands by the well-documented plight of Partulidae in Polynesia and Achatinellinae in Hawaii (Lydeard et al., Reference Lydeard, Cowie, Ponder, Bogan, Bouchet and Clark2004; Régnier et al., Reference Régnier, Fontaine and Bouchet2009). Cowie (Reference Cowie2002) drew attention to the decline of island endemics and replacement of native land snail faunas by introduced species transported widely by people. Endemic land snails of the genus Poecilozonites on the isolated archipelago of Bermuda in the North Atlantic are similarly threatened. The genus has a long evolutionary history (Gould, Reference Gould1969, Reference Gould1993), with at least 12 species known from the fossil record (Hearty & Olson, Reference Hearty and Olson2010). Two species, P. bermudensis and P. circumfirmatus, survived to the mid 20th century (Bieler & Slapcinsky, Reference Bieler and Slapcinsky2000), but both have been in decline since the 1960s (Gould, Reference Gould1993; review in Outerbridge & Sarkis, Reference Outerbridge and Sarkis2018). Once widespread and abundant, P. bermudensis (Plate 1) was thought to be extinct by the early 1990s, until the discovery of an isolated remnant population in an alley in the city of Hamilton in 2014 and subsequently on Port’s Island in 2017 (Outerbridge & Sarkis, Reference Outerbridge and Sarkis2018). The initial decline coincided with the introduction of predatory snails as part of pest management efforts intended to control introduced snails in agricultural crops, including Euglandina rosea, which has caused the demise of native land snail populations on islands in many parts of the world (Gerlach et al., Reference Gerlach, Barker, Bick, Bouchet and Brodie2021). The introduction of snail-eating flatworms (Platyhelmintes, including Bipalium vagum), rodents such as rats Rattus spp., and free-ranging chickens Gallus gallus domesticus, together with habitat destruction, are thought to have contributed to further declines and to have impeded recovery (Outerbridge & Sarkis, Reference Outerbridge and Sarkis2018). Both extant Poecilozonites species are categorized as Critically Endangered on the IUCN Red List (Ovaska & Outerbridge, Reference Ovaska and Outerbridge2019a,Reference Ovaska and Outerbridgeb).
An adult Poecilozonites bermudensis snail.

Here we report on recovery efforts to secure viable populations of P. bermudensis through conservation interventions, using snails from a remnant wild population and reintroductions from an ex situ colony to several localities within the species’ indigenous range in Bermuda (i.e. reintroduction sensu IUCN/SSC, 2013). The main objective was to contribute to the long-term aim of establishing self-sustaining populations at a minimum of 10 sites within the Bermuda archipelago, as specified in the recovery plan for the species (Outerbridge & Sarkis, Reference Outerbridge and Sarkis2018). An additional objective was to document the successes and failures at different localities and their possible causes, to aid in further conservation efforts for this and other species. We carried out the reintroductions within the context of broader ecosystem restoration, focusing primarily on nature reserves. Our approach was two-fold: (1) intensive monitoring of releases at one site, Nonsuch Island, to document aspects of ecology, distribution and population trends of the snails over time, and (2) releases of snails to sites deemed suitable based on our initial results from Nonsuch Island and also from Port’s Island (Copeland & Hesselberg, Reference Copeland and Hesselberg2021). The successful ex situ conservation programme at Chester Zoo, UK, initiated in 2017 from snails originating from the Hamilton alley, allowed us to release > 100,000 snails at 27 localities, including 11 small isolated offshore islands and 16 main island sites connected to each other by a series of bridges.
