Non-technical Summary
Classification of fossils is largely a matter of convenience; their phylogeny reflects their evolutionary history. Nevertheless, the most useful classifications reflect the evolution of a group. Currently the blastozoan family Gomphocystitidae includes only two genera, Gomphocystites and Pyrocystites, which were both directly attached in life and gathered food using five unilateral spiral ambulacra. Most blastozoan ambulacra are straight and bilateral, having erect brachioles alternating left and right regularly. Curved or spiral blastozoan ambulacra are always unilateral with evenly spaced brachioles only on the outside of the food groove. So, do the spiral ambulacra of Gomphocystites and Pyrocystites reflect common ancestry or the functional requirements of efficient filter feeding? To form an efficient filter, the brachioles should be equally spaced along the ambulacrum. Functional analysis shows that it is geometrically impossible to have both regularly alternating and equally spaced brachioles along a curved or spiral food groove. Thus, unilateral spiral ambulacra reflect functional requirements not phylogeny. Spiral gomphocystitid ambulacra increased food gathering capacity by inducing currents that processed larger volumes of seawater. Gomphocystites ambulacra are composed of uniserial adambulacral plates, each of which bears a single brachiole. Using this character to define the Gomphocystitidae shows that Fungocystiites, Celticystis, and a new genus, Fresticystis, can also be assigned to the family, but Pyrocystites cannot because it lacks uniserial adambulacral plates. The four recognized gomphocystitid genera differ in the details of their oral plating. This review also provides accurate morphological data on which to base future phylogenetic analyses.
Introduction and historical review
Although cladistics has become central to many aspects of paleontology, the fact remains that most fossil taxa were described before cladistics had been thought of. Characters that are currently deemed significant may not have been recorded or may have been described inaccurately (Paul and Toom, Reference Paul and Toom2021a). The most sophisticated cladistic analysis is likely to produce poor results if the basic data entered are incorrect. So modern taxonomic revisions of taxa described in the past two centuries can be crucial in preventing such errors. Furthermore, although treating all characters as equal is the default state in cladistic analysis, it is true that some characters are phylogenetically more significant than others. Morphological characters that perform a basic vital function may be restricted in their structure due to functional constraints. If there are only a few solutions to a functional problem, it is inherently likely that the same solutions will have evolved independently several times. So functional analysis of key morphological features may improve understanding of their phylogenetic significance.
The Ordovician to Silurian diploporite family Gomphocystitidae is a case in point. It is here defined by the possession of at least some unilateral spiral ambulacra composed of adambulacral plates, each of which bears a single facet for unknown erect feeding structures (probably biserial brachioles). Using this definition, the genera Gomphocystites, Fungocystites, Celticystis, and Fresticystis nov. are assigned to the family. Hall (Reference Hall1864) described the genus Gomphocystites for Silurian diploporites from New York state and Wisconsin that were characterized by inverted pyriform thecae with a central periproct, a relatively small mouth from which five spiral food grooves spread over the oral surface. The spiral ambulacra were a key character unknown in other diploporites at the time. The specimens from Wisconsin were all dolomitic internal molds, whereas the species from New York, G. tenax Hall, Reference Hall1864, preserved the original calcite plates, which were highly sculpted. Later, Angelin (Reference Angelin1878, p. 31, pl. 9, fig. 20) described a fourth species, Gomphocystis gotlandicus, from the Silurian of Gotland, Sweden. Barrrande (1887, p. 170) described an Ordovician genus, Pyrocystites, almost entirely from internal molds. The type species, P. pirum Barrande, Reference Barrande1887, is a diploporite with an inverted pyriform theca, spiral food grooves, and a smooth thecal surface. Barrande (Reference Barrande1887, p. 157) also introduced the genus Fungocystites, the type species of which, F. rarissimus Barrande, Reference Barrande1887, possesses at least one spiral ambulacrum and has highly sculpted thecal plates similar to those of Gomphocystites.
Miller (Reference Miller1889, p. 249) described Gomphocystis indianensis, from the Silurian of Indiana, USA, and proposed the family Gomphocystidae (sic) (Miller, Reference Miller1889, p. 215), which was neither diagnosed nor described but included the genera Gomphocystites and Hemicosmites von Buch, Reference von Buch1840. The latter genus is a rhombiferan, so it is difficult to understand what Miller thought united the two genera. Jaekel (Reference Jaekel1899, p. 414) accepted the family name Gomphocystidae but included Gomphocystites and Pyrocystites. He illustrated the spiral ambulacra of Gomphocystites (Jaekel, Reference Jaekel1899, p. 415, fig. 87) and an example supposedly of G. glans Hall, Reference Hall1864 (Jaekel, Reference Jaekel1899, pl. 3, figs. 8, 9) from Chicago, Illinois. In addition, he figured Pyrocystites pirum (Jaekel, Reference Jaekel1899, pl. 3, fig. 6) showing spiral food grooves adorally on the theca. Jaekel thought the spiral ambulacra united the two genera, but he also recorded the essential differences between Gomphocystites and Pyrocystites. The spiral ambulacra of Gomphocystites are composed of a uniserial row of adambulacral plates, each of which bears a facet for a feeding structure on the outside of the main spiral food groove; the food grooves of Pyrocystites bear no relationship to the thecal plates they cross.
Bather (Reference Bather1906, p. 36, pl. 2, figs. 15–20) described and illustrated under the name Protocrinus sparsiporus nov. a diploporite from the Naungkangyi Beds (Ordovician) of the Northern Shan States, Myanmar. His figure 15 shows an entire theca in oral view with at least two spiral ambulacra, and figure 19 shows a single isolated adambulacral plate with a curved main food groove entirely within the plate giving rise to a single lateral food groove ending in a facet. This is the structure of adambulacral plates of Gomphocystites, so although this record has been overlooked, Ordovician gomphocystitids have been known since early in the past century. Foerste (Reference Foerste1920, p. 57) added “G.” bownockeri from the Niagaran rocks of Ohio and (Foerste, Reference Foerste1920, p. 56) an unnamed species from the Louisville Limestone near Anchorage, Kentucky. Kesling (Reference Kesling and Moore1968, p. S240) accepted the family Gomphocystitidae to include Gomphocystites and Pyrocystites.
More recently, Bockelie (Reference Bockelie1979) redescribed the Swedish species G. gotlandicus and made it type of a new genus, Celticystis Bockelie, Reference Bockelie1979. He accepted the family Gomphocystitidae but doubted that Pyrocystites belonged in it. Celticystis was distinguished from Gomphocystites by a mouth frame of only four oral plates, not the seven seen in Fresticystis bownockeri, and because the theca was helmet shaped and lacked the aboral extension seen in typical Gomphocystites spp. from North America. Bockelie also added detailed morphological information about the thecal structure and growth of Celticystis gotlandicus (Angelin, Reference Angelin1878). Paul (Reference Paul1984, p. 66) described G. asper, from the Hirnantian (Upper Ordovician) of Wales. Most recently, Sheffield and Sumrall (Reference Sheffield and Sumrall2019) considered Gomphocystites and Celticystis to be synonymous after comparing G. indianensis with C. gotlandicus. Unfortunately, neither is the type species of Gomphocystites, and Bockelie (Reference Bockelie1979) demonstrated several different plate arrangements in other gomphocystitid species. Thus, the first aim of this paper is a taxonomic revision of the family Gomphocystitidae. Since the key character used to unite Pyrocystites with Gomphocystites was the presence of spiral ambulacra, a review of the functional morphology of spiral ambulacra is appropriate along with constituents of the family Gomphocystitidae. The position of Fungocystites from the Ordovician of Bohemia also needs considering since some if not all its ambulacra are uniserial, unilateral, and spiral.
Geologic setting, preservation, and taphonomy
Stratigraphic context
The original examples of Gomphocystites were described from Silurian rocks. Gomphocystites glans and G. clavus Hall, Reference Hall1864 were from the “Niagara Group” of Racine, Wisconsin; G. tenax was from the “Niagara shale” of Lockport, New York. The Racine examples were probably Sheinwoodian (McLaughlin et al., Reference McLaughlin, Mikulic and Kluessendorf2013, p. 17, fig. 2). In New York State, Gomphocystites is restricted to bioherms in the Gasport Formation and has been recorded from two localities at Lockport (Brett, Reference Brett1985, table 1, p. 824). The Gasport Formation is Sheinwoodian (Cramer et al., Reference Cramer, Brett, Melchin, Männik and Kleffner2011). Gomphocystites indianensis Miller, Reference Miller1889, came from the Niagara Group of Jefferson County, Indiana. Foerste (Reference Foerste1920, p. 56) added the horizon as the Osgood Formation. The preservation suggests it came from the Lewisburg Formation of Brett et al. (Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012), formerly the Middle Osgood Limestone (Foerste, Reference Foerste1897), which is Sheinwoodian. Fresticystis bownockeri (Foerste, Reference Foerste1920) occurs in the Cedarville Dolomite of Ohio, which is also Sheinwoodian (C.E. Brett, personal communication, 2025). Fresticystis sp. Foerste (Reference Foerste1920) came from the Louisville Limestone of Kentucky. Brett (personal communication, 2025) suggested the horizon from which the Fresticystis specimen came is probably upper Homerian. Thus, this is the youngest occurrence in Midwest North America.
In Europe, Celticystis gotlandicus occurs in the Högklint Limestone of Gotland, which is Sheinwoodian (Calner et al., Reference Calner, Jeppsson and Munnecke2004). The British species F. asper comes from limestone blocks of Hirnantian (Upper Ordovician) age in channel deposits excavated during the Hirnantian glaciation at Meifod, Central Wales (Brenchley et al., Reference Brenchley, Marshall, Harper, Buttler and Underwood2006). Bather’s “Protocrinus” sparsiporus came from the Naungkangyi Beds (Ordovician) of Myanmar. Originally, the Naungkangyi Beds included the entire local Ordovician succession. The precise age of the rocks from which “P.” sparsiporus came is unknown. According to Aung and Cocks (Reference Aung, Cocks, Barber, Khin and Crow2017), the horizons from which Bather’s (Reference Bather1906) echinoderms came were either Darriwilian or Katian. Finally, Fungocystites Barrande, Reference Barrande1887 is known from the type species F. rarissimus Barrande, Reference Barrande1887 and F. solitarius Barrande, Reference Barrande1887. The latter is an unidentifiable internal mold of an oral area. The type species occurs in the Vinice Formation (Prokop and Petr, Reference Prokop and Petr1999), which correlates with the upper Sandbian international stage (Gutiérrez-Marco et al., Reference Gutiérrez-Marco, Sá, Garciá-Bellido and Rábano2017). Recently, another occurrence of Fungocystites has been reported from the underlying Letná Formation (upper Sandbian) but not illustrated or described (Fatka et al., Reference Fatka, Nardin, Budil, Nohejlová and Zicha2025)
Preservation and taphonomy
With the exception of G. tenax Hall, Reference Hall1864 from New York State, the original species of Gomphocystites were preserved as dolomitic internal molds. This preservation rarely preserves the plate arrangement around the mouth, details of which are highly significant in diploporite systematics. Unfortunately, Miller (Reference Miller1889, p. 249) selected G. glans Hall, Reference Hall1864 as type species of Gomphocystites, which is still known only from internal molds. Bockelie (Reference Bockelie1979) illustrated three significantly different arrangements of oral plating in gomphocystitids (Fig. 1). One formed the basis for his new genus Celticystis (Fig. 1.1); both the others were included in Gomphocystites. Unfortunately, the uncertainty surrounding the oral plating of G. glans means the defining morphology of Gomphocystites sensu stricto remains unknown. Nevertheless, there are reasons for believing it may have resembled that seen in G. tenax (Fig. 1.3).
Interpretive diagrams of oral plating in gomphocystitid genera. (1) Celticystis Bockelie, Reference Bockelie1979. (2) Fresticystis n. gen. (3) Gomphocystites Hall, Reference Hall1864. (4) Fungocystites Barrande, Reference Barrande1887. A–E (3, 4): ambulacra using Carpenter’s (Reference Carpenter1884, Reference Carpenter1891) symbols; An: anus (within periproct); F (4): ambulacral facet; H (1, 2, 4): hydropore; M: mouth (within peristome); PO1–6: perioral plates. (1, 2) Redrawn and reinterpreted from Bockelie (Reference Bockelie1979, fig. 3, p. 159. (3) Redrawn and reinterpreted from Bockelie (Reference Bockelie1979, fig. 11a, p. 165). (4) New.

The specific environmental conditions under which Gomphocystites species lived are known for G. tenax and F. asper. Both were directly attached (cemented) to hard substrates and elevated in reef-like environments. The former occurs in the Gasport Limestone only in reef environments (Brett, Reference Brett1985). The latter was discovered in weathered limestone blocks in a channel excavated in the underlying Katian sediments by the sea-level fall associated with the Hirnantian glaciation (Brenchley et al., Reference Brenchley, Marshall, Harper, Buttler and Underwood2006). These authors stated the cystoids (including “Gomphocystites”) lived on the limestone boulders during the sea-level rise after the glaciation. In addition, Bockelie (Reference Bockelie1979, p. 163) described the environment of C. gotlandicus as a “shallow-water, reef environment.” Such an environment is also consistent with the dolomitized original species of Gomphocystites, which often came from quarries excavated in bioherms in Illinois and Wisconsin.
Materials and methods
Wherever possible, comparative information is derived from photographs of specimens rather than interpretive diagrams. My own interpretive diagrams are either overlain on photographs or produced using a camera lucida and a Wild binocular microscope. Photographs were taken either in stacks manually using a Nikon D300 digital camera and subsequently combining the images using Zerene Stacker software, or using an Olympus Tough TG6 digital camera which, in microscope mode, combines stacked images automatically. Specimens were whitened with ammonium chloride for photography. Stereoscopic images were taken with a Nikon Coolpix camera and a Kyowa binocular microscope.
Repositories and institutional abbreviations
Specimens referred to or illustrated in this paper are in the following repositories: American Museum of Natural History, New York (AMNH, Hall’s type material), Buffalo Museum of Science, New York (BMS, Gomphocystites tenax), Sedgwick Museum, Cambridge, England (CAMSM, Fresticystis asper), Field Museum, Chicago, Illinois, USA (FMNH, Celticystis indianensis and Fresticystis bownockeri), National Museum, Prague, Czech Republic (NMP, Fungocystites rarissimus), Ohio State University Museum, Columbus, Ohio (OSU, Fresticystis bownockeri), Riksmuseum, Stockholm, Sweden (RM, Celticystis gotlandicus), United States National Museum of Natural History, Smithsonian Institution, Washington, DC (USNM, Fresticystis bownockeri).
Systematic paleontology
Class Diploporita Müller, Reference Müller1854
Diagnosis
Blastozoans with multiple dipores in thecal plates.
Remarks
Although Sheffield and Sumrall (Reference Sheffield and Sumrall2019) argued that the class Diploporita is polyphyletic, there are reasons to believe that all diploporites, both ambulacralian and anambulacralian, can be traced back to the Cambrian family Lichenoididae Jaekel, Reference Jaekel1918 (see reconstructions of Peridionites Whitehouse, Reference Whitehouse1941 and Cymbionites Whitehouse, Reference Whitehouse1941 in Jell and Sprinkle, Reference Jell and Sprinkle2021). If this is correct, the class is monophyletic and should be retained. Paul (Reference Paul1972, p. 1) introduced the term “dipores” to include both diplopores and humatipores, the two types of pore structures found in the diploporites. Sumrall and Gahn (Reference Sumrall and Gahn2006) have reported structures very similar to diplopores in the edrioasteroid Thresherodiscus, so it is necessary to specify that diploporites have multiple dipores per thecal plate. In Thresherodiscus, each pore pair is developed in a separate thecal plate, as are the pore pairs of echinoid tube feet, so the homologies of the pore pairs in Thresherodiscus are uncertain. Pore pairs merely allow one-way circulation of body fluids and are therefore efficient in respiratory gas exchange. They have arisen more than once in echinoids, diploporites, and Thresherodiscus.
Subclass Ambulacralia Paul and Toom, Reference Paul and Toom2021
(Paul and Toom, Reference Paul and Toom2021b)
Diagnosis
Diploporites with an oral frame composed of interradial periorals; with five ambulacra composed of adambulacral flooring plates each of which bears a facet for a single uniserial or biserial feeding appendage and was covered by biserial cover plates in life; with diplopores (modified from Paul and Toom, Reference Paul and Toom2021b, p. 237).
Remarks
All diploporites can be grouped into those with a regular ambulacral structure and those without (Paul, Reference Paul, Paul and Smith1988). This division was formalized by Paul and Toom (Reference Paul and Toom2021b) as the subclasses Ambulacralia and Anambulacralia, respectively. In most ambulacralians, the ambulacra are composed of adambulacral plates that alternate left and right along the length of the ambulacrum and each of which bears a single brachiole facet. Such ambulacralians are typified by the family Protocrinitidae (Paul, Reference Paul2024). One or two exceptions to this thecal structure occur, including the family Gomphocystitidae, which is typified by spiral uniserial ambulacra with unilateral ambulacral facets developed only on the outside (left) of the main ambulacral groove. The genus Pyrocystites Barrande, Reference Barrande1887 also has unilateral spiral ambulacra with facets only on the outside of the spiral, but the food grooves are developed on randomly arranged thecal plates. In addition, Pyrocystites has radial not interradial oral plates and other features suggesting it is a sphaeronitid diploporite. Czech specimens currently identified as Pyrocystites include at least two genera. The material needs thorough redescription, which is beyond the scope of this paper.
Superfamily Protocrinitoida Bather, Reference Bather1899
Diagnosis
Ambulacralians with mural ambulacra.
Remarks
In the Treatise (Kesling, Reference Kesling and Moore1968), this superfamily is called the Glyptosphaeritida. Unfortunately, the type genus Glyptosphaerites Müller, 1854 lacks the key characters supposedly defining the superfamily Glyptosphaeritida (Paul, Reference Paul1984; Paul and Fone, Reference Paul and Fone1997) and has several characters that confirm it is a sphaeronitid anambulacralian diploporite (Paul and Toom, Reference Paul and Toom2021b). Paul (Reference Paul1984, p. 65) first recommended that the superfamily be referred to as the Protocrinitida (now Protocrinitoida).
In protocrinitoids the ambulacra form part of the thecal wall (mural ambulacra). The superfamily includes the families Protocrinitidae Bather, Reference Bather1899, Dactylocystidae Jaekel, Reference Jaekel1899, and Gomphocystitidae Miller, Reference Miller1889. In the Mesocystidae Bather, Reference Bather1899, the biserial ambulacra are epithecal on the thecal plates according to Bather’s interpretation of their structure (Bather, Reference Bather and Lankester1900, p. 76, fig. 47). To my knowledge, the ambulacral structure is unknown in the Asteroblastidae Bather, Reference Bather and Lankester1900. No ambulacralian is known to have fewer than five ambulacra.
Bockelie (Reference Bockelie1984) demonstrated that the theca of protocrinitids was composed of three parts: an adoral part including the ambulacra and interambulacra, separated from an aboral cup composed of two or three primary plate circlets by a growth zone where new thecal plates were added. Paul (Reference Paul2024) further modified this concept by describing the adoral part as a “vault” and the “growth zone” as a generative circlet because the term “growth zone” has been used with a different meaning under the Extraxial–Axial Theory (EAT) of echinoderm development (Mooi and David, Reference Mooi and David1997; David and Mooi, Reference David, Mooi, Mooi and Telford1998). The concept of a tripartite thecal structure can also be applied to gomphocystitids. Typically, protocrinitids retain their stem throughout life, but one species, P. fragum von Volborth, Reference von Volborth1855, lost the stem in early development (von Volborth, Reference von Volborth1855; Paul, Reference Paul2024, p. 9). In gomphocystitids, the aboral cup is represented by the aboral extension of the theca below the limits of the ambulacra, which was cemented directly to a hard substrate and, at least in some species, may have lacked any skeletal plates within the attachment ring. The generative circlet is the upper limit of this aboral extension where new adambulacral and interambulacral plates formed. Bockelie (Reference Bockelie1979, p. 159, fig. 2) showed that terminal adambulacral plates sometimes extended onto the ring of basal plates in Celticystis gotlandicus.
Family Gomphocystitidae Miller, Reference Miller1889
(Nom. correct. Pro Gomphocystidae Miller, Reference Miller1889, p. 215)
Diagnosis
Directly attached protocrinitoids with five ambulacra, some or all of which spiral in a clockwise direction from the oral pole with branches only on the left side terminating in a single facet for unknown feeding structures; ambulacra composed of uniserial adambulacral plates; interambulacra uniserial or multiserial; thecal surface highly sculpted, including pits or tubercles for diplopores (modified from Paul, Reference Paul1984, p. 65).
Remarks
Here I take the presence of uniserial, spiral ambulacra as the key new character (apomorphy) defining the family. This character apparently first appeared in Fungocystites Barrande, Reference Barrande1887 from the Vinice Formation (upper Sandbian) of Czechia, which also bears some biserial ambulacra.
Genus Gomphocystites Hall, Reference Hall1864
Synonym Gomphocystis Angelin, Reference Angelin1878, p. 31 (nom. van)
Type species
Gomphocystites glans Hall, Reference Hall1864 by subsequent designation (Miller, Reference Miller1889, p. 249).
Other species
Gomphocystites tenax Hall, Reference Hall1864. Gomphocystites clavus Hall, Reference Hall1864 is regarded as synonymous with G. glans, following Jaekel (Reference Jaekel1899, p. 421). It appears to be an example that grew more symmetrically than typical G. glans.
Diagnosis
Gomphocystitids with an oral frame of six interradial orals; two plates separating the mouth and anus aligned one above the other in the CD interambulacrum; interambulacra narrow.
Occurrence
Silurian (Sheinwoodian), Midwest USA (Illinois, New York, Wisconsin)
Gomphocystites glans Hall, Reference Hall1864
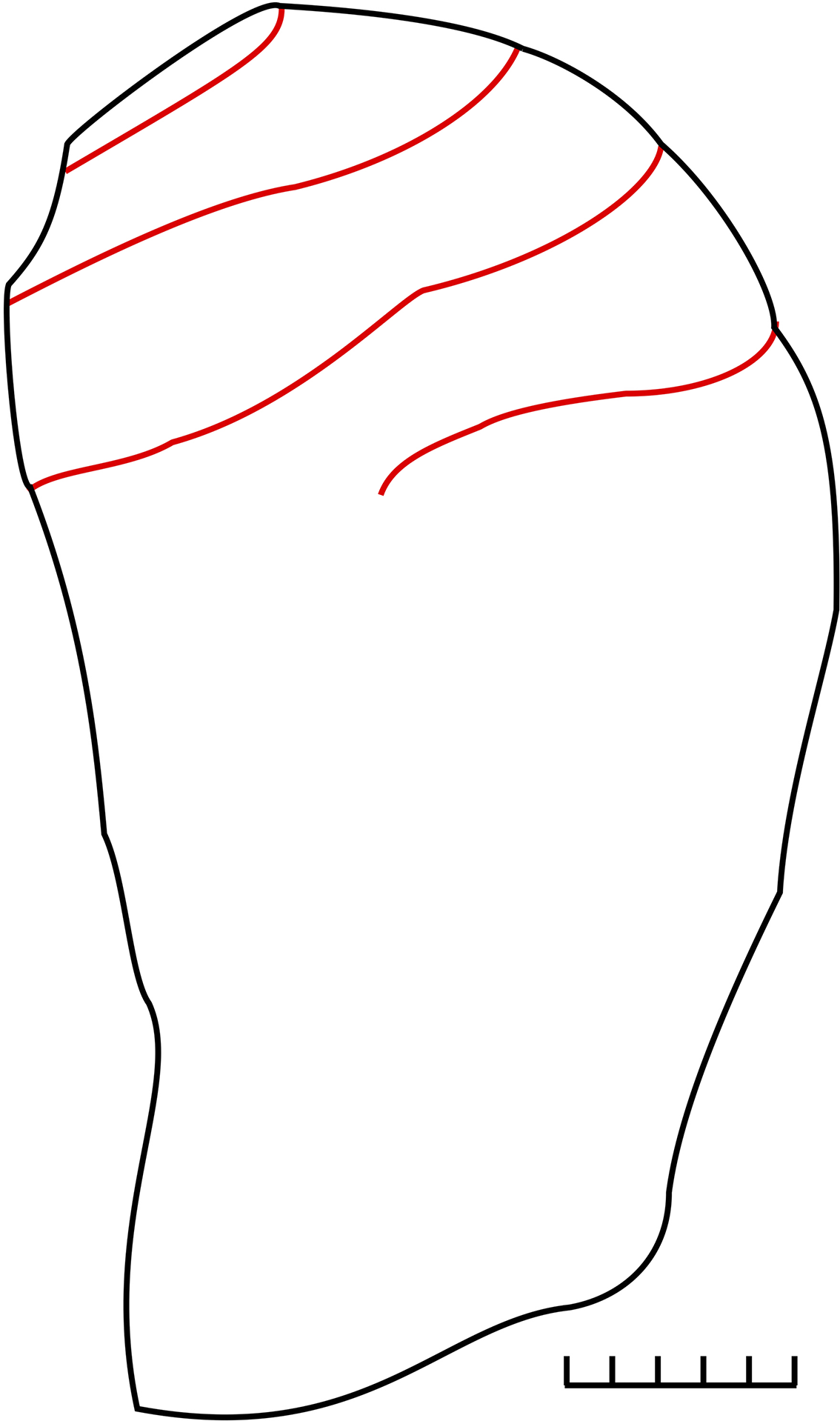
Outline drawing of the holotype of Gomphocystites glans Hall, Reference Hall1864 (AMNH 342), Racine Dolomite, Racine, Wisconsin, USA, showing the thecal outline and spiral food grooves. F G, food grooves; G C generative circlet where new plates were added during growth. In life it is likely that the generative circlet would have been horizontal. Note, parts of all five food grooves can be seen in a single lateral view. Redrawn from Hall (Reference Hall1864, pl. 1, fig. 4).