Methods
Intensive monitoring on Nonsuch Island
We carried out intensive monitoring of P. bermudensis in the 16.3 ha Nonsuch Island Nature Reserve, where ecosystem restoration for the conservation of rare and threatened species and habitats has been in progress since 1962 (Wingate, Reference Wingate and Woodwell1990; Government of Bermuda, Department of Environment and Natural Resources, undated). This island was selected because of the absence of predatory snails, ongoing rat eradication, and presence of restored, relatively intact natural forest habitat. We released P. bermudensis at three sites on the island: (1) old moss-covered limestone wall in mainly dense palmetto Sabal bermudana-dominated shrub forest on east side of the island (Wall), (2) palmetto-dominated forest with patches of olivewood Elaeodendron laneanum saplings and young trees c. 70 m west of the Wall site (Grid), and (3) an old quarry in shrub forest including palmettos c. 50 m north-west of the Grid (Quarry). All were hard releases; i.e. snails were released directly into the environment in small groups, with the groups close to each other. The first release took place at the Wall site in 2016, followed by those at the Grid and Quarry sites in 2017 and 2019, respectively (Table 1). All released snails originated from the Hamilton population either through direct translocation (Wall) or through ex situ breeding (Grid and Quarry). We periodically augmented snails at the Grid with additional releases from the colony at Chester Zoo, which first became available in 2019 (Table 1).
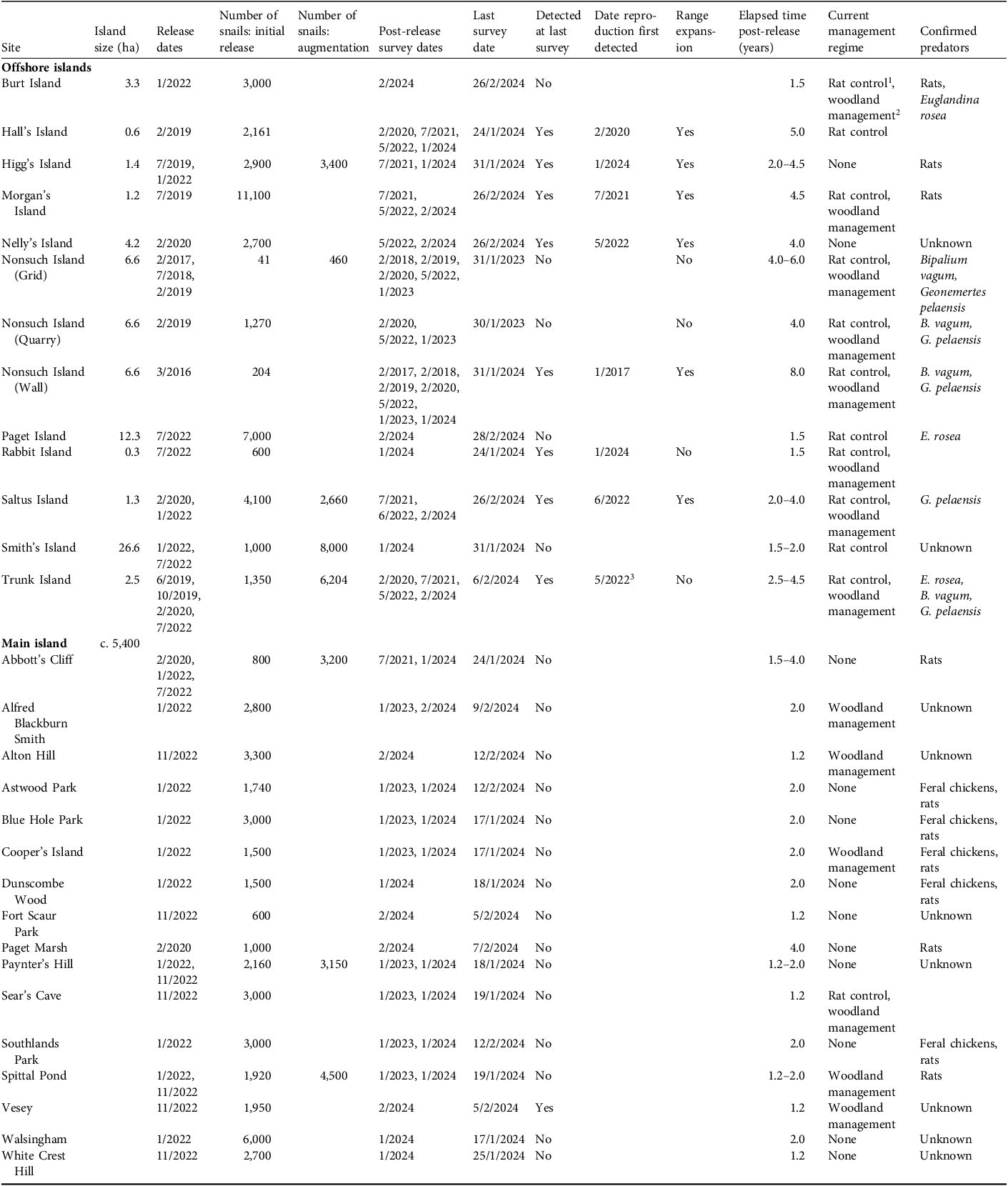
Summary of releases of Poecilozonites bermudensis to offshore and main island sites (Fig. 1) from 2016 to 2022, and results of follow-up monitoring. All released snails originated from a remnant population discovered in Hamilton, Bermuda, in 2014, either through translocations (Nonsuch Island) or since 2019 from ex-situ breeding at Chester Zoo (other sites).

Table 1 Long description
The table presents data on the release and monitoring of Poecilozonites bermudensis snails across different sites. It includes details such as island size, release dates, number of snails, post-release augmentation, survey dates, detection status, range expansion, elapsed time, current management regime, and confirmed predators. The table is organized into columns for each of these variables and rows for each site, providing a comprehensive overview of the snail releases and their outcomes.
1 Control activities include using rodenticides and trapping.
2 Management activities include removing invasive flora and encouraging the growth of native and endemic species.
3 Egg-laying and a hatchling were detected, but no juveniles were found during subsequent surveys.
We conducted surveys at the release sites annually, from 2017 to 2023 (with the exception of 2021), in survey sessions lasting 1–3 weeks. Most surveys took place at night when the snails are active on the surface. In 2017–2020, we marked subadult and adult snails (all with shell width ≥ 15 mm at widest point) for individual recognition with alphanumeric fluorescent micro-tags (alpha tags; Northwest Marine Technology, USA) glued to the shell (Flewitt et al., Reference Flewitt, Williams, Preziosi and Garcia2023) and/or with passive integrated transponders (PIT-tags) inserted into the umbilical cavity and glued in place (Ovaska et al., Reference Ovaska, Outerbridge and Sopuck2025), to obtain information on survival and activity. We estimated population size from mark–recapture data in two 2 × 10 m plots, each surveyed three times 2 days apart, from 27 January to 1 February 2023. One plot was adjacent to the stone wall and included the release site; the other was c. 30 m from the wall, in the shrub forest. In these plots we marked all snails with shell width > 5 mm using ink paint pens (Pilot Super Color, extra fine, permanent ink), using a different colour dot for each session, and calculated population estimates using the Jolly–Seber method (Krebs, Reference Krebs1999). We measured shell width of all snails periodically in portions of the study areas to assess size-frequency distribution.
Fanning outward from the release sites, we carried out surveys for active snails at night and searched refuges, such as fallen palmetto fronds and under rocks, during daylight to obtain information on the area the population was occupying. We measured locations of snails from fixed points within the study sites and converted these to coordinates for area and distance calculations, carried out with QGIS 3.16 (QGIS, 2024). We calculated the area of occupancy of the population at each release site using the minimum convex polygon method, joining the outermost observation points (Moreira et al., Reference Moreira, Calado and Dias2015).
Releases across the archipelago
We selected release sites from a list of locations deemed suitable, focusing on government-owned and private nature reserves, each with varying degrees of habitat management and invasive species control. The offshore islands were surveyed for the presence of predators before any snails were released. The 11 offshore islands ranged from 0.3 ha (Rabbit Island) to 26.6 ha (Smith’s Island); straight line distances to the nearest main island shore ranged from 37 m to 1,575 m (mean 350 m). The 16 main island sites were all located within nature reserves (Fig. 1). Bermuda’s nature reserves provide habitat for a number of the islands’ most threatened species (Government of Bermuda, undated). Introduced predators, including rats and feral domestic chickens, occurred or were suspected to occur at all main island sites but varied in species composition and/or apparent abundance on the offshore islands. As on Nonsuch Island, we released groups of snails directly into the environment without an acclimation period at the site. We selected habitats at the release sites based on observations made by Copeland & Hesselberg (Reference Copeland and Hesselberg2021), including palmetto forest with cracked limestone formations and other structural features that would provide refuges. We released snails during prolonged wet periods and attempted to time the releases after rainfall when the ground was moist. Releases elsewhere than on Nonsuch Island were from 2019 to 2022 with post-monitoring surveys from 2020 to 2024 (Table 1). All released snails originated from the ex-situ breeding colony at Chester Zoo (see Jameson et al., Reference Jameson, Flewitt, Papp, Prince, Richardson and Garcia2020, for details of rearing).