Reference Hall1864 Gomphocystites glans Hall, p. 6, pl. 1, figs. 4, 5.
Reference Hall1864 Gomphocystites clavus Hall, p. 6, pl. 1, fig. 3.
Reference Hall1868 Gomphocystites glans Hall; Hall, p. 310, pl. 12, fig. 14, pl. 12a, figs. 4, 5.
Reference Hall1868 Gomphocystites clavus Hall; Hall, p. 310, pl. 12a, fig. 3.
Reference Hall1870 Gomphocystites glans Hall; Hall, p. 352, pl. 12, fig. 14, pl. 12a, figs. 4, 5.
Reference Hall1870 Gomphocystites clavus Hall; Hall, p. 353, pl. 12a, fig. 3.
Reference Roemer1876 Gomphocystites glans Hall; Roemer, pl. 11, fig. 2a.
Reference Nicholson and Lyddeker1889 Gomphocystites glans Hall; Nicholson and Lyddeker, p. 454, fig. 324.
Reference Haeckel1896 Gomphocystites tenax Haeckel (non Hall), p. 115, pl. 3, fig. 37.
Non Reference Jaekel1899 Gomphocystites glans Jaekel, p. 421, pl. 2, figs. 8, 9.
Reference Grabau and Shimer1910 Gomphocystites glans Hall; Grabau and Shimer, p. 462, fig. 1768.
Reference Bassler and Moodey1943 Gomphocystites glans Hall; Bassler and Moodey, p. 50, 161.
Reference Bassler and Moodey1943 Gomphocystites clavus Hall; Bassler and Moodey, p. 50, 161.
Reference Bockelie1979 Gomphocystites glans Hall; Bockelie, p. 164, fig. 10C–F.
Non Reference Kesling and Moore1968 Gomphocystites glans Hall; Kesling, p. S241, fig. 140.1b (copied from Jaekel, Reference Jaekel1899).
Holotype
AMNH 342 (according to Bockelie, Reference Bockelie1979, p. 164, fig. 10), an internal mold of an almost entire theca with extensive aboral cup.
Material
Hall (Reference Hall1864, p. 6, pl. 1, figs. 4–6) illustrated only the holotypes of G. glans and G. clavus, but Bockelie (Reference Bockelie1979, p. 164, fig. 10) illustrated thecal outlines of another seven paratypes of G. glans and all registered under AMNH no. 2026). Hall (Reference Hall1864, p. 310) stated that all his material of both G. glans and G. clavus came from the Racine Dolomite at Racine, Wisconsin.
Diagnosis
Gomphocystites with a low domed vault and a variable tapered aboral theca.
Occurrence
Silurian (Sheinwoodian), Racine Dolomite Formation, Racine, Wisconsin, USA.
Description
(Modified from Hall, Reference Hall1864, p. 6.) The theca is divided into a relatively low vault containing the five food grooves, and a variable aboral stalk-like cup (Bockelie, Reference Bockelie1979, fig. 10c–k). The vault is gently domed with the height between about 40% (holotype) and 60% of the maximum diameter. The holotype reaches about 72 mm high, of which the aboral stem-like portion is 50 mm on one side and over 60 mm on the other. The food grooves are extensively coiled, such that parts of all five can be seen in lateral view (Hall, Reference Hall1864, pl. 1, figs. 4, 5). Each must curve through approximately three-quarters of a whorl. As a result, the interambulacra are also quite narrow.
Remarks
Hall (Reference Hall1864) did not state the magnification of the two original illustrations of G. glans. However, he did state that this species was one to three inches (25–75 mm) long by one-half to one inch (12–25 mm) at its widest. His illustration of the holotype is 72 mm high, so I presume it was ×1. The preservation is puzzling in that all specimens are supposed to be internal molds and yet traces of the food grooves are visible. Food grooves are incised into the external surface of the plates. Bockelie (Reference Bockelie1979, p. 162, fig. 7) illustrated a stereogram of the internal surface of C. gotlandicus, and no trace of the food grooves can be seen. I suspect that the dolomitization preserved the fill of not only the specimens but also part of the plates, thus preserving impressions of the food grooves. The inner part of the plates is also dolomitized in many Holocystites specimens from the same deposits (Paul, Reference Paul1971, p. 13). Thus, the fossils are not, strictly speaking, internal molds.
Whatever the cause, three basic facts can be deduced about the type species of Gomphocystites. It has a relatively low domed vault with height 40–60% of the maximum diameter, a very variable stalk-like aboral cup forming up to five-sixths of the total thecal height, and food grooves that spiral extensively, giving rise to narrow interambulacra. Gomphocystites tenax is more like G. glans than is any other named species except G. clavus, which is a variant of G. glans with a symmetrical theca. Hence, I am assuming the unique oral plate arrangement in G. tenax, with two plates between the mouth and anus one above the other rather than side by side (Fig. 3), was also present in G. glans and therefore characterizes the genus Gomphocystites.
Lateral profile of Gomphocystites tenax Hall. Reference Hall1864, BMS E25001, Gasport Formation, Lockport, New York, USA, showing thecal profile and spiral food grooves confined to the upper third of the theca. Redrawn from Brett (Reference Brett1985, fig. 5.3).