Locations in Bermuda where Poecilozonites bermudensis were reintroduced, with outcomes as documented during the last monitoring surveys in January–February 2024.

Follow-up monitoring consisted of observers manually searching the litter layer, fallen palmetto fronds, rocks and other cover objects for snails. We collected the first snails encountered until a quota of c. 100 was reached, or no more individuals were found during a search time of 1 h, for measurements of shell width, after which we returned them to their approximate locations. We visually estimated the area where the snails were found and the distance of the farthest observations from the release point. We visited all sites at least once, ≥ 1 year after the release, with the last visit in January–February 2024 (Table 1). The number of surveys per site was 1–4 (excluding Nonsuch Island, where surveys were more frequent), depending on logistics. We evaluated success based on persistence for at least 1 year, evidence of reproduction, juveniles of different size classes present, and expansion of the area of occupancy from the release site.
Results
Intensive monitoring on Nonsuch Island
The reintroduction at the Wall site resulted in the establishment of a self-sustaining population of P. bermudensis. During the first post-release surveys in February 2017, 11 months after the release, we detected hatchling-sized individuals (shell width < 5 mm), indicating reproduction, and all size classes were present in subsequent years. In 2017, adults and subadults (shell width ≥ 15 mm), probably from the original release, formed 63.3% of 158 detections (excluding hatchlings, which were not surveyed quantitatively). The percent of juveniles (shell width 5.0–14.9 mm) changed from 36.7% in 2017 (158) to 92.2% in 2020 (272), 70.4% in 2022 (1,024), and 67.8% in 2023 (883). Large individuals (shell width 20.0–25.0 mm) comprised 27.1% of the sample in 2017 but were rare or absent in subsequent counts (1.5% in 2020, 0.5% in 2022, 0% in 2023).
In 2023, mean population estimates for snails with shell width > 5 mm were 1,870 (95% CI 1,110–4,159) on the plot along the Wall site and 2,639 (95% CI 1,652–5,408) on the plot c. 30 m away. The corresponding means for snails with shell width ≥ 15 mm were 773 (95% CI 395–2,179) and 883 (95% CI 357–3,586). Mean density estimates (snails/m2) derived from these values were 94–132 for all snails and 39–44 for subadults and adults on the two plots, respectively.
The area of occupancy, calculated as the minimum convex polygon around the outermost observation points, increased from the release in 2016 to 2022, with a major expansion from 2020 to 2022 (Fig. 2). By May 2022, the population was distributed over an area of 6,849 m2 and up to 92 m from the release site. We did not observe a notable expansion in 2023 but did not carry out a comprehensive survey in that year.
Expansion of the area of occupancy of P. bermudensis at the Wall site on Nonsuch Island (Fig. 1) from reintroduction in March 2016 to May 2022. Polygons represent the area occupied by the population in each survey year.

In contrast to the Wall site, P. bermudensis failed to establish viable populations at the Grid and Quarry release sites. At both sites individual snails persisted in diminishing numbers for 1–3 years, but we detected no evidence of reproduction at either site. At the Grid site, we detected the last live snails (six individuals, shell width 19.8–22.1 mm) in May 2022, 3.1 years after the last augmentation; surveys in 2023 failed to find live snails. Recaptures of 23 tagged snails caught a year apart showed rapid but variable growth in shell width especially for smaller individuals, but we detected no hatchlings or small juveniles. All detections were < 8 m from the release site. At the Quarry site, 1 year after release in 2020, 51 snails had moved from the quarry to the surrounding forest up to a distance of 10.3 m. All were large (15.0–21.6 mm) and most likely from the original release batch. Surveys in 2022 failed to detect any snails, but one snail (shell width 16.4 mm) was detected in 2023, 14 m from the release site.
Releases across the archipelago
In total, we released 105,970 P. bermudensis to 11 offshore islands and 16 main island sites; of these, 31,574 snails were subsequent reinforcements to eight of the sites (Table 1). Approximately 10% of all released snails were adults or subadults (shell width ≥ 15 mm), 68% were medium-sized juveniles (shell width 10–14.9 mm), and 22% were small juveniles (shell width < 10 mm). The number reintroduced was highly variable among sites, the first release to Nonsuch Island in 2016 being the smallest (204 snails). Once snails bred ex situ at Chester Zoo became available, greater numbers could be reintroduced; the release to Morgan’s Island was the largest (11,100 snails). Each reintroduction usually consisted of a mixture of size classes, but medium-sized juveniles dominated; releases at four sites included no large adults or subadults, and those at 15 sites included small juveniles, as well as other size classes.
During the last monitoring surveys, in January–February 2024, we detected live P. bermudensis on eight offshore islands and at one main island site (Table 1). At the main island site, Vesey Nature Reserve, only three large live snails were found, and no reproduction was observed. In contrast to the main island sites, reintroductions to six of the 11 offshore islands were successful and satisfied all of the criteria for success. In addition to Nonsuch Island, the snails had established self-sustaining populations on Hall’s, Higg’s, Morgan’s, Nelly’s, and Saltus islands, where they were reproducing and had expanded their distribution from the release sites (Table 1). Saltus and Higg’s islands had received reinforcement, but populations were established from the initial releases on the other four islands. The reintroduction to Rabbit Island was probably also successful, but the snails remained clustered near the release site and juveniles were underrepresented in the sample of snails measured (Table 1; Fig. 3). On Trunk Island, we detected live large snails near the release site during the monitoring survey in January 2024 but no juveniles. During an additional visit in September 2024, adult snails were noted laying eggs near the release site in a palmetto patch, but no juveniles of any size class were detected. Despite several releases and an observation of a hatchling in 2022 and egg-laying in 2024, so far there is no evidence of a self-sustaining population.