Gomphocystites tenax Hall, Reference Hall1864
Reference Hall1864 Gomphocystites tenax Hall, p. 6, pl. 1, figs. 1, 2.
Reference Hall1868 Gomphocystites tenax Hall; Hall, p. 310, pl. 12, fig. 15, pl. 12a, figs 1, 2.
Reference Hall1870 Gomphocystites tenax Hall; Hall, 352, pl. 12, fig. 15, pl. 12a, figs 1, 2.
Reference Roemer1876 Gomphocystites tenax Hall; Roemer, pl. 11, fig. 3.
Non Reference Haeckel1896 Gomphocystites tenax Haeckel, p. 115, pl. 3, fig. 37 = Gomphocystites glans Hall, Reference Hall1864.
Reference Bather and Lankester1900 Gomphocystites tenax Bather, p. 77, fig. 48.
Reference Bassler and Moodey1943 Gomphocystites tenax Hall; Bassler and Moodey, p. 49, 161.
Reference Kesling and Moore1968 Gomphocystites tenax Hall; Kesling, p. S241, fig. 140.1c, d.
Reference Bockelie1979 Gomphocystites tenax Hall; Bockelie, fig. 11, p. 165.
Reference Brett1985 Gomphocystites cf. G. tenax Hall; Brett, p. 824, fig. 5.3.
Holotype
AMNH 1700, the upper part of a theca preserving the original plates.
Material
Hall (Reference Hall1864, p. 6, pl. 1, figs. 1, 2) recorded only the holotype from Lockport, New York (AMNH 1700). Brett (Reference Brett1985, fig. 5) illustrated other examples and recorded that this taxon is locally common in the reef facies of the Gasport Formation.
Diagnosis
Gomphocystites with an inverted pyriform theca attached by a relatively broad attachment area.
Occurrence
Silurian (Sheinwoodian), Gasport Formation, Lockport, New York, USA.
Description
(Modified from Hall, Reference Hall1864, p. 6.) Theca inverted pyriform, with moderately domed vault and elongate aboral theca terminating in a relatively large attachment area (Fig. 3); with food grooves curved through about 230° (Fig. 4); with narrow interambulacra, uniserial as far as is known. Mouth frame composed of six plates (Fig. 1.3), of which only PO1 lies in the CD interambulacrum. A second plate occurs below PO1 between the angular mouth and the rounded anus, so the latter is farther from the mouth than in other gomphocystitid genera (Fig. 1.3). The main food grooves extend beyond the last lateral branch to an ambulacral facet (Fig. 5). Thecal surface highly sculpted with diplopores in tubercles.
Spiral food grooves in the holotype of Gomphocystites tenax Hall, Reference Hall1864 (AMNH 1700) showing the spiral angle through which each ambulacrum curves (about 230°). Straight lines represent five radii starting at the origins of the food grooves and all at 72° to each other. Note that each food groove overlaps with the next one clockwise. Currents approaching from any direction would be filtered by part of at least two ambulacra. A–E: ambulacra using Carpenter’s (Reference Carpenter1884, Reference Carpenter1891) notation; An: anus. Based on Hall (Reference Hall1864, pl. 1, fig. 2).

Distalmost part of two ambulacra and adjacent plating in holotype of Gomphocystites tenax Hall, Reference Hall1864, Gasport Formation, Lockport, New York, USA (AMNH 1700), showing uniserial adambulacral plates. Note the food groove extends beyond the last lateral branch (right ambulacrum). Interambulacral plates white. Redrawn from Bockelie (Reference Bockelie1979, fig. 11b, p. 165).

Remarks
Bockelie (Reference Bockelie1979, p. 165, fig. 11a) illustrated the unique oral plate arrangement of G. tenax in which at least two plates separate the mouth and anus but are arranged one above the other (Fig. 1.3). In all other gomphocystitids in which the oral plating is known, the two plates separating the mouth and anus contribute to the margins of both orifices and are side by side in the CD interambulacrum (Fig. 1.1, 1.2, 1.4). The interpretation of the six plates forming the mouth frame as the usual six periorals seems more parsimonious than interpreting PO3 as an adambulacral plate because it bears a brachiole facet, in which case PO3 would be absent. The arrangement in G. tenax is consistent with its narrow interambulacra which, in turn, result from the tight spiral food grooves (Fig. 5). Bockelie (Reference Bockelie1979, fig. 11b, c) illustrated parts of the ambulacra and interambulacral, which were both narrow and uniserial. Food grooves that extend through about 230°, and narrow interambulacra, are characters also seen in G. glans, suggesting that the unusual oral plate arrangement may characterize the type species of Gomphocystites as well.
Genus Celticystis Bockelie, Reference Bockelie1979
(Gomphocystis Angelin, Reference Angelin1878, p. 31, nom. van.; Celtacystis Sheffield and Sumrall, Reference Sheffield and Sumrall2017, nom. van.; Gomphocystites auctt.)
Type species
Gomphocystis gotlandicus Angelin, Reference Angelin1878, by original designation (Bockelie, Reference Bockelie1979, p. 158).
Other species
Celticystis indianensis (Miller, Reference Miller1889).
Diagnosis
Gomphocystitids with mouth surrounded by four perioral plates (PO1, PO3, PO4, and PO6); PO2 and PO5 separated from the oral opening, PO1 and PO6 contribute to the borders of both the mouth and anus; with hydropore shared by PO1 and PO6 and gonopore in PO1; adambulacrals of the C ambulacrum contribute to the anal border; with hemispherical or pear-shaped theca.
Occurrence
Silurian (Sheinwoodian) Indiana, USA, and Gotland, Sweden.
Celticystis indianensis (Miller, Reference Miller1889)
Photographs of Celticystis indianensis (Miller, Reference Miller1889), Lewisburg Formation (?), Jefferson County, Indiana, holotype FMNH UC6019. (1) Oblique posterior lateral view showing thecal shape, rounded periproct, and spiral food grooves. (2) Oral view showing oval mouth at the convergence of the five spiral ambulacra and rounded periproct. (3) Anterior lateral view showing damaged thecal plates. Specimen whitened with NH4Cl sublimate; scale bars = 5 mm.

Oral surface and interpretive diagram of Celticystis indianensis (Miller, Reference Miller1889), Lewisburg Formation (?), Jefferson County, Indiana, FMNH 19708. (1) Oral surface. (2) The same with outlines of plates, food grooves, and principal orifices. (3) Interpretive diagram. Note that only four oral plates form the mouth frame. A–E: ambulacra using Carpenter’s (Reference Carpenter1884, Reference Carpenter1891) symbols; An: anus; G: gonopore; H: hydropore; m: mouth; 1–6: oral plates. Scale bar = 5 mm. Modified from Sheffield and Sumrall (Reference Sheffield and Sumrall2019, fig. 1.3).