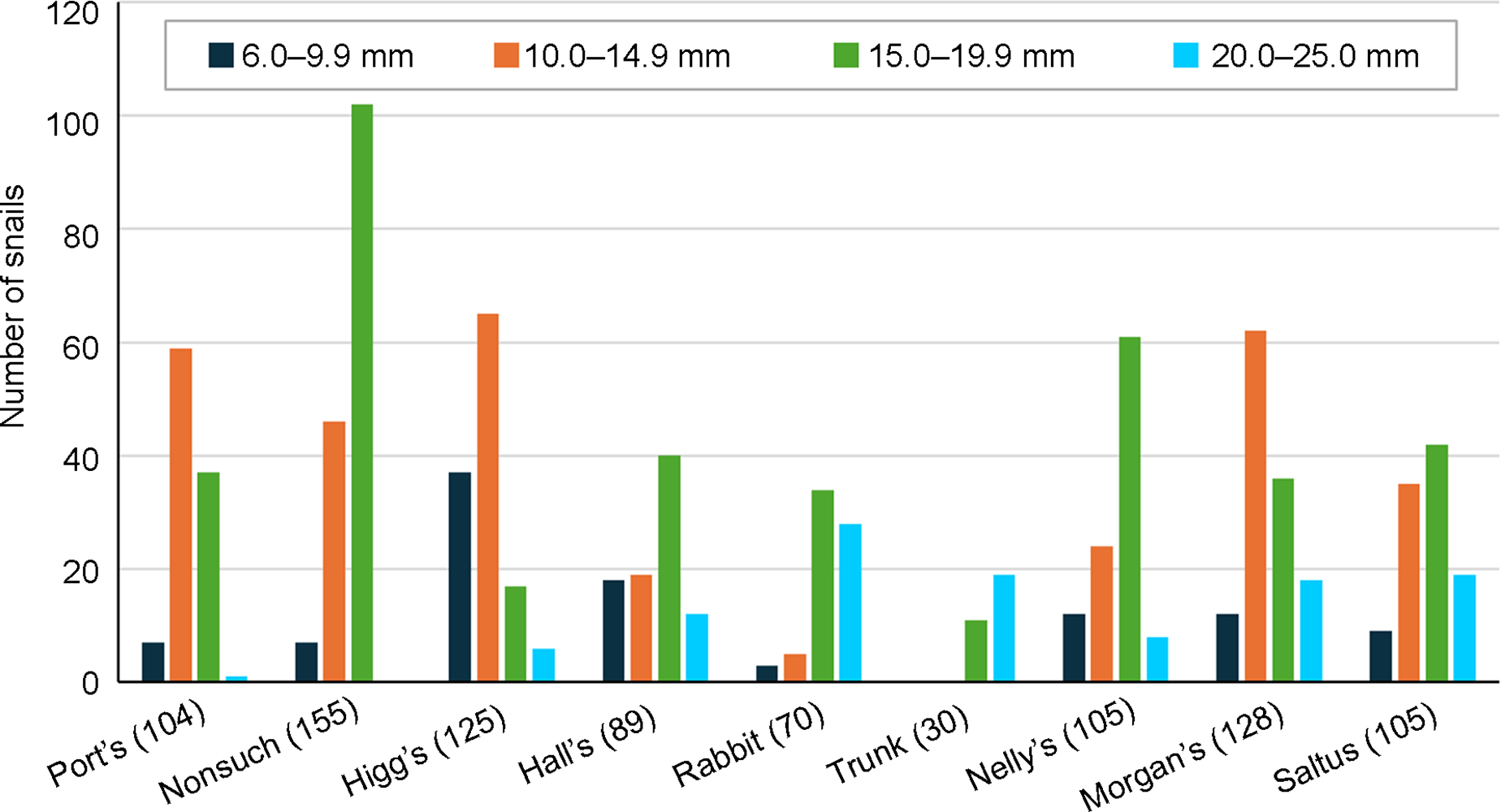
Size-frequency distribution of shell width of P. bermudensis at reintroduction sites during the last post-release monitoring surveys. A sample from the only natural remnant population, on Port’s Island, is included for comparison. Snails < 6 mm in shell width are excluded.

During the last monitoring survey, the proportion of juveniles (6–14.9 mm shell width) in the sample was relatively high at all successful sites, ranging from 34% on Nonsuch to 82% on Higg’s Island; in contrast, the corresponding value on Rabbit Island was 11%, and there were no juveniles in the Trunk Island sample (Fig. 3). There was a preponderance of large snails on Trunk and Rabbit islands, in contrast to Nonsuch Island, the oldest reintroduction, and Port’s Island, with the only remaining natural population, where this size class was virtually absent (Fig. 3).
Discussion
Reintroduction and conservation translocations are often the last resort for preventing extinction or extirpation, as was the case for P. bermudensis. Although a potentially effective tool in land snail conservation, little published information is available on the effectiveness of this approach. A guiding principle for reintroduction is that the threat or threats that caused previous extinction are either eliminated or greatly reduced, to increase the probability of success and avoid wasting effort and resources (IUCN/SSC, 2013). Once established, snail predators are often difficult or impossible to eradicate. Predator-proof enclosures to exclude E. rosea have been used with some success in habitat protection and reintroduction experiments for Partula species on Polynesian islands (Coote & Loève, Reference Coote and Loève2003) and for Achatinella species in Hawaii (Rohrer et al., Reference Rohrer, Costello, Tanino, Bialic-Murphy, Akamine, Sprague and Smith2016; Yeung & Hayes, Reference Yeung and Hayes2018). Such a labour-intensive approach was not feasible for our study.
For P. bermudensis, we chose Nonsuch Island as a site for the initial reintroduction, a model ecosystem for ecological restoration and ongoing invasive species control. Restoration of Nonsuch Island for over 60 years (Wingate, Reference Wingate and Woodwell1990) has involved the elimination of non-native species by selective culling, the gradual reintroduction of native flora and fauna, and the artificial creation of habitats (e.g. ponds) and ecological niches (e.g. cement nesting burrows for the Bermuda petrel Pterodroma cahow) to accommodate the needs of threatened species. Presently, c. 75% of the island is covered in native woodland, much of which is decades old and characterized by deep leaf litter, a feature heavily used by the reintroduced snails. Poecilozonites bermudensis rapidly established a self-sustaining population at the initial introduction site (Wall site) on this island. The disappearance of snails with shell width ≥ 20 mm over time may reflect a density-dependent effect as a result of food limitation at high densities; a survey of a natural population on Port’s Island showed a similar absence of large snails (Fig. 3). Whether the reduction in body size was associated with earlier maturity is unknown. Life history characteristics of this species are largely unknown, but tagged snails on Nonsuch Island were infrequently present for > 2 years (Ovaska et al., Reference Ovaska, Outerbridge and Sopuck2025); assuming maturity in 1 year, generation time may be 2–3 years.
Subsequent reintroductions of P. bermudensis across Bermuda occurred on other small islands with apparently suitable habitat; however, it was impossible to find islands devoid of all predators. Although feral chickens were absent from the offshore islands, rats were present or suspected to be present on many despite ongoing control efforts (e.g. on Nonsuch, Higg’s and Saltus islands), and the suite of invertebrate predators on these islands, especially predatory flatworms, was not fully known. However, due to the isolation and exposure to harsh environmental conditions (e.g. wind and salt spray), invertebrate predators are expected to be less diverse on the uninhabited offshore islands than on the inhabited main island; this is supported by our reconnaissance surveys prior to reintroductions (e.g. E. rosea appeared to persist only on Trunk Island and in low abundance). The successful breeding programme for P. bermudensis at Chester Zoo, including the prolificity of the species in the rearing facility, allowed us to experiment with releases of large numbers of snails to a wide range of sites. A similar approach for species of Partula is in progress in Polynesia, with variable outcomes, based on a successful ex situ breeding programme at Chester Zoo (ZLS, 2025).
Reintroductions of P. bermudensis to six offshore islands were successful, but none of the main island releases resulted in a viable population; reintroduction to a seventh island (Rabbit) is also likely to prove successful given more time. The factors contributing to success are not entirely clear, but we suspect that predation is largely responsible for the death of the snails on the main island sites. Habitat quality and the availability of suitable microhabitats and refuges on the forest floor are probably also important. Restoration activities on the offshore islands may have improved soil quality and leaf litter composition and increased the ability of the natural vegetation to buffer Bermuda’s extreme weather conditions. Competition with introduced gastropods for limited resources (food, retreats) could also be a factor. The introduced milk snail Otala lactea is widespread on the main island and also occurs on Trunk Island, where it appears to be spreading. These snails could potentially compete with P. bermudensis because of their large size, local abundance and similar food requirements. On the main island, rats and feral chickens that prey on snails and disturb habitat are ubiquitous; among invertebrate predators, E. rosea appears to be in decline across the archipelago, but snail-eating flatworms and nemertines are widespread; seven species of planarians and two of nemertines have been identified from Bermuda (Jones & Sterrer, Reference Jones and Sterrer2005). We found shells of P. bermudensis with signs of rat predation at several release sites, but often there was little or no trace of the snails during the post-release surveys. For example, at Spittal Pond, we found only a few shell fragments and no live snails just 2 months after the release.
All but one of the islands with successful reintroductions (Nelly’s) contain native palmetto forest, but although P. bermudensis was associated with this habitat on Nonsuch Island, sheltering under dead fronds and feeding on the nuts (Ovaska, Reference Ovaska2017), the species occurred in a wide range of forested habitats on Port’s Island (Copeland & Hesselberg, Reference Copeland and Hesselberg2021). The availability of refuges such as coral-limestone rocks, where the snails can shelter from predators and the weather, appears to be a key habitat requirement (Copeland & Hesselberg, Reference Copeland and Hesselberg2021) and may account for the reintroduction success on Nelly’s Island, where the forest was composed primarily of introduced species. Human-made structures appear to perform a similar function, and the presence of the old stone wall on Nonsuch Island and the collapsed remains of an old stone building on Nelly’s Island probably facilitated the establishment of the snails.