Stereogram of lateral food groove and ambulacral facet (arrowhead) in Celticystis indianensis (Miller, Reference Miller1889), Lewisburg Formation (?), Jefferson County, Indiana, holotype FMNH UC6019. Note the curved ligament pits and stepped articulation surfaces.

Photograph of Celticystis indianensis (Miller, Reference Miller1889), Lewisburg Formation (?), Jefferson County, Indiana, holotype FMNH UC6019. Lateral view of aboral theca showing no trace of thecal plates. Specimen whitened with NH4Cl sublimate; scale bar = 5 mm.

Reference Miller1889 Gomphocystites indianensis Miller, p. 249, fig. 319.
Reference Bather1899 Gomphocystites indianensis Miller; Jaekel, p. 421.
Reference Foerste1920 Gomphocystites indianensis Miller; Foerste, p. 55, pl. 1, fig. 5a, b.
Reference Bassler and Moodey1943 Gomphocystites indianensis Miller; Bassler and Moodey, p. 43, 161.
Reference Regnéll1945 Gomphocystites indianensis Miller; Regnéll, p. 160.
Reference Regnéll1948 Gomphocystites indianensis Miller; Regnéll, p. 41
Reference Kesling and Moore1968 Gomphocystites indianensis Miller; Kesling, p. S240, fig. 140.1i, j.
Reference Sheffield2017 Gomphocystites indianensis Miller; Sheffield, p. 50, fig. 1.4, p. 145, fig. 3.1.
Reference Sheffield and Sumrall2017 Gomphocystites indianensis Miller; Sheffield and Sumrall, p. 759, fig. 4.1.
Reference Sheffield and Sumrall2019 Gomphocystites indianensis Miller; Sheffield and Sumrall, p. 741, fig. 1.3.
Reference Sheffield, Limbeck, Bauer, Hill and Nohejlová2022 Gomphocystites indianensis Miller; Sheffield et al., fig. 1b.
Holotype
FMNH UC6019.
Material
The holotype plus another well-preserved theca (FMNH 19708).
Diagnosis
Celticystis with pyriform theca.
Occurrence
From the lithology, probably Lewisburg Formation (Brett et al., Reference Brett, Cramer, McLaughlin, Kleffner, Showers and Thomka2012), Sheinwoodian, Jefferson County, Indiana.
Description
The theca is crudely clavate or mushroom shaped with an oval, domed oral surface 32 by 23 mm in the holotype and an asymmetrical, tapering stem-like extension ending in an oblique attachment area aborally; the whole is 33 mm high (Fig. 6). Orienting the oral surface horizontal with the mouth highest suggests that the theca was attached to a steeply sloping surface in life. The oral surface (Figs. 6.2, 7.1) shares five long spiral uniserial ambulacra in a 2–1–2 pattern extending clockwise around it. The mouth is a small, oval opening 2.5 mm across, entirely within the confluence of the food grooves, and surrounded by four plates (PO1, PO3, PO4, and PO6; Fig. 7.3). The ambulacra spiral clockwise and are composed of a uniserial series of small adambulacral plates, each of which bears a short lateral food groove to a sunken brachiole facet (Fig. 8). The anus is a larger, circular opening, 4 mm across, separated from the mouth by a calcite bar 2 mm wide and composed entirely of the two posterior periorals, PO1 and PO6 (Figs. 6.2, 7.1). The hydropore is shared by PO1 and PO6, and the gonopore is a pit in PO1 (Fig. 7.1). Five deep food grooves radiate from the mouth. The anterior, opposite the anus (ambulacrum A of Carpenter, Reference Carpenter1884, Reference Carpenter1891) is single and more widely separated from the others, which occur as two pairs (Figs. 6.2, 7.1). All food grooves are deep, at least 1.1 mm wide adjacent to the mouth and, where well preserved (Fig. 6.2, AB interambulacrum), show lateral ledges for the insertion of cover plates, none of which remains in place. One ambulacral facet in ambulacrum A of the holotype is well preserved. It is divided centrally with the adoral part lower than the aboral (Fig. 8), suggesting the feeding structure was a biserial brachiole. Each part is about one-quarter of a circle with a curved ligament pit. The entire facet is 1.15 mm at its widest.
Ambulacra vary in length due to the available space before encountering the junction between the oral surface and the aboral cup. In the holotype, ambulacrum C is the shortest, curves through about 90°, and does not quite overlap with ambulacrum D. Ambulacrum D is a little longer and curves through a little less than 180°, but the termination is lost due to damage in that part of the theca. Ambulacrum E is by far the longest and curves through about 270° before reversing its curvature terminally as it becomes parallel to the end of ambulacrum A, which itself curves through about 220°. Finally, ambulacrum B is the second shortest and curves through almost 180°, with a sharp right-angle bend before the final third. The interambulacra are typically uniserial except that interambulacrum EA is locally biserial but terminates before ambulacrum E does, so that for the last part of both ambulacra E and A no interambulacral plates lie between the adambulacrals (Fig. 6.1, right). The size of interambulacral plates is typically more variable than that of the adambulacrals as they fill in the spaces between the ambulacra.
Where the theca is damaged (Fig. 6.3), the thecal plates are about 1 mm thick. Throughout the domed vault, the thecal plates are highly sculpted (Figs. 6.1–6.3, 7.1, 8), but in the aboral stalk-like cup of the holotype the thecal sculpture is not visible (Fig. 9). The cup terminates in an oval basal attachment area set at a high angle to the oral–aboral axis.
Remarks
Sheffield and Sumrall (Reference Sheffield and Sumrall2017, p. 758, Reference Sheffield and Sumrall2019, p. 741) rejected the genus Celticystis (their Celtacystis) on the grounds that the oral areas of the type species of Celticystis and Gomphocystites indianensis were very similar, if not identical. I agree with that interpretation, but unfortunately, G. indianensis is not the type species of Gomphocystites, thus Celticystis is not necessarily a junior synonym of Gomphocystites. Not only do we not know the plate arrangement in the oral area of G. glans, the type species of Gomphocystites, but Bockelie (Reference Bockelie1979, figs. 3, 9, 11) demonstrated at least two different oral plate arrangements in Fresticystis bownockeri and G. tenax, neither of which is the same as that seen in Celticystis gotlandicus. Furthermore, Paul (Reference Paul1984, p. 67, fig. 39) illustrated yet another oral plate arrangement in the Ordovician species F. asper. All three of these plate arrangements show an oral frame composed of at least five perioral plates plus some extra plates that Paul (Reference Paul1984, p. 67) interpreted as adambulacral plates that became incorporated into the oral frame.
Genus Fresticystis new genus
Synonym Gomphocystites Foerste, Reference Foerste1920, non Hall, Reference Hall1864
Type species
Gomphocystites bownockeri Foerste, Reference Foerste1920
Other species
Gomphocystites asper Paul, Reference Paul1984 from the Hirnantian (Upper Ordovician) of Meifod, Wales. Gomphocystites sp. Foerste, Reference Foerste1920 from the Louisville Limestone, Anchorage, Kentucky, shares several characters with F. bownockeri but is still imperfectly known.
Diagnosis
Gomphocystitids with pyriform theca in which the vault is as high as or higher than wide and merges into the aboral cup gradually; with seven or more plates forming mouth frame, of which PO1 and PO6 also contribute to the periproct frame; with five relatively open spiral food grooves; with variable interambulacral plating of at least two generations.
Occurrence
Upper Ordovician (Hirnantian) of Wales to lower Silurian (Sheinwoodian-Homerian) of Midwest, USA.
Etymology
Named for Terrence Frest in acknowledgement of his many studies on primitive blastozoans.
Remarks
The key characters of Fresticystis are the relatively high vault, the smooth transition from the vault to the theca, the additional plates forming the mouth frame, and the relatively open spiral food grooves. Apparently, it has a longer stratigraphic range than other genera, being known from the latest Ordovician to the youngest Silurian horizon from which undoubted gomphocystitids have been collected.
Fresticystis bownockeri (Foerste, Reference Foerste1920)
Drawings of Fresticystis bownockeri (Foerste, Reference Foerste1920), Cedarville Dolomite (Sheinwoodian), Cedarville, Ohio, USA, holotype, OSU 8736. (1) Slightly oblique lateral view showing anus (An) and mouth (M) with five ambulacra (A–E) radiating from it in a 2–1–2 pattern. P: primary thecal plates. (2) Interpretive diagram of oral plating showing the mouth frame formed by seven plates (five periorals and two adambulacrals) with the posterior pair of periorals (PO1 and PO6) also contributing to the periproct frame (An). Modified from Bockelie (Reference Bockelie1979, fig. 9, p. 163).

?Reference Jaekel1899 Gomphocystites glans Jaekel (non Hall), p. 421, pl. 2, figs. 8, 9.
Reference Foerste1920 Gomphocystites bownockeri Foerste, p. 57, pl. 1, fig. 6a, b, pl. 2, fig. 9a, b, pl. 3, fig. 1a, b.
Reference Bassler and Moodey1943 Gomphocystites bownockeri Foerste; Bassler and Moodey, p. 49, 161.
Reference Regnéll1945 Gomphocystites bownockeri Foerste; Regnéll, p. 160.
Reference Kesling and Moore1968 Gomphocystites bownockeri Foerste; Kesling, p. S240, fig. 140.1e–h.
Reference Bockelie1979 Gomphocystites bownockeri Foerste; Bockelie, fig. 3b, p. 159, fig. 9a, b, p. 163, fig. 10a, b, p. 164.
Reference Paul1984 Gomphocystites bownockeri Foerste; Paul, p. 66.
Holotype
OSU 8736A, an almost complete theca (Fig. 10).
Material
Foerste (Reference Foerste1920, p. 57) recorded the holotype and a paratype from Cedarville, Ohio (OSU 8736A, B); at least six specimens from Chicago (FMNH UC22944 several specimens; UC22943 two specimens, and UC22495), plus USNM 4523. All partial or complete thecae.
Diagnosis
Fresticystis with a pyriform theca, with multiserial interambulacra that extend below the ambitus.
Occurrence
Cedarville Dolomite (Sheinwoodian) of Cedarville, Ohio, USA. Also reported from the Racine Dolomite (Sheinwoodian) of Chicago, Illinois, USA (Foerste, Reference Foerste1920, p. 58).
Description
Theca pyriform (Fig. 10), up to 40 mm in diameter, with five narrow, uniserial ambulacra in a 2–1–2 pattern (Fig. 10.2) and broader polyserial interambulacra, both reaching the ambitus of the theca. Rounded periproct at the summit about 4 mm across and close to a slightly excentric, elongate peristome, 4 mm wide by 3 mm anterior to posterior. Thecal plates convex, and external molds from the Chicago area show highly sculpted surfaces with diplopores in tubercles (Foerste, Reference Foerste1920, p. 58). The ambulacra are uniserial, but interambulacra are polyserial. The EA interambulacrum is biserial (Fig. 10.1) with larger primary interambulacrals adjacent to the E ambulacrum. The AB interambulacrum is similar but with two or three rows of secondary interambulacrals. The DE interambulacrum appears to have a short row of primary plates in the center with at least one row of secondaries between them and the A ambulacrum as well as two or three between them and the D ambulacrum. Large primary plates (P on Fig. 10.1) and smaller secondaries also occur in the basal cup.
The mouth frame is composed of seven plates (Fig. 10.2), here interpreted as five periorals and two adambulacrals. The two posterior periorals (PO1 and PO6) also contribute to the periproct border. The division of the lateral, paired ambulacra is closer to the mouth edge in the D and E ambulacra than in the B and C ambulacra. In F. bownockeri, the adoral vault continues into the aboral cup smoothly, without any obvious boundary.
Remarks
One diagnostic character of F. bownockeri seems to be the smooth transition from the adoral vault to the aboral theca. Thus, it seems possible that the specimen illustrated by Jaekel (Reference Jaekel1899, pl. 2, figs. 8, 9) as G. glans was a specimen of F. bownockeri. In addition, the vault of F. bownockeri is higher than wide, unlike those of the preceding species in which the height is usually 40–60% of the width. This results in less-tightly coiled food grooves that extend below the widest part of the theca and broader, multiserial interambulacra. In all these characters, Jaekel’s (Reference Jaekel1899) specimen agrees with F. bownockeri rather than with G. glans.
The interpretation of the oral plating shown here (Fig. 10.2) differs slightly from that of Bockelie (Reference Bockelie1979, fig. 3b). The current interpretation is because in Celticystis, PO1 and PO6 contribute to the margins of both the peristome and periproct. Periorals PO2, PO3, and PO4 are identified by their interradial position, and PO5 is the sixth perioral.
Fresticystis asper (Paul, Reference Paul1984)
Photographs of the holotype of Gomphocystites asper Paul, Reference Paul1984, limestone blocks in channel fill within Rawtheyan mudstones, Meifod, Wales (CAMSM X.2001). (1) General view of the theca showing attachment area (below) and one food groove (above). (2) Oblique lateral view showing plating around food groove and tubercular diplopores (arrow). Specimen uncoated; scale bars = 2 mm.

Camera lucida drawing of the oral area of Gomphocystites asper Paul, Reference Paul1984, paratype (CAMSM X.2003), limestone blocks in channel fill within Rawtheyan mudstones, Meifod, Wales. A–E: ambulacra using Carpenter’s (Reference Carpenter1884, Reference Carpenter1891) symbols; Ad: additional oral frame plates (probably adambulacrals); An: anus; F: ambulacral facet of ambulacrum A; G: gonopore; H: hydropore; M: mouth; PO1–PO6: perioral plates. Redrawn from Paul (Reference Paul1984, fig. 39, p. 67).

Reference Paul1984 Gomphocystites asper Paul, p. 66, pl. 26, figs. 6, 7, 9, 10, text-fig. 39.
Reference Brenchley, Marshall, Harper, Buttler and Underwood2006 Gomphocystites asper Paul, Brenchley et al., p. 12.
Reference Lefebvre, Sumrall, Shroat-Lewis, Reich, Webster, Harper and Servais2013 Gomphocystites asper Lefebvre et al., p. 183.
Holotype
CAMSM X.2001, a partial theca.
Material
Three specimens, the holotype CAMSM X.2001 and two paratypes, CAMSM X.2002 and X.2003. All partial thecae.
Diagnosis
Gomphocystites with a pyriform theca, with diplopores developed in prominent tubercles and more common adorally.
Occurrence
Ordovician (Hirnantian), unnamed limestone masses present in channels developed in the Rawtheyan Mudstones and below the Craig Wen Sandstone, exposed in old quarries beside the A495 road at Meifod, Wales (Brenchley et al., 2006).
Description
Theca small, pyriform (26 mm high by 16 mm wide), with distinct attachment area aborally (Fig. 11.1) and food grooves and pustular diplopores adorally (Fig. 11.2). Ambulacra spiral clockwise and are composed of small adambulacral plates about 1.5 by 1.5 mm. Interambulacral plates are larger, reaching 2.5 by 2.5 mm, and usually arranged in double rows. The aboral cup of the holotype below the ambitus is composed of five circlets of large plates reaching up to 4 mm high, except directly above the attachment area. Adambulacrals reach 0.75 mm thick, but interambulacrals are only 0.55 mm in one paratype (CAMSM X.2002). Diplopores are more numerous adorally, with up to eight per interambulacral plate. Only scattered diplopores occur below the ambitus in the holotype. Diplopores are small (0.6 by 0.4 mm), oval, and raised in distinct tubercles (Fig. 11.2, arrow). The individual pores are also oval but with long axes perpendicular to the length of the peripores, and each bears a peripheral rim. As a result, the diplopores often appear to have three pits, the two pores and a depression between them (Fig. 11.2, arrow). The peristome could be seen in one paratype (CAMSM X.2003), where the frame was formed of six perioral plates plus at least five small additional plates (Ad on Fig. 12) originally interpreted as adambulacrals (Paul, Reference Paul1984, p. 67). Orals PO1 and PO6 contribute to the margins of both the peristome and periproct and share a slit-like hydropore (H on Fig. 12). A small, circular gonopore is present in PO1 (G on Fig. 12). Food grooves radiate from the mouth and are about 1 mm wide at the corners but taper to about 0.15 mm wide at their tips. The main food grooves give rise to lateral grooves on the left side only that terminate in kidney-shaped ambulacral facets, one per adambulacral plate. The facets also decrease in size aborally from about 0.8 mm across adorally to 0. 35 mm distally. All food grooves have distinct ledges for the insertion of cover plates, but neither cover plates nor subvective appendages have been preserved.
Remarks
The additional plates in the oral frame and interambulacra formed of a double series of plates both suggest closer affinities with Gomphocystites than with Celticystis. The pyriform thecal shape is also unique among the Gomphocystitidae. Specimens were preserved in the tops of the karstic limestone pillars, which reach over 2 m elevation according to Brenchley et al. (Reference Brenchley, Marshall, Harper, Buttler and Underwood2006, p. 1). In common with other gomphocystitids, G. asper clearly lived in turbulent, well-oxygenated shallow waters as sea level rose and flooded the karst surface after the Hirnantian glaciation (Brenchley et al., 2006).
Genus Fungocystites Barrande, Reference Barrande1887
Synonym Fungocystis Haeckel, Reference Haeckel1896, p. 104 (nom. van.)
Type species
Fungocystites rarissimus Barrande, Reference Barrande1887 by original designation (Barrande, Reference Barrande1887, p. 157).
Other species
Fungocystites solitarius Barrande, Reference Barrande1887, an unidentifiable internal mold.
Diagnosis
Gomphocystitids with pyriform theca, with six plates forming mouth frame, of which PO1 and PO6 also contribute to the periproct frame, with both uniserial and biserial ambulacra.
Occurrence
Upper Ordovician (Sandbian), Vinice Formation, Kladno, Czech Republic (Prokop and Petr, Reference Prokop and Petr1999, p. 64).
Fungocystites rarissimus Barrande, Reference Barrande1887
Stereophotos and interpretive diagram of holotype of Fungocystites rarissimus Barrande, Reference Barrande1887, Vinice Formation, Kladno, Czech Republic, NMP L13065. (1) Stereogram of lateral view of latex cast showing mouth (top), food grooves, and highly sculpted thecal surface. (2) Stereogram of oral view of holotype showing oval mouth and five food grooves, all with ledges for the insertion of cover plates, elongate hydropore, and top margin of circular periproct. (3–5) Oral view (3), the same with superimposed outlines of principal orifices, food grooves, and oral plates (4), and interpretive line drawing (5). A–E: ambulacra using Carpenter’s (Reference Carpenter1884, Reference Carpenter1891) symbols; An: anus; h: hydropore; m: mouth; 1–6: perioral plates 1–6. Note that six plates form the mouth frame, two of which bear the elongate hydropore and contribute to the periproct frame. Scale bars = 2 mm; latex cast whitened with fingerprint powder.

Camera lucida drawings of ambulacra B and C in the holotype of Fungocystites rarissimus Barrande, Reference Barrande1887, Vinice Formation, Kladno, Czech Republic, NMP L13065, showing the main food and ambulacral grooves, lateral facets (F1, F2, etc.), hydropore (H), and mouth edge (M). Note that ambulacrum B is curved in a right-hand spiral and has facets only on the left, whereas ambulacrum C is straighter and has facets on both sides. In both ambulacra, the main food groove extends beyond the last facet.