Island area and the number of snails reintroduced were unrelated to reintroduction success. Releases on both the smallest (Rabbit, 0.3 ha) and largest (Nonsuch, 6.6 ha) offshore islands were successful. Similarly, the smallest (Nonsuch Wall site, 204 snails) and largest (Morgan’s Island, 11,100 snails) releases were successful. Releases at two additional sites on Nonsuch Island (Grid and Quarry sites) failed to establish populations, although the initial results were promising. Individuals grew and some survived for ≥ 1 year, but they either failed to reproduce or the young failed to survive. Whether the failure was due to predation, adverse environmental conditions or unsuitable microhabitats is unknown; factors affecting early survival also require clarification. The snails reintroduced were a mixture of size classes, as age-related survivorship is unknown, but with a preponderance of larger juveniles. Among Achatinella in Hawaii, subadults appear to be the most vigorous and capable of surviving their release (Oahu Rare Snail Working Group, 2007).
As a result of this project, self-sustaining populations of P. bermudensis now occur on six islands, with a population on a seventh island in the initial phase of establishment. Together with Port’s Island where the species occurs naturally, the long-term recovery objective of viable populations at 10 localities across Bermuda could be achieved, increasing the probability of the species’ survival. On the islands where the species is established, the snails are becoming part of ecological processes both through their feeding activities and as prey for litter-foraging songbirds and presumably for native soil invertebrates, thus contributing to ecosystem recovery. The lessons learnt can be applied to other land snails, including P. circumfirmatus, for which reintroduction is currently in progress in Bermuda. Isolated, largely predator-free habitat patches, exemplified by small offshore islands, offer a higher likelihood of success than larger land masses or continuous habitat. Isolation also facilitates safeguarding the species from the introduction of new predators or diseases that could rapidly spread over a continuous land mass or habitat. A priori information on survivorship patterns of different age classes, either in the laboratory or in the wild, and on interactions of refuge type and number with survivorship in the wild, would help in determining the optimal reintroduction numbers and the selection of release sites. Monitoring the fate of releases and potential predator incursions will enhance early detection of impending problems and timely response.
Author contributions
Ex situ breeding, reintroductions, transport of snails to Bermuda: GG; reintroductions, site selection: MEO; fieldwork: MEO, KO; analysis, writing: KO; figures, tables: MEO, KO; revision: MEO, GG, KO.
Acknowledgements
We thank the ectotherm team at Chester Zoo for their propagation efforts and veterinary care, particularly Heather Prince, Tamas Papp, Javier Lopez and Amber Flewitt; the Bermuda Zoological Society for financial support; those who assisted in the field, particularly Alison Copeland, Lawrence Doughty, Tamas Papp and Heather Prince; Penny Rudd, Barbara Outerbridge, Liz Ball, Lissa Ireland, Iri Gill and Jay Redbond for facilitating the repatriation process; Daniel Dickinson for preparing the figures; and Robbie Struan Smith who helped make this study possible. This is a contribution from the Bermuda Biodiversity Project, Bermuda Aquarium, Museum & Zoo. Chester Zoo, UK, provided ex situ breeding facilities, including staff time, operational costs, health screening and staff support for post-release monitoring and shipping of snails to Bermuda, and funding for colour printing of figures. Bermuda Zoological Society provided grants in 2018 and 2020 to KO for monitoring released snails, and funding for publication. Department of Environment and Natural Resources, Government of Bermuda, provided logistics and in-kind support for fieldwork.
Competing interests
None.
Ethical standards
This research abided by the Oryx guidelines on ethical standards. Captive rearing procedures were approved by the Chester Zoo Ethics Committee. The study was conducted under permits issued by the Government of Bermuda’s Department of Environment and Natural Resources.
Data availability
Data used for this study are available from the corresponding author upon reasonable request.




 Open access
Open access