Reference Barrande1887 Fungocystites rarissimus Barrande, p. 157, pl. 17 (1), figs. 1–8.
Reference Haeckel1896 Fungocystis rarissima Barrande, Haeckel, p. 105, fig. 14.
Reference Jaekel1899 Protocrinites rarissimus (Barrande), Jaekel, p. 134, pl. 3, fig. 9, 9a.
Reference Bather and Lankester1900 Fungocystis rarissima Barrande, Bather, p. 74, fig. 94.
Reference Bassler and Moodey1943 Fungocystites rarissimus Barrande; Bassler and Moodey, p. 34, 159.
Reference Kesling and Moore1968 Fungocystites rarissimus Barrande; Kesling, p. S239, fig. 138.2a–g.
Reference Prokop and Petr1999 Fungocystites rarissimus Barrande; Prokop and Petr, p. 64.
Reference Sheffield and Sumrall2019 Fungocystites rarissimus Barrande; Sheffield and Sumrall, p. 744.
Holotype
NMP L13065.
Material
The unique holotype.
Diagnosis
As for genus.
Occurrence
Vinice Formation, Upper Ordovician (Sandbian), Kladno, Czech Republic. According to Gutiérrez-Marco et al. (Reference Gutiérrez-Marco, Sá, Garciá-Bellido and Rábano2017), this correlates with the lower middle Berounian regional stage and thus the late Sandbian international stage.
Description
The theca was directly attached, inverted pyriform, at least 15 mm high by 13 mm maximum width (Fig. 13.1, 13.2), composed of plates with numerous diplopores raised in tubercles.
Thecal plates are highly sculpted but difficult to recognize individually; covered with raised tubercles that are occasionally breached revealing diplopores (Fig. 13.1–13.3). The diplopores are small, only 0.4–0.5 mm axially and with perpendicular canals 0.2 mm in diameter (Fig. 13.1).
The mouth (M on Fig. 13.5) is oval, 2.1 mm wide by 0.8 mm anterior–posterior, within a pentagonal peristome 3.2 by 1.0 mm and surrounded by six perioral plates (1–6 on Fig. 13.5). PO1 and PO6 share a dumbbell-shaped hydropore 2.0 mm long (Fig. 13.2, h on Fig. 13. 5) with a nearly circular pore at each end, 0.3 mm in diameter, and contributing to the borders of both the mouth and anus. Five food grooves, which reach 0.5 mm wide proximally, radiate from the corners of the mouth (Fig. 13.2). They are basically grooves in the external surface of the thecal plates, but with lateral channels for the insertion of cover plates in life (Fig. 13.1, 13. 2). PO1–PO5 each bears the first single, sunken, kidney-shaped, ambulacral facet 0.5 mm across (Fig. 13.5), with a lateral food groove to the left of the main food groove as viewed in growth direction (Fig. 13.2, 13.5). Ambulacrum B has five facets to the left: B1, B3, B5, B7, B9 (B on Fig. 14). Ambulacrum C has eight facets: C1, C2, C3, C5, C6, C7, C8, C9 (C on Fig. 14). In both complete ambulacra, the food groove continues beyond the last ambulacral facet (Fig. 14). Ambulacrum D is curved and appears to have facets only on the left side, but the distal tip is not preserved.
The periproct (An on Fig. 13.5) is apparently circular, about 2 mm across but only partly exposed. Facet C2 is developed on a plate that obscures the lower half or so (Fig. 13.5). The thecal surface is decidedly tubercular with diplopores developed in the tubercles (Fig. 13.1, 13.2).
Remarks
It is frustrating that F. rarissimus is still known only from a single specimen. It demonstrates intriguing characteristics that suggest affinities with gomphocystitids. It is especially regrettable that the plate sutures cannot be made out more clearly, so it is not possible to confirm that in ambulacrum B, the main food groove is entirely developed within the adambulacral plates. That would help confirm the gomphocystitid affinities. Barrande (Reference Barrande1887, p. 157–158, pl. 17(I), figs. 1–6, 8–10) described and illustrated the internal mold, which lacked plating within the basal attachment ring. This is a feature otherwise only reported by Bockelie (Reference Bockelie1979) in the Swedish Silurian gomphocystitid, Celticystis gotlandicus. Although the possession of lateral food grooves only on the outside of a curved ambulacrum is almost certainly a functional requirement (see below), the possession of such ambulacra can be seen as an evolutionary novelty, which suggests Fungocystites has affinities with other gomphocystitids.
All previously described species can be assigned to well-defined genera. The next species is undoubtedly a gomphocystitid in that it possesses uniserial spiral ambulacra, but it is too poorly known to attribute to one of the genera described above. It is therefore attributed to Gomphocystites?
Gomphocystites ? sparsiporus (Bather, Reference Bather1906)
Isolated adambulacral plate of “Gomphocystites?” sparsiporus (Bather, Reference Bather1906), Naungkangyi Formation, Sedaw, Myanmar. (1) Copy of Bather (Reference Bather1906, pl. 2, fig. 19). (2) The same with interpretation of food grooves and facets. (3) Interpretive diagram. Fa: facet for feeding structure; LFG: lateral food groove; MFG: main food groove; m: direction toward the mouth. Scale bar = 2 mm. Note the entire main food groove is within the plate and gives rise to the lateral food groove on the left as viewed in the growth direction of the food groove (i.e., away from the mouth).

Reference Bather1906 Protocrinus sparsiporus Bather, p. 36, pl. 2, figs. 15–20.
Reference Bassler and Moodey1943 Protocrinites sparsiporus (Bather, Reference Bather1906); Bassler and Moodey, p. 37, 184.
Holotype
Specimen A, original designation of Bather (Reference Bather1906, p. 37).
Material
Bather’s type set included five specimens he labelled A–E. Specimen A, a partial theca, is the holotype. Specimen B includes the isolated adambulacral plate illustrated (Bather, Reference Bather1906, pl. 2, fig. 19; Fig. 15). Specimen E is the aboral part of another partial theca showing at least four circlets of thecal plates. The illustration of specimen D (Bather, Reference Bather1906, pl. 2, fig. 18) shows nothing to identify it, and specimen C was not illustrated at all.
Diagnosis
Large “Gomphocystites ?” with a rounded theca, with large adambulacral plates.
Occurrence
Ordovician, Naungkangyi Formation, Sedaw, Myanmar.
Description
Theca large, 68 by 50 mm in the holotype according to Bather (Reference Bather1906, p. 37, pl. 2, figs. 15, 16; Bather’s figures of the holotype are said to be natural size, and the images are 80 by 55 mm). Figure 15 shows a circular periproct 10 mm across and two ambulacra, A and B of Carpenter’s terminology (Carpenter, Reference Carpenter1884, Reference Carpenter1891), both curved in right hand (clockwise) spirals and with two lateral food grooves to ambulacral facets visible in ambulacrum B. Bather (Reference Bather1906, pl. 2, fig. 16) indicated the tips of four of the food grooves in ambulacra A, B, C, and E extended just below the ambitus. The mouth is central in the oral surface, unlike other gomphocystitids in which the periproct tends to be central, and periproct relatively widely separated from it. Bather identified an oval hydropore between them, but the figure does not show this well. Isolated thecal plates (pl. 2, figs. 17, 19) have a sculpted surface covered with pits for diplopores. Adambulacrals are large, reaching 13.5 mm wide by 12.5 mm perpendicular to the food groove (Fig. 15) with a lateral food groove up to 4 mm long. The aboral surface of the holotype is broken, so any aboral stalk or attachment area is unknown.
Remarks
The presence of spiral ambulacra composed of adambulacral plates each with a lateral food groove running to a single ambulacral facet (Fig. 15) confirms that Bather’s materials represent a gomphocystitid diploporite. The holotype is very large compared with all other species, even taking Bather’s more modest dimensions. It is also distinguished from other gomphocystitids by having the mouth central in the oral surface and a considerable distance from the circular periproct, which is lateral. In most other gomphocystitid specimens, the periproct is central in the oral surface with a smaller peristome closely adjacent. The sculpted thecal surface with diplopores in pits also differs from most other gomphocystitids in which the diplopores are in tubercules. Altogether, I suspect this is a valid species, but its generic assignment remains uncertain.
Functional morphology of spiral ambulacra
Blastozoan ambulacra formed a filtration system to gather food from seawater currents using erect brachioles. The basic structure of an efficient filter requires a pattern of equal spaces small enough to trap particles of food. Large gaps in such a system would allow a significant amount of water to pass through unfiltered. Thus, to be efficient, the brachioles needed to be equally spaced along the ambulacra. This would allow the tips of the tube feet from adjacent brachioles to meet, dividing the spaces between the brachioles into a finer series of slits from which the tube feet could trap food. The tube feet themselves would also be equally spaced along the brachioles.
Magnus (Reference Magnus1963, fig. 8a) provided a photograph with a scale that confirms these basic facts and enables measurement of the spacing of pinnules and tube feet in a modern reef-dwelling comatulid crinoid. Spacing of the pinnules averages about 1 in 1.21 mm adjacent to the arm axis and 1.27 mm at their tips. Spacing of lateral food grooves in Fr. asper was measured at 1.5 mm (N = 1) and averaged 2.05 mm in Fu. rarissimus (N = 4). Crinoid pinnules are about 10 mm long and about 0.4 mm in diameter. Brachiole facets on Fr. asper were about 0.4 mm in diameter; brachiole length is unknown as they have never been preserved. Crinoid tube feet were about 0.55 mm long. Since the spacing of gomphocystitid facets is slightly larger than in the modern crinoid, it is reasonable to assume their tube feet were also slightly longer to meet across the larger space.
Modern comatulid crinoids retire into reef crevices during the day to avoid visual predators, and they perch on reef promontories to feed at night (Magnus, Reference Magnus1967). Gomphocystitids were permanently fixed (cemented) to hard substrates and were equally restricted in the positioning of their brachioles, which could be raised to feed and presumably folded down on the theca when resting. I assume gomphocystitids also attached to promontories on reefs as this would allow them to filter food before competing filter-feeding organisms.
In a curved ambulacrum, brachioles on the outside of the curve would be more widely spaced than those on the inside of the curve if they alternated regularly (Fig. 16.1). If the spacing of brachioles were constant, there would be more of them around the outside than the inside of the ambulacrum. Thus, it is impossible to satisfy the requirements of an efficient filter in a spiral biserial ambulacrum. As there is more space around the outside than the inside of the curve, more brachioles can be accommodated on that side of the food groove. Hence, if a unilateral arrangement evolved, it would be better to lose the inside set of brachioles. Furthermore, Magnus (Reference Magnus1963, Reference Magnus1967) showed that modern comatulid crinoids arrange their arms in a planar fan perpendicular to seawater currents with the food groove downstream. So another reason to have unilateral ambulacra with brachioles on the outside of the curve is that currents approaching the sessile blastozoan would first encounter brachioles with the food grooves down current (Fig. 16.2).
Functional morphology of spiral ambulacra. (1) Idealized ambulacral system with five spiral ambulacra to show effect of an ambulacrum with facets on both sides. It is physically impossible to maintain the same spacing between brachioles on both sides of the ambulacrum as well as a regular alternation of facets left and right. The shorter distance on the inside of the spiral means brachioles on this side are more closely spaced. With uniform spacing, more brachioles can be accommodated on the outside of the spiral. (2) Idealized diagram with brachioles only on the outside of the spiral to show that currents (arrows) approaching from any direction first encounter brachioles with food grooves downstream; the preferred orientation seen in living crinoids. (3) Immediate oral area of Celticystis gotlandicus (Angelin, Reference Angelin1878) to show the close correspondence between real gomphocystitid spiral ambulacra and an ideal pentaradiate arrangement (2). Based on Bockelie (Reference Bockelie1979, fig. 5a, p. 160).

Once a curved ambulacrum had turned through 72°, it would reach the start of the adjacent ambulacrum and so produce a continuous palisade of brachioles all around the oral surface (Fig. 16.2). Thus, currents approaching the blastozoan from any direction would encounter brachioles in the precise orientation to filter food efficiently. In addition, as the oral surfaces of blastozoans such as Gomphocystites are dome-like, further overlap of adjacent ambulacra would filter currents at different levels above the substrate and so increase the volume of water filtered (Fig. 17).
Suggested current systems induced by gomphocystitid spiral ambulacra. H is the height of a water column that would inevitably encounter the food gathering system in a unidirectional horizontal current. Long arrows represent the currents deflected downward between the palisades of brachioles. Short arrows represent currents passing through the brachiolar filter. With so much water deflected downward by the spiral ambulacra, additional currents from above the echinoderm would probably be drawn down into the filter, thus increasing the total volume of water filtered.

Comparing these theoretical requirements for efficient filtration systems with actual fossils demonstrates how closely the fossils approach the ideal design (Fig. 16.2, 16.3). Almost all blastozoans with spiral ambulacra turn clockwise (right hand spirals) with facets on the lefthand side, as in Gomphocystites. Nevertheless, exceptions do occur and are very informative. Haimacystis Sumrall, Sprinkle, and Guensburg, Reference Sumrall, Sprinkle and Guensburg2001 is a flattened blastozoan with two unilateral ambulacra that curve in opposite directions. Both ambulacra have facets only on the outside of the spiral. Bistomiacystis Sprinkle and Parsley, Reference Sprinkle, Parsley and Sprinkle1982 is a paracrinoid with four ambulacra, all of which spiral anticlockwise, and again all facets are on the outside of the curve. Alternatively, Canadocystis Jaekel, Reference Jaekel1900 is a paracrinoid with two ambulacra that curve clockwise, and all its facets are on the outside of the curve. Furthermore, all these examples have unilateral ambulacra. Thus, the occurrence of unilateral spiral ambulacra would seem to be due to functional constraints not phylogenetic relationships. Spiral ambulacra have arisen repeatedly among blastozoans. They are present in paracrinoids such as Canadocystis and Bistomiacystis, the eocrinoid Haimacystis, and diploporites such as Gomphocystites and Pyrocystites, which I believe belong in two different families, the Gomphocystitidae and Sphaeronitidae.
The spiral ambulacra of gomphocystitids almost certainly had another effect. All filters present a slight impediment to fluid flow through them because the filter frame itself occupies some of the area of the filter. For example, the gaps in the filter shown by Magnus (Reference Magnus1963, fig. 8a) represent only about 46% of the total area occupied by the arm. Thus, water must “squeeze through” the filter, and its velocity increases temporarily. In gomphocystitids with a filter that spirals down the domed oral surface (Fig. 17), the palisades of brachioles forming the filter would have presented such a barrier. Seawater currents are likely to have been deflected down the interambulacra, especially if the brachioles curved back at their tips to form a roof over the interambulacral spaces. This, in turn, probably induced water from above the top of the gomphocystitids to be drawn down into the filter area and thus enabled the echinoderms to filter a larger total volume of water. Furthermore, water currents higher above reefs usually contain more food particles than those lower down because fewer filter-feeding organisms have removed any food particles. Thus, by deflecting the currents downward, gomphocystitids would not only be filtering a greater volume of water but a potentially richer source as well.
Discussion
The Ambulacralia was defined as a subset of diploporite blastozoans with ambulacra composed of adambulacral plates, each of which bears a lateral branch of the food groove terminating in a facet for erect feeding structures (usually biserial brachioles). Most ambulacralians possess bilateral ambulacra with lateral food grooves alternating left and right regularly along the ambulacrum. In the Protocrinitidae, the main food groove extends beyond the last lateral side branch. and brachioles developed only when the adambulacral plates grew large enough to accommodate a brachiole facet (Paul, Reference Paul2024, fig. 5). The same terminal ambulacral morphology is known in the gomphocystitids Celticystis gotlandicus and Gomphocystites tenax (Bockelie, Reference Bockelie1979). Hence, it seems gomphocystitids possess unilateral versions of typical ambulacralian bilateral ambulacra. If one accepts the possession of similar uniserial ambulacra as an autapomorphy of the family Gomphocystitidae, then Fungocystites is a gomphocystitid even though it has at least one bilateral ambulacrum (C). Furthermore, this suggests that Fungocystites might be the most primitive gomphocystitid since all other genera have five unilateral ambulacra.
The development of spiral morphology imposes a functional constraint on the structure of blastozoan ambulacra, as discussed. Pyrocystites has five unilateral spiral ambulacra with facets only on the outside of the main food grooves. Nevertheless, it is not included within the Gomphocystitidae since it lacks the regular ambulacralian character of uniserial adambulacral plates, each of which bears a single facet; the ambulacra are unilateral but not uniserial. There is no relationship between the position of the facets and the underlying thecal plates in Pyrocystites. This is one of several characters possessed by Pyrocystites that help define the family Sphaeronitidae (Paul and Toom, Reference Paul and Toom2021b, p. 228).
Conclusion
The family Gomphocystitidae is defined by the possession of at least some spiral ambulacra composed of adambulacral plates, each of which bears a single facet. The feeding structures that arose from these facets are unknown, but the divided morphology of the facets suggests they were biserial brachioles. Using this definition, the family includes the genera Fungocystites Barrande, Reference Barrande1887, which has some straight bilateral ambulacra, Gomphocystites Hall, Reference Hall1864, Celticystis Bockelie, Reference Bockelie1979, and Fresticystis n. gen., all of which have five unilateral spiral ambulacra. Celticystis is defined by a small mouth surrounded by four oral plates. Gomphocystites and Fresticystis both have at least five orals forming the mouth frame. In all genera except Gomphocystites, the two posterior orals also contribute to the periproct border. In Gomphocystites, there are two plates between the peristome and periproct but arranged one above the other, so no oral contributes to the periproct border. Fresticystis also has less-tightly coiled ambulacra that extend just below the widest part of the theca, and multiserial interambulacra.
Gomphocystitids are known from the Ordovician of Wales, the Czech Republic, and Myanmar, as well as the Silurian of Midwest USA. “Gomphocystites” californicus Stauffer, Reference Stauffer1930 was reported from the Devonian of California but has been neither described nor illustrated since. Its status is unknown.
Acknowledgments
I thank P. Mayer, Invertebrate Collection Manager, and T. Guensburg, Field Museum, for the loan of the holotype of Gomphocystites indianensis. E. Nardin, Geosciences Environnement Toulouse, France, provided the latexes of the holotypes of Fungocystites rarissimus and F. solitarius. M. Nohejlová, Czech Geological Survey, provided photographs of Czech “Pyrocystites” specimens. M. Riley, Sedgwick Museum, Cambridge, provided access to the original specimens of Fresticystis asper. C. Brett, University of Cincinnati, helped with the finer points of Midwest American Silurian stratigraphy. Comments from E. Nardin, T. Guensburg, an anonymous reviewer, and the editor, B. Lefebvre, improved the original draft.
Competing interests
The author declares none.